
Abstract
The Eastern European Alps boasts highly biodiverse ecosystems and a rich archaeological history. However, there is limited research on the enduring impacts of historical climate change and human activities on plant biodiversity in this region. Using sedimentary ancient DNA, we reconstructed plant and animal dynamics from 8500 years before present (ka BP) around Großer Winterleitensee (Zirbitzkogel, Austria). Variable intensities of human activities since the Middle Bronze Age (~3.5 ka BP) facilitated the persistence of biodiverse Alpine meadow communities and lowered timberlines below their natural limit. Since the end of the Bronze Age (~2.8 ka BP), human activities, particularly pasturing, and increasing temperatures, emerged as significant drivers of plant community dynamics. The introduction of sheep (Ovis aries) did not reduce wild mammal presence, including red deer (Cervus elaphus), hare (Lepus), European mole (Talpa europaea), bank vole (Myodes glareolus), and short-tailed field vole (Microtus agrestis). During the High & Late Mediaeval Period, (~1150–450 BP), cattle (Bos taurus) and horses (Equus caballus) became the dominant domesticates, while all wild mammals except the short-tailed field vole and Eurasian water shrew (Neomys fodiens) declined or even disappeared. These changes are also accompanied by a significant transformation of plant community structure. The ability to determine both plant responses and animal drivers from the same palaeolimnological sequence vastly improves our ability to partition causes of vegetation change over the Holocene. Here, we reveal that plant biodiversity is maintained or increased by moderate cattle grazing. Therefore, non-intensive domesticated stock grazing is essential for maintaining diverse Alpine meadows.
Introduction
The impacts of climate change on alpine plant communities are raising concern due to their potential repercussions on biodiversity, ecosystem functions and services (Liu et al., 2021; Steinbauer et al., 2018; Zu et al., 2021). The rate of climate warming in the European Alps is reaching up to three times the average observed in the northern hemisphere (Hock et al., 2019), which is predicted to result in biodiversity decline in the coming decades (Schwager and Berg, 2019). Pressures from human land use could potentially exacerbate these effects causing further losses (Wang et al., 2018). The Central Eastern Alps, recognized as a biodiversity hotspot harbouring numerous endemic species (Tribsch and Schönswetter, 2003), is now facing significant ecological threats due to both anthropogenic climate change and land use (Schwager and Berg, 2019). Long-term studies are needed to understand the drivers of such changes, but most palaeoecological studies so far do not allow for simultaneous statistical analyses of climate and land use effects.
Recent advances in sedaDNA (sedimentary ancient DNA) methods enable comprehensive reconstructions of the surrounding vegetation and husbandry dynamics using plant and animal DNA from lake sediment deposits, providing an opportunity for statistical testing to elucidate drivers of vegetation changes (Garcés-Pastor et al., 2022; Giguet-Covex et al., 2023; van Vugt et al., 2022). This enhanced understanding can provide pivotal insights into the preservation and management of these invaluable habitats for future generations, particularly regarding the impact of grazing in the management of Alpine ecosystems in the face of ongoing climate change.
From the Last Glacial Maximum (Local glacial history in Supplemental Information) to the Holocene climatic optimum (~9.5–5.5 ka BP (1000 years before present)), the Alps underwent a progressive increase in temperature (Fohlmeister et al., 2013). The short-lived 8.2 ka BP climatic event was characterized by generally cooler conditions (Tinner and Lotter, 2001), along with increased humidity and precipitation (Drescher-Schneider, 2007). Vegetation reconstructions indicate that the 8.2 ka BP event caused a restructuring of terrestrial plant communities (Tinner and Lotter, 2001). Following this event, the climate remained relatively stable until the end of the Holocene climatic optimum at ~5.5 ka BP.
Climate is not the sole driver of plant community dynamics. Throughout the Holocene, Alpine plant communities have faced significant impacts from human land use practices. These include early Holocene vegetation clearance through deliberate burning (~9 ka uncalibrated BP) (Bos and Urz, 2003; Lechterbeck and Rösch, 2021), the artificial lowering of timberlines resulting from Bronze Age (~4.5–3 ka BP) mining (~3.5 ka BP) (Knierzinger et al., 2020) and transhumance (Pini et al., 2017; Schmidl et al., 2005), to modern day recreational hiking (Aziz et al., 2023).
Although the Austrian Neolithic period began ~6.9 ka BP (Löcker et al., 2009), the earliest known human settlement phase in Styria (Figure 1b) dates to ~6.3 ka BP (Wildon Schlossberg) (Brandl et al., 2015). By this time, the adjacent province to the west (Salzburg) already had evidence of Neolithic activity at over 2000 m a.s.l (Gilck and Poschlod, 2019). Austria in general remained sparsely populated until the Late Neolithic (~4.6 ka BP). After which Styria experienced a significant population increase, becoming progressively more inhabited (Brandl et al., 2015). Extensive copper ore mining emerged across Styria during the Neolithic to Bronze Age transition ~4.5 ka BP (Knierzinger et al., 2021). This widespread mining activity led to significant local deforestation (Drescher-Schneider, 2007) and an increased local demand for animal products. In the Late Bronze/Early Iron Age, sheep (Ovis aries) alpine transhumance, the seasonal mobility of pastoral activities across altitudes, became a common practice (Saliari et al., 2020). Following this, during the Iron Age (2.8–2 ka BP), there was a shift in farming practices from sheep to cattle (Bos taurus) alpine transhumance. However, a few strongholds of sheep alpine transhumance remained in East Austria (Schmölcke et al., 2018). The cooler temperatures towards the end of this period likely decreased the viability of cattle farming, leading to a resurgence of sheep as the primary domesticate in Austria (Schmölcke et al., 2018). Throughout the period of the Western Roman Empire (2–1.5 ka BP), the prosperity in Austria was mainly derived from mining (stone, salt, metal ores) and agriculture (Schmidl et al., 2005). Facilitated by a warmer, more stable climate (Marx et al., 2018), farmers of this period took to a reinvestment in cattle alpine transhumance. To support both mining processes and cattle pasturing, further tree clearing was undertaken (Drescher-Schneider, 2007). Following the collapse of the Western Roman Empire ~1.5 ka BP, the Early Mediaeval Period (~1.5 to 1 ka BP) was a time of war and mass movement across Europe, but also of an unpredictable climate (Marx et al., 2018). At the onset of the Early Mediaeval Period, the cooler, wetter climate resulted in a return to sheep alpine transhumance in some areas across Austria (Marx et al., 2018). A new intensification of cattle alpine transhumance took place at the start of the High Mediaeval Period (~1 to 0.7 ka BP), once again causing significant habitat disturbances and plant community turnover (Drescher-Schneider, 1998) that has continued to the present day. This historical reconstruction is based on pollen and archaeological records. It creates a general narrative, but cannot provide species-specific and local biodiversity data for statistically testing the drivers of change: climate and grazing.
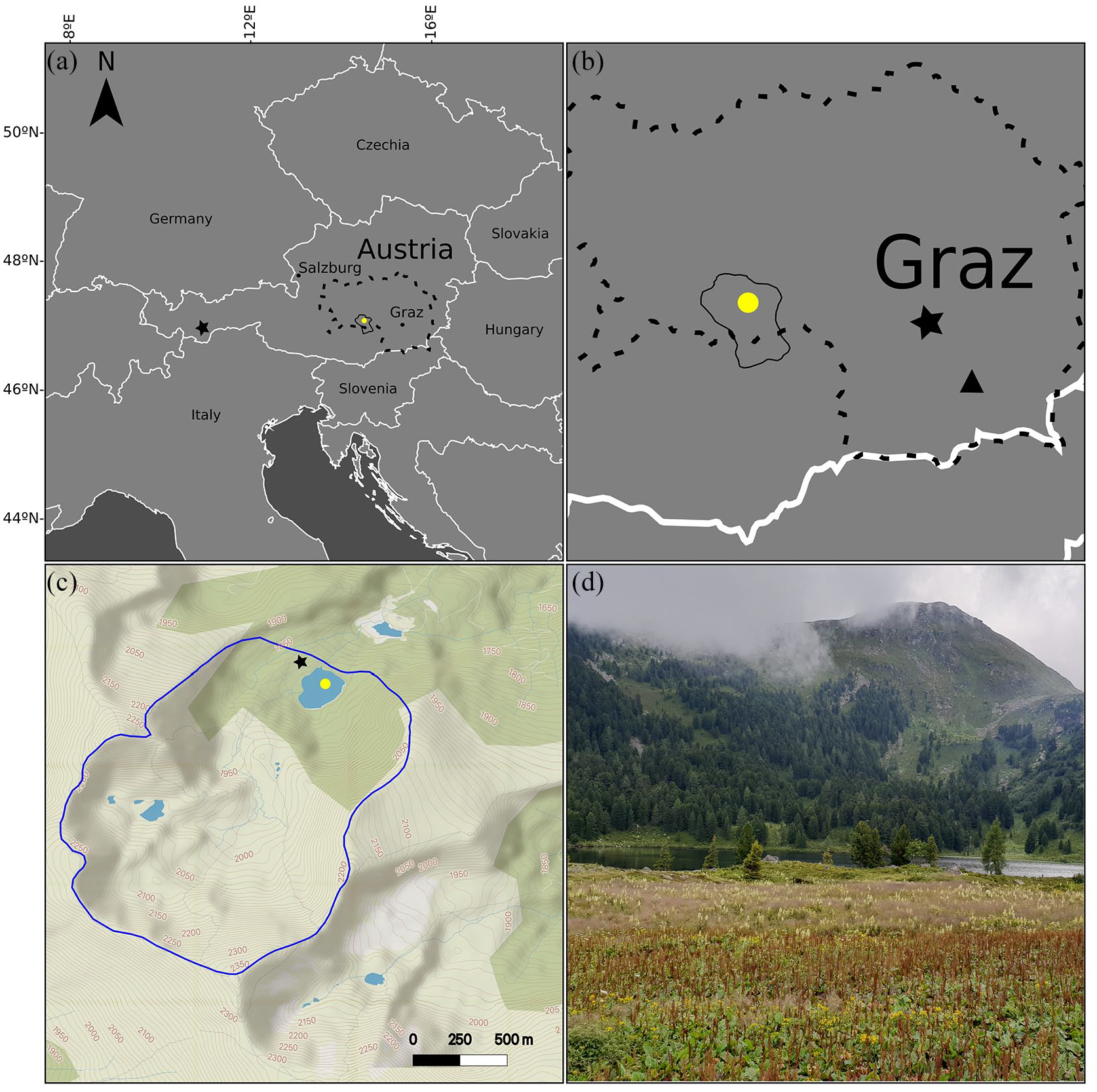
Maps and coring site: (a) A map of Europe centred around Austria, indicating Styria (dashed line), the Seetaler Alps (solid line), Großer Winterleitensee (yellow circle), and Schwarzsee ob Sölden (black star). Map drawn with vemaps.com basemap. (b) A zoomed in image of Styria, (dashed border) and the Seetaler Alps (solid border). The triangle indicates Wildon Schlossberg and the yellow circle, Großer Winterleitensee. Map drawn using vemaps.com basemap. (c) A map of Großer Winterleitensee with the catchment area (blue border), sediment core coring site (yellow circle) and moor core coring site (black star) indicated. Map drawn in QGIS using MapTiler Topo basemap. (d) Photo taken from downslope of the northwestern side of the lake (I. G. Alsos).
In this paper, we analyse plant and mammal sedaDNA from an Alpine lake (Großer Winterleitensee, Easternmost Central Alps) to identify species-level ecological histories and test the main drivers of plant diversity trends. We further aim to identify from when the landscape around Großer Winterleitensee has been used for Alpine transhumance, which animal species were the dominant grazers, and when, in order to resolve the debate concerning human impacts on Alpine biodiversity in the Eastern European Alps. This has further implications for the management of high-altitude Alpine landscapes in general. If biodiversity has increased through the grazing of domesticates then it follows that maintaining grazing is essential to sustain high biodiversity in the face of climate change.
Materials and methods
Großer Winterleitensee is a subalpine lake in Winterleitenkar, Seetaler Alpen, Austrian Alps (47.089653 N, 14.566058 E, 1845 m a.s.l., Figure 1), with an area of 4.5 ha, a maximum water depth of 9.6 m, and a pH of 6.98 (July, 2018). The lake catchment is 2.7 km2 and comprises a diverse landscape including mire, Alpine meadows and dwarf shrubs. On the steep south-eastern facing slope, there is a closed Swiss pine (Pinus cembra) forest with scattered spruce (Picea abies) and larch (Larix decidua) (Figure 1d). The lake is situated in the Styrian crystalline region, which is dominated by metamorphic rock (garnet, mica, schist) with rarer occurrences of plagioclase, gneiss, and pegmatite, and scattered marble and amphibolites on the north-west ridge above the lake (Zernig and Berg, 2018). Trees in this area can grow at a maximum elevation of 1950 m a.s.l, allowing for an elevational difference between lake and upper tree limit of c. 105 m. A moor core was taken within Winterleitenkar (47.0916667 N, 14.5638888 E, 1850 m a.s.l, Figure 1c) for pollen analysis (Zukrigl, 1975). However, with only two absolute dates provided (500 ± 130 and 3320 ± 160 BCE or ~2.5 and ~5.3 ka BP) we are unable to fully align this core with our lake sediment core.
A 4.6 m sediment core was taken from the deepest point of the lake from a floating platform using a Nesje coring system fitted with a 5 m long, 110 mm diameter pipe. For transportation, the core was cut into 1 m sections with the newly exposed ends immediately sealed. A surface sediment core was also taken with a UWITEC gravity corer using a 1 m long, 100 mm diameter pipe. All core sections were transported at 4°C to the Arctic University of Norway in Tromsø (UiT). Prior to opening, the core sections were scanned for magnetic susceptibility using a GEOTEK Multi Sensor Core Logger (MSCL-S) with a point sensor at the Department of Geosciences, UiT. The sections were then cut longitudinally and opened and sampled in a dedicated ancient DNA laboratory, UiT. We took 37 subsamples at 12 cm intervals for sedaDNA analysis from one core half, the other half was archived. During subsampling, samples were also taken for Loss on Ignition (LOI) and plant macrofossil remains were opportunistically collected for radiocarbon dating. A subsample from the sediment/water interface was collected from the surface core in a dedicated environmental DNA laboratory, UiT.
Radiocarbon dating, age-depth model construction and geochemistry
Sixteen terrestrial plant macrofossils were radiocarbon dated at the Poznan Radiocarbon Laboratory (Goslar et al., 2004) using accelerator mass spectrometry (AMS). The calibration of AMS dates used the terrestrial IntCal20 curve (Reimer et al., 2020). The age-depth model was constructed using the Bayesian framework calibration software ‘rbacon’ (v2.5.0) (Blaauw and Christen, 2011), executed in R version 4.3.1 (2023-06-16) (R Core Team, 2023).
The organic content of the sediment samples was measured using the LOI method by Lamb (2004). The archival core halves were scanned using an AVAATECH XRF core scanner with measurements taken at 10 mm resolution using a 1.5 mA current and 10 kV voltage for 10 s. High-resolution imaging was carried out using a Jai L-107CC 3 CCD RGB Line Scan Camera mounted to the XRF scanner. The raw peak area data was normalized using either Ti and Si in order to mitigate the influence of water and matrix effects (Croudace et al., 2006). Both elements are reliable indicators of input of allochthonous material from the catchment. Iron/titanium (Fe/Ti) is used to indicate changing redox conditions, phosphorus/titanium (P/Ti) is used as a weathering indicator and calcium/silicon (Ca/Si) is used as an erosion indicator (Davies et al., 2015). Magnetic susceptibility, measured using a GEOTEK Multi Sensor Core Logger, can be used as an indicator for precipitation (Balsam et al., 2011).
SedaDNA data generation and sequence assignment
All sedaDNA data generation steps mainly followed Rijal et al. (2021). Briefly, all 38 samples, four sampling controls, and four extraction negative controls were extracted using a modified DNeasy PowerSoil kit (Qiagen, Germany). Amplification of the samples used two uniquely dual-tagged generic primer sets. For plant sedaDNA, the trnL P6 loop region of the chloroplast genome was targeted using gh primers (Taberlet et al., 2007). For animal sedaDNA, we targeted the mitochondrial 16S locus using MamP007 primers (Giguet-Covex et al., 2014) including an updated human blocking primer strategy (Garcés-Pastor et al., 2022). Eight uniquely dual tagged (8 or 9-bp) amplicon replicates were created for both plant and animal sedaDNA. PCR reactions and cycling conditions for plants followed Voldstad et al. (2020), while animals followed Garcés-Pastor et al. (2022). We pooled and cleaned PCR products creating two pools, one for each primer set. A sequencing library was created for each pool and sequenced at Genomics Support Centre Tromsø, UiT on an Illumina NextSeq platform (2 × 150 bp, mid-output mode, dual indexing). Following data analysis (outlined below), plant negative control EG23_C003 was indiscernible from the plant sedaDNA samples, while plant sedaDNA sample EG23_087 contained no taxa (Supplemental Figure 4). We suspected a switch between the sample and control. We therefore re-extracted and re-amplified the batch of 20 samples (EG23_B series) in which the switch could have occurred. The resulting library was sequenced using an Illumina MiSeq platform (2 × 150 bp, mid-output mode, dual indexing) at the Faculty of Biosciences, Fisheries and Economics, UiT. For downstream data analyses, we combined the re-extracted samples with the remaining unaffected samples.
Following Rijal et al. (2021), plant sedaDNA data was processed using a bioinformatic pipeline based on the OBITools software package (Boyer et al., 2016). The reference databases PhyloAlps (Garcés-Pastor et al., 2022), Arctborbryo (Soininen et al., 2015; Sønstebø et al., 2010; Willerslev et al., 2014), PhyloNorway (Alsos et al., 2020, 2022) and EMBL release 143 (Kanz et al., 2005) were used. The identified sequences were filtered using custom R scripts (available at https://github.com/Y-Lammers/MergeAndFilter). Sequences were retained if they matched at 100% to at least one reference database, were present in at least three PCR replicates and had a read count of >10. Sequences that were detected in more replicates of the negative control samples than the sediment samples and/or are known common laboratory contaminants (Supplemental Table 3) were removed. Co-occurring sequences were assigned to the same taxon (Alsos et al., 2022). For the animal sedaDNA data, we used the aforementioned pipeline with adjustments outlined in Garcés-Pastor et al. (2022). The single reference database, EMBL release 143 was used. As the concentration of animal sedaDNA is typically lower than terrestrial plant sedaDNA (Murchie et al., 2023), we used more relaxed filtering criteria to maximize detections. Sequences were retained if they were present in at least one replicate, with a read count > 1, and a match of ⩾95% to the reference database. Co-occurring sequences assigned to the same taxon were merged. We discarded sequences matching human (Homo sapiens), whereas sequences matching invertebrates were not used in downstream analyses. Sequences identified as Sus scrofa were discarded as we are unable to distinguish domesticated pig from wild boar at this locus. One of the three unique pig sequences was also present in the extraction control at the same frequency as the sediment samples, leading us to believe this may be contamination from the extraction reagents. Sequences that were only present with a read count of one were also removed.
Climate reconstruction and statistical analysis
We reconstructed the Holocene climate around Großer Winterleitensee using local temperature and precipitation datasets. The temperature reconstruction is based on chironomid assemblage data taken from the Austrian lake Schwarzsee ob Sölden (SoS) (Ilyashuk et al., 2011; Wick and Tinner, 1997), a lake at 2796 m a.s.l, 951 m higher elevation than Großer Winterleitensee. We adjusted the temperature reconstruction using a mean environmental lapse rate of −6.5°C km−1 (Lute and Abatzoglou, 2021). Therefore, by adding 6°C to each SoS temperature point an approximate local temperature reconstruction appropriate for Großer Winterleitensee was created. We reconstructed precipitation at 100-year resolution using the CHELSA-TraCE21k v1.0 model (Karger et al., 2023). Precipitation (kg m−2) refers to the mean mass (kg) of water in all phases (rain, snow, etc.) per square metre per 100 years.
We plotted the proportions of weighted PCR replicates (wtRep) (Rijal et al., 2021) for plant and animal sedaDNA data using the Rioja package (Juggins, 2015) as implemented in R. This conservative measure of abundance avoids patterns of change being masked by read-dominant taxa and thereby highlights the diversity detected (Alsos et al., 2022). We conducted a constrained cluster analysis (CONISS) to identify statistically significant changes in the plant and animal community compositions using the Vegan package (Oksanen et al., 2013) in R.
Box plots were used to determine significant differences in total richness and individual plant growth form richness (Hill number for q = 0) with and without the presence of grazing animals. The data for cattle (Bos taurus), sheep (Ovis aries), horses (Equus caballus), goats (Capra hircus), and red deer (Cervus elaphus) were transformed into binary presence/absence data. Boxplots were plotted using the geom_boxplot() and the error bars were created and plotted using stat_summary() and stat_boxplot() from R package ggplot2 (Ginestet, 2011). The error bars indicate one standard deviation from each group mean. Plant richness is taken as the total count of taxa identified to family level and lower within each sample. Whereas the plant growth form richness is this data separated into each growth form, for example, tree, shrub etc.
To explore the effects of temperature, precipitation and animals (wtRep cow, sheep, horse, goat, red deer) on vegetation changes (plant wtRep data), a redundancy analysis (RDA) was performed using Vegan, and plotted using ggplot2. The drivers were checked for co-correlations that could impact the results using cor(). Horse was highly correlated with cattle, thus it was removed. Scaling of the RDA axes was set to two so the angle between the arrows reflects their relative relationship; an angle <90° representing a positive relationship, 90° a neutral relationship, and >90° a negative relationship. A transformation, such as the Hellinger transformation was not required as the data grouping had already reduced the ecological gradient. Model and constraint significance testing was achieved using anova.cca() in Vegan.
Results
Age-depth model and lithology
The sixteen AMS radiocarbon dates range between 250 ± 90 and 7490 ± 50 years BP (Supplemental Table 1). Two dates were rejected; one had an error margin of 110 years, and the other fell outside of the model. The age-depth model (Figure 2) displays a near linear sediment accumulation rate (SAR), well captured by the Bayesian model but also well described by two linear rates with a break-point at ~2.8 ka BP. At approximately this date the SAR increases 4.5× from 0.2 to 0.9 mm yr−1.
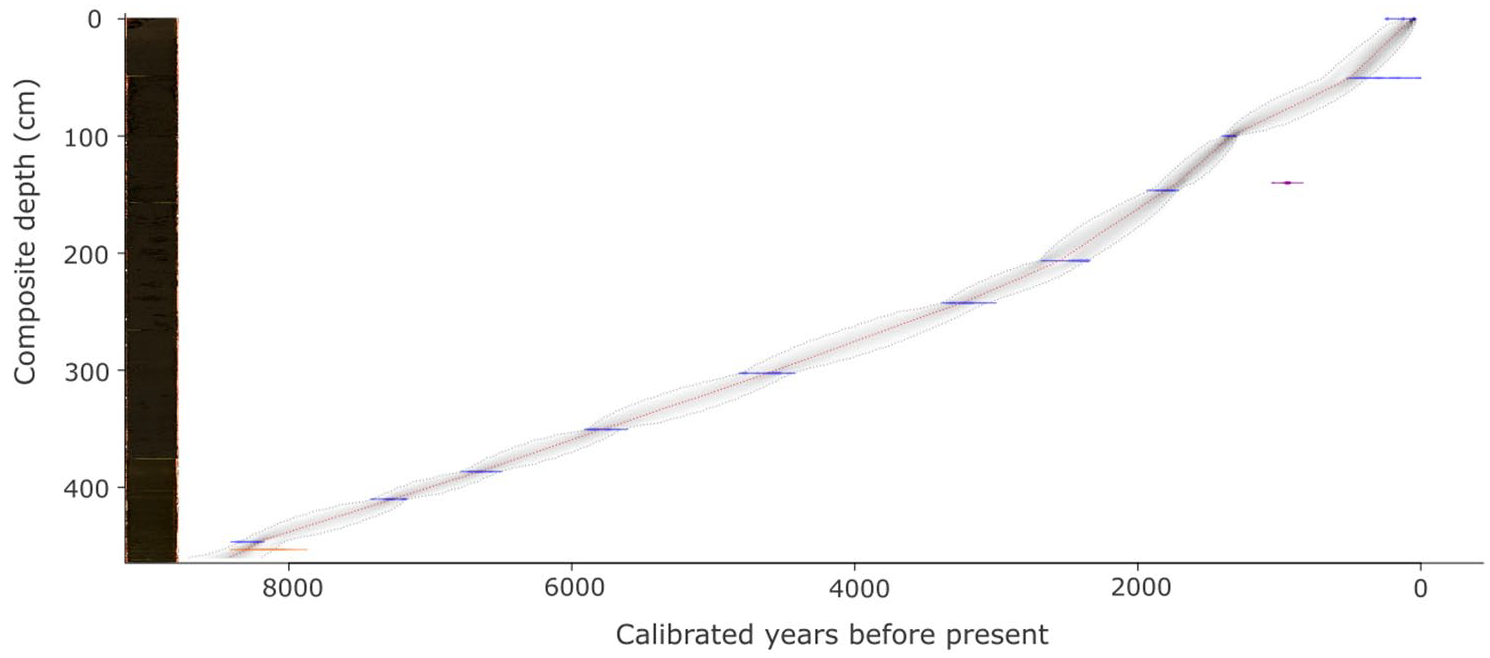
Age depth model, core image and lithology: The age depth model with the calibrated radiocarbon dates (one standard error) in blue. The age-depth relationship is illustrated by a curve: the most probable calendar ages (darker grey), the 95% confidence interval (grey stippled line), and the optimal model derived from a weighted average of the mean (red line). Two dates were discarded, one with an error margin > 100 years (orange) and the other lies outside of the model (purple). The core comprises dark-brown silty-gyttja and some lighter coloured bands throughout.
The core consists of dark-brown silty-gyttja with organic macro-remains and lighter coloured bands throughout (Figure 2). LOI (Figure 3) indicates a core rich in organic matter. In the oldest sample (8.5 ka BP), only 20% of the core comprises organic material. Over the following 3000 years, this percentage increases, culminating in a peak at 5.5–5.2 ka BP (44%) before plateauing and stabilizing around a mean of 43 ± 5%. The youngest sample however is composed of only 27% organic content.
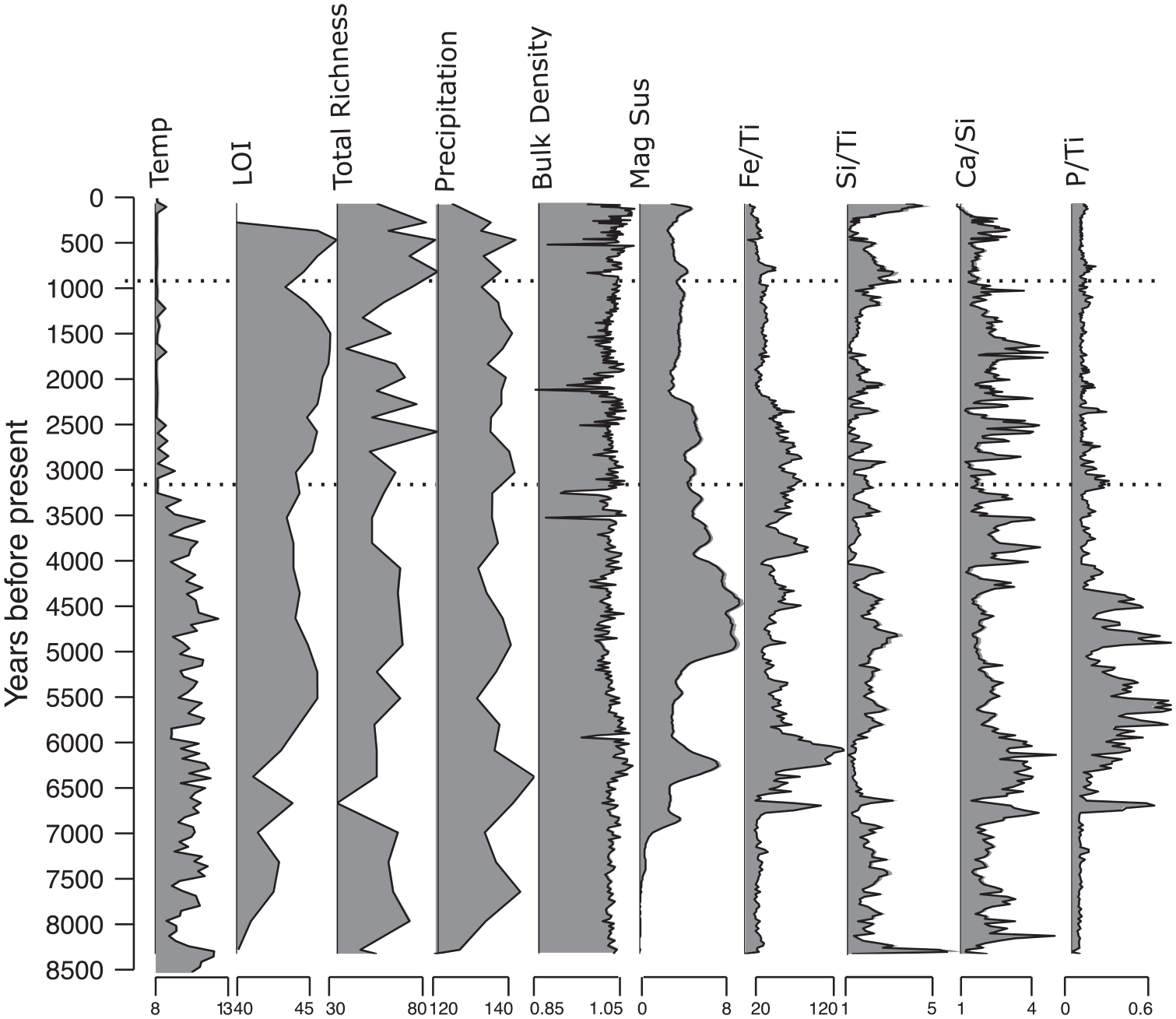
The XRF data with temperature and precipitation data: XRF data of the core taken from Großer Winterleitensee, Austria, alongside mean July temperature (Temp, °C), % loss on ignition (LOI), total plant richness (Hill where q = 0), mean precipitation (kg m−2) of water in all phases, bulk density, and magnetic susceptibility (mag sus). Full XRF data shown in Supplemental Figure 1.
XRF data (Figure 3) reveals an increase in Fe/Ti and Ca/Si input in the early Neolithic period ~6.8 to 6 ka BP, indicative of a period of increased weathering and erosion respectively. The ratio of P/Ti 6.5–4.5 ka BP also signals an alteration in precipitation patterns during this period. However, while a downturn in input takes place, Fe/TI and Ca/Si remain noisy throughout, suggesting that while the transport of material into the lake is still occurring, it is at a reduced rate than before, possibly due to decreased precipitation. The intervals at 2.4–2.2 (Iron Age) and 1.3–1 ka BP (Early Mediaeval Period) are intervals characterized by lower erosion as inferred from Ca/Si. Magnetic susceptibility of this core remains low until ~6.8 ka BP, subsequently peaking at ~4.5 ka BP and then gradually decreasing towards the upper section of the core, suggesting a decrease in precipitation regimes and/or a decrease in input of magnetic materials such as Fe.
Precipitation reconstruction
Modelled precipitation reconstruction data ranges from 117 (8.5 ka BP) to 144 kg m−2 (6.4 ka BP) (Figure 3) with the record starting with the lowest precipitation value (until 6.8 ka BP; mean = 129 kg m−2). This is followed by a peak with the aforementioned highest value and then decreases through the Neolithic period (6.9–4.4 ka BP; mean = 136 kg m−2). The mean precipitation remains relatively stable (mean = 134 kg m−2) until the Modern period, where precipitation decreases once more (600 years BP–present day; mean = 131 kg m−2).
SedaDNA
Plant data
A total of 1,642,012 reads with 100% match to at least one of the reference libraries were obtained after bioinformatic filtering (Supplemental Table 2). Following the collapse of homopolymers and post-identification filtering, we retained 1,626,989 reads, (99%) across 137 unique sequences: 11 identified to family-level, 30 to genus-level and 84 species-level (61%) and 12 to other taxonomic levels (sub-family, tribe, and sub-tribe). These taxa can be broken down into the following growth forms: 8 trees, 16 shrubs, 14 graminoids, 72 forbs, 8 ferns, 13 bryophytes and 6 aquatic plants (Figure 4). The number of taxa per sample ranged between 40 and 81 with a mean and median of 58.5 and 57 taxa respectively. Removed taxa and taxa that are consistently present throughout the core can be found in Supplemental Tables 3 and 4, respectively.
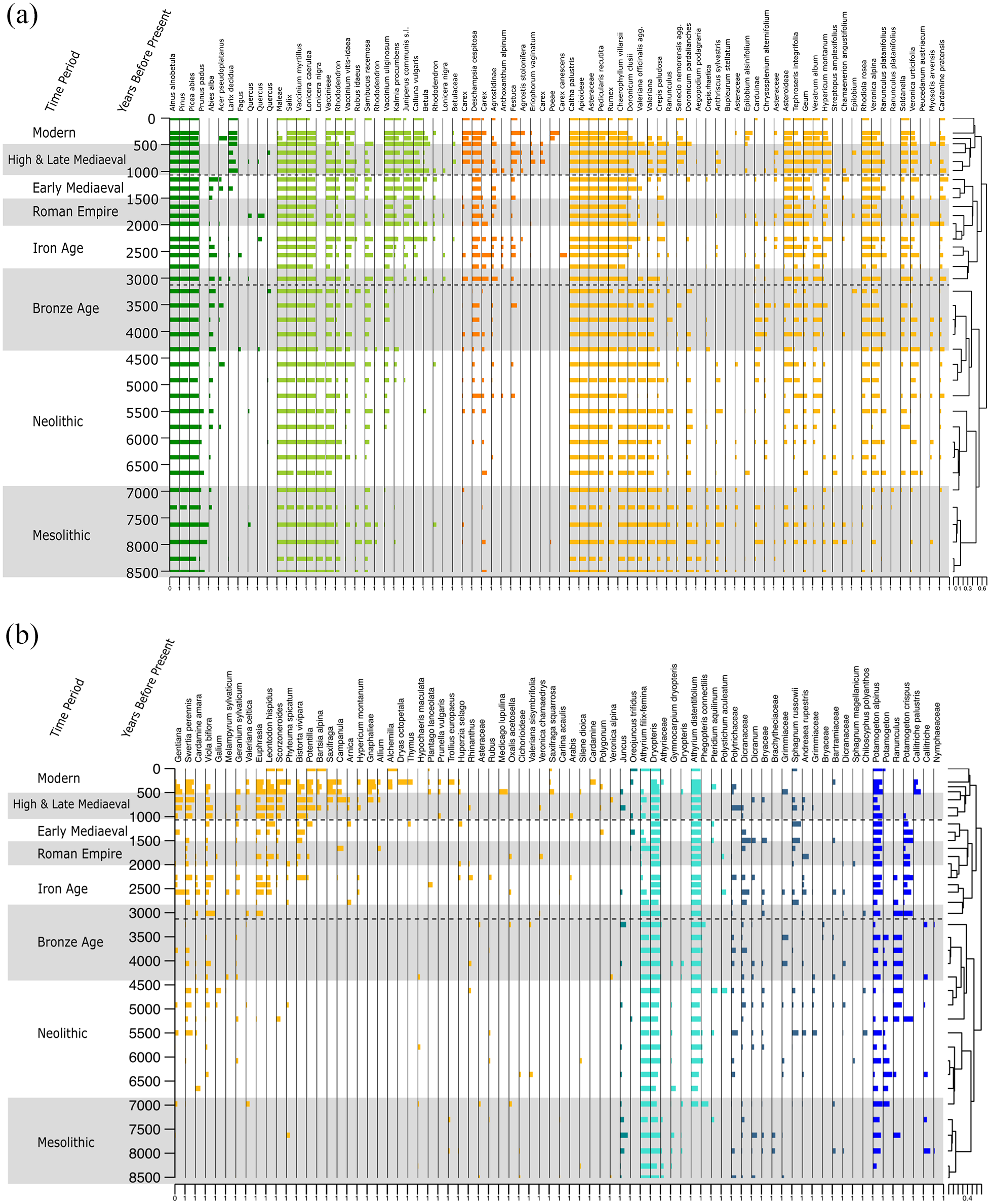
Plant sedaDNA diagram, split into two: A representation of the plant taxa present through time in years before present. Each taxon is represented by its proportion of weighted PCR replicates from the sedaDNA, where 0 represents that the taxon is present in no replicates and 1 represents that the DNA it is detected in all eight replicates. Data is plotted with CONISS statistical zonation using the plant sedaDNA data. Archaeological time periods are alternately shaded grey and white.
Animal data
The animal data comprises 84,062 reads identified with 95% or higher to 246 identified sequences (Supplemental Table 2). Fifteen unique sequences representing 15 taxa were retained after post-identification filtering and haplotype collapsing, comprising 65,152 reads: one taxon at genus-level and 14 at species-level. Whilst these are mammal specific primers, amphibian by-catch does occur. The 15 taxa can be broken down into the following groups: 4 domesticate mammals, 10 wild mammals and 1 amphibian. Red deer (Cervus elaphus) and, from 5.2 ka BP, common frogs (Rana temporaria) were detected consistently throughout the core, being absent in only a few samples across the record (Figure 5).
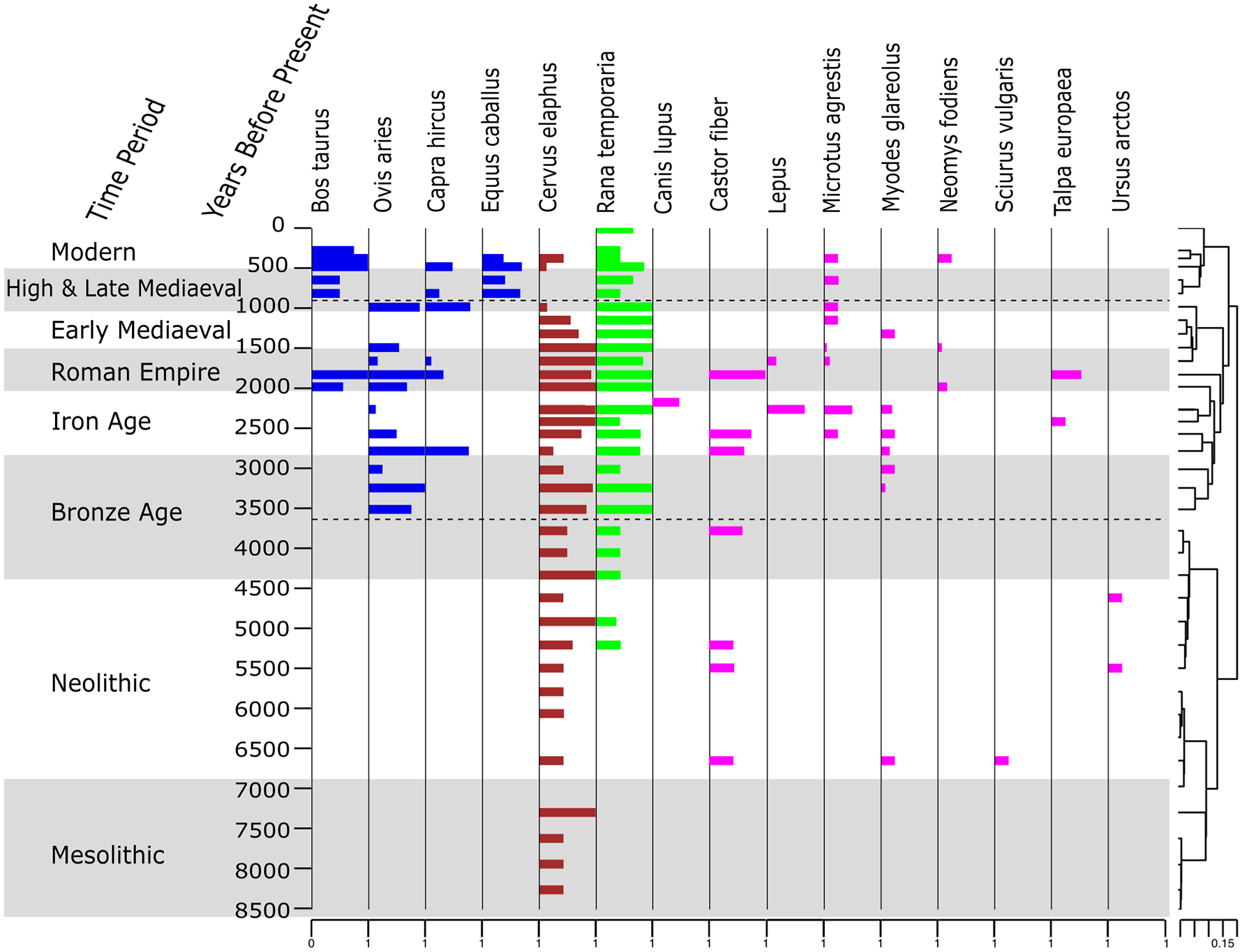
Vertebrate sedaDNA diagram. The proportion of weighted PCR replicates of the vertebrate sedaDNA data where 0 represents sedaDNA present in no replicates and 1 represents all eight replicates. Data is with CONISS statistical zonation using the vertebrate sedaDNA data. Archaeological time periods are alternately shaded grey and white.
CONISS zone analysis
Constrained cluster analysis (CONISS) using a broken stick model of both the plant and mammal data identified three statistically significant zones each according to main changes (Supplemental Figure 3). The breakpoints of the CONISS zones for these two data sets are similar. We chose to use the plant CONISS zones as these zones are based on more taxa than those of the mammal data, giving us more confidence in these zones. We call these zones Plant DNA Zones (PDZ) I–III (Figure 6). For comparison, the CONISS zones for the animals are plotted in Figure 5 (See direct comparison of zones in Supplemental Figure 3).
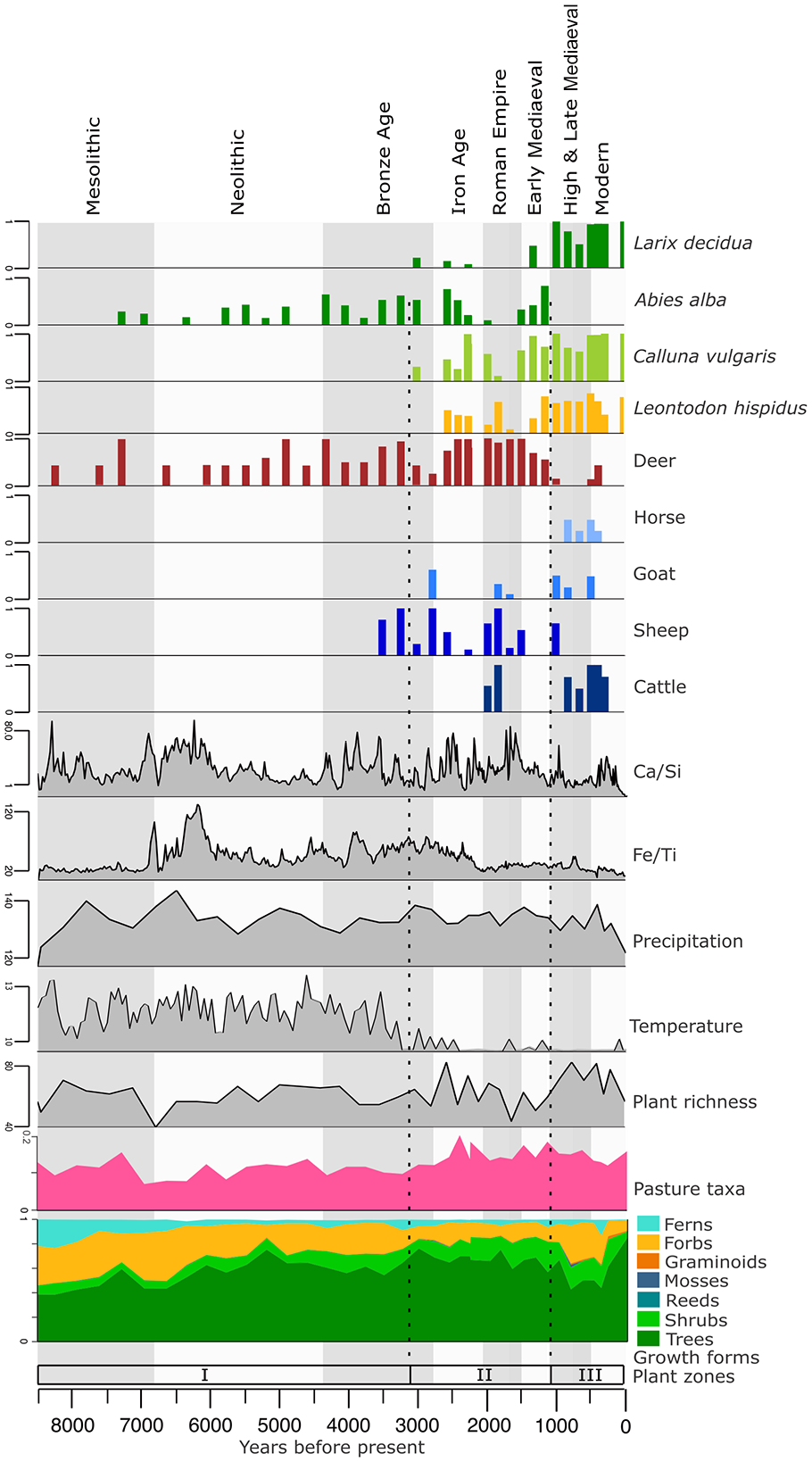
Overall figure: Cultural periods are indicated by the vertical grey and white boxes. Dotted lines are the zones (Plant DNA zones I–III) based on constrained cluster analysis of plant sedaDNA data. Plant taxa that are indicative of the changing environment are plotted with red deer (Cervus elaphus) and four domesticated taxa from the vertebrate sedaDNA data. Also plotted are Ca/Si and Fe/Ti, (indicators of erosion and weathering), precipitation (kg m−2), mean July temperature (°c), plant richness (Hill where q = 0), relative proportion of meadow related taxa, and the relative proportions of plant sedaDNA taxa reads separated by growth form.
PDZ I (459–235 cm, 8500–3010 years BP mesolithic, neolithic, bronze age)
PDZ I is characterized by unstable environmental conditions (Figure 6). At the core’s base around 8.5 ka BP, the models indicate a relatively high temperature (12.02°C), the lowest precipitation value for the record (117.08 kg m−2) and a median starting point for plant richness at 57 taxa. Pasture related plant taxa (Figure 6 and Supplemental Table 5) are present across the entire core, however PDZ I displays the lowest proportion (mean = 10 ± 2%).
Over the Mesolithic period, Fe/Ti remains stable and low suggesting low weathering. Ca/Si indicates variable inputs of material from erosion processes, while becoming more stable towards the end of the Mesolithic period. At the start of this core, the LOI values are at their lowest, possibly due to few aquatic species and/or less erosion at this time. Progressing through the Mesolithic period (until ~6.8 ka BP), the temperature is fluctuating, and the precipitation is highest in the middle of the period. Total plant richness (Hill q = 0, PDZ I mean = 54.5 ± 6.0) fluctuates in tandem with variations in precipitation. Ferns and forbs have high abundance (average 22% and 32%, respectively), whereas trees only constitute an average of 39% of the reads across these samples. As the climate warms and dries ~7.5 ka BP, the proportion of ferns decreases, while that of trees increases. Following a peak of pasture related taxa at 7.3 ka BP, these decrease over the subsequent samples.
The Neolithic Period (6.8–4.6 ka BP) begins with the highest precipitation levels of the core and corresponds with a peak in mineral in-wash (Fe/Ti, Ca/Si & P/Ti; Figures 3 and 6) into the lake and low abundance of trees. The proceeding drier, more stable climate through the Holocene climatic optimum is reflected in an increase in trees, the first appearance of beaver (Castor fibre) and bank vole (Myodes glareolus), as well as the only record of red squirrel (Sciurus vulgaris) at 6.8 ka BP (Figure 5). We note that red deer decreases during the early Neolithic but increases again during the Late Neolithic, when also brown bear (Ursus arctos) appears.
The cooling throughout the Bronze Age coincided with once again an opening of the forest environment with a decrease in trees and an increase in forb and shrub growth forms. In the final 500 years of PDZ I, while detecting the domesticated sheep (Ovis aries), there is an increase of trees and ferns once more with a reduction in shrubs and pasture related taxa suggesting, once again, a closing forest environment. With the exception of one detection at ~6.8 kyr BP, the bank vole is first detected in consecutive samples at ~3.25 kyr BP, coinciding with the aforementioned increase in forest environment.
PDZ II (235–105 cm, 3010–1020 years BP, iron age, Western Roman Empire, Early Mediaeval period)
PDZ II can be characterized as a period where human land use is having a profound effect on plant dynamics. Trees maintain dominance throughout PDZ II with the proportion of reads fluctuating around a mean of 68%. The early phase of PDZ II, is represented by the Iron age (2.8–2.2 ka BP) and plant richness (mean = 59.2 ± 9.2%) and temperature are both decreasing. An initial increase in shrub diversity includes Juniperus communis, Vaccinium uliginosum and Calluna vulgaris indicating habitat disturbance and open land creation, possibly from the pasturing of domesticates. The proportion of pasture related taxa across this zone (Figure 6; mean = 14 ± 0.02%) is highest at 2.5 ka BP at 19%. Pasture related taxa including Calluna vulgaris, and Leontodon hispidus are first detected between ~3 and 2.5 ka BP. Taxa that are present only sporadically prior to PDZ II, are detected more frequently and in more subsequent samples, for example, Deschampsia cespitosa, Juniperus communis s.l. The bank vole is detected from the start of PDZ II until the end of the Iron Age at 2.3 ka BP. The detection of goats (Capra hircus) at the Bronze Age/Iron Age boundary indicates the presence of more domesticate varieties potentially having a more profound effect on the vegetation than sheep alone. After the introduction of goats, the short-tailed field vole is detected in two samples (2.5 and 2.2 ka BP), before being detected once again from 1.7 ka BP until the end of this zone. The presence of Eurasian water shrew (Neomys fodiens), European mole (Talpa europaea), hare (Lepus sp.) and one instance of wolf (Canis lupus) can also be detected in this zone. Abies alba, is a disturbance sensitive species that is not detected when cattle are detected at this lake. The aquatic plant Potamogeton crispus is present almost exclusively in this zone. This species can persist in nitrogen-rich waters, allowing it to thrive in heavily farmed areas where more sensitive species cannot. The onset of the Iron Age (2.8–2 ka BP) sees an increase and then a gradual overall decline in plant richness, although there is no sharp change in plant community structure (Figures 4 and 6).
The climate during the Roman Empire has less precipitation relative to other time periods. However, towards the start of the fall of the Western Roman Empire, a warmer temperature is recorded. Plant richness is overall high through this period, coinciding with the detection of sheep, cattle (Bos taurus) and goats. At 1.8 ka BP, a peak in Ca/Si indicates increased erosion around the lake. The increased erosion, coupled with the detection of multiple domesticates, points to heightened human activity in the vicinity.
The Early Mediaeval Period (1.5–1.1 ka BP) has relatively higher precipitation and lower temperatures. It is characterized by a lower plant richness, however Abies alba is detected throughout. Of the domesticates, only sheep are present at the start of this time period. After this, sheep also are no longer detected in the record. Red deer however, maintain their presence around the lake.
PDZ III (105–0 cm, 1020–0 years BP, High & Late Mediaeval period, modern age)
PDZ III is defined by significant human disturbance and demarcates the discontinuation of Abies alba. Of the wild mammals, only red deer, short-tailed field vole, and the Eurasian water shrew are detected. Red deer however, are only detected in two samples in PDZ III. This could be due to an increased effect of human activity around the lake, suggesting that throughout previous domesticate grazing periods, the natural community structure has not been so greatly impacted. Pasture related taxa remain at around 14 ± 2% throughout this zone, indicating that the grazing of the domesticated animals is maintaining the openness of the alpine pasture and halting the expansion of the forest. LOI values are low at this time, possibly due to lower in-lake organic production as the nitrogen indicator Potamogeton crispus is no longer detected. Plant richness remains high throughout PDZ III.
At the start of the High & Late Mediaeval Period richness is increasing alongside forbs and graminoids which increase from 15% to 30% and >0.1% to 1%, respectively. Sheep and goat are detected during the start of the High & Late Mediaeval Period, with input of Ca/Si indicating increased erosion. while cattle and horse (Equus caballus) are detected later during the Modern Age. The proportion of tree reads decline, meaning that the relative biomass of trees around the lake is decreased, suggesting some tree clearing. This is in contrast to the previous periods of pasturing.
Drivers of changes
Box plots
The box plot (Figure 7) shows that forb richness is significantly higher when there are cattle present compared to when there are not. Samples are plotted with colour representing the different CONISS Zones. There are three outliers in the without cattle group, these three are all samples with sheep present.
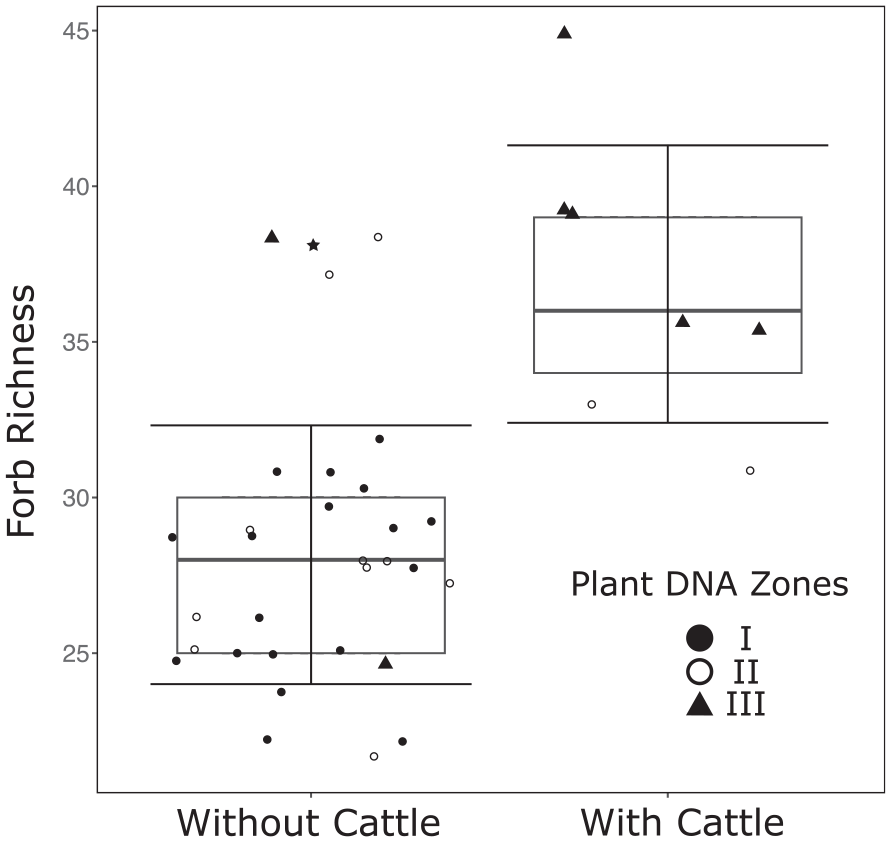
Boxplot: A visual representation showing the statistically significant difference in forb richness (Hill were q = 0) with and without the presence of cattle (Bos taurus). The Error bars indicate one standard deviation and the points represent the constrained cluster analysis Plant DNA Zones I–III. The outliers are indicated by the star.
RDA
The RDA (Figure 8) environmental variables (cattle, goat, sheep, red deer, temperature and precipitation) account for 35% of the variation in plant composition across samples. Only the variables cattle (F = 2.80, p = 0.01) and temperature (F = 9.51, p = 0.001) are shown to be significant; the other variables were not and cumulatively accounted for only 3% of the variation in the plant composition. All explanatory variables are included in this RDA plot to emphasize the patterns of the data, however significant variables are in bold. The cluster clearly separates samples belonging to the three different time periods, PDZ I–III. Both RDA axis 1 (24%) and RDA axis 2 (6%) are most affected by temperature and cattle. The non-significant grazers (sheep, horses and red deer) also fall on RDA axis 2, except goats, which falls on RDA axis 1. RDA axis 1 separates samples by plant CONISS zones (PDZ I-III), and therefore temporally. RDA 2 separates the samples by animal type showing that sheep cause a similar non-significant effect on the vegetation to that of red deer, and an opposing effect to that of cattle. Thus, cattle cause a different effect on the vegetation from that of large native mammals.
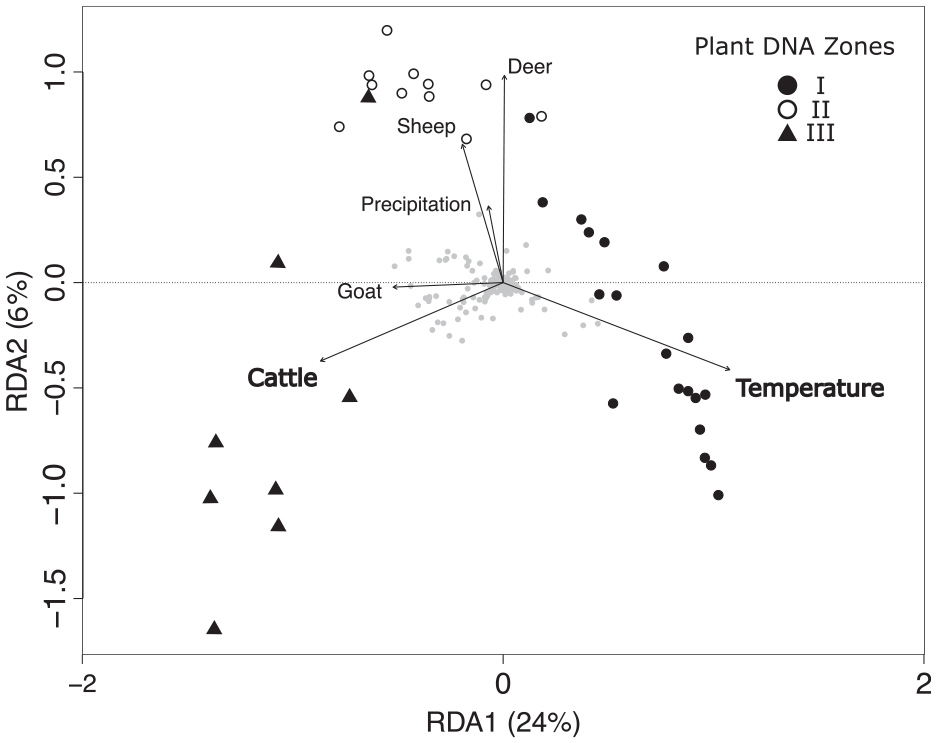
Redundancy Analysis (RDA): Redundancy analysis of the plant sedaDNA taxa (quantified as 0–8 weighted PCR repeats) composition across the explanatory variables representing environmental (precipitation (kg m−2), temperature (°C)) and animal factors. The sample points’ represents the constrained cluster analysis Plant DNA Zones I–III, the grey dots, the plant taxa. The significant variables that explained the plant diversity pattern are shown in bold.
Discussion
Dynamics before the detection of domesticated animals
Climate drove vegetation composition before the introduction of domesticated animals. Climate change is complex as there are different aspects to which plants can respond. Locally, at Großer Winterleitensee, and regionally across the wider Austrian Alps (Moser et al., 2005), precipitation does not have a significant impact on vascular plant richness. Precipitation however, does have a significant effect on plant richness at Sulsseewli (Garcés-Pastor et al., 2022). The Northern experience relatively high humidity that decreases as we move more towards the interior of the Alps, resulting in a more continental climate (Fink, 1993; Moser et al., 2005). While there is increased humidity in the Northern Alps, there is also greater variability in precipitation through the year causing pronounced drier months (Scherrer et al., 2022). Plant richness in environments with prolonged drier periods, like at Sulsseewli, is more likely to be affected by precipitation regime changes (Korell et al., 2021) than plant richness in environments with more stable regimes. Precipitation in the Seetaler Alpen is more evenly spread throughout the year (Hiebl and Frei, 2018) and therefore no relationship between precipitation and plant richness is found. We did not observe any strong increase in plant richness over time, in contrast to results from pollen records from the wider Alps (Giesecke et al., 2019) Taxa expansion occurred at different rates in the Central Eastern Alps relative to the rest of the Alps due to many factors including a more continental climate (Ilyashuk et al., 2011; Wick and Tinner, 1997), human impact (Latałowa and van der Knaap, 2006; Rey et al., 2013; Schwörer et al., 2015), and interspecific competition (Ravazzi, 2002). Pollen analyses revealed the expansion of Picea abies in the Eastern European Alps between 10 and 9 ka BP (Drescher-Schneider, 2007) compared to that of the expansion in the Western Alps at 6–5 ka BP (Latałowa and van der Knaap, 2006; Thöle et al., 2016). This is corroborated with P. abies detected from the start of the Großer Winterleitensee record at ~8.5 ka BP, contrasting with its late detection around ~6 to 5.5 ka BP (Garcés-Pastor et al., 2022; Thöle et al., 2016; Tinner and Theurillat, 2003) in the Swiss Alps. The mass expansion of P. abies into the Swiss alps was facilitated by anthropogenic activities such as fire, grazing and logging (Schwörer et al., 2015). Abies alba, a taxon that is limited by late frosts and low spring precipitation, is not detected at Großer Winterleitensee until ~7.4 ka BP, some 600 years delayed from presence at Lago Sangiatto in the Italian Alps (van Vugt et al., 2022). This suggests that the change in precipitation regimes and a more oceanic climate (Heiri et al., 2003) allowed for A. alba to establish successfully in the catchment of Großer Winterleitensee. Multiproxy palaeoecological studies in the Central and Western Alps suggest that summer high-altitude pasturing was already taking place by the Late Neolithic (Dietre et al., 2014, 2020; Garcés-Pastor et al., 2022; Schwörer et al., 2015). However, no evidence of this is found at Großer WInterleitensee. From the Middle Neolithic the tree reads decline, we can assume that relative biomass of the trees, and thus forest cover, around the lake is decreasing. The decline is corroborated by the local pollen core with tree pollen decreasing at one of the two given dates, ~5.3 ka BP (Zukrigl, 1975). This can be attributed to a known wetting (Zukrigl, 1975) and cold phase ~5.5–5 ka BP in the Alpine region (Ilyashuk et al., 2011; Thöle et al., 2016; Wick and Tinner, 1997). Plant richness was at a low during this period in agreement with general Alps pollen analyses showing a lower richness in Early compared to Middle and Late-Holocene (Giesecke et al., 2019). While some pasture related taxa are present around the lake, they are at their lowest level during this period. Indicating that before domesticates were introduced to pasture, a less diverse alpine meadow was already present. The consistent detection of wild grazers such as red deer, together with a rich forb community suggests that the wild grazer population densities were high enough to maintain the openness of such a meadow.
After the introduction of domesticated animals
The introduction of sheep around the lake during the late Bronze Age aligns with the notion that vertical alpine transhumance systems began during this period in the Eastern Alps (Gilck and Poschlod, 2019; Schmidt et al., 2002). By the Bronze Age, sheep products were integral for daily life (Grömer and Saliari, 2018). Wool was a valuable resource during the Middle Bronze Age, used for textiles and tradable goods (Schmölcke et al., 2018) and sheep meat and milk products were well established dietary staples for the communities of what is now Austria (Schmölcke et al., 2018). A shift in land use around Großer Winterleitensee is reflected in the change of sediment accumulation rate (SAR) from ~2.8 ka BP, the Bronze Age/Iron Age boundary. It coincides with an increased representation of domesticates in the sedaDNA record and significant vegetation change. Human land use, pasturing (Giguet-Covex et al., 2014; Rapuc et al., 2024), and increased precipitation (Arnaud et al., 2016) can increase be drivers of changing SAR. The opening of the landscape and greater pasturing could in increase erosion, thereby increasing the SAR and allowing more sedaDNA to be transported into the sediment from the wider catchment area. The delay in SAR increase from the first introduction of sheep during the Bronze Age, and no great change in precipitation indicate that domesticate pasturing and human disturbance is intensifying. Later, during the Roman Period, the presence of cattle, sheep, and goats are detected. The wealthy Romans in Central Europe were known for their advanced culinary culture and their riches afforded them the luxury of a varied and complex diet (Bakels and Jacomet, 2003) consisting of goat and sheep meat and milk products from all three animals. Wool manufacturing also played a crucial role in the Roman economy (Schmölcke et al., 2018), with what is now Austria becoming a centre for textile manufacturing (Gostenčnik, 2013). The leather of sheep, cattle and goats were used for garments throughout the Roman Empire, especially those worn by Roman military personnel (Grömer et al., 2017). All of these factors could have led to the intensification of alpine transhumance of these animals around Großer Winterleitensee. Towards the end of the Roman Period and into the Early Mediaeval period, tree clearing may be related to the mining of Noric iron and a large human population (Drescher-Schneider, 2007). During the High & Late Mediaeval Period, the abandonment of sheep farming after 1 ka BP coincided with the intensification of cattle farming and dairying, and the introduction of horses around the lake. Horses during this period were used for, among other things, rural heavy transportation (Henning, 2014) and could have been used to cart heavy items to and from the area around Großer Winterleitense, such as logs from tree clearance processes.
While there is an overall increase in plant taxa from pre to post introduction of domesticated mammal grazing, much of the local flora already had arrived prior to human alteration of the landscape. Conversely, in the Western Alps richness is increased greatly due to the presence of domestic animals (Thöle et al., 2016), especially cattle (Garcés-Pastor et al., 2022). At Großer Winterleitensee, there is a reorganization of the existing plant communities around the lake. The introduction of domesticated animals to an area can heavily modify vegetation and soil through several factors caused by animals at varying intensities; husbandry, grazing, excretion (fertilization) and trampling. Cattle and sheep have largely differing grazing methods, while sheep prefer to graze on forbs and close to the ground surface, cattle prefer to eat more moderately high level forage (Cutter et al., 2022). Another way in which cattle can facilitate forb growth is impeding the growth of trees and shrubs, thus diminishing the forest expansion (Wieczorkowski and Lehmann, 2022). We observed the highest mean total richness in the periods of cattle presence. The trampling of cattle has a strong influence on ground cover by forming bare patches (Hiltbrunner et al., 2012) suitable for the germination of seeds and the establishment of less dominant taxa such as pasture related taxa Leontodon hispidus and Calluna vulgaris (Mitchell et al., 2008). Seeds could arrive via wind transportation, but also deposited through cattle faecal matter (Traba et al., 2003) A potential way cattle presence caused a significant change to forb richness may be the larger body size and differing grazing patterns compared to both sheep and goats and the usually larger population density than the horses. Conversely, at Sulsseewli both sheep and cattle that showed a significant relationship with plant richness (Garcés-Pastor et al., 2022), possibly due to a higher sheep density than at Großer Winterleitensee. While both sheep and cattle have a trampling effect (Chai et al., 2019), the cattle would have a greater impact as they are larger and heavier than sheep (Yang et al., 2019). Despite the differences, both these lakes display that light to moderate grazing of either animal increases plant biodiversity.
Abies alba was not detected around Großer Winterleitensee at multiple periods, suggesting significant human disturbance likely by tree removal, browsing from domesticates, and forest regeneration prevention. Abies alba germinates within thick humus layers, so regenerates best in forest environments. It is also a browsing intolerant taxon, where dense populations of even wild ungulates can significantly hinder its growth and subsequent regeneration (Frei et al., 2024; Kupferschmid et al., 2015; Unkule et al., 2022). While A. alba populations demonstrate resilience mostly to the presence of sheep, there is a disappearance of A. alba once at 2.8 ka BP and then every time cattle are present. The A. alba population declines in the Bernese Alps, Switzerland (Rey et al., 2013; Schwörer et al., 2015) at 5.5 ka BP, attributed to anthropogenic disturbances such as fire, grazing and logging, and the expansion of Picea abies, a taxon that is ever present around Großer Winterleitensee. In the Lepontine Alps, Italy (van Vugt et al., 2022) the A. alba population declines and 5.1 ka BP, also attributed to intense human land use. The final collapse of A. alba at Großer Winterleitensee is ~1 ka BP coinciding with heavy disturbance and tree removal around the lake for grazing of domesticates. Larix decidua is seen to establish a persistent population around Großer Winterleitensee instead, at ~1 ka BP. Normally, L. decidua stands would occur where soil is exposed, as the seeds of L. decidua are dispersed by wind and thus are very efficient at colonizing bare ground and disturbed habitats (index of colonizing success of 7/10) (Prach et al., 2017). While L. decidua is an efficient colonizer and more robust against disturbances than A. alba, it was also spread by humans to form larch forest-meadows for grazing and timber production (van Vugt et al., 2022). Species-rich L. decidua stands were already detected by the Bronze Age in the neighbouring Bernese Alps, Switzerland (Rey et al., 2013) and Lepontine Alps, Italy (van Vugt et al., 2022). While a smaller presence of L. decidua is detected around the lake at these times, a continued strong detection of L. decidua occurs much later around Großer Winterleitensee, in the High & Late Mediaeval Period, suggesting an extremely long delay in the creation of these stands.
Wild animals around the lake
Contrary to subalpine lake Sulsseewli record (Garcés-Pastor et al., 2022) the introduction of domesticated animals did not coincide with the disappearance of red deer (Cervus elaphus). This may be since the domestic grazing pressure around Großer Witnerleitensee was relatively low during this period, unlike Sulsseewli. Thus, enough tree coverage was maintained, or habitat disturbance was not intense enough, and the red deer population was able to maintain a presence at the lake. From the High & Late Mediaeval Period, ~1 ka BP, the red deer are no longer detected. Red deer live between woodland and open grassland areas, suggesting the intensification of farming and escalation of tree removal forced the red deer to retreat from this lake, and it did not fully re-establish or the population was kept low due to hunting. Other wild animals are also affected by the plant growth forms around the lake. The bank vole’s preferred habitat is forests with high canopy and shrub cover (Hille and Mortelliti, 2011) while the field vole’s preferred habitat is forests with dense herbaceous cover and wet meadows (Mathias et al., 2017). We see that the bank vole disappears from the record during the Roman Period, however the field vole remains through the intensification of human land use.
Implications for conservation
The preservation of Alpine meadows has become increasingly important in recent years with respect to climate change and Alpine pasture abandonment (Piccinelli et al., 2020; Thöle et al., 2016). The conservation of Alpine pastures not only directly impacts plant biodiversity, but also other organisms dependent upon the open grassland (Cutter et al., 2022). In the past, prolonged heavy grazing has led to significant harm, causing impoverished soils and decreased biodiversity (Cislaghi et al., 2019). However, the undergrazing, or complete abandonment of these pastures leads to an expansion of trees and progressive ecological succession to closed forest (Cislaghi et al., 2019). Compounding this, increasing temperatures are further promoting the growth of trees (Snell et al., 2022) and therefore leads to a decrease in plant biodiversity. Our data shows that the light to moderate grazing of cattle increases not only forb richness, but also total plant richness. This is in agreement with the intermediate disturbance hypothesis which suggests that moderate levels of habitat disturbance allows for highest plant species diversity (Hobbs and Huenneke, 1992). Furthermore, it shows that grazing by sheep has a similar effect on the plant diversity to that of red deer. Moderate levels of cattle or sheep alpine transhumance should be continued to encourage the maintenance of these pastures and upkeep of plant biodiversity. Especially in a manner that could help mitigate the promotion of climate change on tree growth.
Conclusion
This sedaDNA-based research from Großer Witnerleitensee emphasizes the importance of interdisciplinary approaches to unravel complex ecosystem dynamics over long timescales. By using a multi-proxy approach, we are able to tease-apart and evaluate the impacts of climate change, native mammals and alpine transhumance on the vegetation at high taxonomic resolution. Our study represents the first in the Eastern European Alps identifying how and when alpine transhumance occurred using sedaDNA and DNA-barocoding technology. Our findings demonstrate that moderate-intensity cattle grazing significantly enriches the plant community. The insights gained from this research contributes to a deeper understanding of the historical and ongoing drivers of Alpine plant communities, thereby providing a foundation for informed conservation efforts and sustainable, evidence-lead, management practices.
Supplemental Material
sj-docx-1-hol-10.1177_09596836241307304 – Supplemental material for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps
Supplemental material, sj-docx-1-hol-10.1177_09596836241307304 for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps by Scarlett Zetter, Sandra Garcés-Pastor, Youri Lammers, Antony G Brown, Kevin Walsh, Tomasz Goslar, Sébastien Lavergne, Eric Coissac, PhyloAlps Consortium, Andreas Tribsch, Peter D Heintzman and Inger Greve Alsos in The Holocene
Supplemental Material
sj-eps-2-hol-10.1177_09596836241307304 – Supplemental material for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps
Supplemental material, sj-eps-2-hol-10.1177_09596836241307304 for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps by Scarlett Zetter, Sandra Garcés-Pastor, Youri Lammers, Antony G Brown, Kevin Walsh, Tomasz Goslar, Sébastien Lavergne, Eric Coissac, PhyloAlps Consortium, Andreas Tribsch, Peter D Heintzman and Inger Greve Alsos in The Holocene
Supplemental Material
sj-eps-3-hol-10.1177_09596836241307304 – Supplemental material for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps
Supplemental material, sj-eps-3-hol-10.1177_09596836241307304 for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps by Scarlett Zetter, Sandra Garcés-Pastor, Youri Lammers, Antony G Brown, Kevin Walsh, Tomasz Goslar, Sébastien Lavergne, Eric Coissac, PhyloAlps Consortium, Andreas Tribsch, Peter D Heintzman and Inger Greve Alsos in The Holocene
Supplemental Material
sj-eps-4-hol-10.1177_09596836241307304 – Supplemental material for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps
Supplemental material, sj-eps-4-hol-10.1177_09596836241307304 for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps by Scarlett Zetter, Sandra Garcés-Pastor, Youri Lammers, Antony G Brown, Kevin Walsh, Tomasz Goslar, Sébastien Lavergne, Eric Coissac, PhyloAlps Consortium, Andreas Tribsch, Peter D Heintzman and Inger Greve Alsos in The Holocene
Supplemental Material
sj-eps-5-hol-10.1177_09596836241307304 – Supplemental material for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps
Supplemental material, sj-eps-5-hol-10.1177_09596836241307304 for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps by Scarlett Zetter, Sandra Garcés-Pastor, Youri Lammers, Antony G Brown, Kevin Walsh, Tomasz Goslar, Sébastien Lavergne, Eric Coissac, PhyloAlps Consortium, Andreas Tribsch, Peter D Heintzman and Inger Greve Alsos in The Holocene
Supplemental Material
sj-eps-6-hol-10.1177_09596836241307304 – Supplemental material for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps
Supplemental material, sj-eps-6-hol-10.1177_09596836241307304 for SedaDNA shows that transhumance of domestic herbivores has enhanced plant diversity over the Holocene in the Eastern European Alps by Scarlett Zetter, Sandra Garcés-Pastor, Youri Lammers, Antony G Brown, Kevin Walsh, Tomasz Goslar, Sébastien Lavergne, Eric Coissac, PhyloAlps Consortium, Andreas Tribsch, Peter D Heintzman and Inger Greve Alsos in The Holocene
Footnotes
Author contribution(s)
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We thank Kristin Heggland (Tromsø) and Roland Kaiser (Salzburg) for invaluable assistance with fieldwork and Karina Monsen (Tromsø) for assistance with the geological measurements. Scarlett Zetter, Sandra Garcés-Pastor, Peter D. Heintzman and Inger G. Alsos were supported by a Research Council of Norway grant 250963/F20 for the ECOGEN project. Inger G. Alsos was also supported by The European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme grant agreement no. 819192 for the IceAGenT project. Sandra Garcés-Pastor was also supported by the Beatriu de Pinós Programme (BP-2021-00131) and a fellowship from “la Caxia” Foundation (ID 100010434, fellowship code LCF/BQ/PI24/12040011). Peter D. Heintzman acknowledges support from the Knut and Alice Wallenberg Foundation (KAW 2021.0048 and KAW 2022.0033). Andreas Tribsch was supported by ABOL (Austrian Barcode of Life) via an HRSM-project supported by the Bundesministerium für Bildung, Wissenschaft und Forschung (BMBWF) in Austria.
Consortia
The PhyloAlps Consortium, Sébastien Lavergne, Eric Coissac, Charles Pouchon, Cristina Roquet, Wilfried Thuiller, Niklaus E. Zimmermann, Adriana Alberti, Patrick Wincker, Martí Boleda, Frédéric Boyer, Anthony Hombiat, Christophe Perrier, Rolland Douzet, Jean-Gabriel Valay, Serge Aubert, France Denoeud, Bruno Bzeznick, Ludovic Gielly, Pierre Taberlet, Delphine Rioux, Céline Orvain, Maxime Rome, Rafael O. Wüest, Sonia Latzin, John Spillmann, Linda Feichtinger, Jérémie Van Es, Luc Garraud, Jean-Charles Villaret, Sylvain Abdulhak, Véronique Bonnet, Stéphanie Huc, Noémie Fort, Thomas Legland, Thomas Sanz, Gilles Pache, Alexis Mikolajczak, Virgile Noble, Henri Michaud, Benoît Offerhaus, Cédric Dentant, Pierre Salomez, Richard Bonet, Thierry Delahaye, Marie-France Leccia, Monique Perfus, Stefan Eggenberg, Adrian Möhl, Bogdan-Iuliu Hurdu, Paul-Marian Szatmari, Mihai Puşcaş, Jan Smyčka, Patrik Mráz, Kristýna Šemberová, Michał Ronikier & Marek Slovák.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.