
Abstract
The Central Highlands region of Madagascar is currently dominated by open and mosaic ecosystems, and the region is targeted for afforestation projects. At present, little is known about the origin of these ecosystems and whether or not they are ancient or anthropogenically derived mosaics. This paper provides new insights into the environmental history and drivers of change in the Central Highlands of Madagascar during the last 6300 years through palaeoecological records from Lake Dangovavy. Our results suggested that open and mosaic ecosystems have been present in the landscape for at least 6000 years that is, 4 cal. ka BP before human settlement (2 cal. ka BP). Between 6.3 and 2 cal. ka BP landscapes were characterized by a mosaic ecosystem dominated by a matrix of open montane grassland and ericoid shrubland with extent forest patches, associated with seasonal variation of rainfall, and fire. A matrix of ericoid shrubland remained abundant in the landscape despite an increase of fire regime and herbivory activities between 2 and 1.1 cal. ka BP. Major changes in the vegetation were recorded during the last millennium with a shift towards a more open C4-dominated grassland associated with high fire frequency and herbivory activities, most likely linked to human influence. Our findings support the notion of the natural presence of open ecosystems on the islands. These shed light on the landscape history which should be considered when setting conservation goals with a particular focus on areas targeted for tree plantations and fire bans like the Central highlands of Madagascar.
Keywords
Introduction
Open and mosaic ecosystems harbour a wealth of biodiversity yet are often undervalued in biodiversity conservation (Bond and Parr, 2010; Veldman et al., 2015). These ecosystems are often assumed to be secondary degraded forests, which makes them targets for afforestation and other carbon storage schemes. However, there is growing evidence showing that many open and mosaic ecosystems are natural and existed long before any human interventions, supporting high biodiversity richness, as well as soil carbon stocks (Bond et al., 2019; Fernandes et al., 2016; Tölgyesi et al., 2022; Veldman et al., 2015). Distinguishing ancient from derived ecosystems has important management and conservation implications. Misclassification of open and mosaic ecosystems as degraded forests can lead to neglect of their conservation and subsequent loss of unique biodiversity (Bond et al., 2019).
Paleovegetation studies of open and mosaic ecosystems in the tropics and subtropics remain scarce and it is therefore difficult to distinguish ancient from anthropogenically derived mosaics. Focussing on Madagascar, a tropical world biodiversity hotspot, mosaic and open ecosystems, particularly grasslands, are currently abundant, but their origin and history are debated. One area of particular controversy is the Central Highlands of Madagascar, which is dominated by grasslands with localized forest patches mainly confined to ravines (Vorontsova et al., 2016). The nature and origin of the landscape are not clear, and it is still debated whether these grasslands are natural or anthropogenically derived (Bond et al., 2008, 2023; Lowry et al., 1997; Solofondranohatra et al., 2018; Vieilledent et al., 2018; Vorontsova et al., 2016). Some authors argue that open ecosystems were naturally maintained by natural fire and herbivores (Hansford and Turvey, 2022; Solofondranohatra et al., 2020), while others argue that they are the result of anthropogenic activities associated with the use of fire and pastoralism (Erdmann, 2003; Vieilledent et al., 2018).
Humans may well have existed in Madagascar since 10,000 years ago (Hansford et al., 2018); but the timing of human settlement was only recorded at ca. 2 cal. ka BP (Burney et al., 2004; Douglass et al., 2019; Pierron et al., 2017; Radimilahy and Crossland, 2015). The few existing palaeoecological records from the highlands have demonstrated the abundance of ericaceous heathland in the last glacial (Samonds et al., 2019) and open ericoid grassland during the Early Holocene (Burney, 1987c; Straka, 1996). Herbivore fossils were abundant in the region, as recorded at the Tsaramody site (Samonds et al., 2019), and fire was suggested to be more frequent during the Early Holocene (Burney, 1987b). The complex interplay between climate, and fire and their impact on open ecosystems is still not well understood (Burney, 1987b; Gasse and Van Campo, 1998, 2001). The interaction between fire, herbivores, climate, human activity and shifting vegetation patterns remains tentative in the region. More information on the origin and history of the landscapes of the highlands, as well as their drivers of change, particularly prior to human settlement, is therefore urgently needed in order to distinguish between natural and anthropogenically derived ecosystems and to guide the management of this area.
Compiling high-resolution evidence that would allow reconstruction of environmental change in the Central Highlands of Madagascar is critical to understanding the nature and drivers of change of the current open and mosaic ecosystems and therefore their conservation management. Here, we present a high-resolution reconstruction of Holocene environmental changes in the ecosystems of the Central Highlands of Madagascar using a sediment core from Lake Dangovavy in the Vakinankaratra region. Specifically, we analyse multiple proxies to reconstruct past vegetation, fire history and herbivore activities using fossil pollen and stable carbon isotopes, microscopic charcoal and dung fungal spores, respectively. The following research questions are addressed in this paper:
(1) How dynamic were ecosystems around Lake Dangovavy from the Mid-Holocene to the present?
(2) How did fire and herbivory activities change over time prior to and after human settlement?
(3) How did vegetation respond to changing fire regimes, and herbivory activities before and after human settlement?
(4) What implications do the findings have for conservation and management?
Methods
Study area
Lake Dangovavy is a small lake of approximately 600 m wide and 4 m deep situated at −19.287569, 46.857446, and 1700 m a.s.l. near Faratsiho in the Vakinankaratra region of the Central Highlands of Madagascar (Figure 1). The region is characterized by a subhumid climate alternately affected by cool and dry winters, and warm and wet summers (Burgess et al., 2004; Rajeriarison and Faramalala, 1999). It has a mean annual temperature of about 20°C and receives a mean annual rainfall between 1200 and 2000 mm (Razafindratsima, 2019). The substrate is characterized by ferralitic and hydromorphic soils (Sourisseau et al., 2016). Modern vegetation in the region is characterized by the dominance (ca. 65%) of grasslands with some fragmented patches of woodlands and subhumid forests at high and mid-elevation (Burgess et al., 2004; Rabarivola et al., 2019) forming a wooded grassland-bushland mosaic (Moat and Smith, 2007) (see Figure 1). Surrounding Lake Dangovavy, the vegetation is dominated by grassland (particularly, the C4 grass Aristida rufescens, Poaceae) with some shrubs and vestige of trees. Shrubs were represented by Elephantopus scaber, Helichrysum sp. (Asteraceae), Agauria polyphylla, Erica goudotiana, Vaccinium secundiflorum (Ericaceae) and Weinmannnia sp. (Cunoniaceae). The trees are represented by Ilex mitis (Aquifoliaceae), and Aphloia theiformis (Aphloiaceae), Kotschya perrieri (Fabaceae), Syzygium jambos, Syzygium sp. (Myrtaceae), Maesa lanceolata (Primulaceae) and Tina striata (Sapindaceae). Individual native Uapaca cf. bojeri trees are found in the upland, as well as plantations of exotic trees, such as Acacia spp., Eucalyptus spp. and Pinus spp.
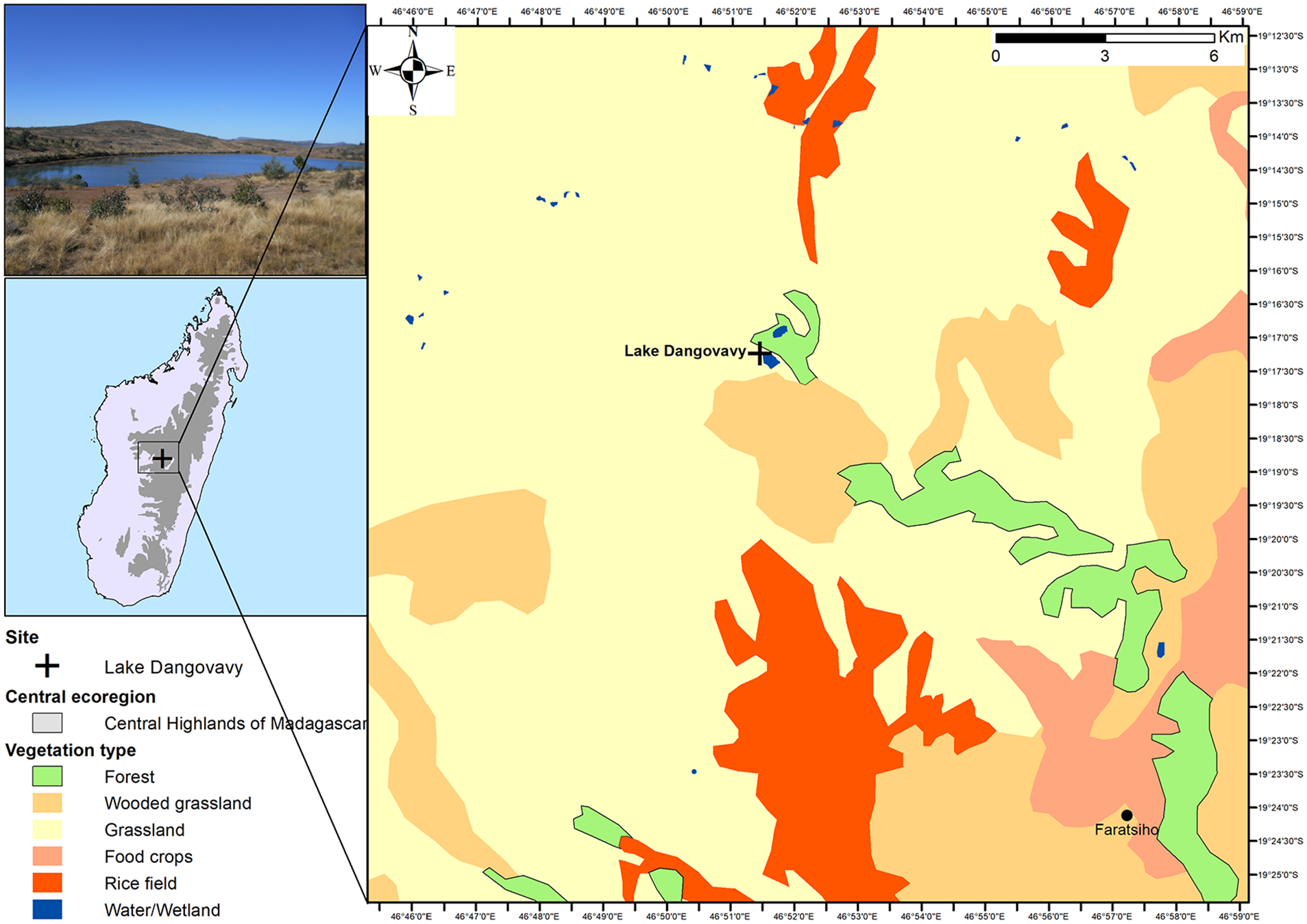
Location of Lake Dangovavy. Ecoregion adapted from Nusbaumer et al. (2010); and vegetation cover based on Moat and Smith (2007).
Sampling and dating
Using a Russian corer, a 110 cm long sediment core was collected 150 m from the edge of Lake Dangovavy at an approximate 1.4 m depth of water (Aaby and Digerfeldt, 1986; De Vleeschouwer et al., 2010). The lithology of the sediment core was described using a simplified version of Troels-Smith (1955) and the Munsell soil colour charts (Munsell Colour Company, 1954). Subsampling was conducted at 4 cm intervals for pollen, charcoal, and dung fungal spore analyses.
Accelerator mass spectrometry (AMS) 14C dating was conducted in seven (07) bulk sediment samples at different depths to describe the chronology of the sediment core (Table 1). The choice to date the bulk sediment was driven by its strong correlation with the pollen deposit, which serves as the primary proxy in this research, even though macrofossil plants were also present. Samples were pre-treated with acid for carbonate removal and sent to the iThemba LABS facility in Johannesburg (South Africa) and Beta Analytic Inc., Miami (United States of America). The dates were calibrated to years before the present (cal. years BP) using the Southern Hemisphere calibration curve ‘SHCal20’ (Hogg et al., 2020) within the open-source statistical software programme R v.4.3.0 (R Core Team, 2023). The age-depth models were conducted using the package ‘rbacon’ v. 2.5.8 (Blaauw and Christen, 2011) in R v.4.3.0 (R Core Team, 2023). Bacon age models help to find outliers in the data by reconstructing the accumulation histories of sedimentary deposits. The chronology was used to identify sediment accumulation change over time and outliers.
Uncalibrated and calibrated radiocarbon dates from Lake Dangovavy.

Indicates dates that are excluded from the age-depth model and considered as outliers.
Pollen, stable carbon isotopes (δ13C), stable carbon and nitrogen isotope composition (C/N), charcoal and dung fungal spore analyses
Pollen and stable carbon isotope (δ13C) analyses were used to understand vegetation dynamics in the area over time. Pollen analysis was based on the standard procedures using HCl, NaOH, HF and acetolysis (Bennett and Willis, 2001; Ferguson et al., 1976). Lycopodium tablets with a known concentration (9666 per tablet) were added to each sample prior to the analysis to estimate concentration and influx (Stockmarr, 1971). A minimum standard count of 300 pollen grains (excluding Cyperaceae and other aquatic types) was made (Maher, 1972). Pollen value was expressed in the percentage of total terrestrial pollen counted. Pollen identification was based on the modern pollen reference slides, photographs and theses from the University of Antananarivo in addition to published pollen atlases like Gosling et al. (2013) and Schüler and Hemp (2016). Samples for δ13C and δ15N isotope analysis were pre-treated with HCl and analysed at the Stable Light Isotope Laboratory at the Department of Archaeology, University of Cape Town. The purpose of the analysis was to identify the variation of C3 and C4 plants in sediment deposits (δ13C) (McCarroll and Loader, 2004). Stable carbon isotope and nitrogen composition (C/N ratio) were used to determine the origin of organic matter (Cloern et al., 2002) whether it came from aquatic plants with low C/N ratio (<12) or terrestrial plants with a high ratio, that is, >12 (Bonn and Rounds, 2010).
Charcoal analysis was used to reconstruct fire frequency in the area over time. Microcharcoal (<150 μm) is supposed to represent a regional signal, while macrocharcoal (>150 μm), being heavier, is supposed to reflect local fire (Carcaillet et al., 2001). Microcharcoal was identified and counted on pollen slides while macrocharcoal was counted under a stereomicroscope in a petri-dish. Charcoal is expressed in units of concentration that is, cm2.cm−3 for microcharcoal and particles.cm−3 for macrocharcoal. Charcoal accumulation rate was not used considering that influx is related to the sediment accumulation rate, which is often complex in the context of tropical ecosystems.
Dung fungal spore analysis was used to determine herbivory activities in the area over time. In this paper, they are represented by Sporormiella, Sordariaceae, Coniochaeta and Podospora taxa assumed to be coprophilous and used in previous studies of megafauna extinction and introduction of livestock in Madagascar (Baker et al., 2013; Burney et al., 2003). Dung spores were identified and counted on pollen slides and are expressed as concentration (spores.cm−3).
Numerical analyses
Cluster analysis based on the Euclidean distance was performed using the function ‘chclust’ in the R package ‘Analogue’ to identify the significant stratigraphic changes in the pollen assemblage of the Dangovavy sediment core (Birks and Gordon, 1985; Simpson, 2007).
Palynological richness, evenness-detrended palynological richness and β-diversity were used to evaluate biodiversity changes in Lake Dangovavy during the last 6300 years. Palynological richness is the number of pollen types per sample with a constant total pollen count and is estimated using rarefaction analysis (Birks and Line, 1992). It has been used by several studies to evaluate change in past plant diversity (e.g. Colombaroli et al., 2013; Giesecke et al., 2012; Meltsov et al., 2011). However, it can be influenced by pollen production and dispersion, and therefore evenness-detrended palynological richness was also considered in this paper. The index of evenness is independent of the number of taxa (Smith and Wilson, 1996) and considers equitably pollen types highlighting rare taxa (Giesecke et al., 2014). In addition, β-diversity was calculated to analyse changes in pollen assemblage composition in Lake Dangovavy and surrounds during the study period.
Ordination analysis was used to identify gradients in the vegetation composition during the mid-Holocene around Lake Dangovavy as already recognized and applied in past studies. First, the untransformed pollen data were analysed to extract the gradient length of the first axis of the Detrended Correspondence Analysis (DCA) (Birks and Gordon, 1985). Since the gradient was short from Lake Dangovavy (1.3 SD), a linear-based method, that is, principal component analysis (PCA) with square-root transformation of the pollen percentage data was used in this paper.
Open-source statistical software programme R v.4.3.0 (R Core Team, 2023) with the package ‘vegan’ (Oksanen et al., 2022) was used for all numerical analyses.
Results
Lithology and chronology
Based on a modified version of Troels-Smith (1955), the sediment core was composed of 10 different stratigraphic layers (SLs), named SL-I to SL-X from the bottom (110 cm) to the top (0 cm). The sediment mostly consists of brown organic clay mud with plant fragments, pieces of charcoal, coarse fine sand, and gravel in some layers (Figure 2). SL-I (110–93.5 cm) is brown and clayey with some plant fragments. SL-II (93.5–86 cm) is grey and clayey with some fine sands in addition to plant fragments. SL-III (86–82.5 cm) is still grey and clayey with an abundance of coarse sand and gravel. SL-IV (82.5–60.5 cm) is darker grey and clayey with rootlets, fine plant fragments and coarse sand. SL-V (60.5–47.5 cm) is similar to the first layer (SL-I), and SL-VI (47.5–38 cm) to the third layer (SL-III). SL-VII (38–29.5 cm) is calcareous grey with some plant fragments, while SL-VIII (29.5–18 cm) remains calcareous grey but less plant fragments. From 18 cm to the top, the sediment is darker clayey with the presence of plant fragments. SL-IX (18–4.5 cm) and SL-X (4.5–0 cm) differed by the presence and absence of fine sands, respectively.
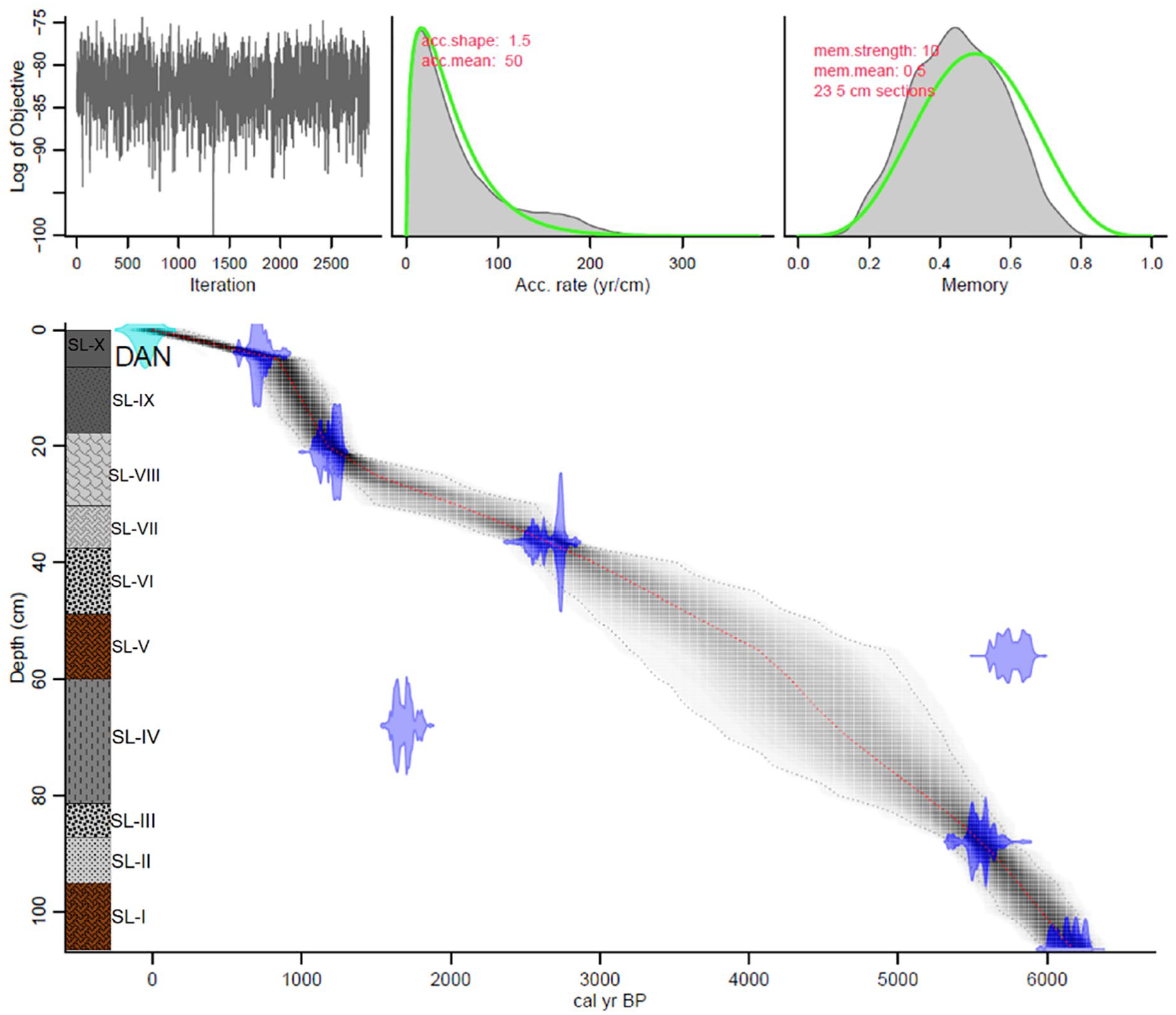
Lithology and age-depth model of the Lake Dangovavy sediment core, based on seven calibrated radiocarbon dates.
The bottom of the Dangovavy sediment core was dated at ca. 6.3 cal. ka BP. The age-depth model based on the seven (07) AMS radiocarbon dates suggests a mean sediment accumulation rate of 59.3 ± 7.3 years.cm−1 throughout the core but a large decrease of 163 years.cm−1 was noted from 4 cm to the top of the core. Two outlier dates, Beta–554609 (68 cm) and IT–C–1710 (56 cm), were excluded automatically by ‘rBacon’ as they represented reversals in the sequence. This may indicate contamination, sediment mixing (e.g. due to bioturbation or root penetration), reworking of sediment, or deposition inconsistencies, such as hiatuses.
Pollen, stable carbon isotopes, charcoal and dung fungal spores
Based on the cluster analysis, five (05) statistically significant pollen assemblage zones were identified, representing major vegetation shifts in Lake Dangovavy over the last ca. 6.3 cal. ka BP. These pollen zones will form the basis for the discussion of the pollen, stable carbon isotopes, charcoal and dung fungal spore results. Dates of the transitions between zones are tentative due to uncertainties in the age-depth model. Figure 3 represents the pollen diagram of the abundant terrestrial pollen taxa (greater than 2%) alongside the measure of δ13C and concentrations of charcoal and dung fungal spores during the Mid-Holocene.
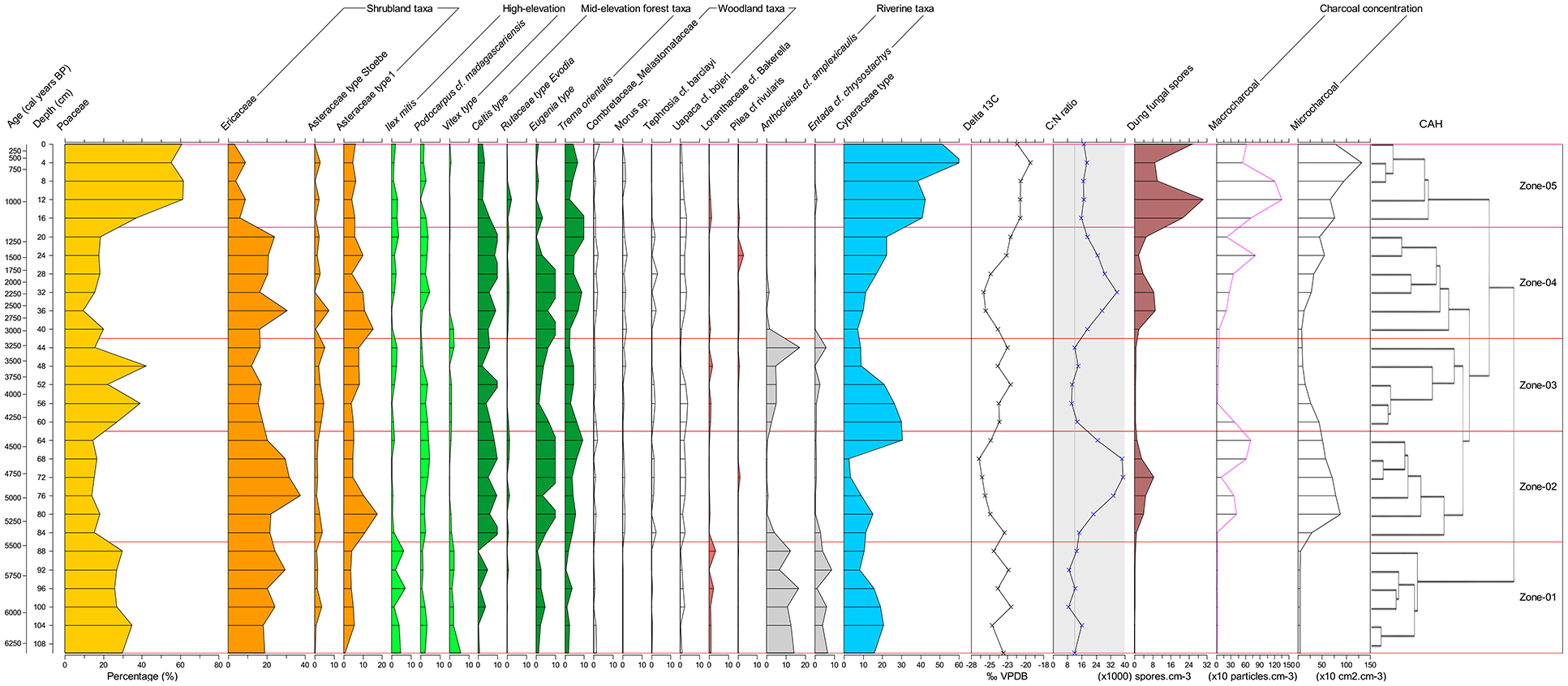
Pollen diagram of Lake Dangovavy grouped in their ecological affiliations (pollen taxa ⩾ 2%) characterized by five (05) statistically significant pollen assemblage zones: Grassland taxa (yellow), shrubland taxa (orange), open woodland taxa (white), riverine forest taxa (grey), high-elevation forest taxa (lawn green), mid-elevation forest taxa (dark green). The line graphs indicate the results of δ13C stable isotope (circle symbol) and C/N ratio (cross symbol) analyses, while dung fungal spores, macrocharcoal, and microcharcoal concentrations are displayed as brown, purple, and black silhouettes.
Pollen zone 1 (ca. 6.3–5.5 cal. ka BP)
The pollen assemblage was dominated by grassland and ericoid shrubland taxa characterized by the abundance of Poaceae (29.2 ± 1.4%) and Ericaceae (22.7 ± 1.8%) pollen, respectively. An abundance of riverine forest taxa was also recorded during this period represented by Anthocleista cf. amplexicaulis (12.7 ± 1.4%), and Entada cf. chrysostachys (5.1 ± 1.1%). Pollen from high-elevation forest taxa represented by Ilex mitis (4.2 ± 0.9%) and Vitex type (2.6 ± 0.6%) was slightly abundant during this period. Pollen from mid-elevation forests and open woodland taxa were scarce (ca. 2%). On the other hand, stable carbon isotope (δ13C) results showed a mean value of −23.7 ± 0.4‰ during the period, while the C/N ratio value was low (11.7 ± 1.1). The charcoal concentration was very low with a mean value of 15.3 ± 1.2 charcoal particles.cm−3 for macrocharcoal and 43.8 ± 1.6 cm2.cm−3 charcoal area for microcharcoal. Dung fungal spores were also very low during this period with a mean value of 0.2 ± 0.1 ×103 spores.cm−3.
Pollen zone 2 (ca. 5.5–4.3 cal. ka BP)
This second pollen zone was characterized by an increased abundance of ericoid shrubland, compositae and mid-elevation forest taxa. Ericaceae pollen showed a mean value of 27.2 ± 3% and Asteraceae a value of 9 ± 2.3%. Mid-elevation forest taxa were represented by Celtis type (8.5 ± 1%), Eugenia type (9.4 ± 1.8%) and Trema orientalis (5.6 ± 1%). A slight increase of open woodland taxa represented by Uapaca cf. bojeri (2.4 ± 0.3%) was also noted. However, Poaceae decreased to half of the previous period with a mean value of 15.7 ± 0.7%. Riverine and high-elevation forest taxa also decreased with an abundance of less than 2%. The δ13C isotope values were slightly lighter than in the previous period with a mean value of −25.3 ± 0.6‰ and this was accompanied by a high stable C/N ratio value (28.7 ± 4.4). Charcoal concentration increased more than tenfold during this period whereby macrocharcoal and microcharcoal increased to a mean value of 366 ± 122 charcoal particles.cm−3 and 623.4 ± 95.7 cm2.cm−3 charcoal area, respectively. An increase of dung fungal spores was also recorded with a mean value of 2 ± 0.6 ×103 spores.cm−3.
Pollen zone 3 (ca. 4.3–3 cal. ka BP)
The third pollen zone was characterized by dominance of grassland with peaks of Poaceae pollen at ca. 4 cal. ka BP (39.3%) and 3.5 cal. ka BP (42.2%). Ericoid shrubland and mid-elevation forest taxa decreased during this period compared to the previous zones. Ericaceae, Celtis type, Eugenia type and Trema orientalis pollen decreased to 16 ± 1%, 6 ± 1.5%, 4.3 ± 1% and 4.2 ± 0.7% respectively. Pollen from high-elevation forest taxa remained low. The relative abundance of pollen from riverine forests increased to 8.8 ± 3.6% with a peak of Anthocleista cf. amplexicaulis (17.7%) and Entada cf. chrysostachys (5.6%) at the end (ca. 3 cal. ka BP). The open woodland remained stable with a dominance of Uapaca cf. bojeri (2.3%). δ13C isotope results were slightly more positive with a mean value of −23.4 ± 0.3‰ while C/N ratio was low (12 ± 0.8). Charcoal concentration (macro and microcharcoal) was 5- to 10-fold higher than zone 1 but lower than zone 2 with a mean value of 100 ± 68.4 charcoal particles.cm−2 and 205.2 ± 64.4 cm2.cm−3 charcoal area for macrocharcoal and microcharcoal, respectively. Macro and microcharcoal concentration were higher at the beginning of the period and dropped at the end. Dung fungal spores dropped to a mean value of 0.5 ± 0.1 ×103 spores.cm−3 during this period.
Pollen zone 4 (ca. 3–1.1 cal. ka BP)
This pollen zone was marked by a return abundance of ericoid shrubland, Compositae, open woodland and mid-elevation forest taxa. Ericaceae and Asteraceae pollen showed a mean value of 21.5 ± 2.2% and 11.9 ± 2.7%, respectively. Both taxa peaked at ca. 2.6 cal. ka BP with a maximum value of 30.6% and 9.6% for Ericaceae and Asteraceae, respectively. Uapaca cf. bojeri was still dominant in the open woodland with a mean value of 2.5 ± 0.4%. Mid-elevation forest taxa remained characterized by Celtis type (8.5 ± 1%), Eugenia type (7.5 ± 2) and Trema orientalis (6.5 ± 1.2%). During this period, Poaceae pollen was low with a mean value of 16.7 ± 1.5%. High-elevation forest taxa remained low (<2%) and riverine forest taxa were almost absent (<0.5%). δ13C isotope results were more negative (−24.4 ± 0.6‰) while the C/N ratio increased with a mean value of 25.7 ± 2.6. Charcoal concentration was higher than zone 3 with a mean value of 311 ± 115 charcoal particles.cm−3 and 298.5 ± 74.8 cm2.cm−3 charcoal area for macrocharcoal and microcharcoal, respectively. Dung fungal spores increased during this period with a mean value of 5 ± 1.3 × 103 spores.cm−3.
Pollen zone 5 (ca. 1.1 cal. ka BP to present)
This last period was characterized by a large increase in the abundance of grassland taxa with Poaceae pollen reaching a mean value of 55.1 ± 4.6%. A peak of Poaceae pollen was recorded at ca. 900 cal. years BP with a value of 61.8%. On the other hand, other taxa decreased significantly in the pollen diagram. The ericoid shrubland and Asteraceae shrub decreased to a mean relative abundance of 6.2 ± 1.2% and 6.7 ± 1%, respectively. The Uapaca woodland, which was more stable in the previous periods, dropped to a mean relative abundance of 1.8 ± 0.5%. The high-elevation forest taxa and riverine forest taxa were very low to almost absent. Mid-elevation forest taxa were also low except Trema orientalis (5.6 ± 1.5%), which was more or less stable compared to the preceding period. The stable carbon isotope (δ13C) value during this last period tended to a more positive value (−20.9 ± 0.3‰). The ratio of C/N was moderately high compared to the previous period with a mean value of 16.9 ± 0.5. Charcoal concentration was very high both for macrocharcoal and microcharcoal with a mean value of 883 ± 165 charcoal particles.cm−3 and 896.3 ± 115.5 cm2.cm−3 charcoal area, respectively. Peaks were recorded at ca. 900 cal years BP with 1356 charcoal particles.cm−3 and 1321 cm2.cm−3 charcoal area for macrocharcoal and microcharcoal, respectively. Dung fungal spores were also very high during this period with a mean value of 19.3 ± 4.2 × 103 spores.cm−3 with a peak of 30.5 × 103 spores.cm−3 at ca. 900 cal. years BP.
Rarefied pollen richness, pollen evenness and compositional change
Rarefied pollen richness in the Dangovavy sediment core ranged between 24 pollen types (ca. 5.9 cal. ka BP) to 41 pollen types (ca. 1.1 cal. ka BP) with a mean value of 33 ± 2 pollen types in the entire period (Figure 4a). Between ca. 6.3–4.3 cal. ka BP (Pollen zone 1–2), pollen richness showed a mean value of 30 ± 2 pollen types. It increased (36 ± 1 pollen types) between ca. 4.3 and 1.1 cal. ka BP (Pollen zone 3–4). A peak of 41 pollen types was recorded consecutively at ca. 2.9 and 1.1 cal. ka BP. From ca. 1.1 cal. ka BP to the present period (Pollen zone 5), a slight decrease in pollen richness was recorded with a mean value of 34 ± 2 pollen types.
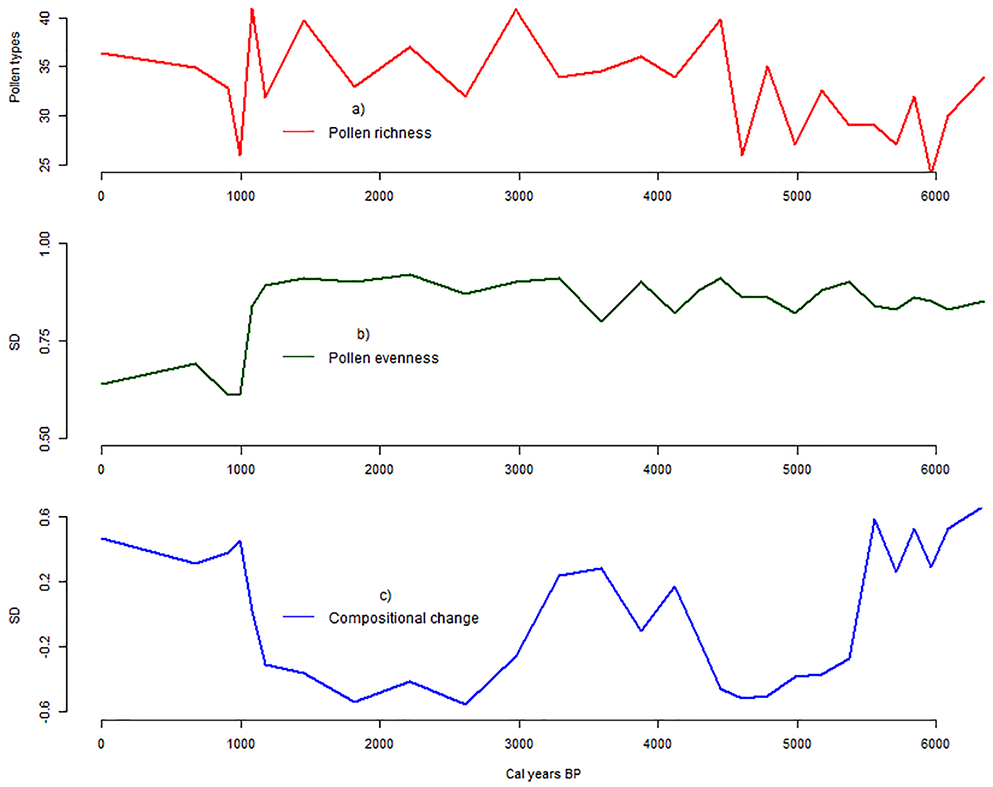
Biodiversity changes: (a) Rarified pollen richness, (b) evenness and (c) compositional change (Beta diversity) in Lake Dangovavy over the last ca. 6300 cal. years BP.
In parallel, between 6.3 and 1.1 cal. ka BP (Pollen zone 1–4), no long-term trends were recorded in the pollen evenness (Figure 4b). It remained fairly constant with a value of ca. 0.8 SD. However, based on the β-diversity calculation, the taxonomic composition differed at ca. 5.5 cal. ka BP (between Pollen zone 1 and 2); 4 cal. ka BP (within Pollen zone 3); 3 cal. ka BP (between Pollen zone 3 and 4) and at ca. 1.1 cal. ka BP (between Pollen zone 4 and 5, Figure 4c). From ca. 1.1 cal. ka BP to the present period (Pollen zone 5), a slight decrease in pollen evenness was recorded with a value of 0.6 SD. This variation was associated with the last change in taxonomic composition based on β-diversity calculations.
Ordination
PCA axis 1 and PCA axis 2 (noted as PCA1 and PCA2 hereafter) explained 57.8% of the variation in the distribution of pollen taxa in the Dangovavy sediment core (Figure 5). Poaceae, Anthocleista, Entada, Ericaceae, Celtis and Syzygium pollen taxa contributed most to the axis components. PCA1 explained 31.2% of the variance in the pollen data. It was related to dryness where riverine forest taxa (Anthocleista and Entada) and Poaceae had a high axis 1 score, whereas upland taxa represented by Celtis, Ericaceae, Trema, Syzygium and Weinmannia, had a low axis 1 score. In addition, PCA1 showed an elevation gradient where high-elevation forest taxa such as Faurea, Ilex and Vitex had high axis scores, while mid-elevation forest taxa such as Celtis, Syzygium and Trema orientalis had low axis scores. PCA2 explained 26.6% of the distribution of the pollen data. It was related to fire regime where high fire regime taxa such as the grassland taxa (Poaceae) had a high axis 2 score, whereas low fire regime taxa such as the ericoid shrubland (Ericaceae), forest (Celtis and Syzygium) and riverine forest taxa (Anthocleista, and Entada) had a negative axis score.
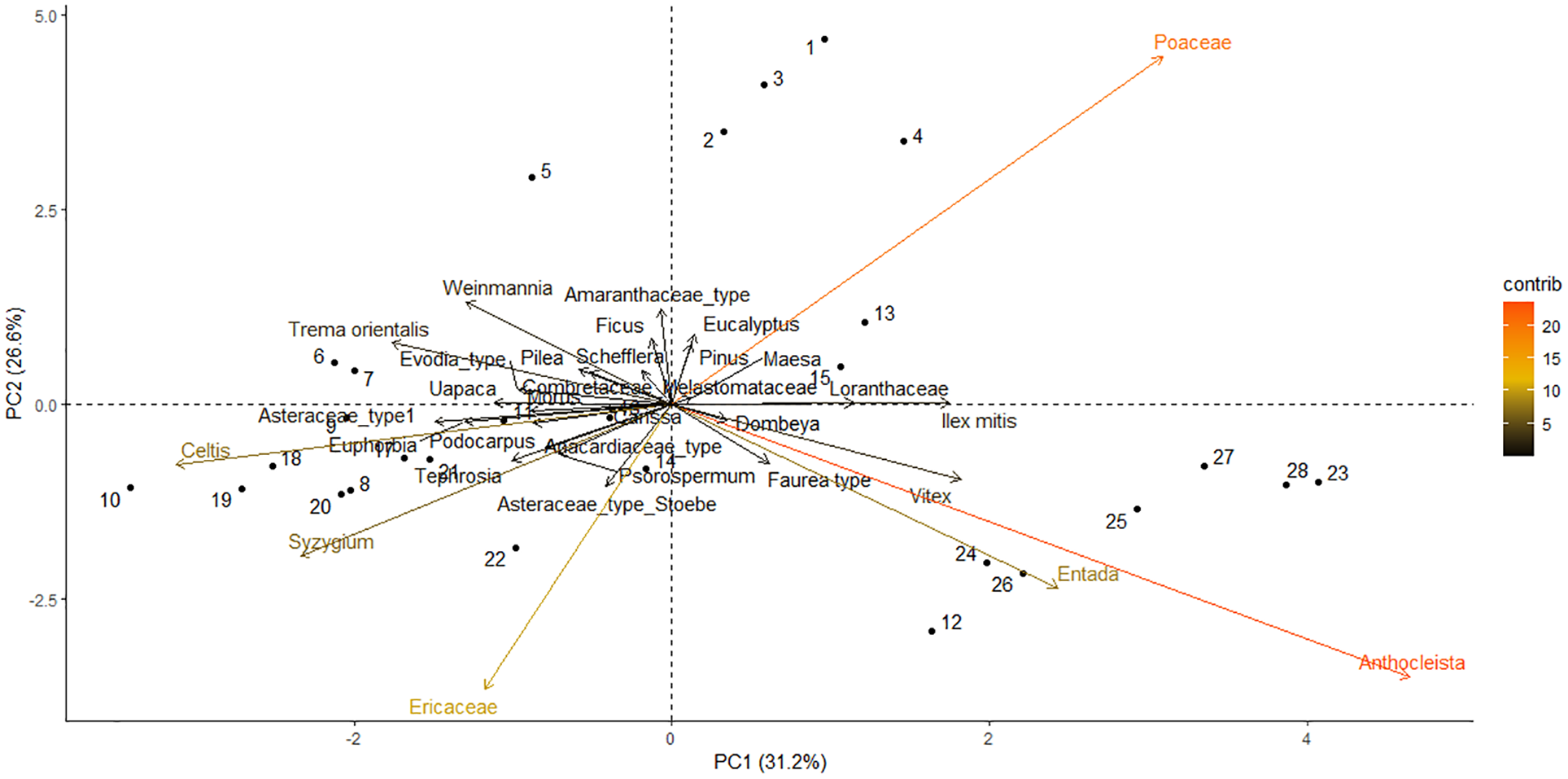
A Principal Component Analysis (PCA) determining the distribution of abundant and characteristic pollen taxa in a sediment core sampled from Lake Dangovavy. The colour gradient represents the contribution of each taxa to the axis components (lowest contribution = black; highest contribution = orange).
Discussion
The vegetation cover and its nature in the Central Highlands of Madagascar has been highly debated over the last two decades. Reconstructions have been reported on vegetation in the region; however, these records had some chronological issues or with low-resolution reconstruction (Burney, 1987c; Samonds et al., 2019; Straka, 1996). Here, we present the first high-resolution reconstruction of Holocene environmental changes in the ecosystems of the Central Highlands of Madagascar, based on a sediment core from Lake Dangovavy in the Vakinankaratra region. The sediment core from Lake Dangovavy is dated for about ca. 6.3 cal. ka BP based on radiocarbon dates of the bulk sediments. Despite some uncertainties, complexity in the age-depth model and the presence of hiatus (e.g. Broothaerts et al., 2023; Razanatsoa et al., 2021; Voarintsoa et al., 2017b) which is common in tropical and sub-tropical environments, the proxies used have allowed to identify the major changes in the vegetation and retrace the environmental history of the areas around Lake Dangovavy prior and after human settlement 2 cal. ka BP.
Environmental change in Lake Dangovavy, Central Highlands Madagascar, during the last ca. 6.3 cal. ka BP
Prior to human settlement (at least from 6.3 to 2 cal. ka BP)
Our palaeoecological record shows that open and mosaic ecosystems were already established around Lake Dangovavy and surrounds at least in the last 6.3 cal. ka BP, more than 4000 years before human settlement. The vegetation was dynamic and was characterized by mosaic ecosystems comprising forest patches of variable extent in a matrix of open grassland and ericoid shrubland, where grassland increased significantly with fire and herbivory activities in the past 1000 years (Figure 6).
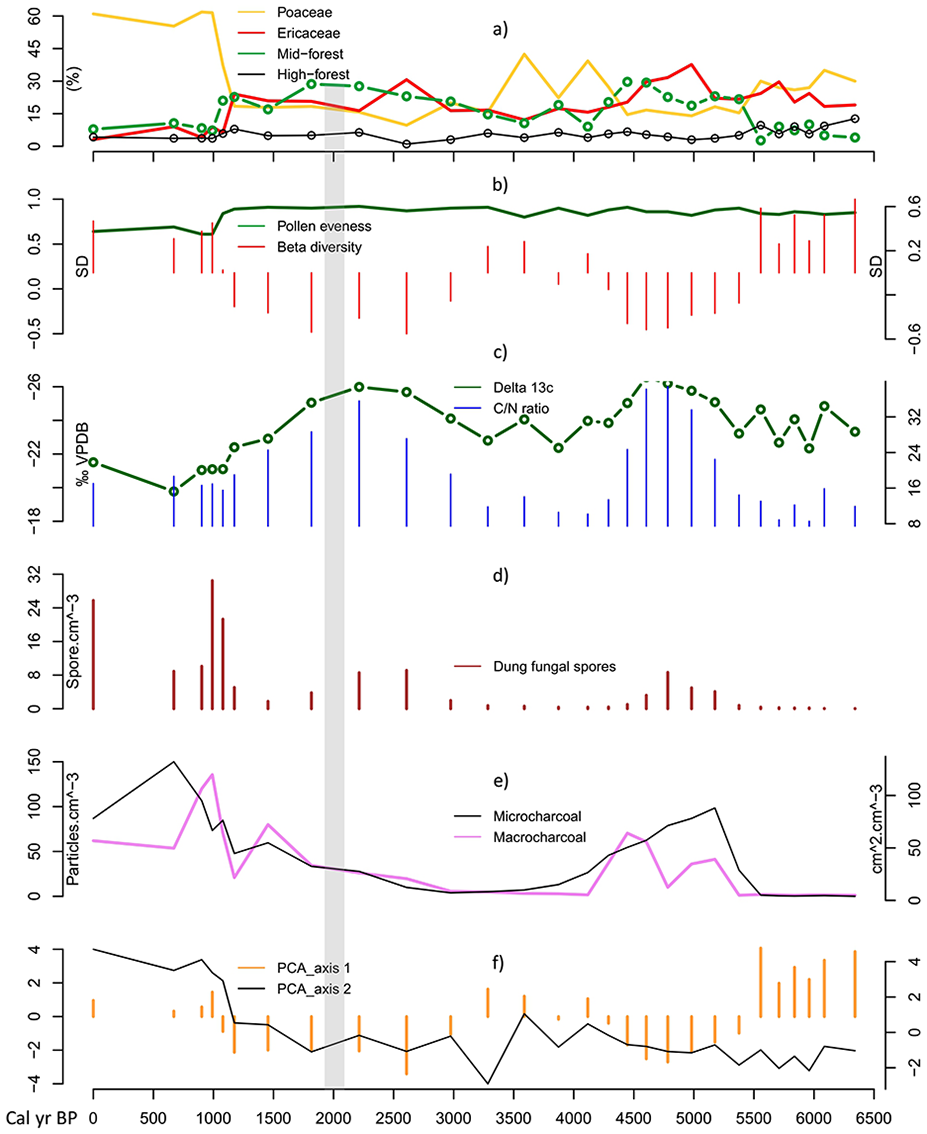
Summary of environmental change in Lake Dangovavy during the last ca. 6.3 cal. ka BP. (a) Relative abundance of grassland and ericoid shrubland (Poaceae, Ericaceae), and forest taxa (high- and mid-elevation forest), (b) Biodiversity changes, (c) Carbon and nitrogen isotopes, (d) Dung fungal spores concentration, (e) Charcoal concentrations, and (f) Principal Component Analysis (PCA) axes. Period of human settlement at ca. 2 cal. ka BP is indicated by a grey bar.
Between ca. 6.3 and 5.5 cal. ka BP, the pollen record suggests a mosaic ecosystem, where open grassy and ericoid vegetation co-existed alongside pollen from riverine and higher elevation forests (Figure 6a). The more negative value of δ13C isotope is consistent with the presence of C3 plants, which could be shrubs, woody plants (i.e. the forest elements mentioned above), or aquatics plants as justified by the low C/N ratio (Bonn and Rounds, 2010). The presence of C3 grass in the area was also possible but this needs further analysis on the measurements of grass-pollen δ13C to distinguish between C3 and C4 grass pollen (e.g. Nelson et al., 2008). Our new record shows the importance of the co-existence of grass and ericoid pollen, suggesting an open landscape with grassland and heathland elements as described in the Madagascar pollen rain studies (Burney, 1988; Razafimanantsoa and Razanatsoa, 2024). It seems likely that forest types were restricted to riverine and higher elevations, with grassland and heathland dominating the landscape. Charcoal (macro and microcharcoal) and dung fungal spores data (Figures 6d and 6e) indicate that fire regimes and herbivory activities were very low in the area during this earliest period. The restriction of forests might be associated with seasonally dry conditions in the area as suggested by the positive value of the PCA1 during this period (Figure 6f). Based on the literature, the Central Highlands of Madagascar experienced drier conditions in the Early- Mid-Holocene as justified by the sediment cores collected in Lake Tritrivakely that suggested an ephemeral swamp through mineral-magnetic properties, pollen, and diatom analysis (Gasse et al., 1994; Gasse and Van Campo, 2001; Williamson et al., 1998). This drier period was in parallel with the southern Madagascan and Southeast African climates showing an anti-phase climate relationship to northwestern Madagascar and Eastern Africa during the Mid-Holocene (Chevalier et al., 2017; Wang et al., 2019).
The period between ca. 5.5 and 4.3 cal. ka BP is marked by a compositional change in vegetation cover (Figure 6b). Ericoid shrubland and mid-elevation forest taxa expanded along with an increase in woodland taxa characterized by Uapaca cf. bojeri (see previous chapter). Riverine forests almost disappeared, while montane grassland was reduced to half its former value compared to the previous period. Expansion of forest taxa was associated with the slightly lighter value (−25.4‰) of δ13C which is justified from terrestrial sources based on the high C/N ratio during the period (Figure 6c). This change was associated with an increase in fire regimes and herbivory activities reflected by the charcoal (macro and microcharcoal) and dung fungal spore records (Figures 6d and 6e). Despite the regional increase of fire, forests and shrubs might persist in fire-free refugia and wetter areas, and maintain themselves through fire-vegetation feedback as commonly observed in savannas elsewhere (Huntley, 2023; Staver et al., 2011).
With an abundance of ericoid shrubland, woodland and mid-forest taxa, montane grasses decreased, which was intensified by increased grazing in the area during this period. Together, the abundance of shrubs and woody elements account for the more negative δ13C isotope value during this period. This expansion of shrubs and forests is associated with a negative value of the PCA1 (Figure 6f) and could be also associated with an increasing humidity in the region during this period. In fact, around 5 cal. ka BP the Central Highlands and Southwest of Madagascar supposedly shifted to more humid conditions (Burney, 1987c, 1993, 1996). This was at the end of African Humid period that is, the climate began to dry which might confirm the anti-phase climate relationship of Central Highlands Madagascar to East Africa previously mentioned. However, more palaeoclimatic studies are still needed in the region to confirm this theory. A more humid environment may have increased biomass production, explaining the increase in fire at the regional scale, as indicated by the microcharcoal values.
A second compositional change was recorded between 4.3 and 3 cal. ka BP (Figure 6b). Expansion of grassland in the mosaic ecosystem occurred at this time, with the highest value of Poaceae pollen at ca. 4 cal. ka BP. Forest and ericoid shrubland decreased compared to the previous period (Figure 6a). It was associated with a slight trend towards a more C4 signal from the stable carbon results. These might suggest the onset of C4 grass expansion in the highlands. Local and regional fire frequency were slightly higher at the beginning of the period which may have promoted an increase in C4 grass. Our findings suggest an abundance of C4 grass, which corresponded to a positive value of the PCA1 (Figure 6f), and was possibly associated with the 4.2 cal. ka BP dry event in the south east African monsoon and Madagascar (Scroxton et al., 2020), the warmest and driest period on the island (Gasse and Van Campo, 1998; Virah-Sawmy et al., 2010; Wang et al., 2019). A climatic and fire threshold might have been reached when the combination of fire with drought led to an ecosystem shift with a decline in heathland and a shift to a more open grass-dominated system. Herbivory activities were low during the period, which might be linked to reduced local animal density and functional species extinction in the landscape (Gill et al., 2009) possibly due to the drought event that occurred. No human influence was recorded in the region during this period.
A resurgence of ericoid shrubland, woodland and forest taxa was recorded in the area between 3 and 2 cal. ka BP (Figure 6a). This marked the third compositional change in the vegetation cover (see Figure 6b). Grassland decreased to half the abundance compared to the previous period. This was associated with a more negative value of δ13C isotope and a high value of C/N ratio (>12) confirming the dominance of C3 terrestrial plants (Figure 6c). Fire frequency and herbivory were low during this period, possibly contributing to the increase of shrubs and forests in the area at this time, as seen in the African savanna (Roques et al., 2001). This period is associated with a negative value of the PCA1 suggesting a more humid period (see Figure 6f). In the literature, this period corresponded to a more cool and humid period in the Central Highlands of Madagascar as suggested by the analyses of mineral-magnetic proxies (Williamson et al., 1998). These cool conditions could also have promoted the abundance of ericoid shrubland as observed during the glacial periods in the Central Highlands region (Burney, 1987c; Gasse and Van Campo, 1998, 2001).
Post human settlement (2 cal. ka BP to present period)
Between ca. 2 and 1.1 cal. ka BP, the vegetation around Lake Dangovavy remained dominated by a mosaic of ericoid shrubland, and mid-elevation forest taxa (Figure 6a). The negative value of δ13C isotope suggests an abundance of shrubs and trees within this period. Fire and herbivory activities started to increase slightly in the area compared to the previous period. However, vegetation in the area was resilient despite these changes, based on the pollen evenness analysis (Figure 6b). Woodland taxa such as Uapaca cf. bojeri, and Combretaceae were moderately abundant during the period (see previous Chapter), taxa which are suggested to be resistant to moderate fire in the area (Burney, 1987a). Therefore, fire was not frequent enough to burn savanna trees and this mosaic landscape might act to limit fire spread creating fire refugia for the ericoid shrubland. The cool and humid climate recorded in the region until ca. 1 cal. ka BP (Gasse and Van Campo, 1998) likely promoted the expansion of shrubs in the area as recorded in the region during the glacial period (Burney, 1987c; Gasse and Van Campo, 1998, 2001). Similar patterns of vegetation were recorded in the region as recorded surrounding Lake Tritrivakely during this period (Burney, 1987c). Despite the beginning of the increase in fire and herbivores in the area, suggesting human impact, it is likely that human population density was low during that period (Beaujard, 2011).
During the last ca. 1.1 cal. ka BP, more open grassland expanded into the area surrounding Lake Dangovavy, accompanied by a massive decrease in ericoid shrubland, and forest taxa (Figure 6a). This change was associated with a shift to more C4 plants according to the carbon isotope results, marking the last compositional change in the vegetation cover in the area (Figures 6b and 6c). This was accompanied by a decrease in pollen evenness suggesting vulnerability of the vegetation. This period is supposedly more drier than the previous period as indicated by the positive value of the PCA1 (Figure 6f). Based on the literature, a regional dry period was recorded at ca. 950 cal. ka BP as indicated by recorded pollen and diatom values (Gasse and Van Campo, 1998; Virah-Sawmy et al., 2010). Fire regime and herbivory activities were very high compared to the rest of the past ca. 6000 years (Figures 6d and 6e). This result suggests a local increase of human impact in the area during the last millennium through fire use and pastoralism, promoting the abundance of grassland. Our findings show similarity to other sites recorded in the highlands region with an expansion of grassland around 1 cal. ka BP (Burney, 1987a, 1987c; Straka, 1996). Similar trends have been recorded almost across the entire island, for instance in the Southwest (Razanatsoa et al., 2022), and in the Northwest suggesting climate and human activities (Railsback et al., 2020; Voarintsoa et al., 2017a) as drivers.
Conservation and management implications
Our data highlights the presence and importance of open and mosaic ecosystems characterized by montane grassland, and ericoid shrubland around Lake Dangovavy from ca. 6.3 cal. ka BP to the last millennium. This finding supports the hypothesis of the presence and abundance of natural open vegetation with forest patches along rivers and in fire refugia, rather than continuous forest in the Central Highlands of Madagascar (Bond et al., 2008, 2023; Solofondranohatra et al., 2018; Vorontsova et al., 2016). Though C4 grasslands expanded later in the record during a drier period and again when human impact increased, it is important to note that open and mosaic ecosystems dominated by a matrix of grasslands and ericoid shrubland are a natural part of the highlands landscape. Conserving these ecosystems is worthwhile, especially the ericoid shrubland, which deserves focused research priorities (Hackel et al., 2024). These habitats should not be assumed to be degraded and replanted as part of the government’s afforestation programme, which primarily focuses on the Central Highland’s open spaces (Lacroix et al., 2016). The decision to afforest the area needs careful re-evaluation, especially with regard to monoculture plantations of exotic trees such as Acacia spp., Eucalyptus spp. and Pinus spp. Exotic species would harm local biodiversity and increase biomass fuel, which would intensify fires and possibly put any remnant forest patches in jeopardy by slowing down native species recovery (Baohanta et al., 2012).
In addition to curbing the spread of alien plantations, however, it is also important to distinguish anthropogenic grasslands, which became more prevalent in Lake Dangovavy and surrounds during the last 1000 years in association with increasing human impact, from the ancient mosaics and heathlands that pre-dated this period. Hence, to encourage the restoration of ericoid shrubland in its natural habitat and to safeguard the remnant forest, careful fire management that will return burning to its historical range of variability is required in the area. As natural fire has occurred surrounding Lake Dangovavy and possibly the entire region for thousands of years before human arrival (Burney, 1987b), banning fire is not necessary. Instead, new measures to not exceed the threshold for Ericaceous vegetation and forest patches should be developed in order to save the remaining fire-adapted trees, including the Uapaca cf. bojeri. In fact, this species plays an important role in the region as it is exploited by local people for textile sources and food (Kull, 2002; Razafimanantsoa et al., 2013). Therefore, conservation projects in the area should focus on the restoration of such taxa as occurred in the former Ericaceous heathland but not the use of exotic trees currently promoted in the region (Lacroix et al., 2016). Meanwhile, given that locals rely on fire to support their way of life (Kull, 2000), fire management should concentrate on strategies that are sufficient to preserve the diversity of the environment and be done in conjunction with the local populace.
Conclusion
This paper provides new evidence of environmental change in the Central Highlands of Madagascar over the last 6 cal. ka BP from Lake Dangovavy, which highlights changes before and after human settlement in the region and informs conservation and restoration management. The palaeoecological records presented here offer a better understanding of the debated origin of the vegetation in the region and contributes to explaining the role of fire and herbivory activities in shaping the current landscape. Through this multi-proxy investigation, the palaeoecological data showed that the area has been dominated by open and mosaic ecosystems comprising forest patches of variable extent in a matrix of open grassland and ericoid shrubland at least 4000 years before human settlement. It highlighted the abundance of montane grassland and heathland in the area and suggests that the forest was restricted to riverine and higher elevations before 5.5 cal. ka BP. Expansion of heathland, woodland and forest, and then grassland to the mid-altitude with the onset of grass expansion, were recorded until 4.3 and 3 cal. ka BP, respectively in the area. Ericoid shrubland and forest taxa resurged to mid altitude between ca. 3 and 2 cal. ka BP, and even after human settlement more precisely at ca. 1.1 cal. ka BP. The vegetation remained resilient within these periods despite its dynamism and changes in regional climate. In parallel, fire was naturally present in the landscape at ca. 5 cal. ka BP where no human activities were recorded in the region. Variation of fire over time conditioned particularly the relative abundance of montane grass versus ericoid shrubland with grasses increasing as Ericaceae decreased and vice-versa. Modern examples of the changing dominance of ericoid shrublands versus C4 grasslands are common, for example at Ambohitantely Special Reserve. The fire events were possibly associated with climate-vegetation-fire feedback allowing for the maintenance of the mosaic landscape in the region. Herbivory activities fluctuated over the period peaking from ca. 5 and again at 2 cal. ka BP possibly maintaining openness of the mosaic, with a major surge of dung fungal spores from 1.1 cal. ka BP associated with the introduction of pastoralism and expansion of grassland. During the last millennium, vegetation in Lake Dangovavy and surrounds shifted to a more grassland-dominated landscape most likely through the influence of human activities as reflected by a high concentration of charcoal and dung fungal spores. Our results highlight the importance of understanding landscape history prior to establishing conservation and reforestation/restoration projects and caution against the current trends of afforestation with alien species. It further contributes to calls worldwide for greater recognition of the value and history of open and mosaic ecosystems to biodiversity and ecosystem services, including carbon storage and grazing resources.
Footnotes
Acknowledgements
The authors would like to thank the Mention Foresterie et Environnement de l’Ecole Supérieure des Sciences Agronomiques, Université d’Antananarivo Madagascar for facilitating field work permits in Madagascar. We acknowledge iThemba Laboratory at the University of Johannesburg for running free samples for radiocarbon dating and Stable Light Isotope Laboratory at the University of Cape Town who ran the samples for carbon content analysis. Special thanks go to Dr. Estelle Razanatsoa for her initial discussions and her assistance with proofreading the manuscript, as well as to Mrs. Samantha Venter for her thorough review.
Author contribution(s)
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the Applied Centre for Climate and Earth Systems Science (ACCESS NRF) [grant number UID 98018]; The Palaeontological Scientific Trust (PAST), Johannesburg, South Africa and the NRF/African Origins Platform [grant number 117666].