
Abstract
Tota is an Andean lake located in the Altiplano of the Eastern Andes of Colombia where socio-politically hierarchical societies of the Herrera and Muisca, flourished for millennia. To them, the lake and surrounding forest were places used for diverse of activities including religious rituals. In this study we produced a multi-proxy paleolimnological reconstruction using diatoms, isotopes, and geochemistry, to try to understand the lake’s pathways of change in response to natural climatic variations and anthropogenic activities. The diatom record is dominated by tychoplanktonic Staurosirella dubia and planktonic Aulacoseira species including species A. cf lirata, A. granulata, A. distans, and A. ambigua. Diatoms were grouped into functional groups and used to infer limnological changes that were further complemented with the geochemistry of the sediments to reconstruct the past environment. Results show three main periods in which the lake changed significantly, these are dated from ~800 to 1200, 1200 to 1500, and 1500 to 1900 CE. A correlation with the archeological record of the region, ethnohistoric accounts and climate suggests that these changes occurred simultaneously with changes in archeological stages, the Spanish arrival, and more recently by the industrial revolution, and the Little Ice Age.
Keywords
Introduction
The Altiplano of the Eastern Andean Cordillera of Colombia (northern South America), where Northern Andean societies flourished, is a high-land terrain of complex topography and remarkable environmental diversity with abundant fresh water lakes, rivers and wetlands (van der Hammen, 1974). In the Late-Holocene (around 2500 cal yr BP) agriculturalists settled and subsequently gave rise to the Herrera (about 400 BC–1000 CE) and Muisca (early 1000–1200 and late 1200–1600 CE) societies (Delgado, 2017, 2021; Delgado Burbano, 2012; Fajardo and Argüello, 2024; Rivas et al., 2024). Several landscape features such as hills, lakes and caves, were considered sacred by the Muiscas (Correa, 2004; Gamboa, 2010) and many lakes and wetlands were not only essential resources for subsistence, but also held significance in myths concerning their origins. These natural elements were pivotal in rituals that reinforced the religious and sociopolitical authority of the elite (Correa, 2004; Gamboa, 2010; Langebaek, 2019; Quintero-Guzmán, 2023). Offerings of wood figures, gold, and pottery are commonly found in archeological sites around these lakes reflecting their social and religious importance (Francis, 1993; Quintero-Guzmán, 2023). For instance, Guatavita Lake (about 100 km south of Tota) was particularly important. Fray Simon (1565] 1981[1565]), described cultural and religious rituals conducted at the lake, including offerings, cultural human sacrifice and burials . While Guatavita Lake was a major ceremonial sanctuary, Lake Tota was considered secondary (Quintero-Guzmán, 2023). However, the wetlands surrounding Lake Tota and nearby valleys hosted an independent and strong center which was part of the Sogamoso chiefdom. This chiefdom was one of the most influential political and religious entities in the Eastern Cordillera also of relatively high population density and early population clustering (Fajardo, 2016; Langebaek, 2019).
During the prehispanic and colonial periods, significant changes in life style, economic and social dynamics including increasing the clustering of the population, rising in pollution and extensive land use, had a direct impact on the indigenous populations (Fajardo, 2016; Larsen et al., 2001) and the surrounding landscape, including lakes (Fajardo, 2016; Gamboa, 2010; Langebaek, 2019).
Recent studies show that neotropical lakes are highly sensitive to direct and indirect anthropogenic stressors (Benito et al., 2022; Luethje et al., 2023; Vélez et al., 2021) and that they respond to stressors differently according to depth. Lakes shallower than 7 m change gradually to a novel diatom state whereas lakes, deeper than 14 m, fluctuate between states of similar diatom composition and then shift to a novel diatom assemblage. Activities such as the capture and deviation of water for irrigation, deforestation, and increase in population size and settlements, and subsequent organic waste, have impacted the lakes’ surface areas, water nutrients and transparency leading to changes in the distribution of native macrophytes and increased concentration of dissolved organic carbon (DOC). Additionally, the extra load of nutrients mainly from erosion, resulted in heightened primary productivity (Luethje et al., 2023; Salgado et al., 2019; Vélez et al., 2021). All these stressors have been gradually eroding the integrity of the aquatic ecosystems, leading to the emergence of novel lake conditions and ecological associations. Unfortunately, these transformations are taking place without a comprehensive understanding of the natural dynamics of the lakes.
Lake Tota, located in the mountains of Boyacá, Colombia, serves as a remarkable example of a ritualized lake by native societies that has undergone limnological changes since pre-colonial times (Cardozo et al., 2014; Correa-Metrio et al., 2022), and from which there is limited knowledge about its historical pathways and responses to natural and human influences. As the second largest and deepest lake in Colombia, spanning ~55 km2 with a maximum depth of 62 m (Gibson et al., 2019), it has been a landmark of significant cultural and environmental importance (Fonseca and Reyes, 2015). However, over the past decades, this lake has undergone notable degradation due to the excess influx of nutrients from agriculture and fish farming, decrease of its surface area to accommodate the ever-expanding agriculture, and from waste waters from a close municipality (Aquitania) (Secretaria de salud de Boyacá, 2012). However, our understanding of how the current limnological state of this valuable lake relates to baseline conditions and how it responded to natural dynamics amidst historical human pressures, dating back to pre-colonial times, remains limited.
In this study, we employ a multi-proxy paleolimnological approach (diatoms, isotopes, and geochemistry) to reconstruct the historical pathways of limnological change in the lake’s in response to natural climatic variations in contrast to historical anthropogenic activities. By examining these proxies from a core extracted from the southwest margin of the lake, and integrating archeological and ethnohistoric information, we aim to unravel the historical environmental changes and shed light on the interplay between natural and human factors in shaping Tota Lake’s ecosystem.
Study site
Lake Tota is located on the Eastern Cordillera of Colombia (5°28′ 13″ N–5° 39′ 14″ N and 72° 50′’ 38″W–73° 00′ 00″ W) between 2950 and 3900 m above sea level (m asl), and constitutes the source of water for over 200,000 inhabitants (Cañón and Valdes, 2011; Corpoboyacá UPTC, 2015; Nuñez et al., 2014). The lake has a surface area of 55.1 km2 that is divided into two sub-basins named the large lake (Lago Grande) and the small lake (Lago Chico) (Corpoboyacá UPTC, 2015) and maximum water depth of 65 m in the large lake (average 30 m) (Cañón and Valdes, 2011; Corpoboyacá UPTC, 2015; Nuñez et al., 2014; Figure 1). Average annual precipitation is of 810–1200 mm and varies from unimodal with higher precipitation (and lower evaporation) occurring between June and August (Cañón and Valdes, 2011; Nuñez et al., 2014) and maxima in precipitation in June–August, to bimodal in the south with maxima in March–May and September–November (Nuñez et al., 2014). Precipitation is controlled mainly by the annual migration of the Inter-Tropical Convergent Zone (ITCZ), and the Trade winds that enter Colombia from the northeast laden with moisture collected in the tropical Atlantic (Gimeno et al., 2020). A record of 50 years of lake level variations shows oscillations between maxima of 3015.70 and minima of 3013.69 m asl (Nuñez et al., 2014), the later associated with El Niño Southern Oscillation (ENSO) (Cañón and Valdes, 2011); water levels increase up to 40 cm during the rainy season but the overall trend is of a gradual decrease since 1987 likely due to civil hydrological modifications (Cañón and Valdes, 2011); it is worth noting that groundwater input is minimal (Cañón and Valdes, 2011).
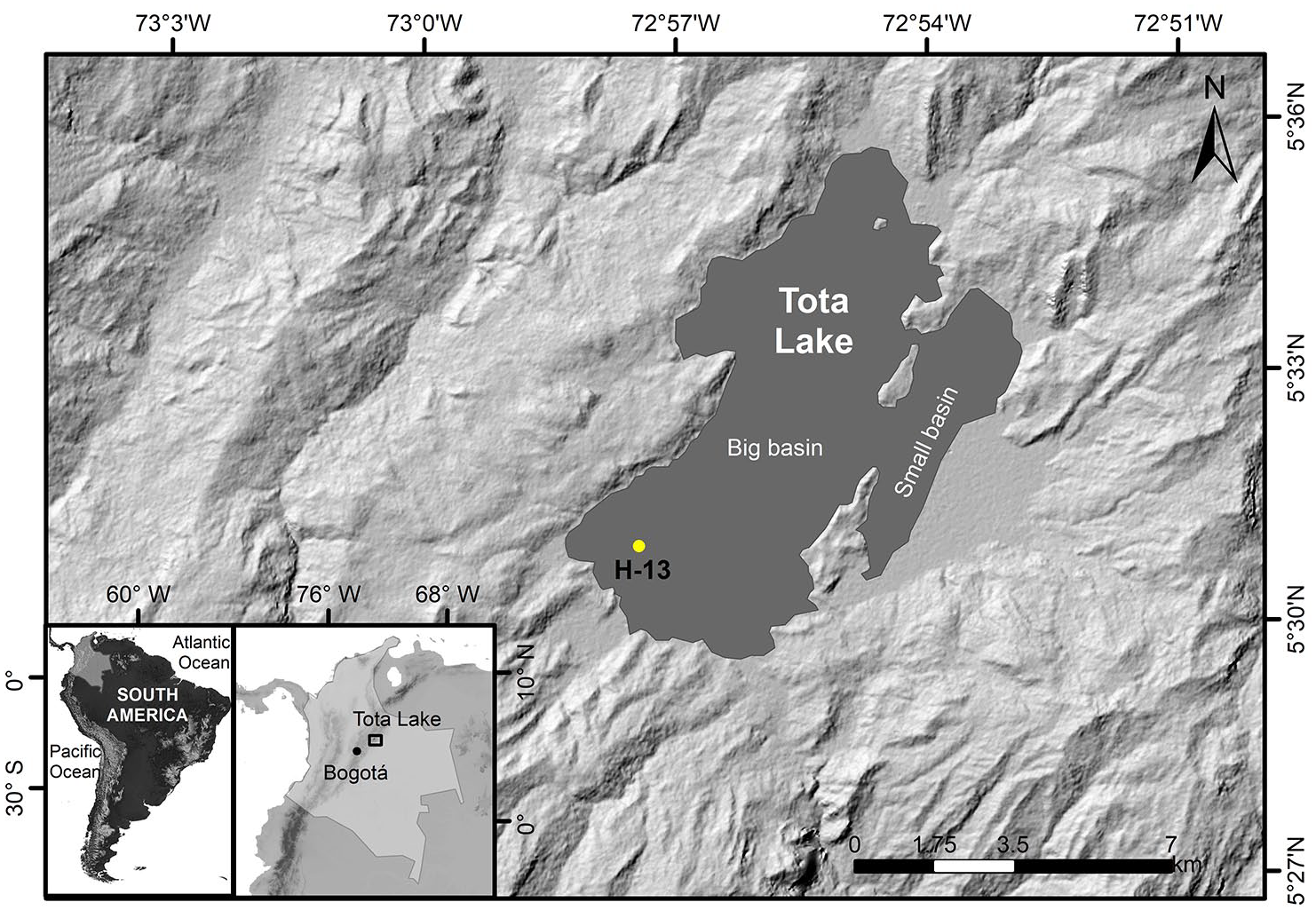
Lake Tota and its watershed, and location of the study (core H-13) site in the context of Colombia and South America (the bottom inserts).
The lake has a pH of 6.6, and prior to 2018 it had low conductivity and high transparency (Secchi depth of 9.7 m), hence that it has been classified as oligo-mesotrophic with point sources of phosphorous, nitrogen and carbon related to specific streams that run through urban or agricultural areas (Aranguren-Riaño et al., 2018; Gonzalez Angarit et al., 2008). Phosphorous is the limiting nutrient and its concentrations are slightly higher in the large lake (Corpoboyacá UPTC, 2015). Modern eutrophication is gradually increasing due to anthropogenic activities on the watershed being farming a constant source of N influx. Lake metabolism, this is, the fluxes of P, N and C are affected by seasonal variations in water levels (Corpoboyacá UPTC, 2015). For instance, TN increases when the lake level is high, likely from the extra N entering the lake via tributaries (Corpoboyacá UPTC, 2015). The negative correlation between TP and Chlorophyll-a and positive between Chlorophyll-a and TN, is suggesting that P, more than N is controlling primary productivity in the lake.
Lake Tota has a water storing capacity of about 1650 cubic millimeters (mm3) (Cañón and Valdes, 2011; Corpoboyacá UPTC, 2015; Nuñez et al., 2014). It is a polymictic (i.e. irregularly stratified throughout the year) warm lake, with a period of deep mixing induced by the Trade Winds during the rainy season (Cañón and Valdes, 2011; Corpoboyacá UPTC, 2015). According to Cardozo et al. (2005) Lake Tota contains a highly diverse algae flora that according to the authors is due to the lakes’ rather stable physico-chemical conditions through time.
A recent geophysical survey in Tota indicates that the lake’s sediment accumulated in the lake’s bottom reach up to 400 m of thickness and may be more than 1 million years old (Gibson et al., 2019). Paleoenvironmental and paleolimnological reconstructions show that its water levels are sensitive to millennial changes in climate, mainly from anomalies in precipitation and evaporation (P/E) that in turn affects lake’s ecology by controlling macrophyte extension and water and nutrients (Cardozo et al., 2014; Correa-Metrio et al., 2022).
Methods
Diatoms were extracted from a core (H-13) collected from the southwest part of the lake by a team from the Paleoclimatology and Sedimentology Laboratory from Indiana University, Indianapolis (IUPUI) (Figure 1; Gibson et al., 2019). This core is 40 cm long and was dated and analyzed for pollen, total organic carbon and total nitrogen, grain size, and geochemistry (Bird et al., 2024; Correa-Metrio et al., 2022). Diatoms were sampled and analyzed every cm between 10 and 39 cm; the upper 10 cm were not recovered in the extraction. Diatoms were grouped into planktonic (sum of all Aulacoseira species and single-celled Cyclotella, Lindavia and Discostella), tychoplanktonic (all species of Fragilaria sensu lato and former Fragilaria spp), and benthic epiphytic and motile species. This grouping allows to explore limnological change related to mixing, DOC, littoral dynamics and macrophyte extension, and stress. Aulacoseira species are robust diatoms that form long chains and require water turbulence to be suspended in the water column, whereas single celled planktonic species, for their relative high surface area to volume ratio, do well under reduced water mixing (Winder et al., 2009) induced by warming of the epilimnion and or changes induced by an increase DOC (Brown et al., 2017). Tychoplanktonics are commonly associated with littoral conditions, particularly in the Colombian Andes (Catalan and Donato Rondón, 2016; Vélez et al., 2021), whereas motile species have the capacity to move and proliferate when conditions deteriorate (stress) for other diatoms that cannot move away from disturbance (Berthon et al., 2011; Passy, 2007).
Ten samples (12, 14, 18, 20, 24, 26, 30, 32, 36, and 38 cm depth) were analyzed for trace element concentration at the University of Regina, Canada using a Rigaku NEX QC + EZ XRF Spectrometer. The sediment was manually disaggregated in a mortar and analyzed in a 32 ml diameter cup, where the sediment was flattened using a manual press; cups were sealed with polypropylene. Samples were run with three filters: low, medium and high, for 2 min each time. Final measurements are presented in weight percentage for main elements and ppm for trace elements. These measurements were used as a proxy for relative concentration of a given element between samples. The following element rations were used as a guide to infer changes in the lake and its watershed (in parenthesis) according to Davies et al. (2015): Ti/Ca (detrital input), Mn/Fe (water column oxygenation), Zr/Fe (flooding), and sedimentary P was used as an indication of excess or consumption of P on the water column by aquatic plants (higher concentration of sedimentary P would indicate more P on the water column, lower concentration would indicate that P is consumed in the water column).
Core dating and geochemical stratigraphy for this core (H-13) has been already published in Correa-Metrio et al. (2022). The Age model was produced with a combination of two AMS 14C on charcoal samples (Table 1) and of 210Pb and 137Cs measured in the upper 13.5 cm (Correa-Metrio et al., 2022) modeled using Bchron (Haslett and Parnell, 2008). Radiocarbon ages were calibrated with the IntCal20 curve (Reimer et al., 2020) and Pb and Cs ages were estimated using a Constant Rate of Supply (Appleby and Oldfield, 1978). Geochemical analyses were performed on bulk sediment include total organic carbon (TOC), total nitrogen and carbon (TN and TC, respectively), and 15N and 13C. Values for depths 23, 25, 28 cm for TC, TOC, TN were not measured so an average between values from the immediate upper and lower depths was used.
Results from radiocarbon dating.
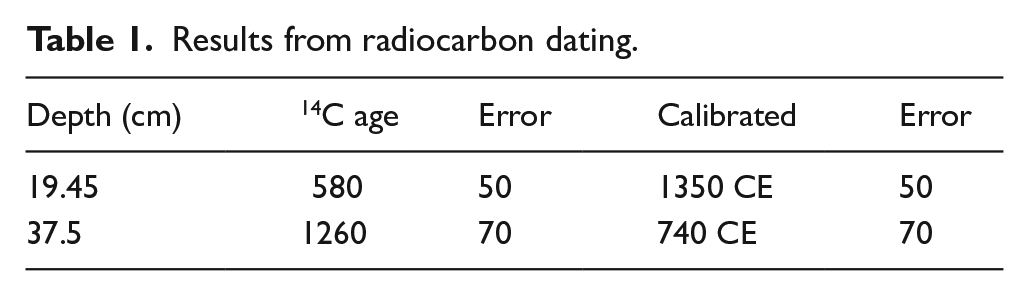
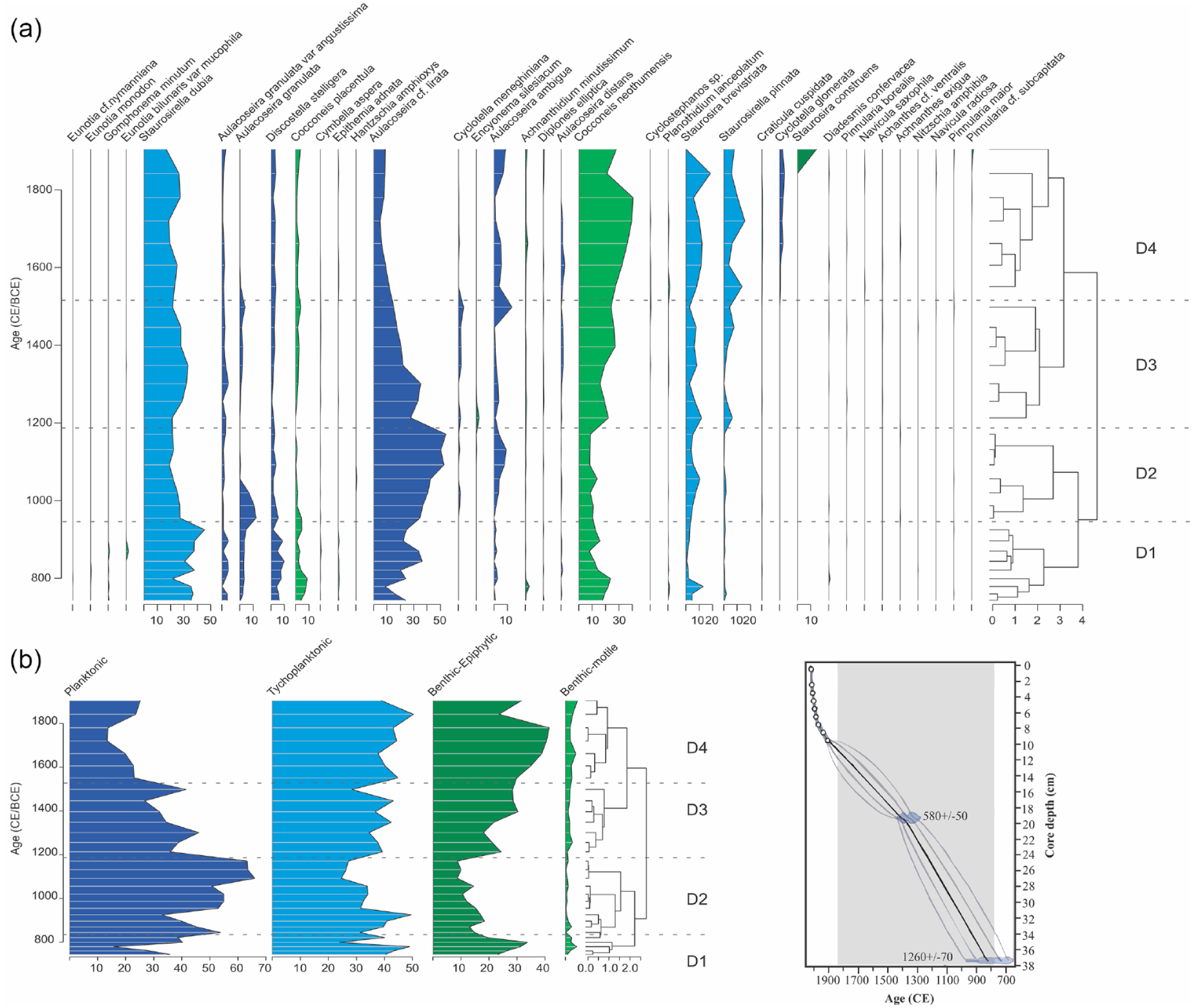
To complement the limnological history inferred from the multi-proxy analyses, we use the record of aquatic plants obtained from the work by Correa-Metrio et al. (2022), Dryad repository at https://datadryad.org/stash/datast/doi:10.5061/dryad.hx3ffbggd. We grouped plants into floating (Sagittaria, Hydrocotile), ferns (Polypodeacea and Pteridophytes), Moss (Selaginella and Lycopodium), emergent (Cyperacea, Polygonium, Typha) and submerged (Myriophyllum, Isoetes, Potamogeton).
Data Analysis
A multiple factor analysis (MFA) (Pagès, 2002) was used to assess the main temporal gradients of change in the diatoms (functional groups), the aquatic vegetation (pollen) and sediment geochemistry (XRF, C/N, and isotopes). MFA allows to cluster the biotic and abiotc data into distinct groups and assess simultaneously the amount of variation explained by each group. The data were standardized during the analysis to reduce skewness and rescale the data into comparable units (Pagès, 2002). The MFAs were performed in R using the package FactoMineR (Pagès, 2002).
Results
The age modeling indicates that the core H-13 covers the last ~1300 years (740 CE). The diatom record (Figure 2) is composed mainly of tychoplanktonic Staurosirella dubia and planktonic, chain forming Aulacoseira species including A. cf lirata, A. granulata, A. distans, and A. ambigua, and epiphytic (Cocconeis neothumensis), and motile species (Achnanthes, Gomphonema, Nitzschia, Navicula, and Craticula spp). Cluster analysis on these groups of diatoms indicate the presence of four main stratigraphic zones (Figure 2) (age in brackets): (1) 39–36 cm (740–800 CE), (2) 35–24 cm (800–1200 CE), (3) 23–17 cm (1200–1500 CE), and (4) 17–10 cm (1500–1900 CE). Zone 1 is composed of variable proportions of all groups. Common species include A. cf lirata, S. dubia, Staurosira brevistriata, Cocconeis neothumensis, C. placentula, A. granulata var. angustissima, and Discostella stelligera. Zone 2 is composed mainly of planktonic (chain forming species A. cf lirata and A. granulata) and tychoplanktonic species which increase and decrease to the top, respectively, and of C. neothumensis. Zone 3 is composed of tychoplanktonics (with increase in Staurosirella pinnata), planktonics (chain forming and single celled) which are significantly reduced compared to the previous zone; epiphytics (Cocconeis, Epithemia, Encyonema spp) and motile species (Nitzschia, Navicula, Craticula, Achnanthidium, etc) increase to the top. Zone 4 is composed mainly of epiphytic Cocconeis neothumensis, tychoplanktonics and colonizer Achnanthidium minutissimum; single cell planktonic diatoms increase gradually and include increases in Cyclotella meneghiniana, Cyclostephanos sp. and the only appearance of Lindavia glomerata. Chain forming planktonic decrease to minimum values in the record.
Diatom diagram (a) and diatom sums according to functional groups (b) with diatom zones (D#) indicated by dashed lines. On the right, age model from Correa-Metrio et al. (2022).
Results from the main element and ratios are presented in Figure 3. Ti/Ca (detrital input) increases gradually from bottom to top with values ranging from 0.5 to 0.9 between 740 and 1500 CE, and above 1.3 after 1700 CE. Concentrations of Mn/Fe (oxygenation) varies from 0.013 to 0.015 between 700 and 1500 CE) and decrease to 0.0099 after 1700 CE. Values for Zr/Fe (flooding) ranges between 0.007 and 0.009 except for a value of 0.0034 at 810 CE. Sedimentary P (productivity dynamics) increases from 0.13 to 0.3 between 810 and 1100 CE and then decreases to around 0.1 after 1700 CE.
Main element ratios used in the study with diatom zones D# (a), and aquatic plants (b) taken from Dryad repository at https://datadryad.org/stash/dataset/doi:10.5061/dryad.hx3ffbggd (Correa-Metrio et al., 2022).
15δN is relatively constant at around 3‰ from 700 to 1500 CE. After this time and until 1900 it increases to 4‰ (Figure 4). After 1900 CE it reaches maxima values of >10‰ 13δC gradually changes from values around −21‰ at the beginning of the record to values close to −24‰ after 1900 CE; there is slight short-lived change to a heavier value at 1400 CE. C/N has values between 10 and 11 from the beginning of the record until about 1200 CE, after this time, values gradually decrease reaching the minimum value (7.8) at 1850 CE, after this time it increases again with values between 9 and 10.

Geochemical stratigraphy of H-13 core.
15δN varies around 2 and 3‰ from 700 to 1400 CE, after this time it gradually increases to 9 in the 20th century (Figure 4). TN ranges between 0.5 and 1 except for the period between 900 and 1200 CE when it increases to 1.5 and 2. C/N ratio has values between 9 and 10 from 700 to 1200 CE and descend to 7.7 at around 1840 CE; it increases again to close to 9 after 1900 CE. TOC and TN have their greatest variations at the beginning of the record between 900 and 1150 CE.
Environmental reconstruction
The most common and abundant diatoms of the record include tychoplanktonic Fragilaria sensu lato (dominated by S. dubia), epiphytic C. neothumensis and planktonic Aulacoseira species (Figure 2). Fragilaria sensu lato, including S. dubia, are common diatoms in the phytoplankton of the lake today and have been for millennia (Cardozo et al., 2005, 2014). Fragilaria spp., are also the most common species in neighboring Andean lakes Fúquene and Siscunsí, and in many other oligo-mesotrophic and transparent Andean lakes (Catalan and Rondón, 2016) most of them shallow basins and with abundant littoral aquatic vegetation (Salgado et al., 2019; Vélez et al., 2003, 2021). The abundance of these species through the record likely reflects the position of the core not very far from the vegetated littoral (represented by the abundance of tychoplanktonic and epiphytic) but under >40 m of water depth (represented by the abundance of planktonics) (Figure 1).
Between 740 and 800 CE (zone 1) the very diverse diatom composition, variations in the proportions of plankton/tychoplankton and in motile species are reflecting variable limnological and or hydrological conditions. These variations could be related to localized hydrological changes as suggested by the increase in flooding (Zr/Fe), the presence of A. granulata var. angustissima, a common riverine species (O'Farrell et al., 2001), and of C. placentula (Figure 2), an epiphytic species that for its habit to live horizontally and firmly attached to the surface makes it particularly tolerant of high energy currents and of physical disturbance (Berthon et al., 2011; Passy, 2007).
Between 800 and 1200 CE (zone 2), the abundance and increasing trend of chain forming planktonic species (Aulacoseira spp in particular A. cf lirata) suggest increasingly good mixing and turbulent water conditions, mainly after 900 CE. A. cf. lirata the most abundant Aulacoseira species, is a robust diatom (diameter ranges between 20 and 30 microns) that likely required turbulence to stay suspended in the water. Greater concentrations of P in the sediment, and relative high values of C/N (10–11) suggest excess of P in the system, and mixed autochthonous and allochthonous sources of organic matter, respectively.
Single cell planktonics Cyclotella meneghiniana, D. stelligera, and D. pseudostelligera present in zones 1 and 2, are known to tolerate turbidity and inhabit waters rich in nutrients (Mejia, 2011; Montoya-Moreno et al., 2013). In particular, D. stelligera is associated with higher levels of DOC (Brown et al., 2017; Malik and Saros, 2016) and with deforestation in nearby lakes Fúquene and Pedro Palo (Vélez et al., 2021), and other lakes (Correa-Metrio et al., 2016; Costa-Böddeker et al., 2012; Köster et al., 2005). Recent increases in this diatom in Ecuadorian lakes has been used as evidence of thermal stratification in paramo lakes due to global warming (Michelutti et al., 2015). Thus that these increase in this species, particularly in zone 1, could be indicating increases in DOC, in turbidity and nutrients waters. C/N in zones 1 and 2 has the highest values in the record, suggesting input from erosion and lake productivity.
Between 1200 and 1550 CE (zone 3), the change in composition of the diatom community in respect to the previous 500 years suggests a shift in the ecosystem and ecology of the lake. The notable decrease in A. cf lirata can be related to changes in the water column induced by reduced turbulence and or water transparency, as mentioned before this is a robust diatom that required turbulence; the increase in epiphytic species suggests a shift in macrophyte composition and or extension of the plant cover. The aquatic pollen shows that floating plants are significantly reduced after ~1200 CE while emergent, submerged, fern and mosses recover and increase their abundance after a significance reduction at ~1300 CE (Figure 3). Motile species also increase that could be related to ecological stress, as motile diatoms thrive over the non motile for their capacity to avoid stress by moving away from it. Since this guild is affected negatively by turbulence (Passy, 2007) it could also be indicating a reduction in water turbulence, also explaining the decrease in Aulacoseira spp. The decrease in P in the sediments could be indicating more uptake in the water column either by plants or other algae. More in-lake productivity is also suggested by the low C/N values, possibly related to changes in land use in the watershed as suggested by trend to more negative values in δ13C (Figure 3) or in the macrophyte composition and abundance, or both. Detrital input (Ti/Ca) continues to increase. All these changes suggest an expansion of the littoral zones of the lake, and overall unfavorable conditions for chain forming plankton, likely due to a reduction in water levels, turbulence, and mixing.
From 1550 to 1900 CE (zone 4), the continuous increase and peak in motile diatoms and of colonizer Achnanthidium minutissimum, also associated with disturbance by agropastoralism in the Ecuadorian Andes (Benito et al., 2019), indicates that ecological stress from the previous zone continues and likely intensified. The maximum abundance of epiphytic diatoms indicate macrophyte extension while the only appearance of L. glomerata and Cyclostephanos likely indicates increase in nutrients (Anderson, 1990; Brown et al., 2017); erosion continues to increase and periods of anoxia seemed more recurrent at the same time that the lake was more productive (low C/N and sedimentary P). Variable conditions were identified in the small lake basin where A. minutissum and Cocconeis spp also increased (Cardozo et al., 2014). All these seem to indicate a period of stress and disturbance, littoral conditions and a more productive lake. After 1900 CE the increases in TN and 15δN likely reflect the start of an increasing cultural eutrophication from human waste and from fertilizers from agriculture and aquaculture, practices that characterize Tota today.
Results from the MFA analysis (Figure 5a) revealed three lakes states: one with tychoplanktonic and benthic species and more productive autochthonous organic matter (negative axis DIM2), another one initially with planktonic diatoms, floating plants and flooding and then with more diverse plants and tychoplanktonics, and a third one with more benthic diatoms, where erosion and nitrogen content (15δN) are more prominent. The later aligned mainly along the positive axis of DIM1. The MFA factor map (Figure 5b) shows a gradual change from zone 1 to 4, with zone 4 in a more distant position from the rest of the zones (all samples from this zone in the positive axis of DIM 2).
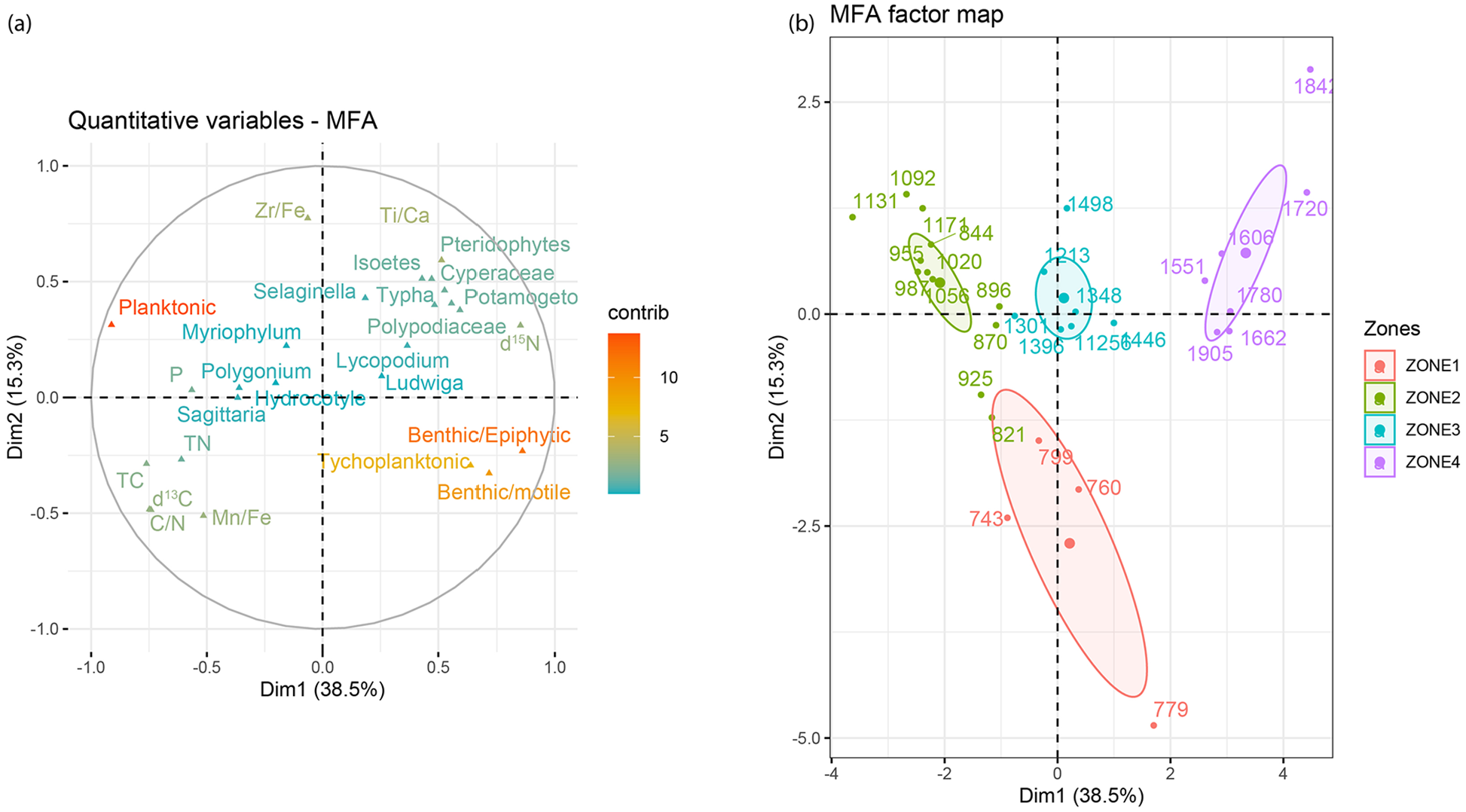
Multi factor analysis for a databased composed of diatoms, aquatic plants, and geochemistry of the core (a) and MFA map for all samples (b, numbers reflect time in CE).
Results from the analyses reveal key moments of limnological and ecological change in the studied basin of Lake Tota. The first change is dated at 1200 CE. At this time the lake changed from mainly a planktonic diatom state with floating plants (zones 1 and 2) (MFA results; Figure 5a) to a state with a more prominent benthic signal (zone 3) and littoral vegetation (Figures 3 and 5a). This change is also observed in the terrestrial vegetation that changed from a diverse forest with open vegetation to a less diverse composition with more abundance of montane and submontane forests after 1250 CE (Correa-Metrio et al., 2022).
To try to explain what could have happened at 1200 CE, natural (climate, lake dynamics, and ecology) and anthropogenic causes need to be considered. In the following lines, we provide an archeological and a paleoclimatic context for this period and discuss this change under possible anthropogenic and or natural causes.
Distinct archeological indicators such as settlement patterns and cemetery size, 14C chronology, paleogenomics, land use practices, and agricultural infrastructure, point to an important increase in population size during the early Muisca (1000–1200 CE) but specially during the late Muisca period (1200–1600 CE). With the population growth there was a significant increase in sociopolitical, economic and religious complexity (Table 2) (Argüello, 2015; Boada, 2007; Boada Rivas, 2018; Delgado et al., 2021; Delgado Burbano, 2012; Fajardo, 2016; Kruschek, 2003; Langebaek, 1995, 2019). This growth was particularly pronounced in the Sogamoso Valley, where Total Lake is located, and where high population numbers were maintained until the Colonial period (Fajardo, 2016; Langebaek, 2019; Tables 2 and 3). The estimated Muisca population by the time of the Spanish arrival was more than 3 million (Francis, 1993), although ethnohistoric records suggest population sizes ranged from 1200.000 to 2400.000 persons (Langebaek, 2019). Population density and occupation area in the Sogamoso Valley peaked in the early Muisca with 16.9 persons/km2 and 224.98 ha occupied (Table 2). Along with this increase, the production of crafts such as textiles, pottery and goldsmithing, intensified and diversified (Argüello, 2015; Boada Rivas, 2018; Delgado, 2021; Fajardo, 2016; Langebaek, 2019). Pottery manufacture required wood, the main source of heat for the ovens, and as such deforestation was rampant (Mora-Pacheco, 2012). Muiscas had two types of gardens, small scale with diverse produce that sustained a household, and large scale, communal, used to feed a larger community and that consisted of specialized, few produce (Mora-Pacheco, 2012) and terraces or camellones with drainage systems were built for garden protection against floodings (Boada Rivas, 2018). Soils were fertilized with decomposed waste from previous harvest by planting a mixed garden involving natural nitrogen fixators such as beans, and adding animal dung (Szpak, 2014), and parcels were left to rest uncultivated for few years. All these activities highlighting an impressive agricultural infrastructure compatible with both extensive agriculture and population density (Table 3) thus, a significant land use alteration. Lakes on the other hand, were also used. Many lakes and wetlands were ceremonial centers for rituals and sacrifices in addition of being sources of sustenance. Lake Tota for instance was an important source of animal protein (Langebaek, 1987) and a religious center throughout the Muisca period, well until the Colonial period (Mora-Pacheco, 2012; Quintero-Guzmán, 2023), and even today. Interestingly, settlement patterns in Sogamoso and Tunja chiefdoms during the late Muisca period suggest more interaction between such settlements and the promotion of the Tota capitanía (zybyn in Muisca, translates to a concept similar to a chiefdom), as turning more important or independent socio-politically (Fajardo, 2016; Fajardo and Argüello, 2024). This can be interpreted as a more profuse effort of local chiefs to increase and demonstrate power where the Lake Tota played a key role through feasting and offerings.
Sogamoso valley population estimates. a

Data from Fajardo (2016); min, average, and max values for absolute number of inhabitants.
Area of occupation for several settlements across the Altiplano Cundiboyacense.

The limnological change at 1200 CE is also correlated with the transition between two important periods of climate change: the Medieval Climate Anomaly (MCA) and the Little Ice Age (~950 to 1250 and ~1400 to 1800 CE, respectively) (Mann et al., 2009) from which there is not a final consensus on how they were manifested in the in the northern neotropics. Given that the region around Tota is highly influenced by the ITZC and its dynamics with the Tropical Atlantic and the Trade Winds, we compared our record against others records from the Caribbean, Panama, Venezuela, and North-East Peru, also affected by changes in the Atlantic Ocean. In Venezuela, the record from Cariaco, indicates overall high precipitation during the MCA and dry conditions in the LIA (Haug et al., 2001) reflecting the position of the ITCZ north and south respectively, and a record from the Venezuelan Andes indicate cooler and wetter conditions during the LIA (Polissar et al., 2006). In Cuba wet conditions prevailed during the MCA while dry conditions during LIA, caused by a cold state in the Atlantic Multidecadal Oscillation (Fensterer et al., 2012). A record from Nicaragua, also a proxy for teleconnections between the Pacific and the Atlantic, is interpreted as indicative of a La Niña-like state and a positive state in the North Atlantic Oscillation (NAO) during the MCA, and a El Niño-like state and negative NAO during the LIA (Stansell et al., 2013). This agrees with a reconstruction of the tropical state of the Pacific that indicates a La Niña state and low variance during the MCA and El Niño like state with high variance during the LIA (Jiang et al., 2023). High variability during the LIA has also been reported from Barbuda where periods of rainfall and drought alternated on interannual and interdecadal scales (Burn et al., 2016) which would agree with the Spanish chronicles of the Tota region in that report frequent droughts and floods (Mora-Pacheco, 2019). A record from Northeast Peru suggests that conditions during the LIA were wet, due to a cooling of the Tropical North Atlantic in the boreal spring. The above suggests that the climatic manifestation of the MCA, LIA in northern South America is still not resolved. Based on changes in terrestrial vegetation around Lake Tota, Correa-Metrio et al. (2022) conclude that the MCA was warm and dry, inferred from the abundance of herbaceous and open vegetation of the record, and that the LIA was cold and relatively wet, as indicated by the increase in arboreal and forest vegetation, and that the transition between these two climate states (~1200 CE) was wet and colder, compared to the MCA. Warmer and drier conditions during the MCA and wetter and colder conditions during the LIA in the Eastern Cordillera of Colombia are also proposed by Bird et al. (2024) based on sediment grain size and geochemistry.
The second period of limnological change occurred after 1500 CE and is correlated with the arrival of the Spanish and the well documented transformation of Lake Tota’s watershed during this time: significant increase in deforestation from the expanding population (Tables 2 and 3), agriculture and husbandry and the foundation of villages and subsequent erosion and animal and human waste (Cuéllar, 2010). At this time, deforestation and desiccation of wetlands and lake littorals were common practices used to accommodate the increasing number of cattle ranches, and the expansion of agriculture and settlements (Cuéllar, 2010; Fajardo, 2016). For instance, the close by towns of Tota and Cuitivá were founded in 1535 and 1570 CE respectively.
Discussion
Based on the occupation history of the area, the limnological shifts identified here can be representing the gradual increase population density and consequent changes in land use concomitant with socio-economic expansion and diversification. Excess of sediment and organic matter produced by deforestation, soil fertilization and agriculture, may have altered lake’s DOC, warming the epilimnion and reducing water mix, affecting the P/N cycles and increasing macrophyte extension. It is worth noting that pollen grains from maize have not been detected in the record from Lake Tota (Correa-Metrio et al., 2022). Their absence could be explained by a number of reasons including maize crops being planted father from Lake Tota and closer to the nearest city of Tunja, one of the biggest and most socially complex chiefdoms and not in Tota’s watershed, and diet habits, and although maize was important in the Muisca diet, recent studies derived from stable isotope analyses on bone and teeth, indicate that people from the Herrera and Muisca periods consumed maize, C3 plants and animals, in similar proportions or even with higher amounts of C3 resources (plants and herbivores) (Delgado, 2021; Rivas et al., 2024).
Not only Lake Tota, but other lakes in the Muisca region (Table 3) were also affected by human activities. Fúquene suffered from deforestation and drainage alteration (Mora-Pacheco, 2012; Vélez et al., 2003), Pedro Palo, south of Lake Tota, was not a ceremonial center but it was located along a trade route to the western lowlands and it was impacted by deforestation and burning by indigenous activities as far back as 2000 year BP that altered DOC and created a shift in the planktic/benthic diatom ratio (Vélez et al., 2021). In other parts of South America, lakes were also affected by human practices. In Laguna de Los Cóndores, another ceremonial lake in Peru, anthropogenic erosion in the watershed impacted lake’s water quality (Matthews-Bird et al., 2017). In Lake Llaviucu in Ecuador, pastoral and agricultural activities, including burning, affected the lake and left their marks on the lake’s record (Benito et al., 2022).
The manifestation of the climatic changes in northern South America related to the MCA and LIA is still debated, however based on pollen and grain size and geochemical records Correa-Metrio et al. (2022) and Bird et al. (2024) suggest that conditions during the MCA were warm and dry, shifted to cold and relatively wet during the LIA. The shift to a benthic state in zones 3 and 4 could be due to an expansion of the macrophyte cover from increasing lake levels and the inundation of previously exposed littoral areas.
The MFA factor map (Figure 5b) suggests that the changes between diatom zones and lake states have been gradual, and that zone 4 could be representing a novel lake state. Records from mountain lakes in the neotropics show that many lakes in this region are in a novel diatom state induced by anthropogenic activities (Vélez et al., 2021). The shifted to a new state after 1500 CE in this record could be a novel state in the lake. Since the record analyzed here could have been affected by human activities that have been present throughout the recorded interval, pre-human baselines conditions for the lake could not be identified. A longer archive is needed to look at natural pathways of change.
Implications and conclusions
Three main limnological shifts in Lake Tota were identified in this study at 1200, 1500, and 1900 CE. These changes occurred simultaneously with the change from the Herrera to the Muisca archeological stages, increase in indigenous population, the Spanish arrival and the industrial revolution, respectively, and with the MCA and the LIA climatic periods. Besides the changes in the lake occurring with changes in human activities, the role of climate change can’t be ruled out. Either triggered by anthropogenic activities and or climate change, the Lake Tota has responded sensitively to external forcing causing a change in the diatom flora planktonic to benthic.
The gradual eutrophication of the lake today, primarily caused by agricultural and aquacultural activities is of great concern (Aranguren-Riaño et al., 2018) and may represent a completely new state unprecedented in the history of the lake. These human interventions are disrupting the lake’s ecological balance and have set it on a path toward eutrophication, where excessive nutrient inputs will lead to increased primary productivity and subsequent changes in the lake’s ecosystem structure and function. If left unchecked, the lake may eventually reach a novel state from which it will be difficult to reverse the eutrophication process.
Footnotes
Acknowledgements
Our thanks to Rebecca Jamieson, University of Regina for running the XRF analyses, the Smithsonian Tropical Research Institute Senior Fellowship for supporting JE, the “Corporación Auntónoma Regional de Boyacá (CORPOBOYACÁ)” for all their assistance during field work and core extraction.
Author contribution(s)
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Sciences and Engineering Research Council of Canada Discovery Grant, the United States National Science Foundation [EAR 1231445649] and Indiana University-Purdue University, Indianapolis, IN (RSFG & IDF).