
Abstract
This paper presents the results of stable carbon (δ13C) and nitrogen (δ15N) isotopic analysis of bulk bone collagen from faunal remains from the Bronze Age site of Kaymakçı in western Anatolia. We use the isotopic values in conjunction with zooarchaeological data and contextual archaeological information to examine human-environment interactions; in particular, animal management and husbandry. Middle and Late Bronze Age agricultural systems in western and central Anatolia remain poorly understood, and this research aims to contribute to rectifying this research gap. We obtained stable isotopic values from both wild and domestic species, including deer, hare, birds, catfish, dogs, pigs, caprids and cattle. The δ13C values range from −22.0‰ to −16.9‰ and the δ15N values range from 2.3‰ to 10.3‰. For cattle, the δ13C values suggest that some specimens were subject to different management strategies, likely related to feeding or herding patterns or even the movement of animals in and around the region through some exchange mechanism. There appears to be no fixed location or strategy for the management of caprids and, furthermore, there is evidence to suggest that the herding/management areas of some caprids overlapped with those of wild animals such as deer and hare. Variation in δ15N values for pigs may indicate that whilst some animals in the sampled assemblage were free-roaming, others were penned. For the pigs and some other herbivorous domesticated species variation in δ15N values may also point to the exploitation of the wetland areas around Lake Marmara.
Keywords
Introduction
The communities of Bronze Age Anatolia witnessed the formation of larger and more densely organized settlements with more centralized governance structures. By the later Middle Bronze Age (MBA) and Late Bronze Age (LBA) of the second millennium BCE, these had either become woven into, or the people were at least certainly aware of, the first Anatolian empires, notably that of the Hittites (Bachhuber, 2012; Bryce, 2005, 2011; Glatz, 2009; Özdoğan, 2023; Roosevelt and Luke, 2017). The Aegean world, to the west, also saw the rise of the regionally powerful Minoans and Mycenaeans (Mountjoy, 1998). Region-specific leadership exerted control and influence over their immediate and larger territorial landscapes from large settlement centres. As part of these control structures, extensive agricultural systems that included field agriculture (i.e. predominantly cereal crops) and animal husbandry (including ante-mortem product exploitation) became more common, and these agricultural and staple finance economic systems were integrated into networks of exchange in goods such as metals and ceramics, and likely also textiles (Bachhuber, 2012; Diffey et al., 2020; Özdoğan, 2023; Roosevelt and Luke, 2017; Styring et al., 2017). The beginnings, subsequent developments, and intensified structure of deliberate and focussed animal management and crop cultivation is a defining part of human history.
The consequences of these overlapping strategies included radical changes in the dynamics between human societies and the natural world resulting in incalculable impacts not only on past human societies but also their environments (Goudie, 2019). In this context it is, therefore, important to understand how these Bronze Age settlements and their respective mechanisms of management leveraged local landscapes, agriculture, and agricultural systems to provide for increasingly large and diverse populations. Whilst there has been an abundance of research in the region, MBA and LBA agricultural systems (including animal management and husbandry) in western and central Anatolia remains poorly understood, with research so far privileging centres such as Troy and Hattusa (Marston et al., 2022 − for Hattusa see Diffey et al., 2020, Von Den Driesch and Pöllath, 2004 − for Troy see Riehl, 1999; Riehl and Marinova, 2008; Uerpmann, 2003). Excavations and environmental research at the Bronze Age site of Kaymakçı in western Anatolia are providing an important focus to address this deficit (see Irvine et al., 2024; Marston et al., 2022; Roosevelt et al., 2018; Shin et al., 2021; Slim et al., 2020;). This study aims to contribute further to rectifying this gap in regional understanding. Here we use stable carbon and nitrogen isotope analysis of faunal osteological material (on extracted bulk bone collagen) from stratigraphic excavations at Kaymakçı to investigate human-environment and human-biome interactions around the site, focussing on animal management and husbandry, in particular. We demonstrate that the data from Kaymakçı confirms the importance of micro-regional analyses in assessing the dynamics of Mediterranean river basins with diverse ecological niches, including wetlands, plains, and mountainous uplands. We show that the Kaymakçı community benefitted from and expanded on such proximate variation.
This study is just one part of the larger joint German Archaeological Institute-Koç University Research Center for Anatolian Civilizations (DAI-ANAMED) ‘Humidity and Society: 8,500 Years of Climate History in Western Anatolia’ project, part of and supported by the DAI ‘Groundcheck’ research cluster. It also provides some of the first applications of stable isotope analysis to faunal osteological remains to shed light on second millennium BCE animal management practices in the region. Whilst animal management practices have been one of the more commonly investigated themes utilizing isotopic analyses in Anatolia, faunal isotopic data from western Anatolia, particularly for the MBA and LBA periods is still relatively scarce (Irvine and Özdemir, 2020; Yazıcıoğlu and Irvine, In Press). Investigating animal management practices is a key facet in permitting us to explore how human and animal populations interacted with their environments as well as providing an understanding of the economic foundations of past communities. The study of archaeological fauna allows us to gain a better understanding of the living landscapes of the past; providing proxies for past environmental conditions, insights into ecology and behaviour, as well as a better understanding of the activities and decisions of past human groups that depended on those animals (Britton, 2018; Jones and Britton, 2019).
Thus, in this study, by combining biomolecular stable isotope analysis of faunal osteological material (bulk bone collagen) in conjunction with the evidence from more traditional zooarchaeological and contextual analyses, we aim to contribute to the understanding of the lives of the Bronze Age peoples of Kaymakçı and, in particular, their animal management practices. This, furthermore, will provide us with a way to investigate the dynamics of the past economy as well as human interactions with their ecological and environmental surroundings at the site and in the immediate surrounding environs.
Site and biogeographic information
Kaymakçı is located in the Marmara Lake Basin on the northern edge of the middle Gediz River Valley in western Anatolia (Figure 1). Under the direction of the Kaymakçı Archaeological Project (KAP), excavations at the site since 2014 have revealed a primary occupation period in the second millennium BCE, with several local MBA and LBA phases ending with thorough abandonment perhaps well before the end of the 13th century BCE (Roosevelt, 2024; Roosevelt et al., 2018). There are six contemporary citadels surrounding the lake basin, and Kaymakçı is the largest with a fortified area of 8.6 ha (Roosevelt et al., 2018; Roosevelt and Luke, 2017). Inside the main citadel fortifications walls demarcate an inner citadel and other areas, and the traces of terraces and other remains suggest that there was a dispersed extramural settlement at least down to the lakeshore and surrounding plains (Roosevelt and Luke, 2017). A large part of the inner citadel appears to have served as a centralized facility, whilst decentralized storage units appear associated with structures of mixed domestic and small-scale industrial utility spread across the site (Marston et al., 2022; Roosevelt et al., 2018; Shin et al., 2021). Due to its size, location, finds, and chronology, Kaymakçı must have been a regional capital in this area, which was known to the Hittites as the Seha River Land (Roosevelt and Luke, 2017). During the LBA, this indigenous kingdom was a vassal of the Hittite Empire at some points, whilst at other times it was aligned with the regional powers of the Aegean (Beckman, 1999; Bryce, 2003; Gander, 2017; Hawkins, 1998; Heinhold-Krahmer, 1977; Meriç, 2021).
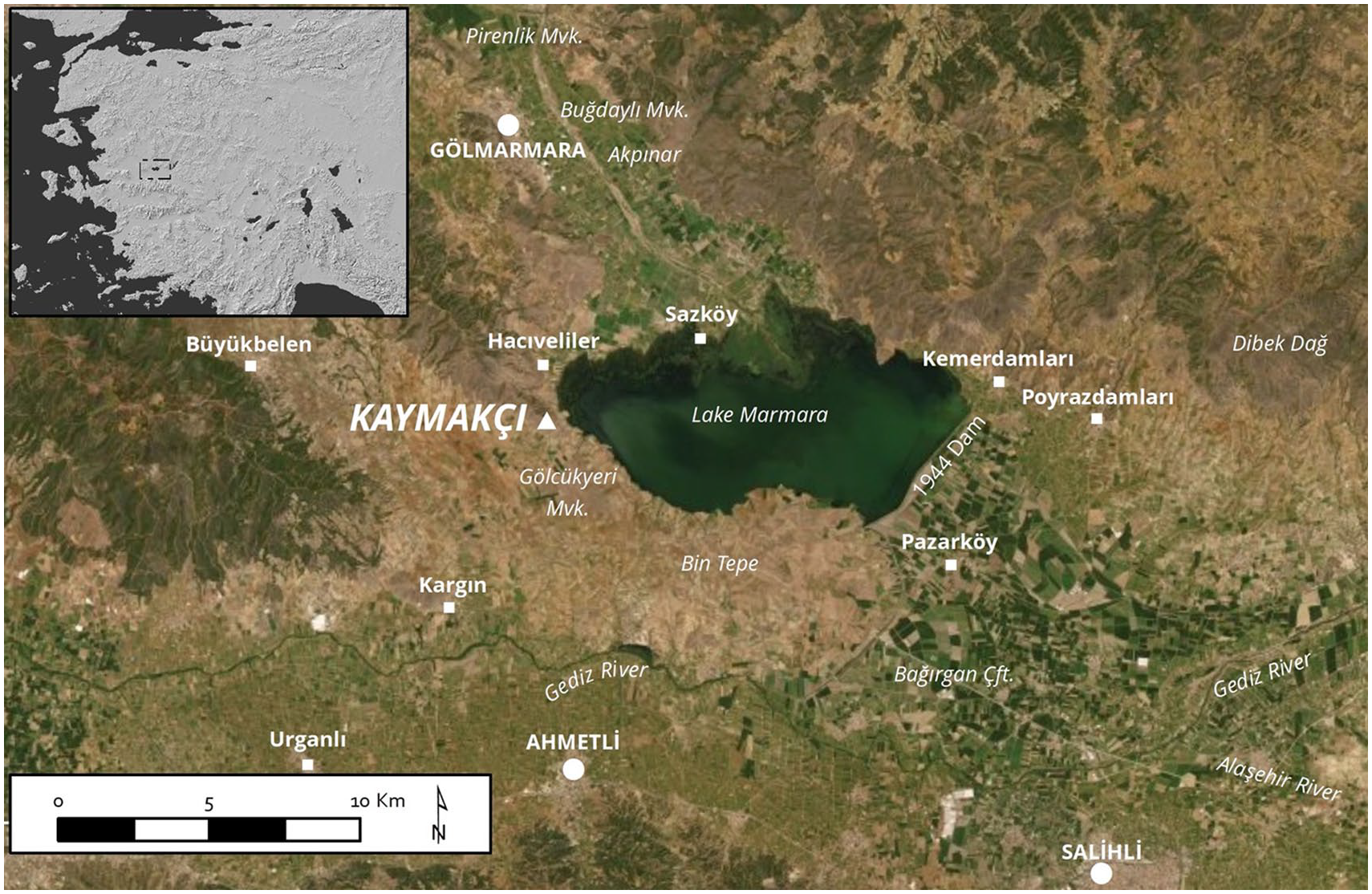
Regional map showing the location of Kaymakçı on the edge of the Marmara Lake Basin in the broader Gediz River valley. The inset shows the location of the map in western Türkiye (© Gygaia Projects).
Kaymakçı lies in the Mediterranean coastal woodland zone with a climate consisting of hot summers with humid and temperate winters, which makes it a particularly good environment for agriculture (Marston et al., 2022; Shin et al., 2021). The current landscape cover in the uplands around the site consists of open woodlands of deciduous and evergreen oak, steppe pastureland, and agricultural plots of annual crops, olives, and grapes (Marston et al., 2022). With increasing elevation, woodland density increases, as well, with pine forests dominating the mid- to upper elevations of the Bozdağ mountains along the southern fringe of the Gediz valley, for instance. Following anthracological analyses, oak species dominate the wood charcoal assemblage from Kaymakçı, and it is likely that the wooded landscape in the Bronze Age would have been similar to the current day (Marston et al., 2022). Local annual precipitation in the second half of the 20th century CE averaged 500–600 mm per year, with the summer months of June to September having the least precipitation (Marston et al., 2022; Roosevelt, 2009). Archaeobotanical research at Kaymakçı has revealed the presence of cereals, pulses, and fruits (Irvine et al., 2024; Shin et al., 2021). Cereals include barley, free-threshing wheat, einkorn, and emmer wheat, and pulses include bitter vetch, chickpea, grass pea, lentil and common vetch. Grape and fig are the only two fruits found at the site to date (Shin et al., 2021).
In the second millennium BCE, at least three major rivers, the Kumçay to the north and the Alaşehir that fed into the Gediz to the southeast, were the primary hydrological sources in the region, each fed by local mountain streams and permanent springs. In addition, historical documentation reveals that seasonal flooding created vast bodies of water in the plain, forming lake-like features for several months of the year (Çelik et al., 2024). Evidence indicates that at least one body of water, Lake Marmara, was stable throughout the year, even if waxing and waning in full extent and thus providing ideal conditions for a dynamic wetland zone. Kaymakçı sits overlooking the western edge of the lake basin. Diverse evidence confirms that the basin would have been particularly important (with regards to both agriculture and fishing) for the inhabitants of the site and other local contemporary communities; this pattern persisted through history until recent times (Luke, 2019; Luke et al., 2017; Luke, 2024; Marston et al., 2022; Roosevelt et al., 2018). In the Bronze Age this importance is demonstrated in one aspect by the evidence for fishing, in the form of carp and catfish bones and fishing equipment, including numerous bone gorges and at least one bronze fishhook (Roosevelt et al., 2018: 670, 678). There was likely an abundance of well-watered land for field agriculture with wetlands and other bodies of water in the area, some of which were spring fed, and others associated with seasonal flooding from the lake (Çelik et al., 2024; Gul et al., 2013; Irvine et al., 2024; Marston et al., 2022; Shin et al., 2021). Ethnohistorical research, too, confirms that prior to the 1950s transhumance flourished in the Gediz basin, with annual pastoral cycles between the mountain ranges (Luke et al., 2017).
Previous zooarchaeological research at Kaymakçı
Zooarchaeological research at Kaymakçı since 2014 documented more than 14,000 specimens from dry-sieved samples representing a diverse faunal spectrum of more than 40 distinct taxa from freshwater bivalves to brown bear (Marston et al., 2022; Roosevelt et al., 2018; Slim et al., 2020). Fish remains are numerous and visibly dominate the heavy fraction from floatation samples. This is not surprising given the site’s strategic proximity to Gediz wetlands. Wels catfish (Silurus glanis) and a variety of carp species have been identified ubiquitously in both dry- and wet-sieved units (Marston et al., 2022), with some individuals reaching lengths of 1 m or above. The presence of deer in the assemblage is significant, and most are fallow deer (Dama dama), as their body size and antler morphology indicate (Marston et al., 2022). Still, domesticated taxa dominate the dry-sieved samples. Numerically, pigs are the most abundant species identified in these samples (Slim et al., 2020), but cattle seem to have provided most of the meat obtained from farm animals (Fındıklar et al., 2024). The inhabitants of the settlement enjoyed these resources also thanks to the proximity of the wetlands. Cattle and pigs (and wild boar) require significantly more water than caprines to survive, so in pre-Roman sites in the eastern Mediterranean and southwest Asia, they tend to occur more frequently in lush environments than in arid regions (see, e.g. Çakırlar and Marston, 2019). Kill-off patterns reconstructed using bone density, fusion and tooth eruption and wear, and pig body size analysis suggested a variety of husbandry strategies involving multiple pig populations co-existing at Kaymakçı (Slim et al., 2020). The kill-off patterns of pigs suggest both intensive and extensive husbandry. The presence of perinatal piglets supports the idea that at least some (pregnant) pigs were kept within the site while the large number of younger pigs were possibly provisioned to the site. Along with adult males and females, and body size analyses, the pig assemblage of Kaymakçı presents to be phenotypically pluriform with diverse pig management strategies being employed at the site (Slim et al., 2020). The kill-off patterns of cattle, on the other hand, despite being an important source of beef, suggested that a restricted and concise provisioning model was applied for cattle husbandry. Juvenile and even young cattle are absent from the settlement deposits. While only adult cattle seem to have been brought to the site, beef from complete carcasses were (re-)distributed across the settlement, as indicated by the body part distributions (Fındıklar et al., 2024).
Isotope zooarchaeology: Theory and applications
Isotope zooarchaeology is now a reasonably well-established sub-field of bioarchaeological isotope studies and has been proven to be adept in clarifying complex aspects of human-environment interactions and human-animal economic, dietary, socio-political, and socio-cultural relationships (Britton, 2017; Jones and Britton, 2019; Makarewicz, 2016; Pilaar Birch, 2013; Zangrando et al., 2014). It has enhanced our ability to reconstruct the dynamics of past ecosystems, paleoenvironmental and palaeoclimatic conditions, and the economic and animal exploitation strategies employed by past humans (Jones and Britton, 2019; Makarewicz, 2016; Pilaar Birch, 2013; Zangrando et al., 2014). For shedding light on the multitude of approaches taken to animal management by past human groups isotopic analyses of domestic species has proven extremely useful, and while studies on wild animals are becoming more commonplace, the study of domestic species has continually been at the centre of bioarchaeological investigations (Jones and Britton, 2019). Isotopic analyses of teeth, micro- and macro-faunal bones (from both wild and domestic species), and shell have significantly contributed towards our ability to characterize the specific environments within the immediate environs and wider regions where archaeological sites are located (e.g. see Leichliter et al., 2017; Prendergast et al., 2017; Richards et al., 2017; Somerville et al., 2018; Worthey et al., 2022).
Faunal remains are noteworthy because their isotope values tend to reflect their ecology and the contemporary environmental locale, and their presence in the archaeological record is a direct result of human activities (Jones and Britton, 2019; Pilaar Birch, 2013; Zangrando et al., 2014). As a result, the isotope signatures of wild animals can provide evidence of animal ecology and environmental conditions (Britton, 2017; Makarewicz, 2016; Pilaar Birch, 2013; Zangrando et al., 2014). Isotopic analyses of the preserved remains of domestic animals are better fit to provide indications of anthropogenic modifications to their environments, diets, physiologies, and life-histories, including domestication, mobility, pastoralism and transhumance, foddering and feeding practices, and the compositions and sources of consumed water and vegetation (Makarewicz, 2016; Pilaar Birch, 2013; Zangrando et al., 2014).
The δ13C and δ15N values of bulk bone collagen mostly reflect dietary protein inputs and, due to collagen turnover in the bones, provide an average of an animal’s diet over the course of its life (Hedges et al., 2007).
The δ13C values of consumed plants at the base of the foodweb are reflected in the δ13C values of animal bone tissue, and thereby their δ13Ccollagen values (DeNiro and Epstein, 1978). Plants derive carbon from atmospheric CO2, which is fixed in plant tissues through photosynthesis, and thus δ13C values vary across different photosynthetic pathways (Bender, 1971; O'Leary, 1988). Most plants, including temperate shrubs, trees, grasses, and leguminous species utilize the C3 (Calvin-Benson) photosynthetic pathway, with δ13C values ranging from −35 to −20‰ (O'Leary, 1988). Plants which utilize the C4 (Hatch-Slack) photosynthetic pathway mostly include dry-adapted grasses that occur in environments with low moisture and warm temperatures and have δ13C values ranging from −15 to −7‰ (O'Leary, 1988). It should be noted, however, that for this study the exact δ13C values for local C3 and C4 plants is not known or defined. δ13C values can also vary across a single plant species due to several environmental factors, such as sunlight levels, water availability, temperature, salinity, altitude, and the canopy effect (Heaton, 1999; Middleton, 2018). Previous research has demonstrated that the δ13Ccollagen values of mammals are enriched by a trophic discrimination factor of ca. +5.0‰ relative to the δ13C values of their dietary inputs: Δ13Ccollagen-edible tissue (Ambrose and Krigbaum, 2003; Cheung and Szpak, 2021; Lee-Thorp et al., 1989; Sullivan and Krueger, 1981; Tykot, 2004). The δ13Ccollagen values of herbivores will, ultimately, depend mainly on factors relating to their diet and environment. Different locations and even seasonal inputs may be identifiable, and variations in δ13C between, or even within, species may be due to feeding habits: feeding in different locations, being foddered with different plant material, feeding under forest cover, or in wetter environments (Towers et al., 2011). Another significant factor is intense grazing which can alter the C3/C4 ratio of grasslands and may ultimately affect δ13C values (Derner et al., 2006).
Nitrogen isotope ratios, when applied to animals and humans, are generally used to interpret trophic level and marine resource consumption (Towers et al., 2011). The δ15N values of bone collagen (δ15Ncollagen) are primarily influenced by the δ15N values of dietary nitrogen, with environmental factors also having an indirect effect (DeNiro and Epstein, 1981). The δ15Ncollagen values can reflect environmental conditions through variations in the δ15N values of the soil and plants at the base of the foodweb (Somerville et al., 2018). One very general example is that higher δ15Ncollagen values can be exhibited in hot and dry environments relative to cooler and wetter ones due to a variety of factors, including soil and plant δ15N values being negatively correlated with mean annual precipitation and positively correlated with mean annual temperatures (Ambrose, 1991; Amundson et al., 2003; Cormie and Schwarcz, 1996; Craine et al., 2015; Handley et al., 1999; Hartman, 2011). δ15N values in soil and plants, and thereby animals, can also be variable at the local level due to factors such as local topography, including altitude, and spatial differences in local soils and plants, humidity, salinity, and local inputs of nitrogen (both natural and anthropogenic) including the deposition of organic matter (e.g. animal faeces and/or guano), sea spray effect, etc. (Bogaard et al., 2007; Hedges and Reynard, 2007; Middleton, 2018; Somerville et al., 2018; Towers et al., 2011). The δ15N values of bulk bone collagen are generally enriched by around 3‰ compared to that of the diet, but this can be variable due to several factors, such as species, age, trophic level, amounts of dietary protein input and physiological stress (Hedges and Reynard, 2007).
Materials and methods
For this stable isotope study, a total of 92 faunal bone fragments were sampled. Both wild (fish, birds, deer, hare) and domestic animals (caprids, cattle, pigs, dogs) were selected for isotopic analysis. The samples come from different areas of the site and different contexts (see Figures 2 and 3 and Supplemental Material 1), but all date to local LBA phases, dating roughly between the late 18th/early 17th and 15th/14th centuries BCE, as determined by a combination of relative ceramic phasing and radiocarbon analyses (Roosevelt, 2024).

Quickbird satellite image of the citadel of Kaymakçı, showing the locations of excavation areas from which faunal remains examined in this study derive (© Gygaia Projects).
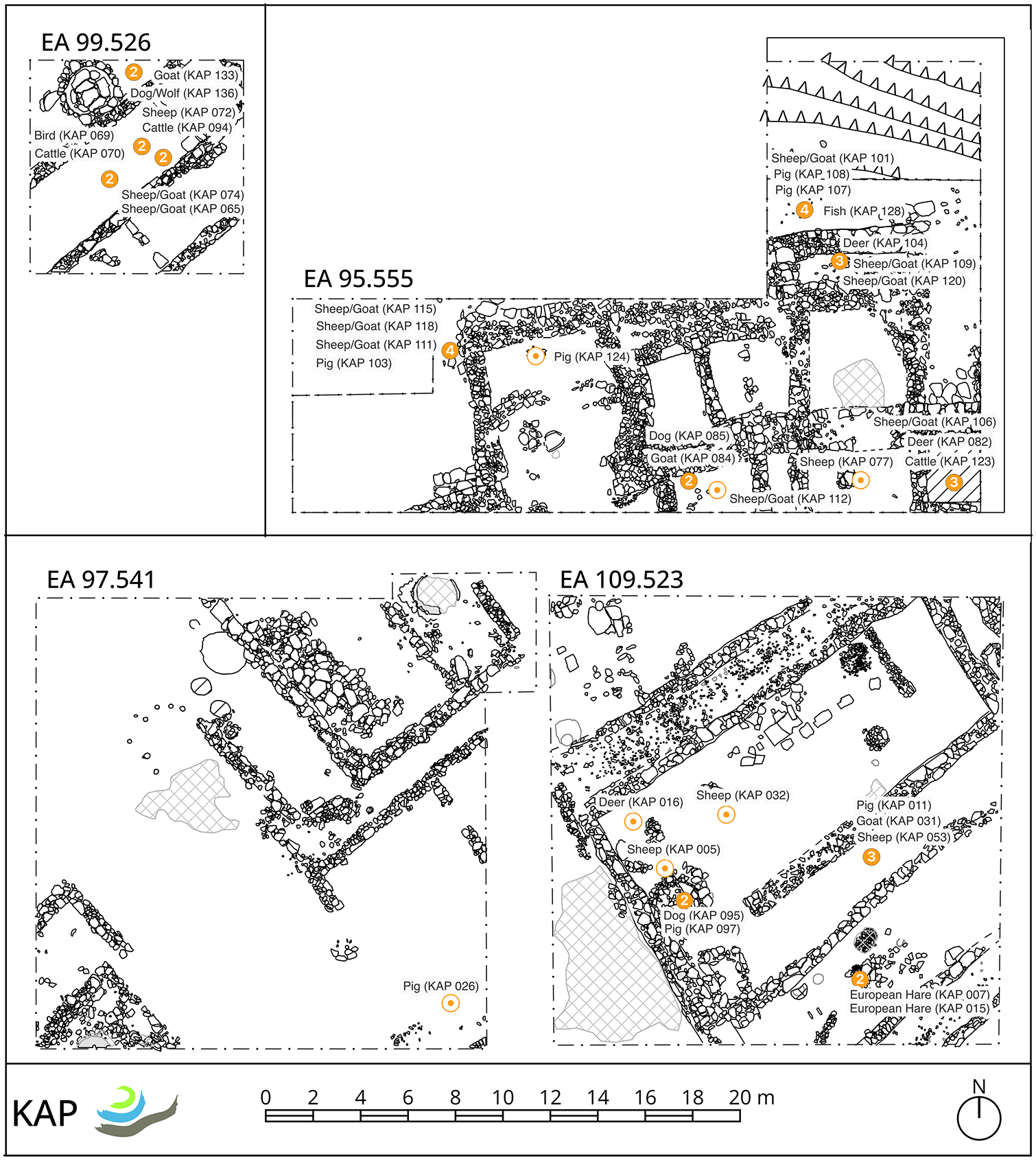
Plans of excavation areas 95.555, 97.541, 99.526, and 109.523 at Kaymakçı, showing the locations of faunal remains examined in this study (© Gygaia Projects).
For the taxonomic identification of the faunal material, an on-site reference collection was consulted, along with specialist publications including Schmid (1972), for mammals, Brinkhuizen (1989) and Radu (2005), for fish, and Cohen and Serjeantson (1996), for birds. To distinguish between sheep and goat, we followed Zeder and Lapham (2010), for postcranial bones and Zeder and Pilaar (2010), for teeth.
Prior to collagen extraction the external surfaces of the faunal bone samples were subjected to mechanical cleaning and in some instances, where necessary, they were cut into smaller pieces (ca. 3 cm in length) using a circular diamond saw. Collagen extraction of the faunal remains was performed at the ARHA (Archaeology and History of Art) laboratory of Koç University, İstanbul, Türkiye and followed a standard modified Longin (1971) protocol: demineralization of the bone samples in 0.5 M HCl (0.3 M HCl for more delicate bones such as those from the fish and birds), gelatinization in a pH 3 HCl solution at 70℃ for 48 h in a heating block, filtration using a 9 μm polyethylene Ezee-filter, followed by freezing and then lyophilization in a freeze dryer (Telstar LyoQuest Plus) for 48 h. Isotopic analysis was performed at TENMAK (Turkish Energy, Nuclear and Mineral Research Agency) on a ThermoFisher Scientific Flash EA 1112 coupled to a Delta V Plus mass spectrometer.
The reference standards used for IRMS analysis of samples at TENMAK were B2215 and IAEA-600 (δ13C), which have known values of −24.92‰ and −27.77‰, respectively. For δ15N, the same standards were used (B2215 and IAEA-600), in addition to IAEA-N-1, which has known δ15N values of 4.26, 1.0, and 0.4 ‰, respectively. All samples were run in duplicate and the standard values consistently fell within acceptable ranges for the known reference values. The analytic precision was 0.12‰ for δ13C and 0.27‰ for δ15N. The resulting δ13C and δ15N values were processed using Microsoft Excel.
Quality control of the extracted and analysed collagen was assessed following the criteria set out by Ambrose (1990), DeNiro (1985) and van Klinken (1999), who have determined that collagen with C:N atomic ratios lower than 2.9 or higher than 3.6, %C values outside of the range of 10%−47%, and %N values outside the range of 5%−17% is likely to have been altered, contaminated, and affected by diagenesis.
Results
From the initial 92 samples, only 38 passed the ancient bone collagen quality criteria defined above. All other samples (i.e. those that did not meet the quality criteria) were discarded from further analysis and are not discussed in this paper. Two of the samples have marginally low %N values but have acceptable %collagen and %C values, as well as acceptable C:N atomic ratios (Ambrose, 1990; DeNiro, 1985; van Klinken, 1999). Only when sample values fell outside of the acceptable ranges for all three quality checks (%C, %N and C:N) were they discarded from further analysis, as C:N values in ancient collagen still remain a strong gauge of collagen preservation (Guiry and Szpak, 2021).
The full set of δ13C and δ15N values and associated isotopic, zooarchaeological, and archaeological data can be found in the Supplemental Material 1, and the δ13C and δ15N values are plotted in Figure 4. The means and standard deviations, ranges, minimum, and maximum values for δ13C and δ15N values by species (for those whose n ⩾ 2) can be found in Table 1 and the means and standard deviations are visualized in Figure 5. Overall, the δ13C and δ15N values for the entire sample assemblage range from -22.0‰ to -16.9‰ and 2.3‰ to 10.3‰, respectively. All δ13C and δ15N values presented and discussed in this paper are from bulk bone collagen (i.e. δ13Ccollagen and δ15Ncollagen) and the values are given in parts per mil (‰) relative to the appropriate standards: V-PDB for δ13C and AIR for δ15N (Schoeninger and DeNiro, 1984).
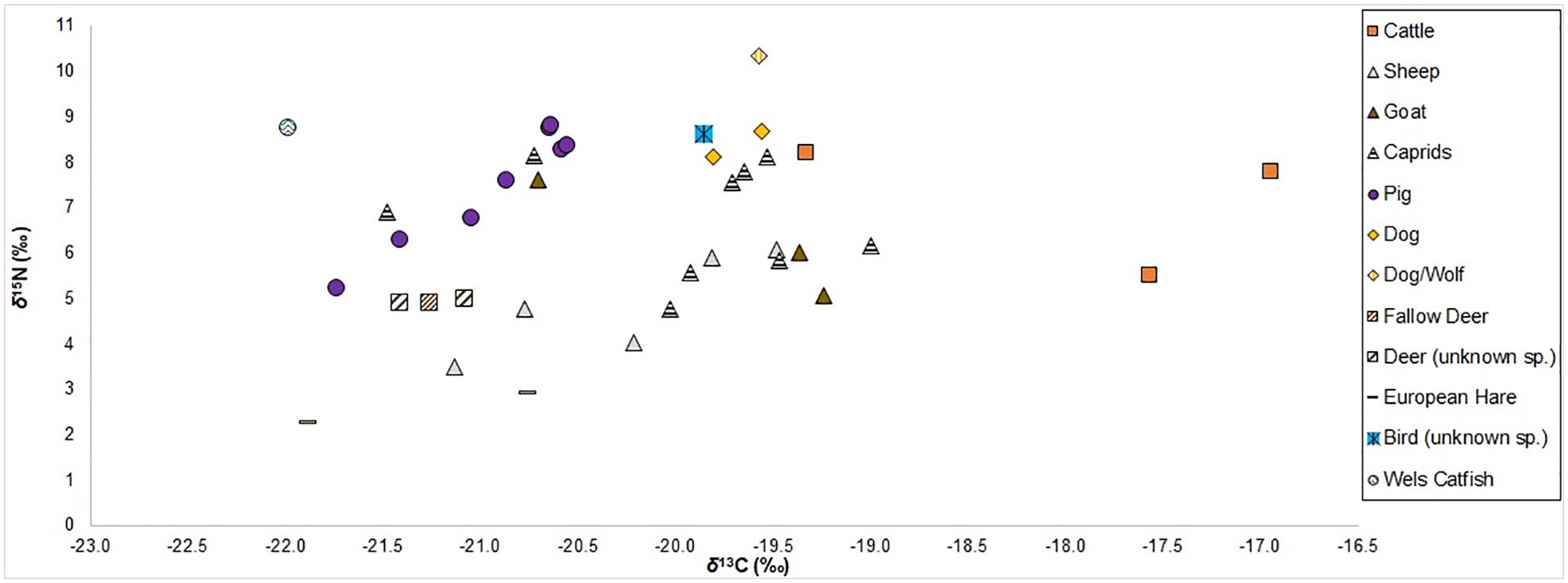
δ13C and δ15N values plotted for all faunal remains from Kaymakçı examined in this study.
δ13C and δ15N means and standard deviations (SD), ranges, and minimum and maximum values of species sampled from Kaymakçı examined in this study.

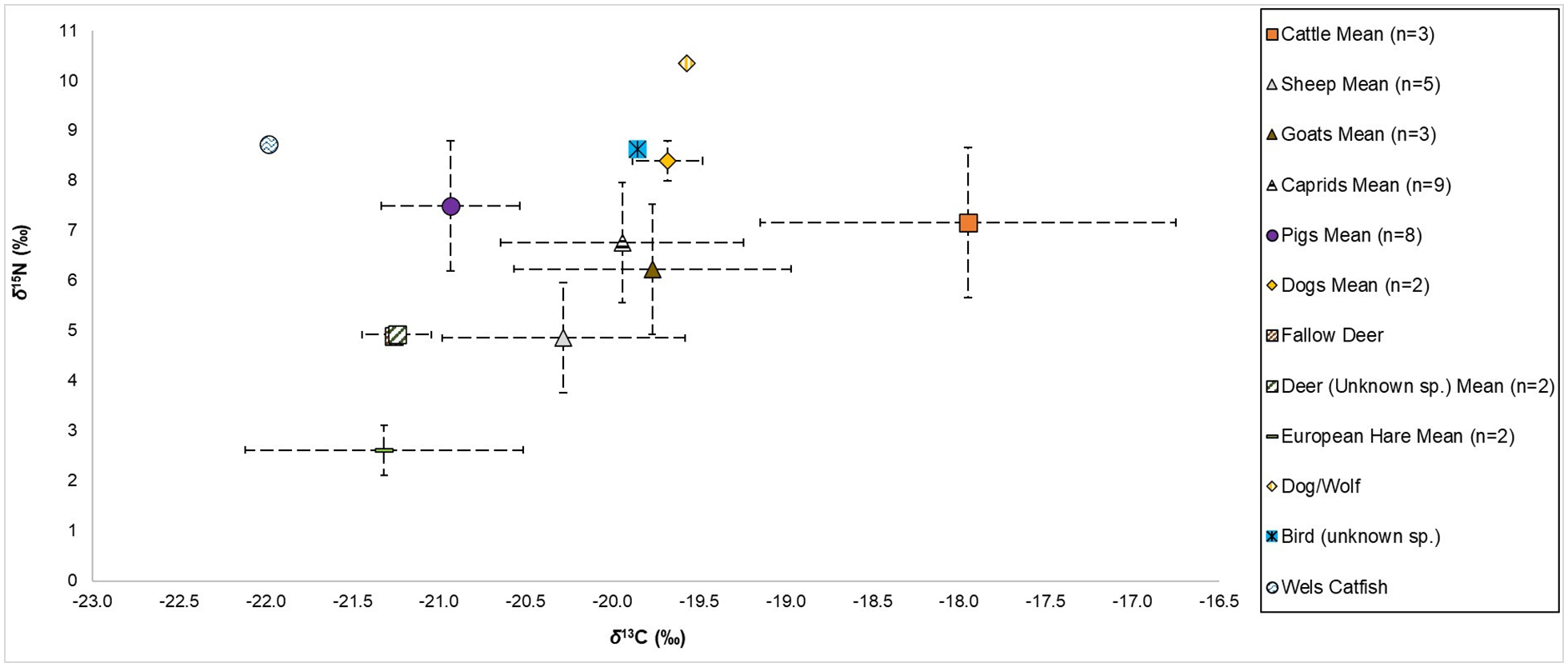
Means and standard deviations of δ13C and δ15N values, plotted for all faunal remains from Kaymakçı examined in this study.
Discussion
As can be seen from the stable isotope values (Supplemental Material 1, Table 1 and Figure 4), there is a reasonably large, and significant, spread in both δ13C (5.1‰) and δ15N (8.0‰) values across the species, and even within some species. This significance for intra-species range in values is particularly true for cattle (2.4‰ for δ13C and 2.7‰ for δ15N), and to a lesser extent for sheep (1.6‰ for δ13C and 2.6‰ for δ15N) and goats (1.5‰ for δ13C and 2.5‰ for δ15N). For pigs this is true for the δ15N values (3.6‰). There is a relatively large range for caprids too (2.5‰ for δ13C and 3.3‰ for δ15N), although, in theory, these ranges may be skewed by the fact that this caprid assemblage could include both sheep and goats which have different feeding patterns (grazers vs browsers). However, from the data in hand it does not appear that there were significantly different and distinct herding and/or management strategies according to species for sheep and goats (see further discussion below).
Conversely, there is a relatively tight clustering for the terrestrial herbivorous wild animals (deer and hare), with an overall range of 1.1‰ (−21.9‰ to −20.8‰) for δ13C and an overall range of 2.7‰ (2.3–5.0‰) for δ15N. This suggests that these wild herbivorous terrestrial animals were exploiting and feeding in very similar, or even the same, habitats and environments. It should be noted here that whilst one deer sample was definitively identified as fallow deer (Dama dama) the other two deer samples in this study could not be firmly identified to species level. However, these deer of unknown sp. (see Supplemental Material 1, Table 1, Figures 4 and 5) are very likely to also be fallow deer (due to this being the most ubiquitous deer species in the Kaymakçı faunal assemblage and the similarity of their isotope values to that of the firmly identified fallow deer specimen). There is a chance that these unidentified deer species may be red deer (Cervus elaphus), but they are definitely not roe deer (Capreolus capreolus). Fallow deer is native to the Anatolian peninsula, and is a species of deer that has been translocated and managed since the early Holocene (Baker et al., 2024). The frequency of fallow deer in archaeological bone assemblages in western Anatolia increases through time, which can be attributed to changes in the landscape (mainly deforestation due to human activity), the symbolic meaning of the species, and possibly direct management (i.e. controlled diet and mobility) (Çakırlar and Atici, 2017; Hubbard, 1995).
The patterning of the stable isotope values of the terrestrial herbivorous wild animals may also mean that we can tentatively, given the small sample size, identify a wild/natural unaffected range of terrestrial herbivorous faunal δ13C and δ15N values ranging from ca. −22.0‰ to −21.0‰ for δ13C and ca. 2.0–5.0‰ for δ15N. One implication of this is that domestic terrestrial herbivorous species with values outside these ranges are likely to be anthropogenically affected, that is, managed and herded (Middleton, 2018).
Animal management at LBA Kaymakçı
Specific animal management and herding strategies should be expected for domestic animals in Bronze Age Anatolia. At Kaymakçı, the zooarchaeological evidence has indicated a diversity in exploitation strategies, with cull patterns suggesting a variety of practices, ranging from primary exploitation for meat procurement as well as antemortem product exploitation across, between, and within different domesticated species; seasonality in (wild and domesticated) animal exploitation is possible, as well (Fındıklar et al., 2024; Marston et al., 2022; Slim et al., 2020). The stable isotope data obtained from this study allows us to investigate animal management strategies in more detail.
Relatively, and cautiously, speaking (as there are only three sampled cattle specimens), potentially the clearest evidence we have for animal management from the stable isotope values at Kaymakçı regards the cattle. Their δ13C values, in particular, are clearly outside of a hypothesized natural unaffected range (Figure 4) and are, in general, more positive than the values of other terrestrial herbivores, both wild and domestic (Figure 5). Furthermore, there is an indication that there were two distinct management strategies for cattle. Although, with only a few cattle individuals represented this remains a hypothesis that requires further investigation. Two individuals have more positive δ13C values that are >−18.0‰, which would suggest a mixed diet of C3 and C4 plants (Middleton, 2018). This could perhaps be indicative of herding on unmanaged/non-agricultural land or being foddered on collected vegetation that could have included wild C4 plants. One possibility is that they were herded further away from the site and could even be considered ‘non-local’ to Kaymakçı and its immediate environs. We cannot say this for certain, however, without further mobility analysis (e.g. δ18O and/or 87Sr/86Sr isotopic analyses). Even if correct, we cannot be sure whether this pattern results from long(er)-distance herding, pastoralism, or even the movement or exchange of cattle (i.e. cattle being brought to Kaymakçı from elsewhere). Ongoing and forthcoming research into management of cattle in Anatolia by some of the authors of this paper have noted a similar pattern in δ13C values of cattle from multiple regions and time periods in Anatolia.
There are a large number of cattle remains (NISP ca. 10%−15%) in the Kaymakçı faunal assemblage, and they were likely to have been regarded as an important species (Fındıklar, 2021; Fındıklar et al., 2024). It has been suggested, more generally, that cattle would have been prestige animals in Bronze Age Anatolia, principally related to their role as draught animals in agricultural extensification (Arbuckle, 2014). Furthermore, as only richer parts of society could likely maintain these animals, the animals themselves were a source of elite wealth having a direct financial value through their sale, rental, or exchange (Arbuckle, 2014; Kohler et al., 2018). The cattle in our dataset with the more negative δ13C value, which appears more ‘local’ (δ13C value closer to a hypothesized natural unaffected range) may be indicative of these types of cattle, being kept closer to the settlement, herded in the vicinity of Kaymakçı, and possibly being involved in agricultural activities. The use of cattle for traction at Kaymakçı would also make sense considering the likelihood of the Bronze Age model of extensive agriculture being conducted at the site (Fındıklar, 2021; Fındıklar et al., 2024). An antemortem product exploitation strategy of milk and/or traction at Kaymakçı has been suggested for cattle, most of which were culled in adulthood: 9 in 10 survived beyond 3 years of age (Marston et al., 2022). The δ15N value of the ‘local’ individual in the dataset may also point towards the penning of cattle and/or free-roaming herding in the wetlands down by the lake, as suggested for cattle at Kaymakçı by previous non-isotopic studies (Marston et al., 2022).
A ubiquitous presence of bitter vetch, which is often considered to be mainly a fodder crop, has been noted for the citadel areas of the site (Shin et al., 2021). One possible implication for Kaymakçı, therefore, is that bitter vetch was a highly valued and ubiquitous crop as it was either cultivated and/or strategically gathered from wetland boundaries for cattle grazing and fodder, and was thereby an important component in the overall economy and sociopolitical and socioeconomic structures (Irvine et al., 2024). However, it should be noted that for this sampled (cattle) faunal assemblage, whilst their δ13C values would agree with bitter vetch consumption (see Figure 6), their δ15N values do not (see Figure 4) as legumes tend to reduce the δ15N values of consumers (see Lösch et al., 2006).
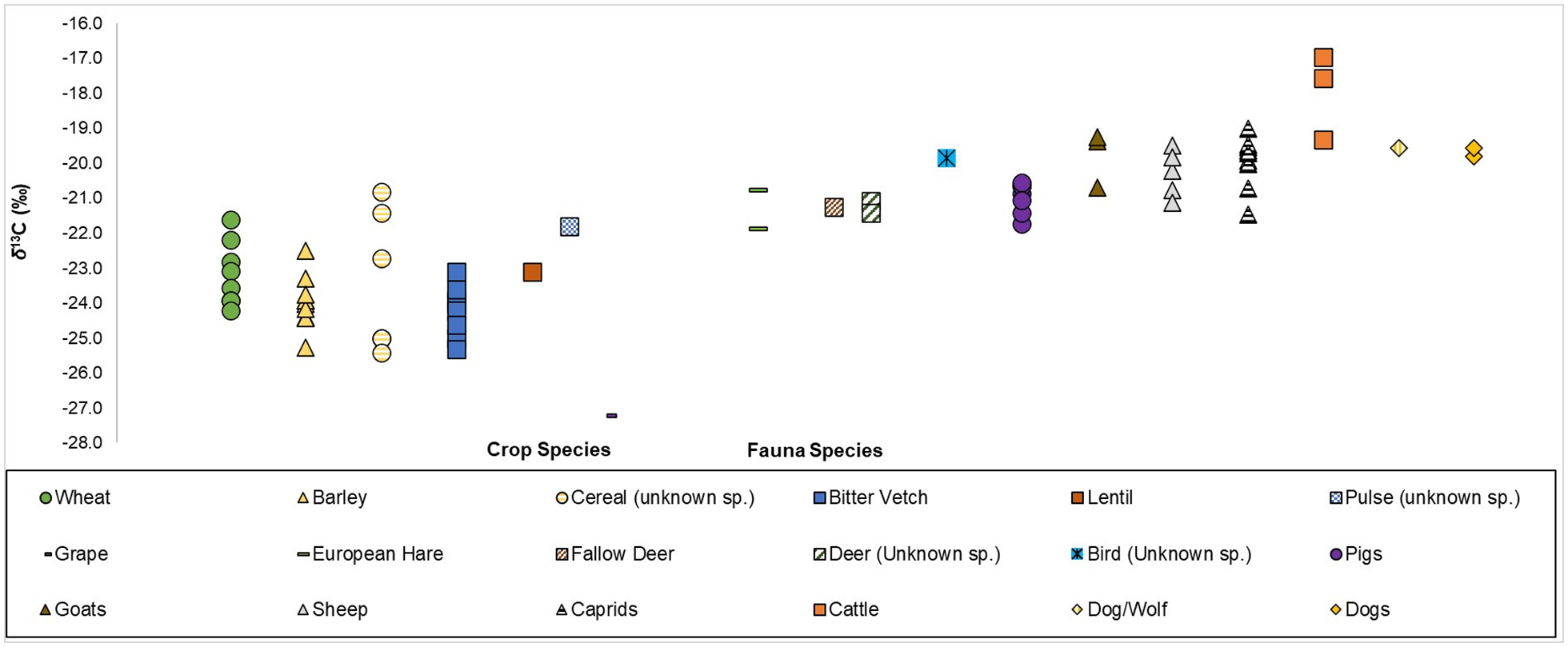
Linear plot of δ13C values plotted for all faunal remains, alongside those of contemporary crops, from Kaymakçı (crop δ13C values from Irvine et al., 2024).
As previously mentioned, the stable isotope values do not appear to suggest distinct management strategies for sheep and goats, and it seems possible that they would have been herded together. This is visible through the overlap in δ13C and δ15N values for sheep, goats, and caprids (Figure 4) – we do not see distinct patterning by species. However, there does not seem to have been a fixed herding location or strategy for sheep or goats, which is visible through the large ranges of δ13C and δ15N values. It seems likely that there was an exploitation of a variety of environments and locations, including the same habitats as those exploited by terrestrial herbivorous wild animals (sheep and goats with similar δ13C values, especially, to those of the deer and hare). Indeed, previous zooarchaeological and environmental investigations conducted at and around Kaymakçı have suggested that there was likely to have been herding and pasturing of sheep and goats amongst the open woodlands (Marston et al., 2022). The stable isotope data would also point towards herding of sheep and goats in other areas around the site (sheep and goats with more positive δ13C values), and likely exploitation of the more humid and saline wetland areas around the lake (sheep and goats with relatively higher δ15N values) (see Britton et al., 2008).
For the pigs, the δ13C and δ15N values would appear to suggest some variety in their management – which corroborates previous zooarchaeological examination of pig kill-off patterns. With some likely to have been free-roaming in open woodland (individuals with values close to the hypothesized natural unaffected range), pigs with slightly more positive δ13C values and δ15N values > 6/7‰ may be indicative of urban penning and/or the penning or free-roaming of pigs down by the lake. The stable isotope values for pigs supports earlier zooarchaeological and environmental investigations regarding pig management at and around Kaymakçı, which suggested that there was likely to have been open woodland free-roaming and urban penning of pig as well as the penning and/or free-roaming of pigs in the wetland areas near the lake (Marston et al., 2022; Slim et al., 2020).
The δ13C and δ15N values of the dogs (and as a result of the inferred trophic enrichment between diet and bone collagen) indicates that the main source of animal protein in their diet likely came from either wild animals (particularly deer), or sheep or pigs roaming freely in the same ecological zones as these wild animals. This would point towards an element of commensality, with the dogs either being fed directly by humans, or feeding on food scraps, perhaps including the discarded remains of these animals. This may, therefore, indicate a phenomenon of interest – this being that there were differences in the exploitation of animals depending on their management strategy (where they were herded), with wild animals and domestic animals free-roaming in the same/similar natural unaffected ecological zones being reared/exploited for primary consumption. Nevertheless, it is difficult to develop this idea further with any certainty due to the small number of dogs (n = 2) and the lack of human stable isotope values.
One final aspect to consider about animal management strategies at Kaymakçı is the relation between the δ13C values of contemporary Bronze Age crops (Irvine et al., 2024) and the δ13C values of the fauna examined in this study (Figure 6). As stated earlier, the δ13Ccollagen values of mammals are enriched by a trophic discrimination factor of ca. +5.0‰ relative to the δ13C values of their dietary inputs. This allows us to say that it is possible that wild (deer and European hare) and domestic animals grazed on crops/crop stubble and that domestic animals were possibly foddered on crops (Figure 6). However, it would seem more likely that if/when crops were used as fodder, the fodder would have been predominantly barley and/or bitter vetch, rather than wheat.
Environment and niche exploitation
The stable isotope data presented here has demonstrated that a variety of animal management strategies were employed at LBA Kaymakçı. However, this is not unexpected as there is a diverse range in local environments, meaning that there is the potential for versatility in animal husbandry management strategies at and around Kaymakçı. Animal husbandry strategies can be heavily influenced by the environments available to past human populations; thus, studies of animal management strategies such as this one can provide insights into the ways in which particular strategies may have been adapted and tailored to the specific conditions encountered (Jones and Britton, 2019). Multiple ecological zones are open to exploitation around Kaymakçı, including oak forests, lacustrine and riparian wetlands, and open steppe woodlands (Marston et al., 2022). The isotopic analysis of wild and domestic fauna in this study would appear to further emphasize that the Bronze Age people of Kaymakçı and the Marmara Lake Basin exploited all these available ecological zones. This versatility in ecological and environmental exploitation, with regards to the agricultural economy, has also been observed in crop management and arable agricultural systems at Kaymakçı, as it likely enabled risk-averse management strategies as well as the maximization of agricultural productivity and output (Irvine et al., 2024).
Previous studies into Leporidae have determined that they are generalist herbivores with mixed diets of grasses, forbs, shrubs and succulents (Somerville et al., 2018). Additionally, their δ15N values are most likely affected by soil salinity rather than moisture or temperature variables (Somerville et al., 2018; Ugan and Coltrain, 2011). This factor, and the low δ15N values of both the European hare and the deer sampled in this study, would seem to indicate that these animals fed not in the wetland areas around the lake but rather in the hilly open woodland areas around the lake basin. This means we can also likely place the free-roaming pigs and caprids in these areas too.
With regards to the other terrestrial wild species examined in this study (other than the deer and European hare), there is little we can say about local environmental and ecological conditions, as we only have δ13C and δ15N values for singular specimens. However, these animals (unknown species of bird and a possible wolf) were likely local and lived and ate in areas around the site. What this all indicates is that there was an overlap in the exploitation of ecological zones by both wild species and anthropogenically affected and managed domestic species, further emphasizing the exploitation of multiple ecological zones by the LBA inhabitants of Kaymakçı, as well as possibly suggesting that they encroached into ‘wild’ habitats to implement selected agricultural strategies.
Conclusion
In conclusion, we can say that the people of LBA Kaymakçı employed a variety of animal management strategies with regards to domestic species (cattle, caprids, pigs), in part due to the diverse range of environments and multiple ecological zones open to exploitation around the site. Furthermore, there does not appear to have been necessarily ‘fixed’ or homogeneous management strategies for specific species, with multiple ecological zones and strategies being exploited and employed at an intra-species level for cattle, caprids, and pigs. How representative Kaymakçı is of LBA animal management strategies in western Anatolia, and even Anatolia more generally, is currently unclear due to the relative lack of comparable data. However, one hypothetical pattern observed in the stable isotope values of this study that does seem to ring true across the larger region is distinct management strategies for cattle, with the presence of both ‘local’ cattle and other cattle that were managed differently, possibly herded and/or managed further away from the sites where they were recovered, including here at Kaymakçı. This is observed in the collated and examined δ13C values of cattle across time from the Neolithic to the Byzantine periods, across multiple regions of Anatolia, and is the topic of ongoing research.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836241297077 – Supplemental material for Stable isotope analysis of faunal remains from Bronze Age Kaymakçı, Western Anatolia
Supplemental material, sj-xlsx-1-hol-10.1177_09596836241297077 for Stable isotope analysis of faunal remains from Bronze Age Kaymakçı, Western Anatolia by Benjamin Irvine, Christina Luke, Canan Çakırlar, Tunç Kaner, Rana Özbal, Şengül Fındıklar and Christopher H Roosevelt in The Holocene
Footnotes
Acknowledgements
We would like to thank the Manisa Museum Directorate and the General Directorate of Cultural Heritage and Museums of the Ministry of Culture and Tourism, Republic of Türkiye, for permissions for excavations, sampling, and analyses. We are grateful to the directors and staff of the Turkish Energy, Nuclear and Mineral Research Agency (TENMAK), Ankara, Türkiye, for their helpful responsiveness and careful analyses. For assistance and guidance in field and laboratory work, we acknowledge all participants of the 2014–2019 seasons of the Kaymakçı Archaeological Project. For assistance with the lab work at Koç University we express our gratitude to Ebru Kaner, Sevil Kandemir, Şebnem Turhan, Berfin Dolançay, and Melis Yordamlı. Several researchers have significantly contributed to the Kaymakçı dataset. We especially acknowledge the laboratory efforts of Francesca Slim in identification and analysis of the broad zooarchaeological dataset to help contextualize her MA research on pigs from Kaymakçı. Ongoing analyses of more recent collections by Safoora Kamjan and Tuğçe Yalçın will feature in future publications. Finally, it is the excavators who have made this possible, both those who live nearby as well as the seasonal team. Here our heartfelt thanks to the communities of Büyükbelen, Hacıveliler, and Tekelioğlu, and the students of the Kaymakçı team. BI acknowledges a joint DAI-ANAMED Environmental Archaeology Fellowship held during the research and data collection of this study. This research was supported by Koç University, the Merops Foundation, private donors, and the ‘Groundcheck’ research cluster of the German Archaeological Institute (DAI) through a collaborative project of the German Archaeological Institute-Istanbul and Koç University Research Center for Anatolian Civilizations called ‘Humidity and Society: 8,500 Years of Climate History in Western Anatolia’. With respect to the latter project, the authors thank Felix Pirson for the pleasant and fruitful collaboration.
And finally, thank you to the two anonymous reviewers, whose comments and suggestions helped to improve the quality of this paper
Author contributions
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
ORCID iDs
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.