
Abstract
This paper presents new data derived from pollen, starch and radiocarbon samples that were collected from six Aboriginal earth (oven) mounds and middens on the Calperum and Pike floodplains, Murray-Darling Basin (MDB), South Australia (SA). Analyses of these samples were undertaken in order to enhance our understanding about the Holocene lifeways of Aboriginal people living in this region. The results from these analyses, combined with published data about the mounds’ contents, relevant ethno-historical information and climate data, allow us to infer that Aboriginal people adopted, in this region, an innovative food production system about 3800 years ago. Further, we argue that the timing of the change suggests this was in response to adverse El Niño Southern Oscillation (ENSO) related weather patterns and consequently fluctuations in both terrestrial and aquatic food resources.
Keywords
Introduction
It has been argued that Australian Aboriginal populations broadened their diets during the mid-Holocene in response to environmental challenges, associated demographic and/or social factors (Edwards and O’Connell, 1995; Haberle and David, 2004; Williams et al., 2015a, 2015b). 1 The nature of such responses included a more intensive use of previously marginal regions, and the targeting of subsistence resources such as emergent macrophytes, eels, toxic nuts and grass seeds which require extensive processing and/or enhanced logistical strategies to maximise the nutrition available (Edwards and O’Connell, 1995; Haberle and David, 2004; Jones, 2023; Jones et al., 2022; Lourandos, 1983, 1997; Lourandos and Ross, 1994; Pardoe, 1995). Expanded food harvesting and production systems led to broad spectrum diets (BSD) (see below).
In some floodplain environments of the Murray-Darling Basin (MDB), in south-east Australia (Figure 1), it has been argued that there was a mid-Holocene focus on specific wetland features together with the introduction of innovative management and processing, including the targeting of established emergent macrophyte habitats, the use of ground ovens, and the digging and harvesting of aquatic rhizomes for cooking and consumption (e.g. Gott, 1999; Jones et al., 2022). The resultant archaeological signature of these activities was the creation of earth oven mounds near swamps, billabongs and anabranch creeks, consisting mainly of ashy sediment, charcoal and clay heat retainer nodules, with smaller but variable quantities of bone, mussel and other gastropod shell (Balme and Beck, 1996; Berryman and Frankel, 1984; Brockwell, 2001a, 2001b, 2005, 2006a, 2006b, 2009; Brockwell et al., 2017; Coutts et al., 1976; Godfrey et al., 1996; Johnson, 2004; Jones, 2016, 2023; Jones et al., 2017, 2022; Klaver, 1988; Lane, 1980; Martin, 2006, 2011; Pardoe, 2003; Pardoe and Hutton, 2020; Westell and Wood, 2014). Such techniques allowed the efficient extraction of a previously marginal and low trophic plant food, which emerged as traditional resources declined. Factors possibly involved in this transition include climate variation, demographic pressure and over-predation of traditional food resources. This paper aims to further explore the relationship between Holocene diets, climate and socio-economic innovation via the analysis of new data derived from pollen, starch and radiocarbon samples that were collected from six Aboriginal earth (oven) mounds on the Calperum floodplain in South Australia’s (SA) Riverland region (Figures 1–3).
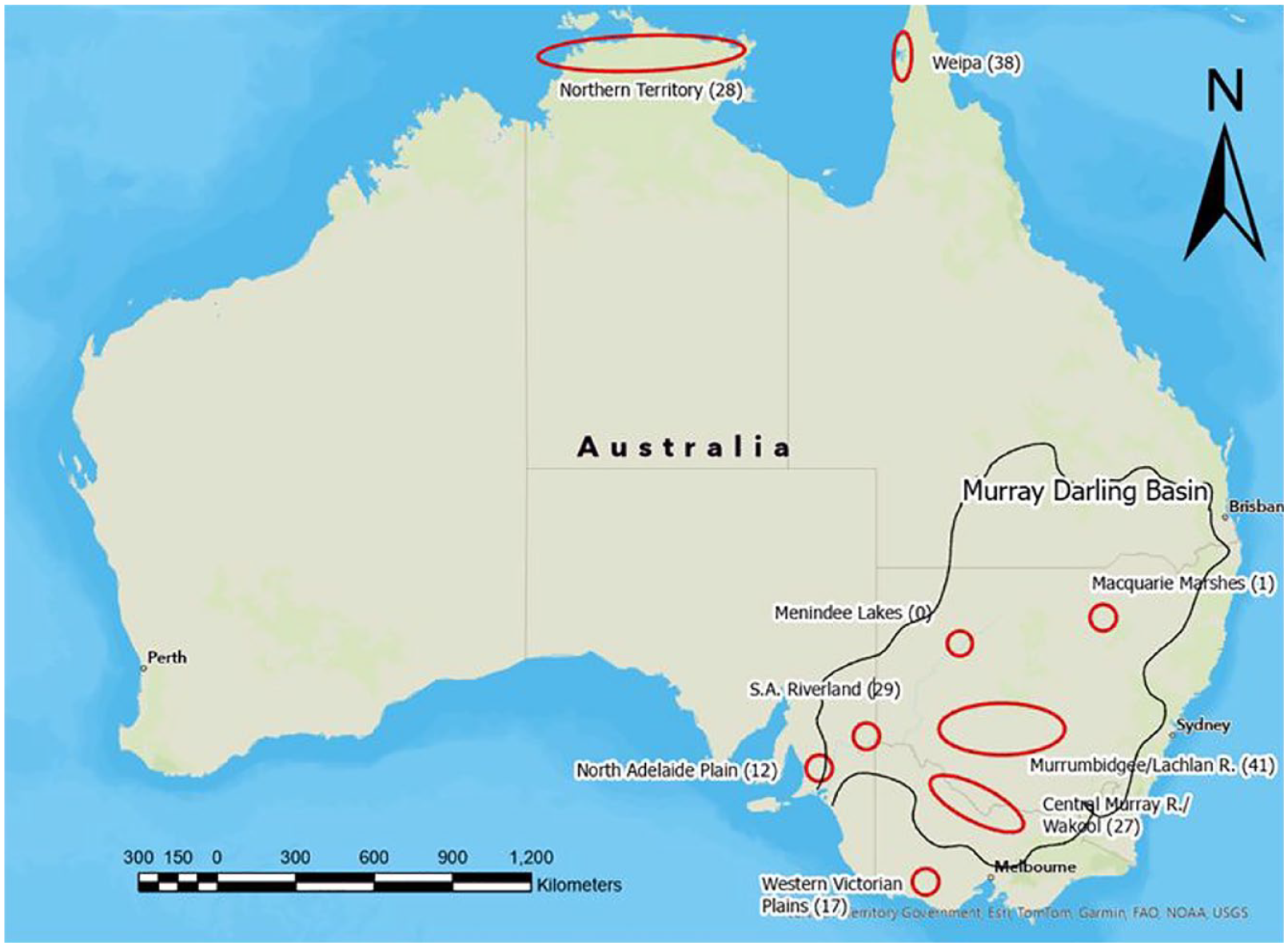
An approximation of earth mound precincts in northern and south-eastern Australia showing the number of 14C dates by region where available (Balme and Beck, 1996; Berryman and Frankel, 1984; Brockwell, 2001a, 2001b, 2005, 2006; Brockwell et al., 2017; Coutts et al., 1976, 1979; Godfrey et al., 1996; Johnson, 2004; Jones, 2016; Jones et al., 2017; Klaver, 1998; Martin, 2006; Westell and Wood, 2014; Williams, 1987).
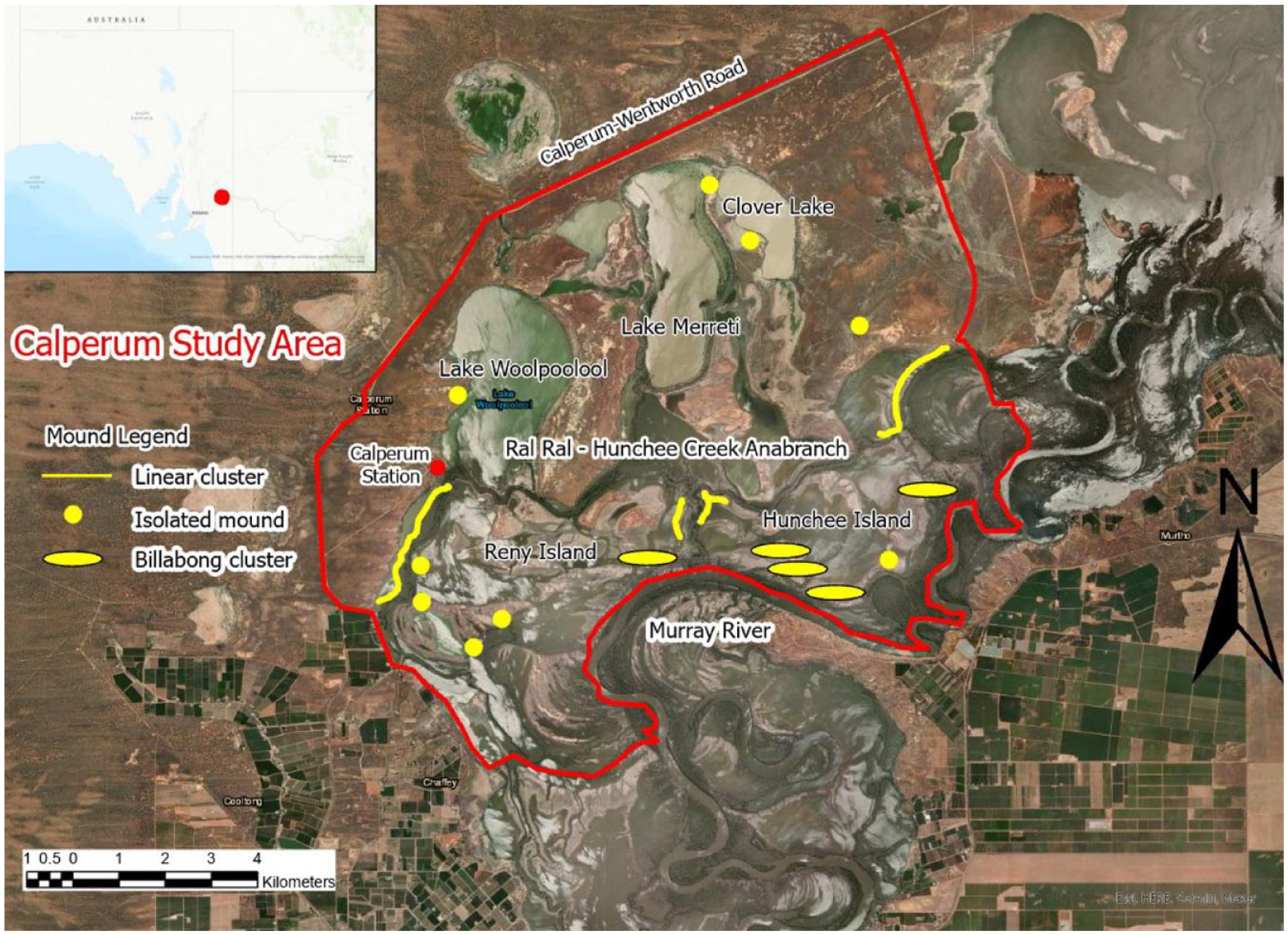
The Calperum study area, including the location of isolated and groups of mounds.
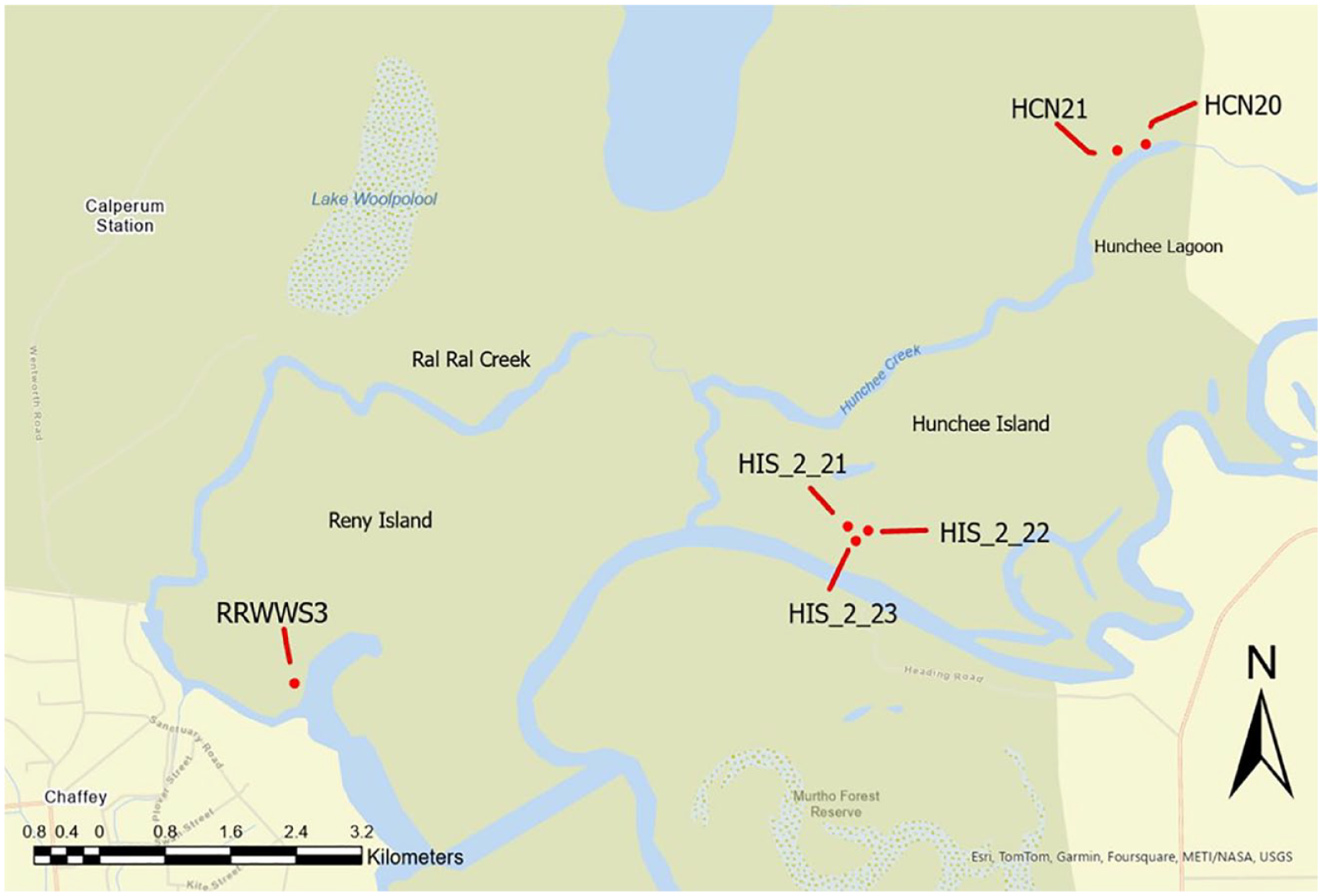
Locations of excavated earth mounds.
Australian Aboriginal earth mounds
Earth mounds in Australia have been recorded in a number of discrete geographic areas including western Victoria, the northern Adelaide plains, the MDB, the Northern Territory and in north-west Cape York Peninsula (Figure 1). They are typically found near water sources in or near areas that were regularly inundated and have dates of less than 5000 cal BP. Earth mounds are one expression of a typological continuum of open sites that have been under-utilised as a potential source of information on regional socio-economic trends during the mid-to Late-Holocene (see Jones, 2023; Jones et al., 2022).
The global context
BSD are associated with a hypothesis which proposed a transition in human subsistence systems during the terminal phase of the last glaciation that was labelled ‘the broad-spectrum revolution’ (BSR) (Flannery, 1969). This was a diverse phenomenon involving a range of food resources and subsistence systems which developed (or propagated) over millennia in a variety of locations, around the world (Edwards and O’Connell, 1995; Zeder, 2012). This phenomenon has been variously attributed to the influence of intra-group social change and the development of complex social organisation, the introduction of new technologies, sedentary behaviours, demographic change, new food storage strategies and climatic change associated with the terminal Pleistocene in western Asia (Edwards and O’Connell, 1995; Flannery, 1969). The BSR became synonymous with dietary diversity, resource intensification and stress associated with population growth and resource availability (Zeder, 2012).
Socio-economic intensification and BSD in Australia from the mid-Holocene
Edwards and O’Connell (1995) examined BSD in Australia with reference to the arid zone (i.e. that region where annual evaporation equals or exceeds precipitation), which constitutes ~70% of the continent. This region has been the subject of a number of studies into occupational history and lifeways, processing technologies and resource exploitation, focussing on responses to environmental changes and socio-economic drivers (Edwards and O’Connell, 1995: 774). Edwards and O’Connell (1995) attributed the global expression of the BSR to ‘very general processes’, active in diverse locations within a relatively similar time frame. In Australia, however, the archaeological record suggests that the ‘revolution’ occurred significantly later, from the mid-Holocene. Such evidence and interpretations indicate that climatic conditions associated with the terminal LGM did not trigger changes in diet breadth at that time, prompting a continuing debate as to the reasons for this apparent temporal anomaly (Frankel, 1995).
In Australia, the evidence for a post-5000 cal BP social and economic ‘intensification’, often cited in support of a socially driven ‘continental level’ hypothesis for change, includes the intensive exploitation of marginal and low trophic foods such as seeds, cycad nuts, terrestrial and aquatic underground storage organs (USO), inter- and intra-regional trade, fish traps, fresh water eel management, diffusion of art styles and increases in population, sedentism and social complexity (David et al., 2006; Frankel, 1995; Lourandos, 1983, 1997; see also Beaton, 1985). The ‘continental intensification model’ for Australia is associated with questions about change and dynamics within past Aboriginal societies, including an emphasis on demographic, socio-economic and socio-cultural factors (Beaton, 1977, 1985; Bowdler, 1981; Lourandos, 1983; Lourandos and Ross, 1994; Ross, 1985; Ross et al., 1992; Williams, 1987). The model presented arguments against traditional passive views of ‘hunter-gatherer’ groups, based on evidence that indicates the potential role of internal influences (Beaton, 1983; Lourandos, 1980; Lourandos and Ross, 1994). Such influences include demographic, social and economic factors affecting Aboriginal societies, particularly during the mid-to Late-Holocene. Hiscock and Sterelny (2023) recently presented a contrasting perspective that considered whether Aboriginal peoples responded to . . . ‘climatically driven reductions in higher-ranked resources’ and consequently reconfigured ‘their economic strategies and social interactions’ from the mid-Holocene.
Ulm (2013: 183–185) provided a general perspective of intensification at the continental level and argued that ‘regional cultural trajectories need to be disarticulated from the continental narrative to enable independent characterisation of local behavioural variability’. He emphasised the need to address chronological control, sampling and taphonomy and suggested that researchers refocus on open sites instead of rockshelter deposits in order to gain a greater picture of regional trends (Ulm, 2013).
Climate considerations
A review of climate research literature (Tables 1 and 2) identifies instances of climate which may have influenced Australian Aboriginal subsistence systems in wetland environments in the MDB that contain earth mounds. The Australian climate is dominated by two key systems; the tropical northwest monsoon, which impacts the north of the continent during the Southern Hemisphere summer, and the westerly weather patterns of the Southern Ocean. In addition to these main influences, other cyclical phenomena also impact rainfall patterns across Australia. These include the El Ni
Correlation of earth mound activity to climatic variation within the MDB.
Sampling detail for the pollen and starch analyses of selected Calperum mounds.
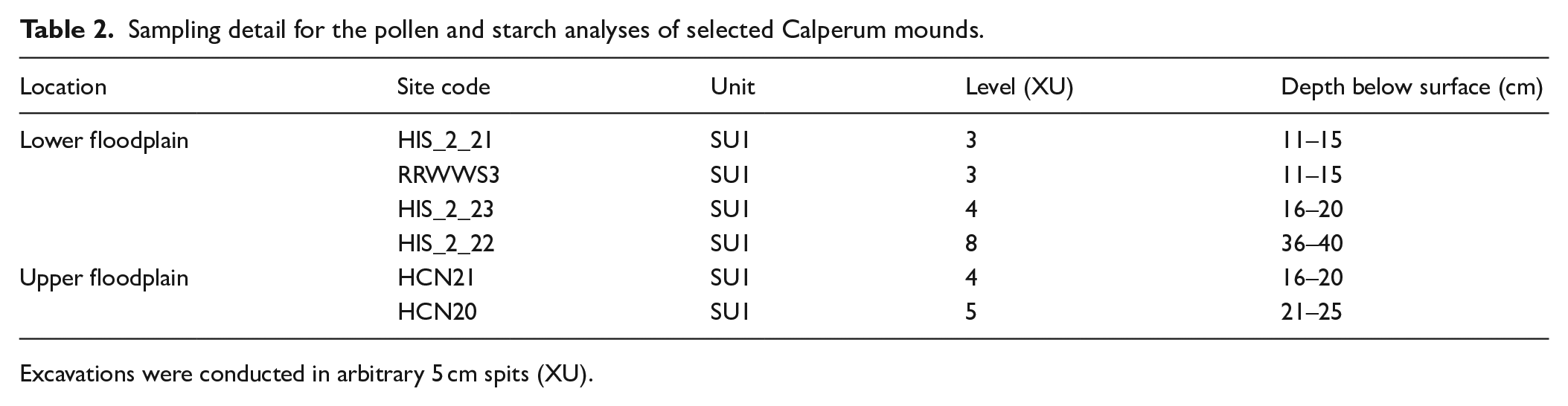
Excavations were conducted in arbitrary 5 cm spits (XU).
Higher precipitation levels evident on the east coast of Cape York, in contrast to the western coast (Stevenson et al., 2015) and Arnhem Land (Fitzsimmons et al., 2013; Hesse et al., 2004), were attributed to easterly trade-winds during the winter dry season, thereby emphasising the influence of geographic and environmental variation on local climate conditions (Luly et al., 2006). In this context, it should be noted that earth mounds have not yet been reported on the eastern side of Cape York, possibly supporting the influence of climatic factors on resource procurement systems as key to their origin and use. A decrease in runoff from about 15,000 to 13,000 cal BP in the south-east of the continent was variously attributed to moisture uptake by returning vegetation, decreased snowmelt and the influence of peri-glaciated ground (Fitzsimmons et al., 2013; Hess et al., 2004).
Generally wetter conditions prevailed from about 7000 to 5000 cal BP in south-western Victoria as indicated by high lake levels (Bowler and Hamada, 1971; Fitzsimmons and Barrows, 2010; Fitzsimmons et al., 2013; Jones et al., 1998) followed by lower discharge from MDB rivers and a trend toward aridity after 5000 cal BP (Gingele et al., 2004, 2007; Petherick et al., 2013; Shulmeister and Lees, 1995). A study of diatoms and macrophyte pollen at Tareena Billabong, at the eastern edge of the Chowilla floodplain in the Riverland of South Australia (Figure 4), suggested Tareena Billabong was a permanent fresh lagoon isolated from the main channel at about 5000 cal BP with reconnection occurring after 3800 cal BP with the likely onset of wet-dry cycles from that time (Gell et al., 2005).
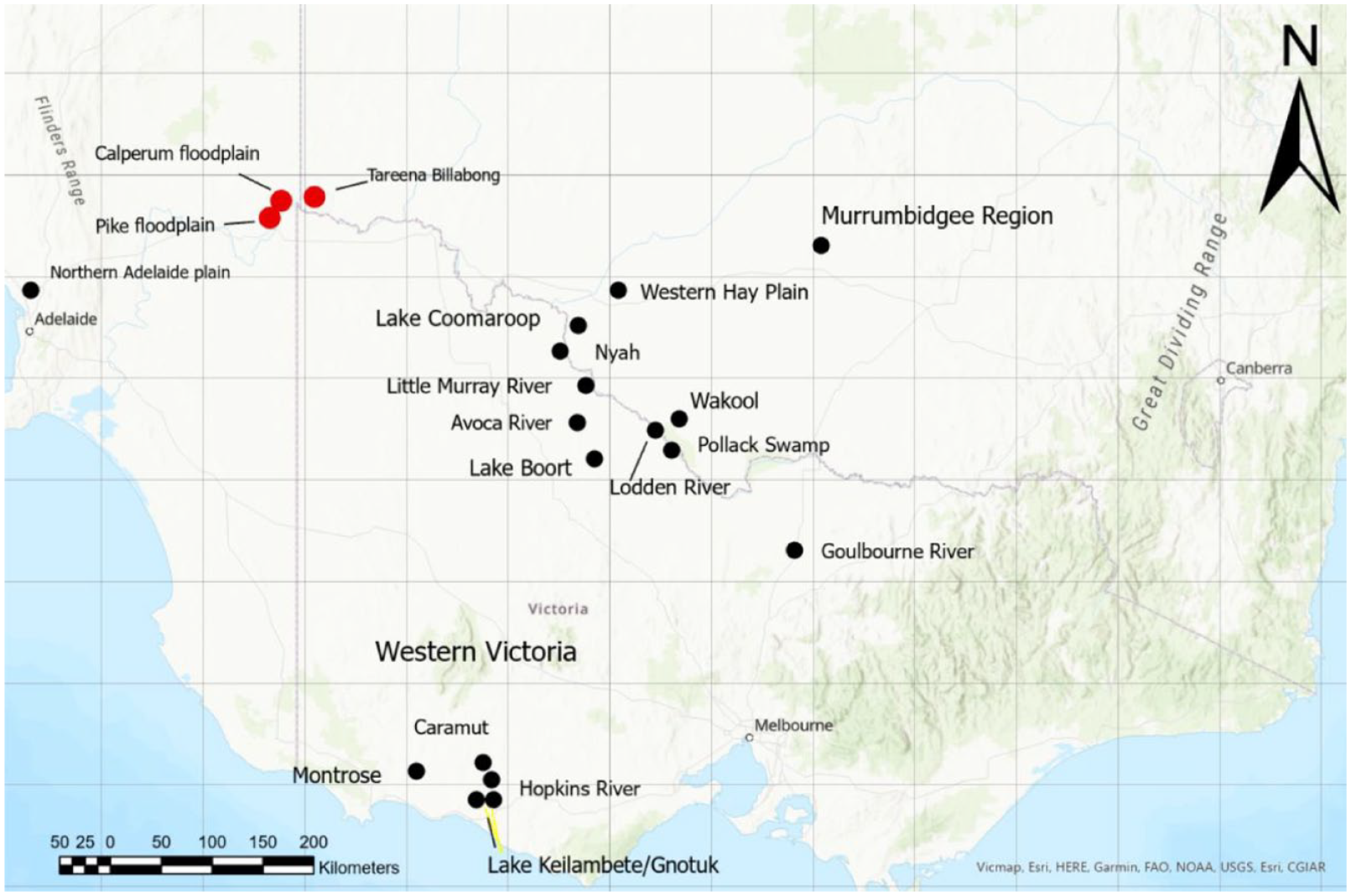
The locations of the Tareena Billabong, Pike River and Chowilla floodplains (red dots), Lakes Keilambete/Gnotuk and the major mound precincts of the central Murray, Murrumbidgee, western Victoria, and northern Adelaide Plains regions (black dots).
As climatic variability and aridity increased, it has been suggested that available resources likely decreased, leading to impacts on group size and diet choice. The demographic analysis by Williams et al. (2015b) suggested a hiatus in the rate of population growth across Australia from 5000 to 4800 cal BP coinciding with the earliest earth mound dates (Figure 5). Thus, the emergence of earth mounds is potentially an earlier response to environmental variability than an intensification of seed grinding and the exploitation of toxic plants. A resumption in the rate of population growth identified by Williams et al. (2015b) after about 3800 cal BP was potentially due to a reorganisation of resource procurement strategies in response to the onset of ENSO induced climatic variability from at least 3800 cal BP.
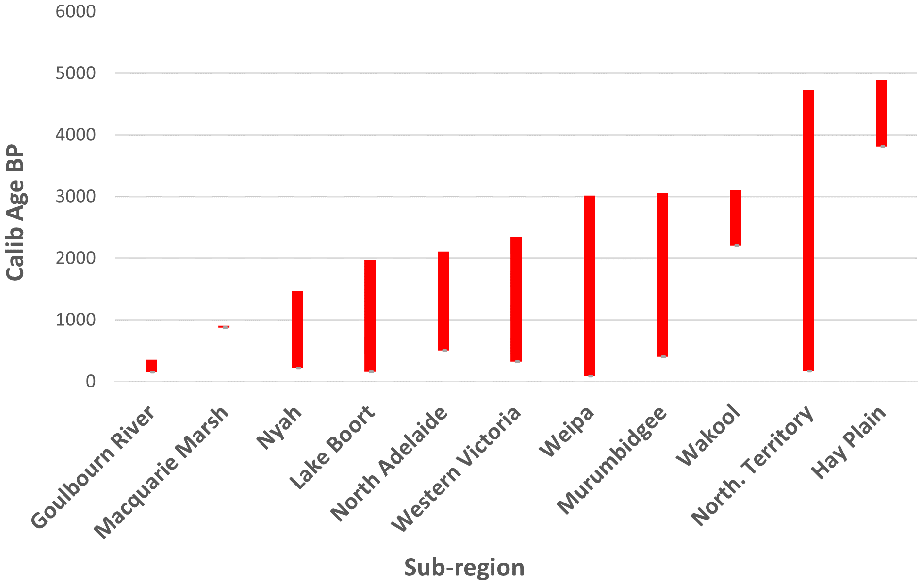
Age ranges of dates across the major mound precincts in Australia (Jones, 2023).2 Calperum dates are excluded. Note the 800 year gap between dates reported by Martin (2006), near Balranald in the Hay Plain, and the MDB mound precincts of Goulbourn River, Macquarie Marsh, Nyah, Lake Boort, Murrumbidgee, and Wakool (Figure 4).
ENSO related climatic variability, in the Australian region from about 5000 cal BP, coincided with the apparently simultaneous appearance of earth mounds in both the north and south of Australia as indicated by available 14C data (Table 1 and Figure 5). The correlation of the apparent emergence of earth mounds over two widely separated regions at about the same time is intriguing, and potentially indicative of a relationship between wider environmental factors, resource procurement and socio-economic transitions at both local and regional levels. Such correlation is key to the identification of potential influences behind the introduction of previously marginal foods (which are often subject to more complex preparation requirements) and the potential broadening of Aboriginal diets at this time.
The Calperum study area
The SA Riverland region contains a number of contiguous anabranch systems between the Calperum floodplain and the SA/Victoria border (Figures 2 and 4). The region is characterised by complex floodplains containing seasonal flow paths, oxbows, ephemeral lakes, lagoons, billabongs and swamps, which can extend 10 km from the main river channel (Figure 2).
Over 60 mounds have been identified within the boundary of the Calperum study area in close association with anabranch channels and the margins of billabongs in the more regularly flooded (lower) parts of the floodplain (Figure 2) (Jones, 2016, 2023; Jones et al., 2017; Westell and Wood, 2014; see Jones et al., 2022 for a detailed summary of the study area).
Prior published research by Jones et al. (2022), which was based on content and sediment analyses of the same six earth mounds that are the subject of this article, revealed that the Calperum mounds contents ‘primarily included anthropogenically burnt clay (heat retainers), charcoal, fragments of freshwater mussel shell and aquatic snails as well as very minor quantities of other faunal material and stone artefacts’. Whilst sediment analyses contributed to the interpretation of mound formation processes (Jones et al., 2022). The minimal amount of faunal material (other than mussel shell and snails), artefacts and a general lack of other archaeological evidence apart from clay heat retainers, confirmed that these features were single purpose and not used as living areas (Jones et al., 2022). This conclusion, combined with the following points, led to the interpretation that the mounds were ‘a food-production procurement strategy based on heat retainer technology and the exploitation of emergent macrophytes’ (e.g. Typha spp. roots) (Jones et al., 2022):
Anabranch channels and the margins of billabongs represent prime habitat for emergent macrophytes such as Typha spp. (bulrush) and Phragmites spp. (common reed).
Ethno-historical accounts indicated that earth mounds were used to cook Typha spp. roots for food and fibre in large quantities in the MDB including in the area adjacent to the study area, as well as other food items (e.g. Beveridge, 1869, 1889; Eyre, 1845: 2; Kirby, 1895; Mitchell, 1839: 2); see also Gott (1982: 61) and Beveridge (1889: 71) for descriptions of the method of starch extraction by Aboriginal people after cooking and the recovery of fibre for the production of string).
Large quantities of clay heat retainer nodules in the Calperum mounds suggested that they resulted from the continuous use of earth ovens in specific locations (e.g. Jones, 2016, 2023; Jones et al., 2017; Martin, 2006, 2011; Westell and Wood, 2014).
The presence of aquatic snail species in mound contents may be inferred as being a bycatch that was transferred to the mounds along with the mud that adheres to the roots of freshwater plants such as Typha spp. (Jones et al., 2022). Snail shells were noted in various levels of all excavated mounds. Quantification and identification of all species of snail taxa found were thus interpreted as a potential proxy for the harvesting of freshwater plants. In total 38 specimens of native freshwater taxa from the Planorbidae family were recorded, including the genera Isidorella (n = 35) and Gryaulas (n = 3) and two specimens of the genus Plotiopsis of the Thiaridae family.
Methods
Pollen and starch analyses, and radiocarbon dating were used in this study to further investigate the potential relationships between the establishment of earth mounds at Calperum, food resource availability and variable weather patterns indicated by climate research within the Australian region from the mid-Holocene (e.g. Gell et al., 2005). In particular, pollen analysis was included to explore correlations with ecological conditions on the Calperum floodplain to those reported and dated by Gell et al. (2005) at the Tareena Billabong located 25 km to the east (Figure 4). Starch analyses were undertaken to investigate the potential presence of macrophyte starches. Radiocarbon dates were obtained (in addition to those presented in Jones et al., 2022) in order to further clarify the ages of operation of the mounds (see below).
Pollen and starch
Pollen signatures were obtained from samples across the six excavated Calperum mounds for a comparison of environmental vegetation profiles present (Table 2). An 800–1000 g sediment sample from each mound was analysed at the Palaeo Research Institute in Golden, Colorado, USA. Levels selected were confined to levels 3–5 except for level 8 of mound HIS_2_22 which was one of three mounds around the Double Thookle Billabong (also known as Hunchee Island South Billabong). The similar physical appearance of mounds HIS_2_22 and HIS_2_23 potentially indicated a shared age range. Consequently, the sample for the former was taken from level 8 to test for environmental continuity via pollen analysis of a deeper stratum. A control sample was not included due to limited funding. A comparison between mounds to test for differences in vegetation present in conjunction with dating was the key purpose, consequently a control sample of background floodplain sediment was not considered to be useful in this context. The analytical methods used are outlined below.
Pollen preparation involved a chemical extraction technique based on flotation as outlined in Cummings and Varney (2021) (see also Jones, 2023). A light microscope was used to count pollen at a magnification of 500×. Pollen preservation results in these samples varied from good to poor. An extensive comparative reference collection housed at the PalaeoResearch Institute assisted in pollen identification to the family, genus and species level where possible.
Pollen aggregates were recorded during pollen identification. Aggregates are clumps of a single type of pollen and may be interpreted to represent either pollen dispersal over short distances or the introduction of portions of the plant represented into an archaeological setting. The aggregates were included in the pollen counts as single grains, as per customary practice. An ‘A’ next to pollen frequency on the percentage pollen diagram notes the presence of aggregates.
The percentage pollen diagram was produced using Tilia software Version 2.1.1. Total pollen concentrations, expressed as pollen per cubic centimetre (cc) of sediment, were calculated in Tilia using the quantity of sample processed in cc, the quantity of exotics (spores) added to the sample, the quantity of exotics counted, and the total pollen counted.
‘Indeterminate’ pollen includes pollen grains that are folded, mutilated or otherwise distorted beyond recognition. These grains were included in the total pollen count since they are part of the pollen record. The microscopic charcoal frequency registers the relationship between pollen and charcoal. The total number of microscopic charcoal fragments was divided by the pollen sum, resulting in a charcoal frequency that reflects the quantity of microscopic charcoal fragments observed, normalized per 100 pollen grains.
The pollen extraction method aims to extract starch granules when they are present. Since starch analysis was requested for these samples and starches were recorded as part of the pollen count, an additional search for starches was conducted. Starch granules are a plant’s mechanism for storing carbohydrates and are found in numerous seeds, as well as in starchy roots and tubers. The primary categories of starches include the following: with or without visible hila, hilum centric or eccentric, hila patterns (dot, cracked, elongated) and shape of starch (angular, ellipse, circular or lenticular). Some of these starch categories are typical of specific plants, while others are more common and tend to occur in many.
Radiocarbon age determinations
The prior record of age determinations for mounds in the MDB used charcoal as the predominant material used for dating (n = 85), with both bone (n = 13) and shell (n = 1) considerably less common (Jones, 2023). In planning the initial dating program for earth mounds at Calperum it was decided to concentrate on mussel shell in the first instance, with follow up dating on selected mounds (HCN20 and HCN21) involving duplicate samples of shell and charcoal. The stratigraphic record for these mounds were complicated by the likely presence of shell lenses at levels 2–4 (Jones, 2023). Additional dating was undertaken to clarify the ages of operation of the mounds and investigate age of deposition of the shell middens, which were potentially established prior to the initiation of the mounds. Charcoal samples were included to assess the degree of mixing that occurred during mound to assist in unravelling the history of deposition in these locations.
The methods used for the samples analysed at ANSTO and Waikato are outlined in Hua et al. (2001). Pre-treatment for charcoal involved the acid-alkali-acid washing method using hot hydrochloric acid followed by sodium hydroxide then hydrochloric acid prior to combustion and graphitisation. The shell samples were cleaned by removal of the shell surface using a dental drill, after washing in de-ionised water the shell was crushed to small pieces and repeatedly subjected to HCl etches to remove 40–85% of the surface of the shell to remove secondary carbonates (see Kirillova et al., 2018). The shells were treated with phosphoric acid and the resulting carbon dioxide was graphitised as described by Hua et al. (2001). The AMS measurements were performed on the Vega 1MV accelerator (Wilcken et al., 2015). The 14C measurements were normalised using NBS Oxalic acid I as the primary standard (Mann, 1983). Stable carbon isotopic ratios were measured on the graphite targets using a Vario Microcube Elemental Analyser and an IsoPrime Isotope Ratio Mass Spectrometer (EA/IRMS), the δ13C was used to correct the measured 14C/13C values to determine the radiocarbon age. All results are reported as percent modern carbon (pMC) and as conventional radiocarbon ages (yr BP) following the conventions of Stuiver and Polach (1977).
Conventional Radiocarbon Ages (CRAs) were calibrated using the OxCal 4.4 program (Bronk-Ramsey, 2009) applying the SHCal20 atmospheric curve (Hogg et al., 2020). The calibrated age-ranges listed in Table 3 are reported at 95.4% probability. Gillespie et al. (2009) sampled freshwater mussel shell from the Murrumbidgee and Darling Rivers located upstream from Calperum Station and concluded that no appreciable reservoir effect would occur in MDB samples. Westell et al. (2020) noted that the intervening geology would limit the entry of older carbonate bearing material that would contaminate younger samples. This was due to the presence of non-carbonate fluvial/alluvial sediments and underlying quartzose sands of the Pliocene Loxton-Parilla Sands.
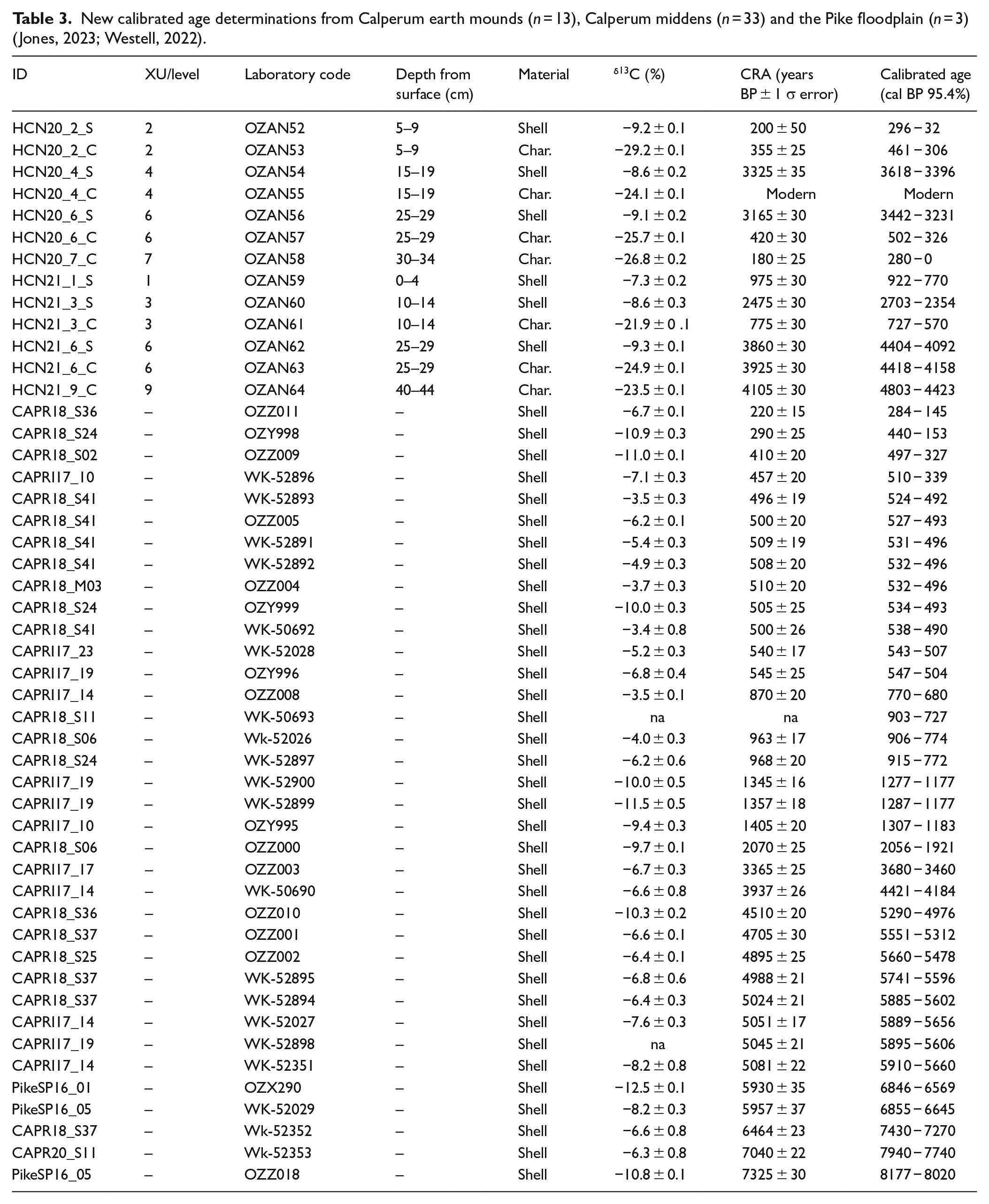
New calibrated age determinations from Calperum earth mounds (n = 13), Calperum middens (n = 33) and the Pike floodplain (n = 3) (Jones, 2023; Westell, 2022).
Results
Eighty one dates, including 53 midden dates (provided by Westell, 2022) and 28 earth mound dates (provided by Jones, 2023), were analysed at the Australian Nuclear Science and Technology Organisation (ANSTO) Centre for Accelerator Science at Lucas Heights and the University of Waikato Radiocarbon Dating Laboratory (Table 3; Supplemental Data).
Pollen and starch data
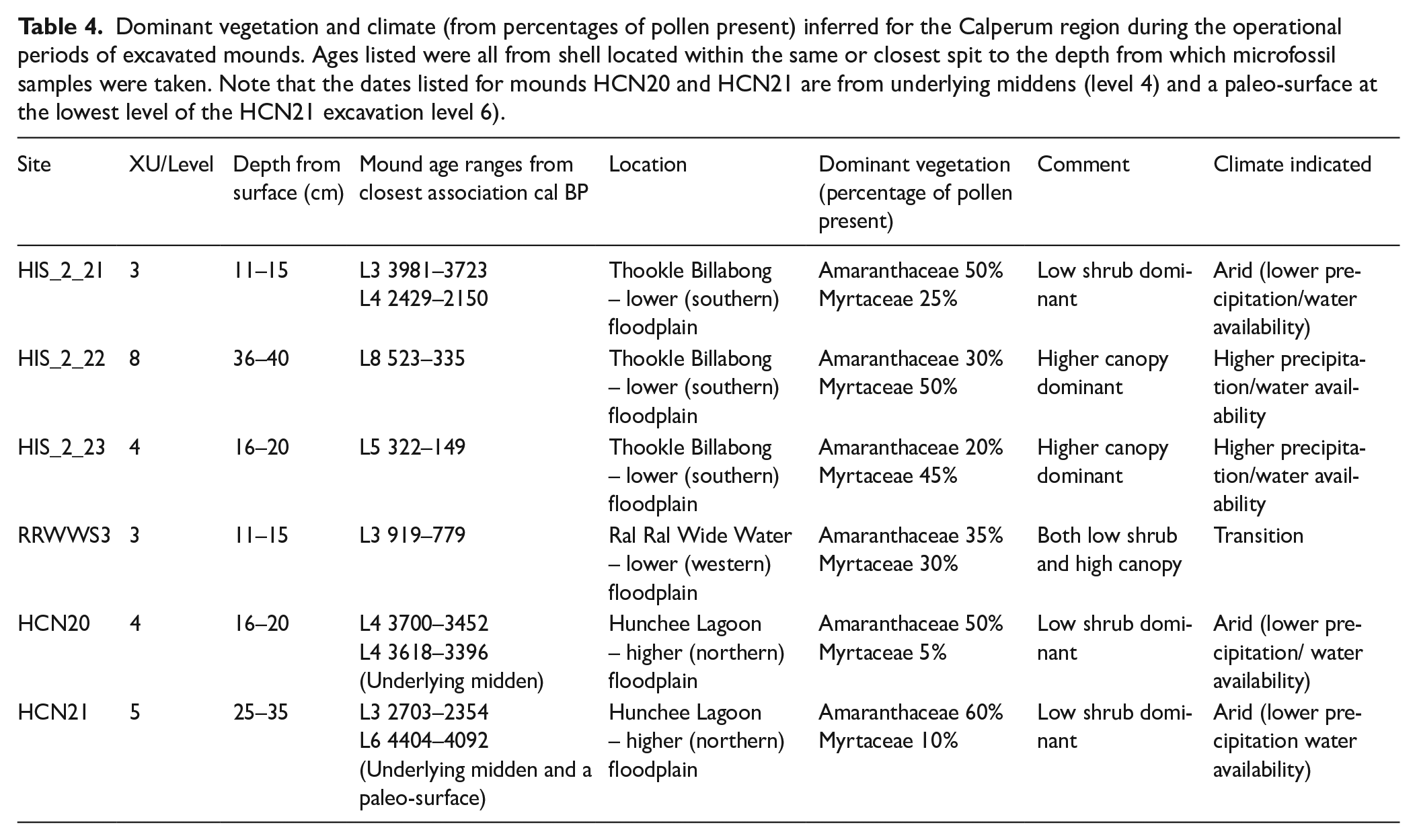
The majority of the pollen is derived from three taxa: Myrtaceae, Amaranthaceae and high-spine Asteraceae (Figure 6). Woody Myrtaceae have aromatic leaves containing oil glands and include Eucalyptus spp., which are certainly represented in this collection of pollen (Chippendale, 1988). No attempt was made to differentiate below the family level for Myrtaceae pollen, due to similarities in morphology and size and the high frequency of this family in the region. The results obtained from the pollen analysis, including the percentage of pollen present from the predominant taxon (Myrtaceae and Amaranthaceae, are presented in Table 4 and Figure 6. It is noted that the older sediments of HIS_2_21, HCN20 and HCN21 were dominated by Amaranthaceae pollen, in contrast to the dominance of Myrtaceae pollen in the younger sediments of mounds HIS_2_22, HIS_2_23 and RRWWS3. Larger quantities of Myrtaceae pollen suggest a more closed canopy, while increased Amaranthaceae pollen suggests more sunlight at ground level and the dominance of smaller species. The HCN20 sample also contained a moderately large quantity of high-spine Asteraceae pollen. Concentrations of microscopic charcoal are greater in the samples from mounds RRWWS3, HCN20 and HCN21 with smaller amounts in descending order from HIS_2_23, HIS_2_21 and HIS_2_22 .
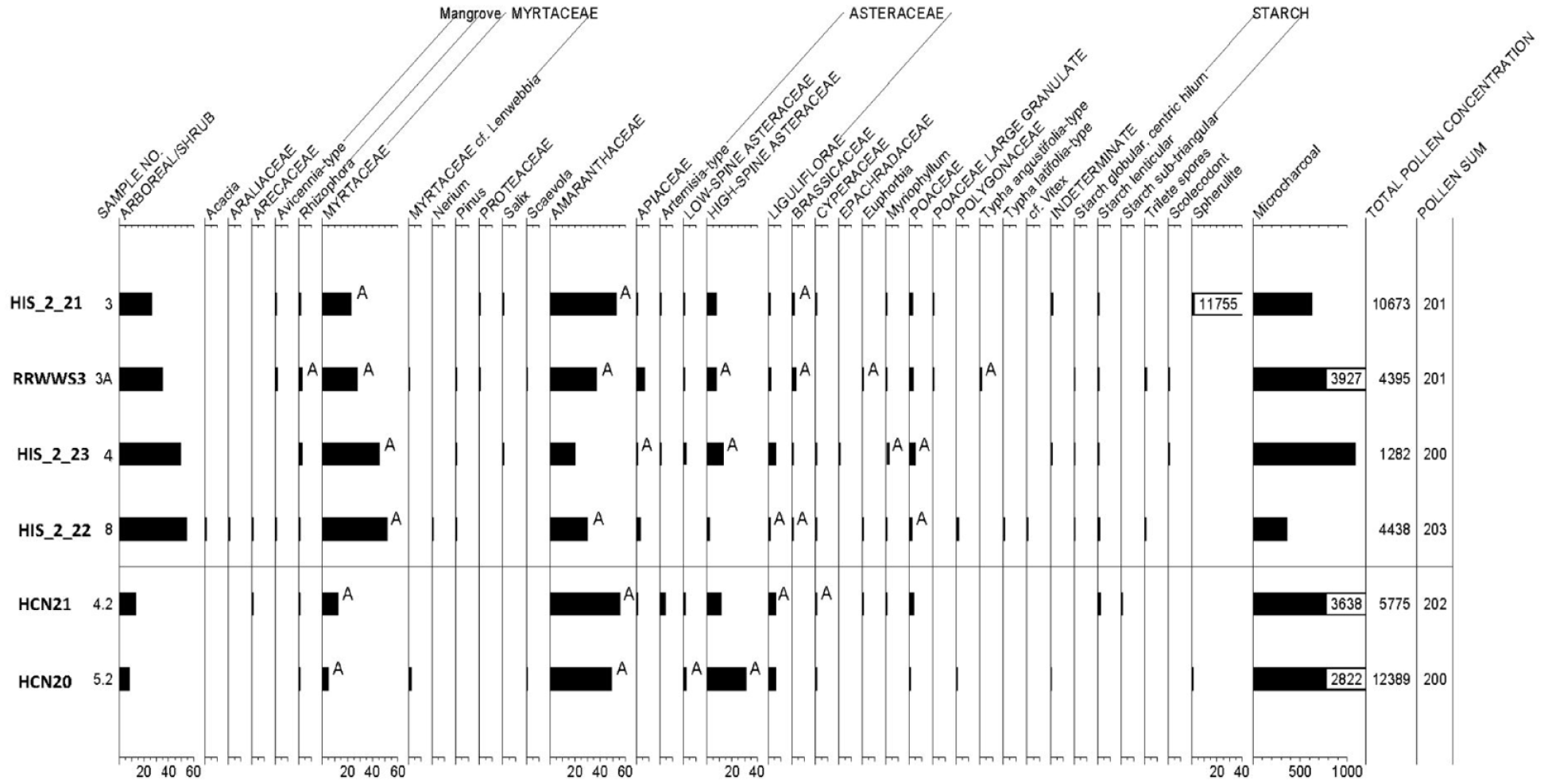
Pollen and starch study results from the six excavated mounds at Calperum. Vertical axis shows the stratigraphic level from which the sample was taken. The horizontal axis shows the percentages of pollen present for each mound sample. ‘A’ next to the pollen frequency on the percentage pollen diagram notes the presence of aggregates.
Dominant vegetation and climate (from percentages of pollen present) inferred for the Calperum region during the operational periods of excavated mounds. Ages listed were all from shell located within the same or closest spit to the depth from which microfossil samples were taken. Note that the dates listed for mounds HCN20 and HCN21 are from underlying middens (level 4) and a paleo-surface at the lowest level of the HCN21 excavation level 6).
The arboreal and shrubby portion of the pollen record includes small quantities of Acacia, Araliaceae, Arecaceae, Avicennia, Rhizophoraceae, Myrtaceae (probable Lenwebbia), Nerium, Proteaceae, Salix and Scaevola. Small amounts of pollen from the mangrove genera Rhizophora and Avicennia. The former were present in all mound samples and the latter in all except for mound HIS_2_23, thus possibly indicating long distance transport of pollen by wind from coastal habitats that contain these genera. Similarly, the presence of Pinus (pine) and Salix (willow) pollen suggests wind borne contamination since the introduction of Salix and Pinus species to SA after 1836. Mounds RRWWS3, HIS_2_22 and HIS_2_23 are characterised by a top layer of fine, ashy and more porous, sediments in contrast to the harder crust found on the Hunchee mounds, and a sealed crust on HIS_2_21 which would potentially inhibit the entry and downward migration of later arrivals such as Pinus, Salix and Nerium pollen. This is potentially complicated by bioturbation as indicated by the presence of Salix pollen at level 3 in mound HIS_2_21.
Non-arboreal pollen other than Amaranthaceae and high-spine Asteraceae (daisy/sunflower) are in small frequencies. Apiaceae, Artemisia, low-spine Asteraceae, Liguliflorae, Brassicaceae, Cyperaceae, Epachradaceae, Euphorbia, Myriophyllum, two types of Poaceae, Polygonaceae, two species of Typha (one that produces single pollen grains and the other that produces tetrads), and cf. Vitex represent members of the umbel family, sagebrush or a similar plant, ragweed or marshelder or a similar plant, a member of the chicory tribe of the sunflower family, mustards, sedges, a member of the Epachris family, spurge, water milfoil, grasses, a member of the knotweed family and vitex or chaste-tree. Wetland indicators include willow, some members of the umbel family, water milfoil, some grasses and Typha. Ferns are represented by small quantities of three morphotypes of spores with scalloped, reticulate and smooth surfaces. A small quantity of scolecodonts were observed in HIS_2_22 and HIS_2_23. HCN20 contained a moderately large quantity of high-spine Asteraceae pollen in contrast with all other mounds.
Spherulites were abundant in HIS_2_21, rare in HCN20, and absent in the remainder of the mounds. They are small, globular forms that display radial birefringent patterns. Although some forms of spherulites are observed in dung, they are not diagnostic for the presence of dung. However, the high count in HIS_2_21 likely reflects the extensive damage and bioturbation by rabbits and consequently high levels of rabbit dung in its sediments.
Starches were observed in five of the six samples examined (Cummings and Varney, 2021). All mound samples except for HCN20 yielded lenticular starches, which are typical of grasses with large seeds and also Marsilea drummondi (nardoo) and Crinum flaccidum (Murray lily) both of which are species that were used by Aboriginal peoples in the MDB as a source of carbohydrate (Aston, 1973: 37–39; Maiden, 1889). Starches that were spherical to sub-angular with a centric hilum were noted in the samples from RRWWS3, HIS_2_22 and HIS_2_23. Although this starch morphotype is typical of many grass seeds, it is also observed in other seeds and some tubers. The sediment from mound HCN21 provided a single small (20 µ) sub-triangular fragment of starch which was slightly angular with a nearly centric or slightly eccentric hilum. At present, this specimen remains unidentified as it was not large enough to adequately match with reference samples. A single centric starch, typical of grass seeds, was observed in sample HCN20.
As outlined above, the recovery and identification of ancient starch in Aboriginal earth oven mounds at Calperum was not significant probably because of the temperatures generated during cooking and the presence of high moisture levels potentially associated with the deliberate addition of water to the earth oven before closing (Eyre, 1845: 2) and/or a high water content in rootstocks derived from an aquatic environment and quickly cooked in mounds established in close proximity to such locations (see Takahashi et al., 1982 for a study of the relationship of moisture content to starch degradation under high temperature conditions).
Low survivability of ancient starch in Calperum mounds is consistent with work conducted at Fort Hood in Texas, by Laurence (2013) who found only 1 fragment of starch out of 27 was related to the past activities of Indigenous peoples. In addition to a lack of ancient starch in burnt rock mounds, Laurence (2013) also found that environmental contamination from modern starch could occur in both field and laboratory contexts. The results of this study, combined with the generally poor preservation of ancient starchy residues and the strong possibility of modern starch contamination from agricultural and natural sources in Australian Aboriginal earth mound environments suggests that there may be little value in pursuing future starch studies to identify plants cooked in earth oven mounds (see also Martin, 2006, 2011).
Radiocarbon dates
The initial radiocarbon dating programs in 2019 and 2020 for earth mounds at Calperum included 13 ages (11 from mussel shell and 2 from charcoal) from the four lower floodplain mounds HIS_2_21, HIS_2_22, HIS_2_23 and RRWWS3 (Jones et al., 2022). Two ages (one from shell and one from charcoal) were obtained later from the higher floodplain mounds HCN20 (WK52024) and HCN21 (WK52025) and were tested at the University of Waikato Radiocarbon Dating Laboratory. A further 13 radiocarbon dates were obtained in early 2021 for mounds HCN20 and HCN21 (Table 3) in order to investigate the provenance of charcoal and the mussel shell lenses found within these mounds (Jones, 2023).
Dates for the lower floodplain range from mound HIS_2_23 at 283–0 cal BP (OZZ572) (obtained from shell) through to mound HIS_2_21 at 3981–3723 cal BP (OZZ566) (also obtained from shell). Dates from the upper floodplain mounds HCN20 and HCN21 range from 4803–4423 (OZAN64) to 280–28 cal BP (OZAN58), both of which were obtained from charcoal (Jones, 2023; Jones et al., 2022). Relevant earth mound radiocarbon dates are presented in association with pollen results in Table 4.
Westell (2022) provided shell dates (n = 53) obtained from shell middens at Calperum and the nearby Pike floodplain that are presented with the available earth mound dates showing a shell age distribution between 5910–5660 (WK-52351) and 284–145 (OZZ011) (Figure 7). Figure 8 details Calperum midden and mound shell age plots (from ~8000 cal BP) in comparison with data from a lake level study in south-western Victorian lakes (Wilkins et al., 2013) and an ENSO activity model (Moy et al., 2002) over the period from 8500 cal BP. A comparison of climate-linked data with shell radiocarbon data was intended to investigate the potential of fresh-water mussel shell availability as a proxy for environmental change at Calperum. For example, the Wilkins et al. (2013) plot suggests increasing aridity in the south-eastern region of the Australian continent after 5000 cal BP and fluctuations in the availability of water in the MDB from about 3800 cal BP (possibly linked to variation in ENSO activity) that persisted until after 2500 cal BP (Moy et al., 2002).
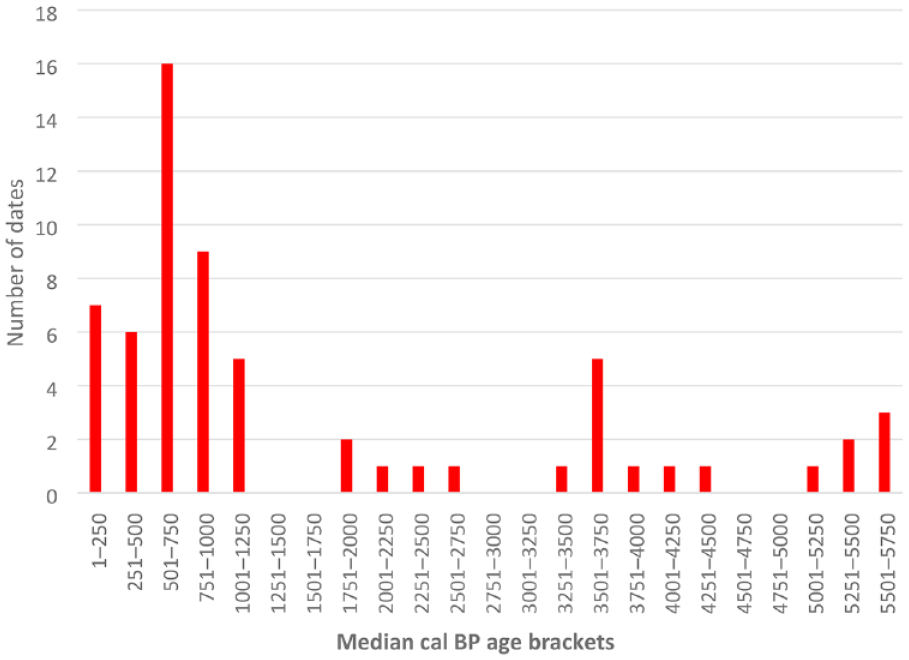
Combined plot of median shell ages from midden and mound sites on the Calperum floodplain from ~5800 cal BP (calculated as a median of the calibrated range) indicating the number of dates per 249 year period (Refer to Table 3 and Supplemental Data; see also Jones, 2023; Jones et al., 2022; Westell, 2022; Westell et al., 2020).
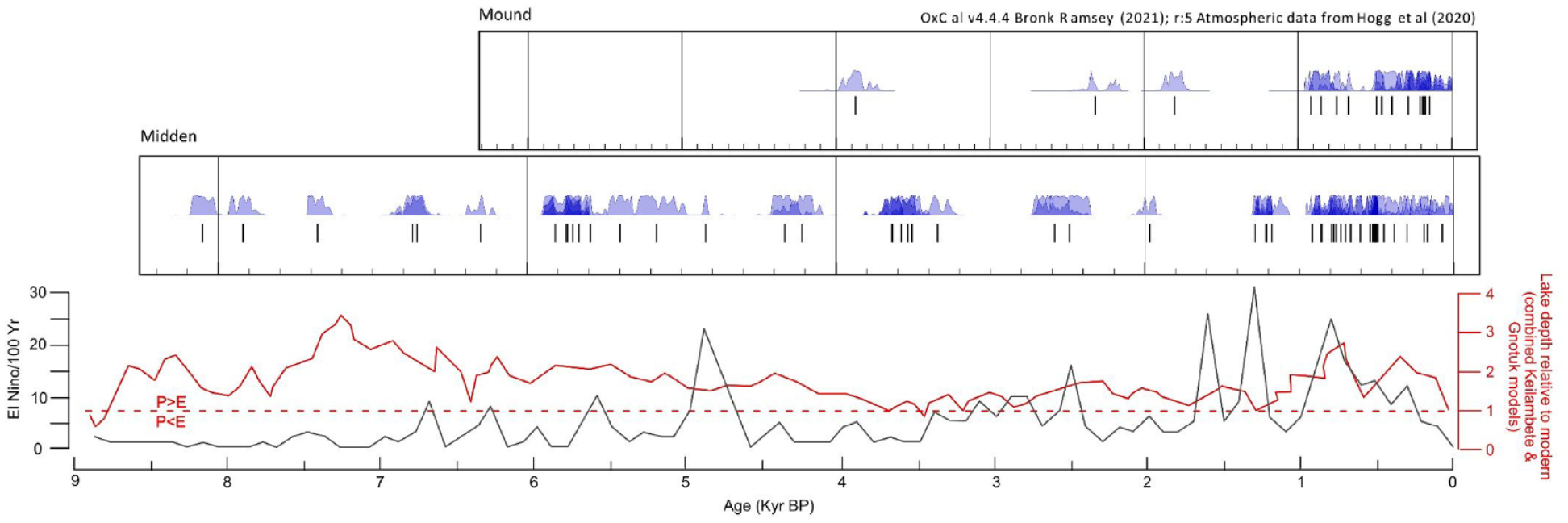
Comparison of shell age ranges from Calperum midden and mound sites from about 8000 cal BP. Vertical lines represent the median for each date-range. The three mound dates shown in the top graph prior to about 1800 cal BP all refer to mound HIS_2_21 (see also Table 3) (Jones, 2023; Jones et al., 2022; Westell, 2022; Westell et al., 2020). The two plots below the shell dates represent a lake-level study in south-western Victoria in Lakes Keilambete and Gnotuk (Figure 4) from Wilkins et al. (2013) and the Laguna Pallcacocha ENSO activity model by Moy et al. (2002) presented on the same chronology. Note that P > E/P < E indicates precipitation greater than/lesser than evaporation and additional dates prior to 5910–5660 cal BP from Calperum (n = 2) and the nearby Pike floodplain (n = 4) have been added to provide continuity with the chronology of the climate plots presented (Westell, 2022; also refer to Supplemental Data).
Note that dates obtained from shell lenses from the HCN20 and HCN21 excavations have been included in the midden data set since these are chronologically unrelated to the formation of these earth mounds.
Discussion
Evidence of climate variation at Calperum
The data from the Tareena Billabong (located at the eastern edge of the Chowilla floodplain – Figure 4) paleolimnological study by Gell et al. (2005: 449) indicated its formation at about 5000 cal BP, during a period of relatively high rainfall and isolation from the main river channel (Figure 8). A climate transition to ENSO driven variable conditions in the Riverland region became active over the period ~3800–2500 cal BP (Gell et al., 2005; see also Bowler and Hamada, 1971; Fitzsimmons and Barrows, 2010; Fitzsimmons et al., 2013; Gingele et al., 2004, 2007; Moy et al. 2002; Petherick et al., 2013; Shulmeister and Lees, 1995; Wilkins et al. 2013). The billabong was reconnected to the main river after 3800 cal BP with the likely intensification of ENSO related wet-dry cycles, episodic aridity and phases of elevated salinity (Gell et al., 2005). Improved conditions and higher water availability developed after 2500 cal BP, indicated by low levels of Chenopodiaceae pollen from ~2500 cal BP (Gell et al., 2005). The pollen results obtained from mounds at Calperum (Figure 6 and Table 4), while lower in resolution, similarly show lower water availability from 3800 cal BP and a later transition in the vegetation present (Amaranthaceae to Myrtaceae), which indicates the onset of more favourable conditions at some time prior to ~1000 cal BP (Figure 8 and Table 4).
Reduced anabranch flows, wet-dry cycling and fluctuations in salinity levels and increased aridity in the Riverland region, likely caused variable water conditions in wetland microenvironments located on wide floodplains, such as at Calperum, from ~3800 cal BP. Given episodic wet-dry conditions, variability in surface water flows and fluctuations in salinity levels (including in anabranch creeks, swamps, billabongs and lagoons), previously productive environments would likely suffer episodic decreases in subsistence resources, including terrestrial animals and some plants, as well as fish, crustaceans, waterbirds, shellfish and some aquatic plants.
Mussel shell dates from middens and earth mound contexts on the Calperum floodplain (Figure 7); Supplemental Data, (if considered as a proxy for mussel availability and consumption by Aboriginal peoples) tentatively suggest fluctuations in floodplain food resources during the period 3800–1300 cal BP. In particular, data obtained from freshwater mussel shell middens at Calperum (Figure 8) potentially suggest regular variation in the availability of mussels in floodplain environments between ~8000 and 1300 cal BP. V. ambiguus is the most likely species found in floodplain environments at Calperum during periods of favourable water conditions (Jones, 2011; Walker, 1981; Westell, 2022). With the onset of drying and saline conditions both the riverine and lacustrine species of freshwater mussel found in the region would be adversely affected (Walker, 2017; Walker et al., 2001). Furthermore, the nature of the life cycle of both V. ambiguus and the river mussel A. jacksonii indicates that numbers of these invertebrates would be seriously disrupted by repeated drying and low water conditions that impact fish stocks, since an early life cycle stage relies on the parasitisation of fish as a host (Walker, 1981, 2017). A report by Sheldon et al. (2020) on the impact of dry conditions on mussel mortality during 2017–2019 in the northern MDB indicated mortality rates of up to 100 per cent during a 2-year drying event. Sheldon et al. (2020) indicated that recovery of mussel numbers in this instance was dependent on the survival of small populations in permanent water holes and ‘on the recovery and movement of native fish populations’ that require the return of sufficient river flow to allow successful spawning. An extended period of fluctuations in river flows over centuries, as potentially occurred after ~3800 cal BP, would have major adverse consequences for mussel habitats located in anabranch, lagoon and billabong environments.
The loss or a reduction in access to aquatic resources and an extended delay in habitat recovery from ~3800 to 2500 and possibly to 1300 cal BP, potentially provided the impetus for a transition in subsistence procurement at Calperum, as indicated by the likely establishment of mound HIS_2_21 at ~3800 cal BP (Figure 8). The likely beginning of a transition in ecological conditions involving low water levels and fluctuations in salinity levels, suggested above for floodplain environments at Calperum from ~3800 cal BP, would have progressively impacted mussel availability. This would have occurred at salinity levels above 3 g L–1 (3000 ppm) (Walker et al., 2001). The growth of Typha would also be impacted but not as severely as mussel growth in the first instance since Typha species are moderately salt tolerant (Beare and Zedler, 1987; Hocking, 1981; McMillan, 1959; Whigham et al., 1989; Zedler et al., 1990). The growth of Typha is reported to be impacted at salinities above 3000–5000 ppm but mortality occurs in the range 10,000–25,000 ppm (Glenn et al., 1995). These salinity sensitivities suggest that mussel survival would be affected prior to that of Typha. Such a significant difference in susceptibility to salinity levels supports the hypothesis that a change in subsistence strategy occurred ~3800 cal BP as an adaptation to deteriorating conditions within the anabranch-fed water features on the floodplain.
The pattern of dates reported in this paper as well as Jones (2023), Jones et al. (2022), Westell (2022) and Westell et al. (2020) (Figures 7 and 8; Supplemental Data) possibly indicate climatically induced fluctuations, and extended periods of disruption, in the supply of mussels (and other resources) in floodplain environments at Calperum. From the evidence discussed above this likely developed from ~3800 cal BP and was active for about 1300 years between ~3800 and 2500 cal BP, as also indicated by the Gell et al. (2005) study, and diminished the availability of mussels in anabranch, lake and billabong environments as a readily available staple food. The anabranch system is the main surface source of water for lakes and billabongs (and consequently floodplain mussel and other animal and plant habitats) on the Calperum floodplain. The period 3500–2251 cal BP demonstrates three shell ages from the total obtained with a gap of about 800 years (Figure 7). These samples were derived from earth mound contexts (from mound HIS_2_21 located close to the main river channel and two from each of the shell lenses (middens) located under mounds HCN20 and HCN21 at Hunchee Lagoon). The age range of 2703–2354 cal BP (OZAN60) for a shell sample from a lens located at level 3 of mound HCN21 suggests that a potential amelioration of climate, a recovery of mussel habitat and a resumption of mussel predation occurred in the Hunchee Lagoon area after ~2500 cal BP.
A potential scenario
Based on the evidence presented, mussel availability and consumption likely fluctuated during the period ~3800 to ~1200 cal BP at Calperum, becoming increasingly concentrated on the banks of the main river channel where freshwater flows continued, albeit adversely influenced by increased variability and diminished discharge upstream (Bowler and Hamada, 1971; Fitzsimmons and Barrows, 2010; Fitzsimmons et al., 2013; Gell et al., 2005; Gingele et al., 2004, 2007; Petherick et al., 2013; Shulmeister and Lees, 1995). The presence of shell, in the lower floodplain mound HIS_2_21, dating to 3981–3723 (OZZ566), 2429–2150 (OZZ567) and 1864–1729 (OZZ568) cal BP, indicates the continuing availability of mussels in the southern part of Hunchee Island near the main river channel, as well as the initiation and continuity of mound use in this location.
Under the scenario outlined above, a decline in water availability across the anabranch fed system of lakes, swamps and billabongs at Calperum would have led to a decline in both terrestrial and aquatic food resources across the wider floodplain. Aboriginal groups at Calperum would have likely contracted to areas close to the main river channel. Continuing adverse climatic conditions and pressure on declining resources provided the impetus to adopt a food production system by taking advantage of a natural ecological niche involving Typha and a partial reorganisation of labour from traditional foraging activity to intensive activities such as the harvesting of large quantities of roots and the operation of earth ovens, essentially involving a production system rather than collecting. This opportunity likely related to the proximity of the main river channel and a continuity of conditions suitable for the survival of Typha in the nearby Thookle Billabong (Figure 9). This finding aligns with our hypothesis relating early Typha exploitation to the onset of the ENSO weather patterns identified by Gell et al. (2005), a continuing presence of mussels and other aquatic resources and the availability of Typha in the vicinity. The paucity of dates from mussel shell between ~3300 and ~2500 cal BP suggests declining returns from floodplain environments reliant on regular anabranch recharging. This induced a redirection of group and clan resources to the seasonal exploitation of Typha close to the main river channel. The return of wetter and less variable conditions from ~2500 cal BP indicated by palaeolimnological evidence (Gell et al., 2005), the new pollen evidence provided in this paper (Figure 6 and Table 4) and the age range of ~2703–2354 cal BP (OZAN60) obtained for a shell sample from a lens (midden) level 3 of mound HCN21, potentially indicates the return of conditions suitable for the expansive growth of both emergent macrophytes and prey habitat recovery across the floodplain at that time (although continuing fluctuations in water and food resource availability are potentially indicated by the lack of mussel dates during the period between ~1750 and 1300 cal BP) (see Figure 8).

The locations of mounds HIS_2_21, HIS_2_22 and HIS_2_23 during the flooding event of 2022/2023. The main river channel is located immediately to the south.
Under this scenario, after the establishment of the new production system, Aboriginal people on the Calperum floodplain may have progressively targeted new stands of Typha leading to the abundance of mound locations now demonstrated at Calperum. The increase in midden and mounds after ~1000 cal BP indicated by the dating evidence (Figures 7 and 8) suggest the higher availability of water in the environment and a natural increase in the population at Calperum after this time. Alternatively, the higher presence of younger sites in an active floodplain environment may reflect taphonomic factors.
Conclusions
The research has provided new information to consider key debates about the development of Aboriginal societies in the Holocene within the MDB. In particular, via a study of earth mounds on the Calperum floodplain this research has considered the influence of mid-Holocene environmental change on Aboriginal societies, particularly in relation to diets.
It is argued in this paper that the transition in subsistence procurement that took place on the Calperum floodplain was a consequence of long-term fluctuations in the supply of traditional food resources which precipitated a response that involved the establishment of a new food production system. It is proposed here that this may have been enabled through the integration of a new or under-utilised seasonally abundant resource (likely Typha spp.), a logistical change of scale in the use of an existing technology (heat retainer cookery) and active resource management through the use of fire, digging and harvesting (see Gott 1999). The repurposing (and rescaling) of heat retainer technology was the key innovation which may have culminated in the seasonal exploitation of emergent macrophytes on the Calperum floodplain from ~3800 cal BP.
Given the early age of earth mound HIS_2_21 on the lower floodplain at 3981–3723 cal BP (OZZ566) and the chronological data obtained at Calperum, it is highly probable that a logistic increase in the use of heat retainer technology (potentially for the cooking of Typha roots) was in use in the region by this time. Consequently, a transition in subsistence strategy on the Calperum floodplain is indicated after ~3800 cal BP. The establishment of mound HIS_2_21 adjacent to a billabong located ~300 m from the main river channel (Figure 9) at ~3981–3723 cal BP closely correlates to climate change at this time (Gell et al., 2005; Jones 2023). The early age range of 4404–4092 cal BP (OZAN62) for shell in level 6 of the HCN21 excavation is ~400 years earlier than the establishment of mound HIS_2_21, which suggests prevailing environmental conditions at the time supported mussel harvesting and consequently available floodplain resources in general (Figures 7 and 8). As conditions fluctuated Aboriginal people likely responded to ecological change and the consequent reduction in resources in billabong, lagoon and anabranch locations due to low water availability and salinity variations in water features across the floodplain.
The initiation of mound HIS_2_21 at ~3800 cal BP by Aboriginal people likely followed a reduction in floodplain productivity and is evidence of a change in focus to areas adjacent to the main river channel where resources were still available. This would have led to a higher population density in the immediate vicinity of the river and greater pressure on depleted resources. The continuation of wet-dry cycles over an extended period, leading to long-term decreases in resource productivity, provided the impetus for innovation and the implementation of the food production system outlined above.
Under this hypothesis, improvements over time in local ecological conditions would lead to the progressive establishment of mounds in different locations in the floodplain, demonstrating the persistence and value of this production system. In effect, Aboriginal people at Calperum broadened their diet via the addition of a new seasonal food production strategy to their subsistence repertoire, demonstrating cultural, technical and socio-economic innovation. The strong correlation to adverse climate change strongly suggests that behavioural change at Calperum was a function of external environmental challenge and adaptation through innovation and socio-cultural reorganisation at subregional level from ~3800 cal BP. The dating evidence indicates that other regions in the MDB (Figure 5) potentially experienced different conditions and influences that were consequently expressed in different temporal periods (Jones, 2023). As previously noted, this finding reinforces Ulm’s (2006: 256, 2013) point about the importance of regional studies to identify both similarities and differences in the development of mid-to Late-Holocene Aboriginal societies when investigating questions of intensification and changes in diet breadth. However, further research into the ages of mussel shell middens and shell from earth mounds in this region is required to provide a finer chronoloigcal resolution in order to clarify these associations. In addition, further research is required for direct evidence of the primary use of Typha within earth mounds at Calperum as suggested by early historical accounts and other research indicating Typha exploitation in other regions of the MDB. Currently, the only direct evidence of early Typha use in the MDB is the presence of leaf imprints in burnt clay from mound excavations in the western region of the Hay Plain (Martin, 2006). Advances in the detection and analysis of environmental DNA from plant material may have relevance for further investigations.
Supplemental Material
sj-docx-1-hol-10.1177_09596836241266397 – Supplemental material for Aboriginal earth mounds and ENSO on the Calperum floodplain, Murray-Darling Basin, South Australia
Supplemental material, sj-docx-1-hol-10.1177_09596836241266397 for Aboriginal earth mounds and ENSO on the Calperum floodplain, Murray-Darling Basin, South Australia by Robert Jones, Amy Roberts, Craig Westell, Ian Moffat, Geraldine Jacobsen and Linda Scott Cummings in The Holocene
Footnotes
Acknowledgements
The authors sincerely thank all of the fieldwork volunteers and the RMMAC members who participated in the fieldwork for this project. The authors also thank the RMMAC Directors who have supported this work and the staff at Aboriginal Affairs and Reconciliation, Department of State Development, Government of South Australia, for their assistance with permits. The authors thank the Australian Landscape Trust for their ongoing support of this project.
Abbreviations
Accelerator Mass Spectrometry (AMS), Australian Nuclear Science and Technology Organisation (ANSTO), Broad spectrum diets (BSD), El Niño Southern Oscillation (ENSO), Last Glacial Maximum (LGM), Mid-Holocene Climate Optimum (MHCO), Murray-Darling Basin (MDB), River Murray and Mallee Aboriginal Corporation (RMMAC), South Australia (SA).
Author contributions
Robert Jones: Conceptualization; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Writing – original draft; Writing – review & editing.Amy Roberts: Funding acquisition; Methodology; Project administration; Resources; Supervision; Validation; Writing – review & editing.Craig Westell: Conceptualization; Data curation; Funding acquisition; Investigation; Writing – review & editing.Ian Moffat: Methodology; Supervision; Validation; Writing – review & editing.Geraldine Jacobsen: Data curation; Formal analysis; Methodology; Validation; Writing – review & editing.Linda Scott Cummings: Data curation; Formal analysis; Methodology; Writing – review & editing.and River Murray and Mallee Aboriginal Corporation: Supervision; Validation
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding was provided by an Australian Research Council Linkage Project (LP170100479) led by Professor Amy Roberts. Associate Professor Ian Moffat was the recipient of Australian Research Council DECRA Fellowship (DE160100703), Future Fellowship (FT220100184) and a Flinders University Research Investment Fund Grant. Radiocarbon dating at ANSTO was funded by ANSTO research grants AP12533 and AP12957. The authors acknowledge the financial support from the Australian Government for the Centre for Accelerator Science at ANSTO through the National Collaborative Research Infrastructure Strategy (NCRIS). Additional funding was provided to Robert Jones by the Australian Archaeological Association Student Research Grant Scheme and student grants from Flinders University and the Royal Society of South Australia. Craig Westell received financial support for this research from the Australian National Collaborative Research Infrastructure Strategy (NCRIS) (funding for radiocarbon dating conducted at the ANSTO Lucas Heights facility), and the National Landcare Programme (administered by the SAMDB NRM Board). Additional financial support has been provided by Vivienne Wood Heritage Consultant Pty Ltd, the Flinders University College of Humanities, Arts and Social Sciences Research Assistance Grant and the Australian Archaeological Association Student Research Grant scheme.
Ethical approval
This paper is based on PhD research conducted by both Robert Jones and Craig Westell in collaboration with the River Murray and Mallee Aboriginal Corporation (RMMAC) and approved by Flinders University’s Social and Behavioural Research Ethics Committee (Project Number: 6618).
Supplemental material
Supplemental material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.