
Abstract
Although rare, temporally and taxonomically highly-resolved palaeoecological studies with high chronological precision are essential to perform detailed comparisons with precisely dated independent evidence such as archaeological findings, historical events, or palaeoclimatic data. Using a new highly-resolved and chronologically precise sedimentary record from Lago di Mezzano (central Italy), we reconstruct decadal-scale vegetation, species diversity, and fire dynamics, aiming to better understand the linkages between climate, land use, fire, and plant communities from the Neolithic to the Copper Age (c. 5100–3100 cal. BC). Closed, mixed beech-oak forests, including evergreen Quercus ilex, dominated the landscape around Lago di Mezzano during the Neolithic and were disturbed by repeated opening phases, with important implications for lake biogeochemistry and mixing regimes. This was in conjunction with increasing fire activity to promote agro-pastoral practices, as inferred from increasing charcoal, Cerealia type, Triticum type, Hordeum type, Plantago lanceolata type, and Urtica pollen. Fires, on their turn, augmented species diversity (richness and evenness). The comparison of the Mediterranean record from Lago di Mezzano with available continuous and high-precision submediterranean and cool-temperate palynological sequences suggests comparable land use pulses across Southern and Central European regions, most likely in connection with climate change. The outcomes of this study are not only of palaeoecological and archaeological interest; they may also help to improve projections of ecosystem dynamics under future global change.
Introduction
Ongoing global warming and alterations in land use are expected to induce major ecological shifts, involving severe consequences for biodiversity and the composition of plant communities (Newbold et al., 2020; Scheffers et al., 2016). Mediterranean vegetation has been influenced by climate change and humans since prehistoric times (e.g. Carroll et al., 2012; Glais et al., 2017; Lowe et al., 1996; Revelles et al., 2015; Sadori et al., 2004; Tinner et al., 2009). However, because of the long timescales underlying successional patterns after natural and/or anthropogenic disturbances, understanding the current composition of vegetation and predicting future dynamics can be challenging when relying only on short-term observations (Willis and Birks, 2006). An alternative approach is given by palaeoecological investigations of past relationships between vegetation, fire, climate, and land use, which may reveal unique long-term patterns of resilience, vulnerability, and adjustment capacity of plant communities and species diversity. Hence, palaeoecological insights are relevant for biodiversity conservation, ecosystem restoration, and forest management (Gillson et al., 2022; Nieto-Lugilde et al., 2021; Valsecchi et al., 2010; Whitlock et al., 2018).
Late Pleistocene and Holocene environmental, vegetation and fire histories have been extensively studied on lake sediments of Lago di Mezzano in central Italy (Brandt et al., 1999; Giraudi, 2004; Ramrath et al., 1999a, 1999b, 2000; Sadori, 2018; Sadori et al., 2004; Wilkes et al., 1999). These studies revealed the general course of vegetation history, showing that both, climate and human impact determined important changes in vegetation composition and structure over millennia (Sadori, 2018). Particular emphasis has been devoted to the Bronze Age (2550−950 cal. BC; Sadori et al., 2004) after the finding of a submerged settlement (Franco, 1982), but uninterrupted palynological time series, suited to address short-lived disturbance impacts on vegetation (Lang et al., 2023), are still missing at the site. The lake has also attracted scientific interest because of the long duration of sediment accumulation (c. 34,000 years) and because some sediment sections are annually laminated (Ramrath et al., 1999a, 1999b, 2000), which improved the chronological basis of the available studies (Brandt et al., 1999; Sadori, 2018; Sadori et al., 2004; Wilkes et al., 1999).
It has been hypothesized that climate-human interactions generated synchronous land use phases, which altered vegetation structure and composition in Southern and Central Europe since the Neolithic (Rey et al., 2019a). Unfortunately, continuously sampled, high-resolution and precisely dated studies to address this hypothesis are generally rare in the Mediterranean realm and elsewhere in Europe. Nevertheless, some of them (Bisculm et al., 2012; Colombaroli et al., 2008; Lotter, 1999; Pedrotta et al., 2021; Rey et al., 2019a; Rösch et al., 2014; Tinner et al., 1999) could show the long-term effect of land use and fire disturbances on Neolithic vegetation successional patterns, while the existence, extent, and cause of synchronous land use phases remain understudied.
Our study has four main goals. The first goal is to establish a chronology as precise as possible for the study period of interest (c. 5100–3100 cal. BC). The second is to obtain the first uninterrupted palynological time series at Lago di Mezzano to reconstruct decadal-scale vegetation, diversity, and fire-history dynamics spanning from the Neolithic to the Copper Age at a Mediterranean site. The third is to investigate the long-term effect of land use and fire disturbances on Mediterranean plant communities’ composition, structure, and biodiversity. The fourth goal is to match available temperate and submediterranean continuous palynological records c. 5100–3100 cal. BC to test whether the temporal patterns in land use phases and forest recovery are comparable across a wider latitudinal gradient in Southern and Central Europe.
Material and methods
Study site
Lago di Mezzano (42°36′42″N, 11°46′12″E, 452 m a.s.l.) is a mesotrophic to oligotrophic (Ramrath et al., 2000) maar lake located in central Italy, in the northern part of Latium about 30 km from the Tyrrhenian Sea (Figure 1). The volcanic lake formed about 100,000 years ago (Metzeltin and Vezzoli, 1983) and is located in the Caldera of Latera, which is part of the Vulsini Volcanic Complex. The lake’s surface area is 0.5 km2, the maximal water depth reaches 31 m, and the catchment area comprises about 1 km2. A few small inlets, together with rainfall and groundwater, feed the lake. An artificial outlet located on the flat area on the northeastern side, the Fosso delle Volpi, flows into the Olpeta River, which then continues into the Fiora River. Nowadays, this artificial outlet limits seasonal lake level fluctuations to c. 0.5 m. Prior to the land reclamation works of the last century, the long-term evolution of the lake level was characterized by periods when the water level was ±10 m compared to today (Giraudi, 2004). Over the past millennia, indeed, the lake has changed from endorheic to exorheic several times, also altering its shape and size (Giraudi, 2004; Sadori et al., 2004). A temperate climate with relatively cold winters and warm summers characterizes the region around Lago di Mezzano. At the nearby weather station of Valentano (530 m a.s.l.; c. 7 km from the lake), the mean annual temperature is 13.9°C, with mean winter temperatures (December, January, February) of 6.5°C and mean summer temperatures (June, July, August) of 22.3°C. Annual precipitation is c. 1000 mm, with maxima during autumn and winter.
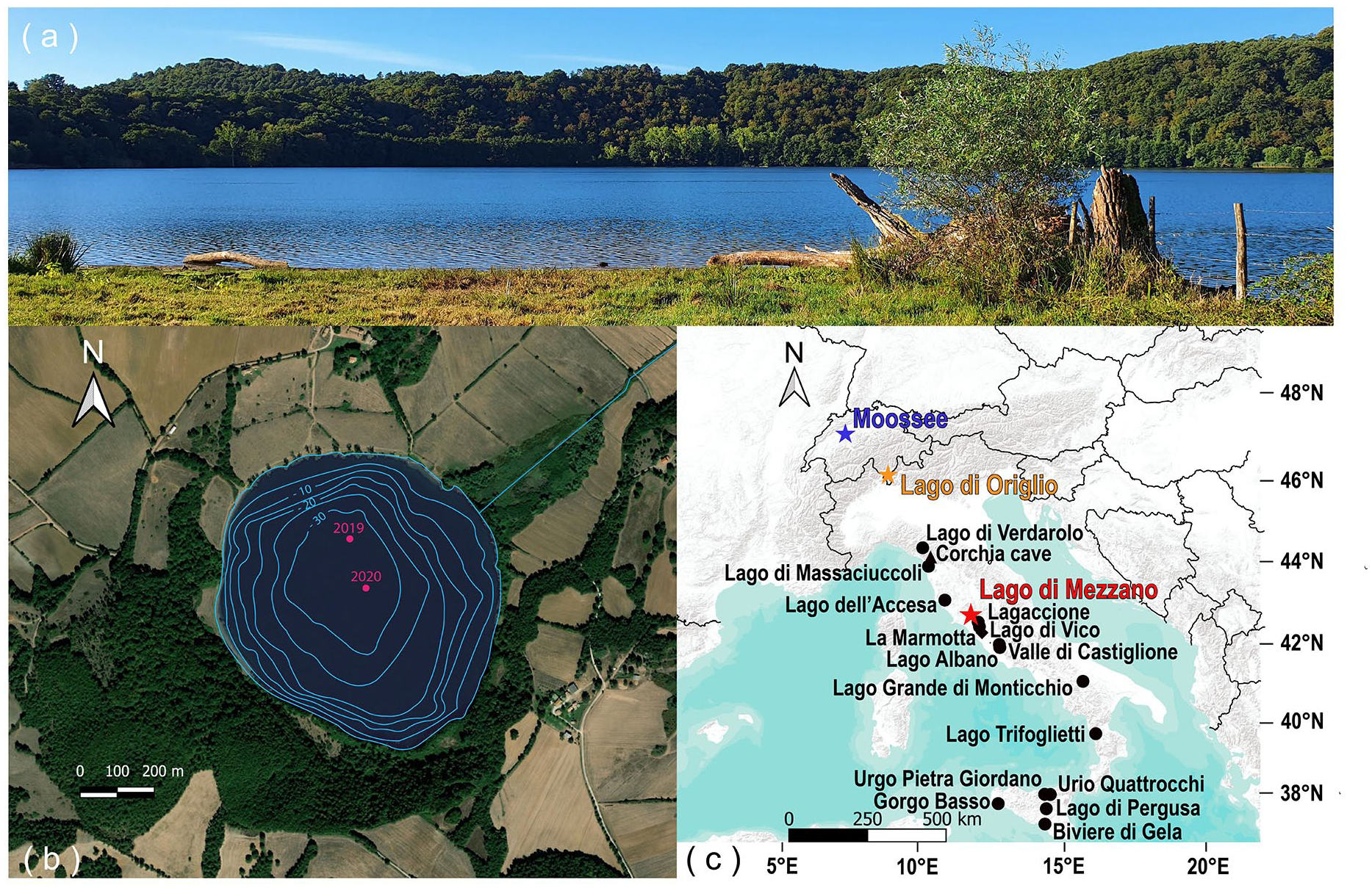
(a) Photograph of Lago di Mezzano (view from the north-east shore, photo Kathrin Ganz), (b) satellite photo of Lago di Mezzano with coring sites (in pink; for this study only 2019 cores are used) and bathymetric map (Ramrath et al., 1999b), and (c) topographic map showing the location of Lago di Mezzano, Lago di Origlio (Tinner et al., 1999), Moossee (Rey et al., 2019a), and other discussed study sites.
A hilly landscape in the volcanic crater surrounds Lago di Mezzano with open lands for cereal cultivation, pastures, and woodlands. On the steep west-southwest slopes in the caldera, mixed deciduous oak forests are dominated by Quercus cerris. Other tree species such as Ostrya carpinifolia, Quercus pubescens, Carpinus betulus, Castanea sativa, Fraxinus ornus, Acer opalus, and Prunus avium, together with shrubs such as Cornus mas, Corylus avellana, Ruscus aculeatus, Acer campestre, Rosa canina, Prunus spinosa, and Ligustrum vulgare, as well as the evergreen liana Hedera helix are also present. Single trees of Fagus sylvatica can be found on the north-facing slopes near the lake, which is significantly below the elevation range of the Apennine beech forests (1000–2000 m a.s.l.; Buonincontri et al., 2023; Pignatti et al., 2017), whereas single specimens of Quercus ilex are present on the south-facing slopes. Riparian vegetation (i.e. Alnus glutinosa, Populus alba, and Salix sp.) populates the lake shores.
Coring and chronology
In May 2019, four parallel sediment cores (MZZ A-D) were retrieved from the deepest part of the lake (31 m water depth, 42°36′47.0″N, 11°46′10.0″; Figure 1b) using an UWITEC piston corer with a diameter of 6 and 9 cm. Four additional cores (MZZ E-H) were collected in September 2020 from the center of the lake (30 m water depth, 42°36′42.8″N 11°46′12.0″E). We first visually correlated the cores based on lithological features. We then refined the correlation using X-ray fluorescence (XRF) and Hyperspectral imaging (HSI) data obtaining a composite length of 1191 cm. In this study, we focus on the section between 687 and 599 cm from the cores MZZ B and MZZ C, which covers part of the Neolithic (i.e. c. 5100–3100 cal. BC) and is composed of laminated sediments (gyttja, carbonaceous silts). In this section, 27 Accelerator Mass Spectrometry (AMS) radiocarbon ages were obtained on terrestrial plant macrofossils (Supplemental Table 1) in the Laboratory for the Analysis of Radiocarbon with AMS (LARA) at the University of Bern. The age-depth model (Figure 2) was calculated using OxCal 4.4 P-Sequence (parameter k = 1; Bronk Ramsey, 1994, 1995, 2001; Bronk Ramsey et al., 2001) and the IntCal 20 calibration curve (Reimer et al., 2020) to allow for Bayesian radiocarbon wiggle-matching and thus very high chronological precision (Rey et al., 2019b, 2023).
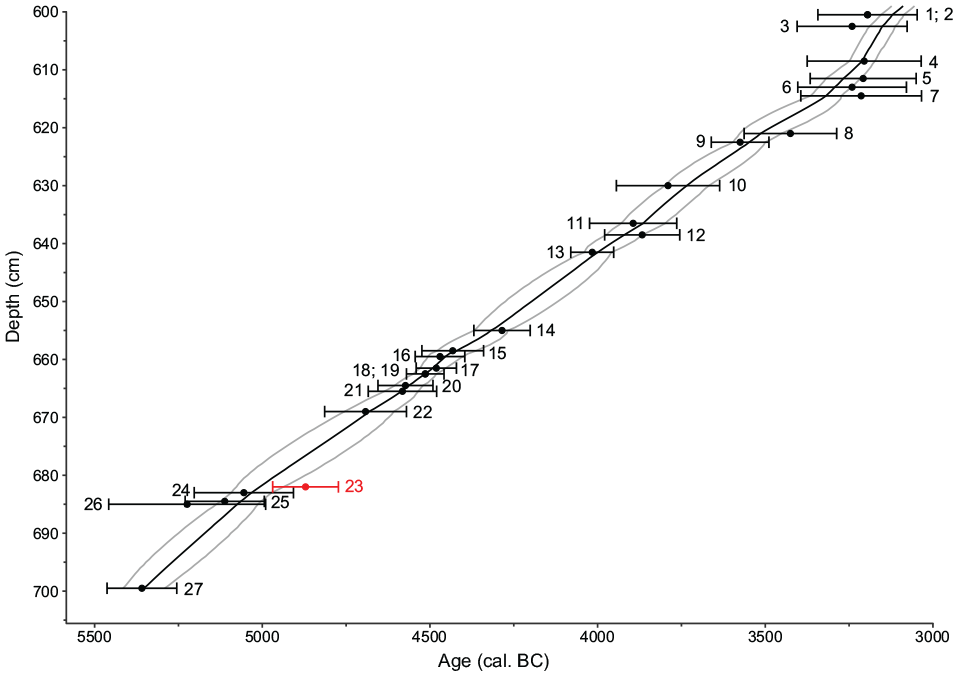
Age-depth model of Lago di Mezzano. Black points represent the calibrated ages of 14C dated terrestrial plant macrofossils with 95% (2σ) error bars (IntCal20; Reimer et al., 2020). The black line is the P-sequence modeled chronology (OxCal; Bronk Ramsey, 1994, 1995, 2001; Bronk Ramsey et al., 2001). Gray lines show the 95% (2σ) confidence interval of the model. The red point represents an age that was treated as an outlier. Ages numbering is following Supplemental Table 1.
Pollen, spores and microscopic charcoal analyses
A total of 177 samples of 0.5 cm3 were collected continuously between 687 and 599 cm for pollen, spores, algae, and microscopic charcoal analyses. This strategy results in a resolution of c. 11 years/sample ±2.7 years (mean ± standard deviation) over 2000 years. Pollen samples were prepared following standard procedure for glycerin (Moore et al., 1991), with HCl, HF, KOH, acetolysis, sieving (0.5 mm mesh size), and decanting. We added Lycopodium tablets to the fresh samples for pollen concentrations (grains cm−3) and influx values (grains cm−2 yr−1) estimation (Stockmarr, 1971). Pollen and spores were identified and counted using a light microscope at a magnification of 400× or 1000× using pollen keys and atlases (e.g. Beug, 2004; Moore et al., 1991; Punt, 1976; Reille, 1992), and the reference collection of the Institute of Plant Sciences of the University of Bern. On average, the pollen sum of each analyzed sample is 687 ± 77 pollen grains, excluding spores and pollen of aquatic plants. Microscopic charcoal particles (>10 µm) were analyzed on pollen slides following Tinner and Hu (2003) and Finsinger and Tinner (2005). Microscopic charcoal concentrations (particles cm−3) and influx values (particles cm−2 yr−1) were calculated with respect to the added Lycopodium spores and/or the depositional time, respectively. The pollen diagram (Figure 3) was divided into local pollen assemblage zones (LPAZ) using optimal partitioning with minimum sum-of-squares (Birks and Gordon, 1985). We assessed the number of statistically significant zones according to the broken-stick method (Bennett, 1996).

(a) Arboreal pollen (AP) percentages of selected taxa and microscopic charcoal influx (particles cm−2 year−1) diagram as well as lithology of Lago di Mezzano. Macrofossils represented as yellow dots (Fagus sylvatica bud scales) and red dots (Alnus glutinosa catkins and fruits). (b) Selected non arboreal pollen (NAP) percentage, spores, microscopic charcoal influx (particles cm−2 year−1), and lithology of Lago di Mezzano. Water plants, ferns, and other spores are excluded from the pollen sum. Palynological richness PRI (=N0), palynological evenness (PIE), and agricultural land use probability index (LUP; Deza-Araujo et al., 2022) are also shown. LPAZ: local pollen assemblage zones. Empty curves show 10x exaggerations.
Biogeochemical analyses
The analysis of geochemical elements was conducted at the Institute of Geological Sciences, University of Bern, using a Cr-tube equipped ITRAX XRF core scanner (Cox Ltd., Sweden). For the laminated sequence considered in this study, measurements were made at a resolution of 200 µm and integration time of 30 s at 30 kV and 50 mA, and are reported as element counts. Particularly interesting are titanium (Ti) as an indicator of terrigenous detrital sediment delivery from the watershed (Haug et al., 2001; Peterson et al., 2000), the ratio silicon–titanium (Si/Ti) as an indicator of biogenic silica (Melles et al., 2012), phosphorus (P) as an indicator of nutrient enrichment (Corella et al., 2012), and iron (Fe) as an indicator of erosion or oxic conditions (Makri et al., 2021).
Hyperspectral imaging (HSI) scans were conducted at the Institute of Geography, University of Bern, using a Specim Ltd. Single Core Scanner equipped with a PFD-CL-65V10E line-scan camera (Butz et al., 2015; spectral resolution of 2.8 nm, ranging from 400 to 1000 nm). Measurements were performed with a spatial resolution of 83 μm and an exposure time of 160 ms (Aperture: 1.9, Spectral bin: 2, FOV: 109.33 mm, speed 0.5, fr: 6 Hz). Data were then post-processed following Butz et al. (2015, 2016, 2017) and Zander et al. (2021): calculation of relative absorption band depths RABD655–685max and RABD845 as proxies for sedimentary green pigments (chlorophyll a and b and diagenetic products, TChl) and pigments of anoxygenic phototrophic bacteria (bacteriopheophytin a and b), respectively. The RABD655–685max index (TChl) represents algae bearing green pigments such as diatoms, cyanobacteria, green algae, and golden algae and is used as an indicator of primary productivity in the lake (Leavitt and Hodgson, 2002). The RABD845 index (Bphe) represents phototrophic purple sulfur bacteria (PSBs) that live at the chemocline in stratified lakes (Butz et al., 2016; Yurkov and Beatty, 1998) and is used as an indicator of hypolimnetic anoxia reaching the photic zone. We performed the calibration of the RABD indices to pigment concentrations (μg/gdry sediment) on 38 sediment samples with a minimum dry weight of 1 g. First, we conducted pigment extractions using 100% acetone following a modified version of the method in Sanchini and Grosjean (2020). Then, we measured the pigment concentrations of the extracts with a spectrophotometer (Shimadzu UV1800) and calculated concentrations using the molar extinction coefficient for bacteriopheophytin a from Fiedor et al. (2002) and the molar extinction coefficient for total chloropigments a from Jeffrey and Humphrey (1975). A linear regression model between the spectrophotometer measurements and the average RABD index values of the sample locations was used for calibration (Bphe = 508.94 * RABD845–503.62, R2 = 0.69; p < 0.01; RMSEP = 3.38 μg/gdry sediment; TChl = 1527.7 * RABD655-685max–1625.7, R2 = 0.79, p < 0.01, RMSEP = 47.3 μg/gdry sediment). Moreover, we used the R850/R900 spectral ratio as a proxy for volcanic lithogenic influx (Saunders et al., 2018). For the profile representation (Figure 4), we averaged elements counts, ratios and concentrations over a 5 mm wide subset.
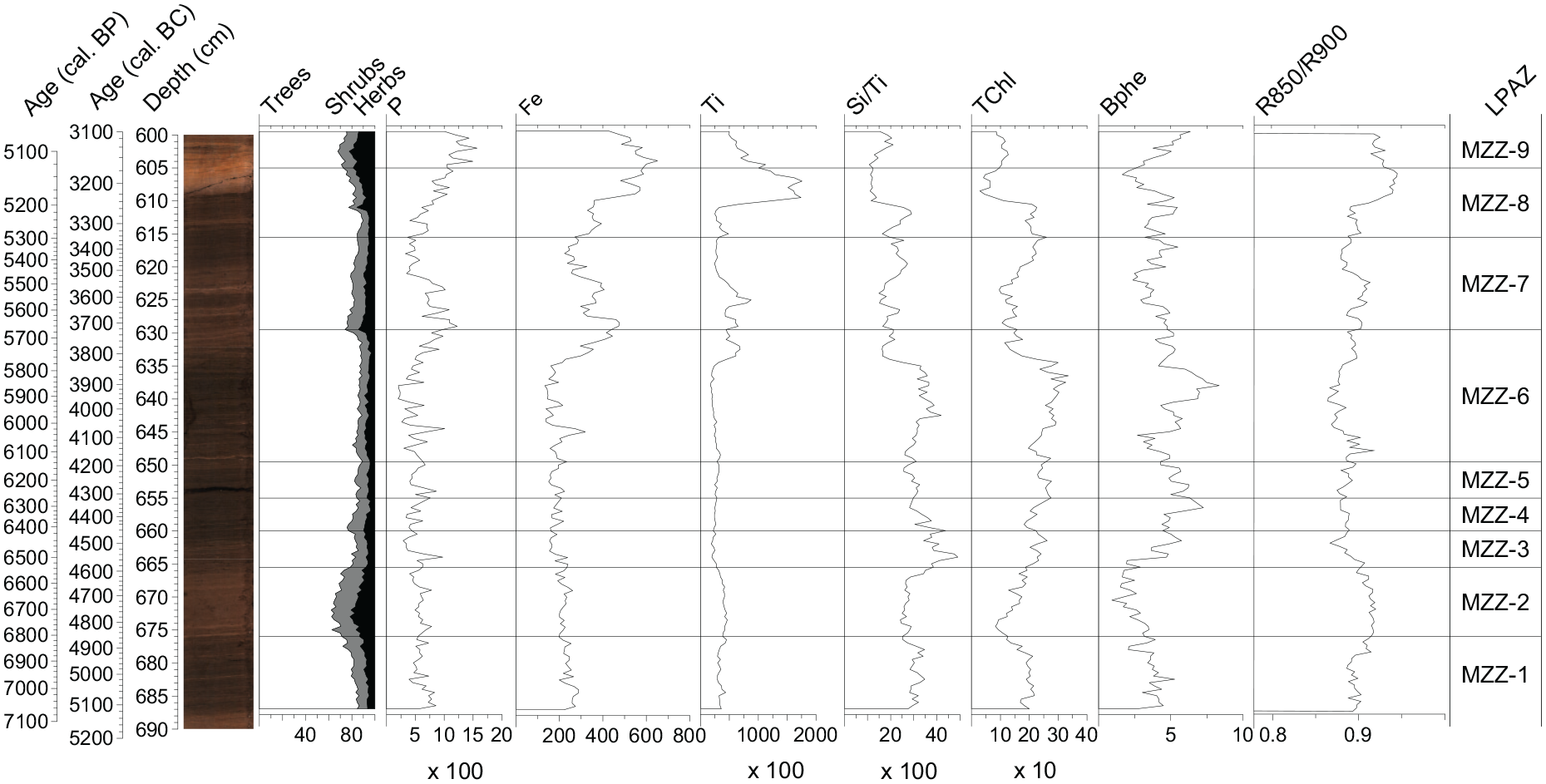
Photograph of the sediment section of Lago di Mezzano and comparison of tree, shrub, and herb pollen percentages, element counts and ratios from X-ray fluorescence (XRF) data (P, Ti, Si/Ti; cts), and hyperspectral imaging (HSI) for total aquatic productivity (TChl; µg/gdry sediment), hypolimnetic anoxia (Bphe; µg/gdry sediment), and volcanic lithogenic influx (R850/R900).
Corchia Cave CC27
The stalagmite CC27 from Corchia Cave, located in the Apuan Alps in Tuscany, central Italy, at about 200 km from Lago di Mezzano, was analyzed for stable isotopes (δ13C and δ18O) and trace elements (Mg, U, P, Y). A composite mean anomaly index was produced as a precipitation indicator by combining the different individual trace element records (Isola et al., 2019; Regattieri et al., 2014). Full methodological descriptions of the geological analyses are provided in Isola et al. (2019), in which the mean anomaly index is presented from 4050 cal. BC (6000 cal. BP) to the present. Here we extend the Corchia cave precipitation proxy to the period 4050–5100 cal. BC to match the other time series.
Numerical analyses
Biodiversity estimations
To investigate biodiversity dynamics around Lago di Mezzano, we calculated the palynological richness (PRI) corresponding to Hill number N0, the probability of interspecific encounter (PIE), and the diversity Hill number N2 (Hill, 1973). PRI is a proxy for species richness (Birks and Line, 1992), whereas PIE (Hurlbert, 1971) is used as a measure of evenness (e.g. Tolmos et al., 2022). The Hill number N2 is an estimate of the taxa abundance distributions and thus it is related to evenness as well (Felde et al., 2016; van Vugt et al., 2022). Evenness-detrended palynological richness (DE-PRI; Colombaroli and Tinner, 2013) accounts for the possible influence of palynological evenness (PIE) on palynological richness (PRI). For the comparison among sites, we re-calculated PRI for Lago di Origlio (submediterranean site; Tinner et al., 1999) and Moossee (temperate site; Rey et al., 2019a) for minimum pollen sums of 500. Rarefaction analyses according to Birks and Line (1992) were performed using vegan (Version 2.6-4; Oksanen et al., 2022), a community ecology package developed to be used with the software R (R Core Team, 2022). PIE was estimated for the first time at Lago di Origlio (this work), while it had been already calculated at Moossee (Rey et al., 2019a).
Land use probability index (LUP) and fire number (FN) estimates
Several plants producing pollen of cultural indicators such as Olea, Castanea, Cerealia t. (including Triticum t. and Hordeum t.; t. = type), and Plantago lanceolata t. are native to Mediterranean regions, and their presence in the pollen diagram is therefore difficult to interpret (Deza-Araujo et al., 2020; Mercuri et al., 2013; Sadori, 2018). To overcome this difficulty and to estimate the intensity of human impact, we used the novel land use probability index LUP (Deza-Araujo et al., 2022). The LUP index summarizes the ecological knowledge on the indicator values of taxa in a probabilistic way for different biomes of Europe, by providing locally suited weights (anthropogenic indicator values, AIV) to pollen of crops, adventives and apophytes. It has been tested with independent palynological evidence spanning from coldest arctic-alpine to warmest thermomediterranean environments as well as with local archaeological evidence (Deza-Araujo et al., 2022). The training and validation sets comprise sites in northern, central and southern Italy. Recently it has been successfully applied to European regions outside the validation range (e.g. Brechbühl et al., 2023). For Lago di Mezzano, we used the AIV for the mesomediterranean vegetation.
To be able to compare fire activity among study sites avoiding bias due to analytical and site-specific morphological differences, we calculated the fire number (FN = # of fires 1000 km−2 year−1) for Lago di Mezzano, Lago di Origlio, and Moossee. FN is calculated by using a charcoal influx-based transfer function, developed by Adolf et al. (2018), which is derived from a continental calibration effort for microscopic and macroscopic charcoal influx (as estimated from annually collected surface sediments) with satellite images of burning. FN is reconstructed within a 40-km radius of the deepest point of the lakes considered in the study (Adolf et al., 2018) and adjusted for an area of 1000 km2.
Time series analysis (cross-correlations)
We calculated cross-correlations to estimate effects of fire (microscopic charcoal influx), pastoral (Sporormiella dung spore influx), and human activities (land use probability index LUP; Deza-Araujo et al., 2022) on vegetation (pollen percentages) and diversity (PRI, PIE). Such multivariate, parametric time-series analyses are well-established in palaeoecology (Clark et al., 1989; Dodson, 1990; Green, 1981; Lang et al., 2023; Tinner et al., 1999) and used to calculate lag or lead effects for one variable (response or dependent variable) in response to another variable (driving or independent variable). Cross-correlation analyses compare the values of the driving and response variables by moving the two sequences of values against each other for a given number of time lags. At each time lag, the correlation coefficient between the values of the two variables is calculated (Bahrenberg et al., 2008). Data were linearly de-trended to remove major trends from the time series, and only variables with different units have been cross-correlated (e.g. influx versus percentages) to avoid artificial correlations due to sedimentation rates or dependence among variables within percentages. In addition to the sequence of Lago di Mezzano (this study), we also re-estimated the cross-correlations at Lago di Origlio (Tinner et al., 1999) and Moossee (Rey et al., 2019a) for the common period 5100–3100 cal. BC. The three analyzed series include 177 lags (1 lag = 11.5 ± 2.7 years) for Lago di Mezzano, 173 lags (1 lag = 11.7 ± 1.4 years) for Lago di Origlio, and 189 lags (1 lag = 10.8 ± 3.3 years) for Moossee. Cross-correlation coefficients were calculated at ±30 lags, corresponding to c. ±340 cal. years, that is, less than one-fourth of the sample number N (Bahrenberg et al., 2008). We tested the significance of the cross-correlation coefficients by calculating ±2 SE (standard errors) of the correlation coefficients, which corresponds to a two-sided significance level (α) of 5% (Bahrenberg et al., 2008; Tinner et al., 1999).
Results and interpretation
Chronology and lithology
The sediments of Lago di Mezzano corresponding to the period 5100−3100 cal. BC are carbonaceous (mainly composed of silty gyttja). The P-sequence age-depth model, which relies on Bayesian statistics (Figure 2), has a mean modeled uncertainty of ±52 cal. years (2σ), while the minimum uncertainty is ±28 cal. years (2σ) around 3100 cal. BC (Supplemental Table 1). The model suggests an approximately constant depositional time of c. 23 years cm−1, with a maximum of 37 years cm−1 at c. 4400 cal. BC and minimum of 9 years cm−1 at c. 3200 cal. BC. One radiocarbon date (sample 23: BE-13286.1.1; Supplemental Table 1, Figure 2) was treated as an outlier since it did not fit with the model in the Bayesian analysis.
Pollen, charcoal, and biogeochemical analyses inferred vegetation, land use, and fire dynamics
Throughout our 2000-year-long record (5100−3100 cal. BC), nine statistically significant local pollen assemblage zones (LPAZ) have been identified, suggesting several important vegetation changes (Figure 3a and b). In general, pollen percentages, concentration, and influx values show similar trends for most trees, shrubs, and herbs (Supplemental Figure 1). Nevertheless, low concentration and influx values for specific taxa such as Quercus robur t., Quercus ilex t. and Carpinus betulus are not mirrored in their percentage values during, for example, LPAZ MZZ-4. This might point to distortion effects such as a percental overrepresentation of these tree taxa during some forest opening phases or alternatively, problems related to pollen deposition affecting both, concentrations and influx.
Tree pollen percentages show high values (>80% of the pollen sum) along most of the sequence, indicating the presence of rather closed forests around Lago di Mezzano during the Neolithic from 5100 to 3100 cal. BC. Among the most important taxa are Q. robur t. (>20% of the pollen sum), Fagus (10−20%), Q. ilex t. (c. 15%), Carpinus betulus (5−10%), Corylus (5−10%), Quercus cerris t. (3−10%), and Alnus glutinosa t. (up to 40%). The unexpected existence of Fagus forests near the lake in a mesomediterranean vegetation context is supported by frequent findings of plant macrofossils such as bud scales (Figure 3a). Similarly, Alnus glutinosa macrofossils testify to the presence of the tree species, likely close to the shores (Figure 3a). Other arboreal taxa with lower values but regular occurrences over the entire pollen record are: Erica t., Ilex aquifolium, Hedera helix, Acer, Ulmus, Tilia, Fraxinus excelsior t., Fraxinus ornus, Ostrya t., Castanea sativa, Pistacia, Phillyrea, Sambucus nigra t., Prunus, Rubus, Frangula alnus, Viburnum, and Salix. The co-occurrence of these taxa belonging to different plant communities and vegetation zones (temperate to Mediterranean) suggests that the vegetation around Lago di Mezzano was highly varied, probably depending on different altitudes (relevant for e.g. moisture along the shores or cold air inversions), soil types, and slope aspect.
Percentages of the shore tree A. glutinosa t. pollen show high and constant values (c. 30%) for most of the record. Exceptions are the zones MZZ-1 to MZZ-4 (Figure 3a), with a collapse between 4850 and 4600 cal. BC (<5%) in MZZ-2, followed by a constant recovery toward values of >30% in MZZ-3 and MZZ-4. During the pollen- and macrofossil-inferred collapse of alder stands, mesophilous Fagus also declined and wetland plants such as Salix, Cyperaceae, Sparganium t., and Pediastrum expanded (MZZ-2), likely pointing to lake level oscillations. Lake eutrophication seems less likely because for instance high Pediastrum values are usually associated to low in lake productivity (as derived from TChl, Figures 3 and 4).
In the following, we summarize zones with similar vegetation history patterns: LPAZ MZZ-2, MZZ-4, MZZ-7, and MZZ-9, which correspond to the time 4850−4600, 4450−4300, 3750−3350, and from 3200 cal. BC onwards. In these zones lower tree pollen percentages (<80%) suggest forest opening phases characterized by marked decreases in Fagus pollen percentages as well as increases of herbaceous taxa, including primary anthropogenic indicators such as Triticum t. and Hordeum t., and secondary indicators such as Plantago lanceolata t., Rumex acetosella t., Urtica, Sporormiella dung spores, and Pteridium aquilinum. In agreement, increased LUP values (Figure 3b; Deza-Araujo et al., 2022) and higher percentages of the light-loving shrub Corylus avellana, suggest increasing pastoral and agricultural activities during these opening phases. Microscopic charcoal influx values, as a proxy for fire activity, are higher during MZZ-2, MZZ-7, and MZZ-9 (peaks around 15,000, 16,000, and 80,000 particles cm−2 year−1, respectively) compared to the rest of the sequence. Using continental transfer functions for microscopic charcoal (Adolf et al., 2018) this translates to FN of c. 2.3, 2.4, and 7.3 fires 1000 km−2 year−1, respectively. Percentages of Pediastrum also increase during these phases, possibly reflecting changes in lake water properties (Jankovská & Komárek, 2000; Masi et al., 2018; Sadori et al., 2015). Simultaneous high values of Ti and R850/R900 (Figure 4) may point toward increasing erosional input. Slash-and-burn and agricultural activities may therefore have been the main cause of the repeated forest openings and the associated erosion (see also results on cross-correlation, chapter “Time-series analyses”, showing strong reductions of trees and increases of cereal cultivation in response to fires).
The first three land use phases of our record (LPAZ MZZ-2, MZZ-4, and MZZ-7) show common successional patterns. Initially, the opening phase is characterized by declines in tree percentages, mainly represented by Fagus (from 14% to 5%) and an increase in herbaceous and anthropogenic taxa. Subsequently, after c. 10 years, pollen from light-loving and disturbance-adapted Corylus shrubs increases as well (from 4% to 10%), followed by Carpinus betulus (from 4% to 10%) about 90 years later. Finally, after c. 170 years from the beginning of the forest opening phase, Fagus reaches high values again (c. 14−15%; Figures 3a and 5), likely revealing repeated successional recovery cycles after forest burning for land use. Q. robur t. and Q. ilex t. remain far more stable than Fagus in percentages, concentrations, and influx values (Figure 3 and Supplemental Figure 1). During the youngest reconstructed land use phase (MZZ-9), Fagus pollen remains constant, while evergreen Q. ilex t. and deciduous Q. robur t. decline markedly (in percentages, concentration, and influx), together with a sharp increase in microscopic charcoal values (reaching 80,703 particles cm−2 year−1 at c. 3190 cal. BC), corresponding to a FN of c. 7.3 fires 1000 km−2 year−1. Such a divergent pattern might indicate different land and forest use practices during this time compared to previous land use phases.
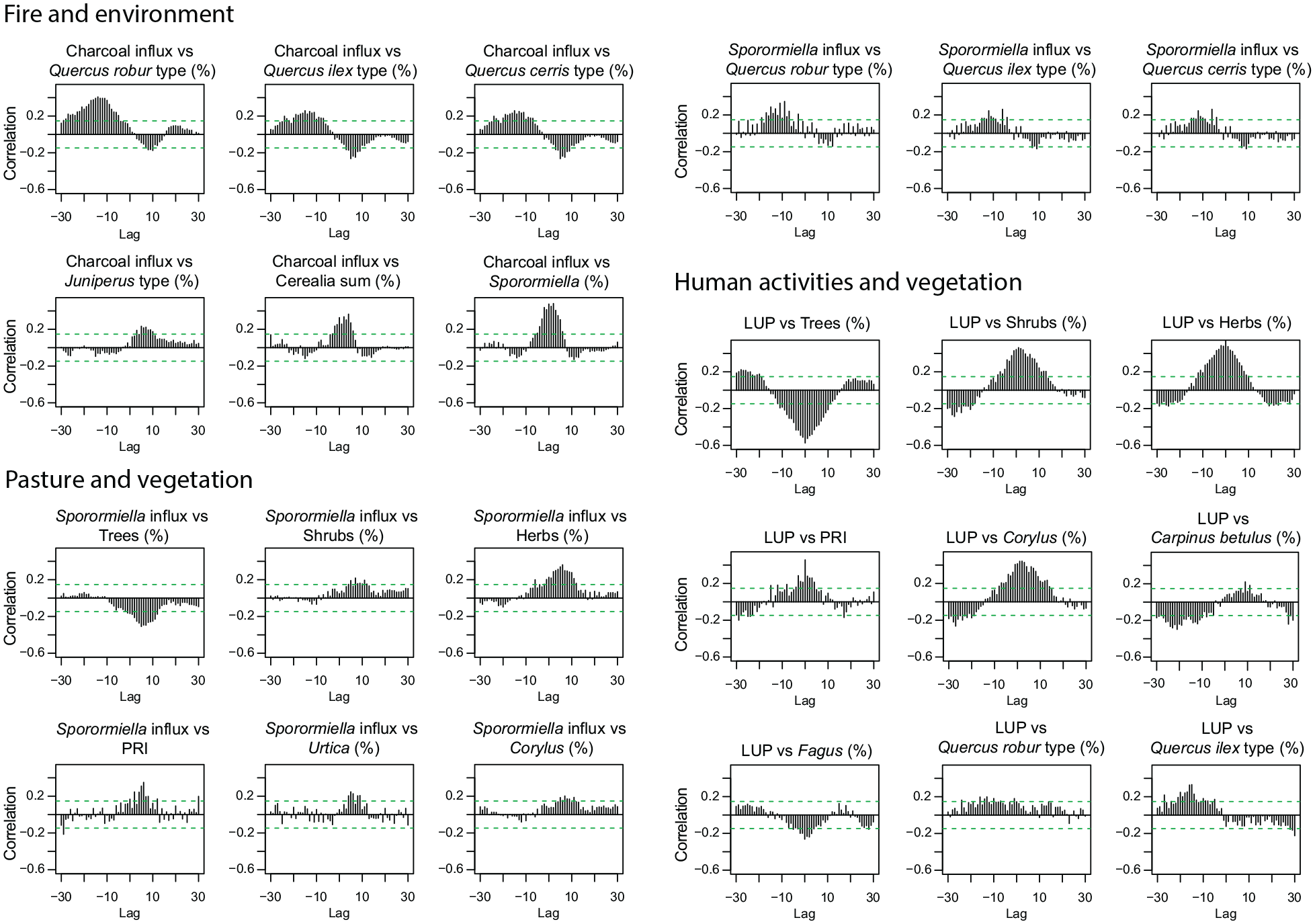
Cross-correlation of microscopic charcoal influx (particles cm−2 year−1), Sporormiella dung spores influx (particles cm−2 year−1) as proxy for pastoral activities, and agricultural land use probability index (LUP; Deza-Araujo et al., 2022) versus selected terrestrial pollen and spores percentages from Lago di Mezzano (5100−3100 cal. BC). All variables were de-trended. 1 lag corresponds to 11.5 ± 2.7 years (mean ± standard deviation). The green dashed lines mark significance level (p < 0.05).
The LPAZ MZZ-1, MZZ-3, MZZ-5, MZZ-6, and MZZ-8 show an opposite trend with phases of forest closure (tree pollen >80%), dominated by Fagus, Q. robur t., and Q. ilex t. Additionally, we observe low microscopic charcoal values (c. 3900−6900 particles cm−2 year−1) in these zones, with few exceptions in MZZ-1 (peaks reaching c. 20,000 particles cm−2 year−1). High values of TChl, Si/Ti, and Bphe were also recorded (Figure 4), pointing toward anoxic conditions and increased primary production (Boehrer et al., 2017; Boehrer and Schultze, 2008) under rather closed forest conditions. Under anoxic conditions, P is often recycled from the sediments back into the water column leading to generally low sedimentary P concentrations during anoxic phases (e.g. Makri et al., 2020; Zander et al., 2021). However, in our record, sedimentary P appears to be controlled by another process as it follows the same trend as indicators for detrital input such as Ti, suggesting P is retained more effectively in the sediment during phases of high detrital input despite water column anoxia as suggested by the presence of Bphe throughout the record. High detrital input may indicate increased soil erosion which would also deliver iron oxyhydroxide to the lake which in turn would scavenge P from the water column and contribute to retaining P in the sediment (Tu et al., 2021). Among the mentioned LPAZ, MZZ-6 (4200−3750 cal. BC) shows some particular characteristics. On one hand, high percentages of tree pollen (>80%) reflect stable forest cover with deciduous forests dominated by beech (Fagus) and oak (Q. robur t.) and evergreen forests with Q. ilex. This is supported by lowest microscopic charcoal influx values in the diagram (average 4280 particles cm−2 year−1) suggesting low forest fire activity in the region. On the other hand, during LPAZ MZZ-6, anthropogenic indicators such as Cerealia t., Triticum t., Hordeum t., P. lanceolata t., Urtica, Pteridium aquilinum and Sporormiella, as well as increased LUP values point to farming activities including forest grazing close to the lake when NAP are slightly higher (i.e. 4150−3900 cal. BC).
Biodiversity reconstruction
Palynological richness (PRI or Hill number N0) shows only slight variations over the whole sequence. However, higher values of PRI generally occur when percentages of tree pollen are low (Figure 3b). This suggests that, although the forests around Lago di Mezzano were very diverse, their opening for cultivation purposes (see increasing of anthropogenic taxa; Figure 3b) created new habitats for species characteristic of open lands. PIE (as a proxy for evenness) and Hill number N2 show opposite dynamics to those of PRI during some periods (e.g. 4700−4500, 4300−4100, 3900−3700 cal. BC; Figure 3b and Supplemental Figure 2), while during other periods they co-fluctuate with PRI (e.g. 4500−4300), suggesting that richness and evenness changes were not linearly linked. Indeed, since PRI and DE-PRI generally show comparable trends, we assume that PIE has no major effects on PRI (Colombaroli and Tinner, 2013). An exception occurs at around 4450 cal BC when PIE-inferred evenness and PRI decrease together, likely resulting from a PRI-distortion related to the dominance of Alnus glutinosa t. during this period (Figure 3b). DE-PRI corrects this palynological bias by producing values higher than PRI, as also documented in calibration studies comparing surface pollen and vegetation (Senn et al., 2022). PRI, PIE, and N2 are generally positively correlated with microscopic charcoal, suggesting a beneficial influence of fires on species diversity and evenness (Figures 3b and Supplemental Figure 2).
Time-series analyses
Cross-correlation analysis of microscopic charcoal influx versus pollen percentages of selected plant taxa shows the relationship between fires and vegetation, disclosing leads and lags. At Lago di Mezzano, microscopic charcoal influx is negatively correlated with all Quercus types found in the pollen sequence (Figure 5). Quercus ilex t. and Quercus cerris t. show a maximum negative correlation at lag +5 (c. 55 years after fire), while Quercus robur t. suffers longer, reaching its fire-induced minimum at lag +10. On the other hand, shrub taxa such as Corylus and Juniperus t. show a maximum positive correlation with microscopic charcoal influx at lag +5, suggesting that fire favored the spread of these pioneer and light-loving shrubs which peaked c. 55 years later.
Sporormiella and Cerealia t. (including Triticum t. and Hordeum t.), as proxies for pastoral and agricultural activities, respectively, show a maximum positive correlation with microscopic charcoal influx at lag +2 and +4, pointing out that land use was supported by fires. Both proxies are also significantly positively correlated with microscopic charcoal influx values at lag −5 and lag −3, suggesting that first human activities began before the forest opening was maximized with fire to expand the area for land use (Tinner et al., 1999). Moreover, the land use probability index (LUP) shows a maximum negative correlation with trees and a maximum positive correlation with shrubs and herbs at lag 0. Together, these findings show that the forest was opened for agricultural and pastoral activities. Cross-correlations between Sporormiella influx values and percentages of plant taxa (e.g. trees, shrubs, herbs, Quercus, Corylus, PRI) show similar patterns as the cross-correlations between microscopic charcoal influx and the same response variables, suggesting a close link between pastoral activities and fire. Hence, increasing pastoralism, as indicated by Sporormiella influx, affected trees negatively and shrubs and herbs positively with a delay between c. 60 and 80 years (Figure 5). Interestingly, Fagus shows a maximum negative correlation with LUP at lag 0 and no correlation with microscopic charcoal influx. This may suggest that the beech forest was not primarily opened with fire to create space for cultivation but also through logging activities. Sporormiella influx impacts PRI with a maximum positive correlation at lag +7, indicating a positive effect of pastoral activities on vegetation diversity. Moreover, LUP is strongly and significantly correlated at lag 0 with PRI, suggesting that Neolithic human activities had a positive impact on vegetation diversity around Lago di Mezzano.
Recalculations of cross-correlations for Lago di Origlio (submediterranean site; Tinner et al., 1999) and Moossee (temperate site; Rey et al., 2019a) confirm earlier results and reveal common patterns with Lago di Mezzano in vegetation-fire-interactions. Microscopic charcoal influx is negatively correlated with percentages of tree pollen at all three sites, although at Lago di Mezzano the maximum negative correlation is delayed by c. 55 years. Shrubs and herbs are also positively correlated with microscopic charcoal influx at all three sites. Among the shrubs, Corylus was particularly fostered by fires at all three sites. Fraxinus excelsior t. and Quercus robur t. are negatively impacted by fire at all three sites. Specifically, the overall pattern of the Quercus robur t. cross-correlation with charcoal is almost identical at Lago di Mezzano and Lago di Origlio, and still similar at Moossee (Figure 6). The short response time of F. excelsior t. to microscopic charcoal influx highlights the high sensitivity of this taxon to fire in all three vegetation types. At Lago di Origlio and Moossee, Fagus is negatively correlated with microscopic charcoal influx values, whereas at Lago di Mezzano it shows no correlation, suggesting other causes of disturbance besides fires. Abies is negatively correlated with a short response time at Lago di Mezzano and Lago di Origlio, whereas at Moossee it responds first positively and later negatively to fire. This might be explained by different fire severities and the sensitivity of this species under submediterranean to mesomediterranean climates and fire regimes. Interestingly, PRI is positively correlated with microscopic charcoal influx at Lago di Mezzano and Moossee and negatively correlated at Lago di Origlio. This finding is likely related to significantly higher fire disturbance (e.g. frequency, intensity) at Lago di Origlio, if compared to the two other sites (Figures 7 and Supplemental Figure 3).
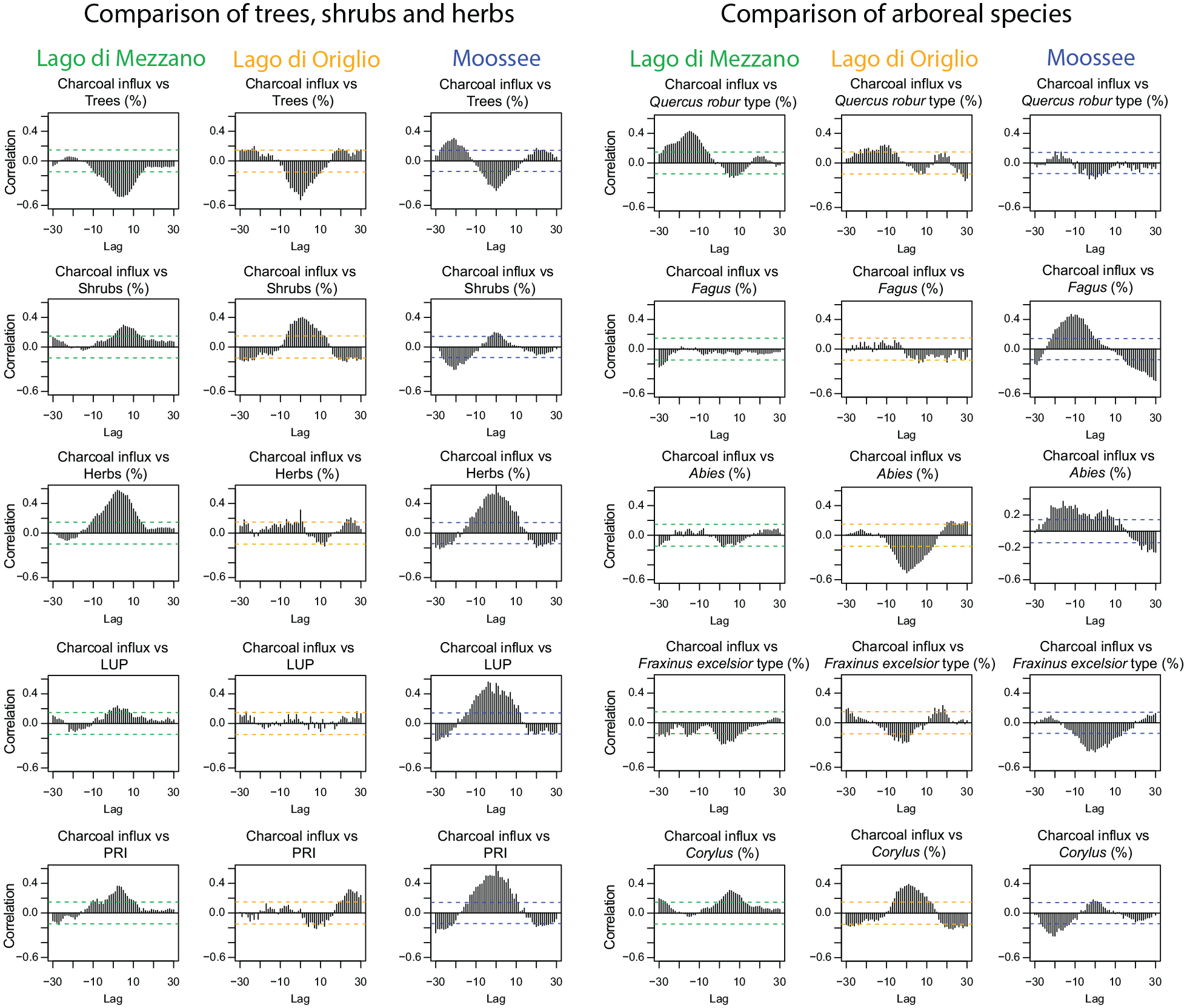
Cross-correlation diagrams of microscopic charcoal influx (particles cm−2 year−1) versus selected terrestrial pollen percentages and indices from Lago di Mezzano, Lago di Origlio (Tinner et al., 1999), and Moossee (Rey et al., 2019a) for the period 5100−3100 cal. BC. All variables were de-trended. 1 lag corresponds to 11.5 ± 2.7 (Lago di Mezzano), 11.7 ± 1.4 (Lago di Origlio), and 10.8 ± 3.32 (Moossee). The horizontal dashed lines mark significance level (p < 0.05).
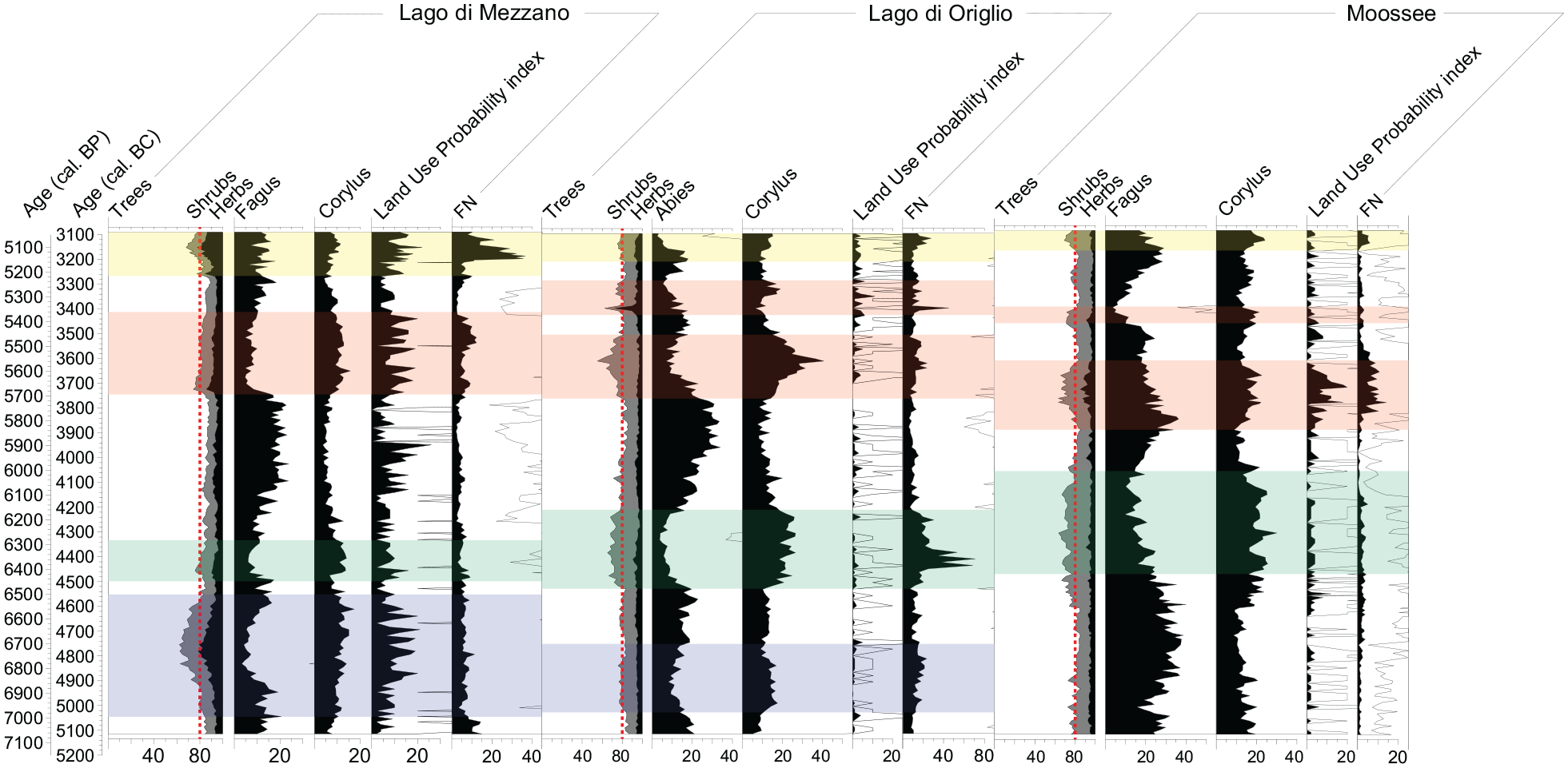
Records comparison from contiguously sampled high-resolution sites Lago di Mezzano, Lago di Origlio (Tinner et al., 1999), and Moossee (Rey et al., 2019a). Main pollen diagrams showing pollen percentages of taxa that are particularly affected during forest opening phases, agricultural land use probability index (LUP; Deza-Araujo et al., 2022) and fire number (FN = # of fires/1000 km2/year; Adolf et al., 2018). Computation of fire number (FN) relies on a transfer function using charcoal influx as derived from a continental calibration effort based on satellite images of burning, according to Adolf et al. (2018). Comparable opening phases are highlighted in colors. Red dashed lines indicate 80% of tree pollen (on total terrestrial pollen sum). Empty curves show 10× exaggerations.
Discussion
The conversion of natural vegetation for land use in the Lago di Mezzano area
During the Neolithic, the co-dominance of evergreen broadleaved Quercus ilex together with temperate deciduous trees such as Quercus pubescens and/or Quercus petraea (within Quercus robur t.), Quercus cerris, and Fagus sylvatica suggests a highly diverse vegetation composition in the Lago di Mezzano area. The diversity of habitats in and around the Caldera of Latera, characterized by different soil types, altitudes, and slope aspects, allowed the formation of a vegetational composition spanning from oromediterranean moisture-loving beech forests, to intermediate submediterranean deciduous oak forests with, for example, Q. cerris and Q. pubescens, to mesomediterranean warm-loving evergreen holm-oak forests. Moreover, our pollen data suggest that secondary trees and shrubs in the Mezzano area comprised, among others, Corylus, Ulmus, Tilia, Acer, Vitis, Fraxinus excelsior (and/or F. angustifolia), F. ornus, Ostrya carpinifolia, Abies alba, Hedera helix, Juniperus, Taxus, Ilex, Phillyrea, Pistacia, and probably also Olea europaea and Castanea sativa. Indeed, stretching from the Tyrrhenian Sea to the Apennines, Latium has a large diversity of species (Blasi, 1994) that were well represented in the Caldera of Latera during the Neolithic (Sadori, 2018).
Our results suggest that the rather dense mesomediterranean to oromediterranean oak-beech forests around Lago di Mezzano were opened repeatedly by fire, cutting, browsing, and grazing during the Neolithic periods 4950−4600, 4450−4300, 3750−3350, and 3200−3100 cal. BC. Disentangling climatic and human influence on fire and vegetation dynamics is one of the major challenges when aiming to understand past interactions between vegetation, climate, fire, and land use. At Lago di Mezzano, all forest opening phases occurred in conjunction with increasing anthropogenic indicators (Figure 3a and b). Our cross-correlation analysis underlines the role of fire and grazing during land use phases (Figure 5) and point to the practice of slash-and-burn activities. This technique has been employed by human populations to create open land for pasture and agriculture since the Neolithic (Clark et al., 1989; Colombaroli et al., 2008; Dietre et al., 2017; Doyen et al., 2016) and does not imply shifting cultivation (Baum et al., 2020). Intriguingly, Mesolithic foraging cultures may already have used fire to open forests to improve the subsistence basis (Heidgen et al., 2022). During the Neolithic, forest opening, in addition to providing space for agriculture and pasture (e.g. crops such as Triticum and Hordeum or adventives and apophytes such as Plantago lanceolata, Poaceae, and Rumex acetosella), created new habitats that could be colonized by many herbaceous plants including Caryophyllaceae, Ranunculaceae, and Asteraceae (Figure 3b).
Our cross-correlations results using LUP as the independent variable (Figure 5) strongly suggest anthropogenic activities as the main drivers of vegetation dynamics during the Neolithic forest opening phases (5100–3100 cal. BC). The Neolithic in Italy started around 8000 years ago (e.g. Bagolini, 1987; Malone, 2003; Palmisano et al., 2017; Persiani, 2009; Petitti and Rossi, 2012; Robb, 2007). For instance, la Marmotta, a Neolithic lake-shore community that was technologically advanced in navigation, construction of houses, agriculture, hunting, and crafts, existed between 5700 and 5300 cal. BC at Anguillara Sabazia on Lago di Bracciano, c. 80 km south-east of Lago di Mezzano (Fermé et al., 2021; Fugazzola Delpino et al., 1993), indicating that local human impacts on vegetation around lakes may have already started as early as 6000–5000 cal. BC.
During land use phases, herbaceous biodiversity increased in the grasslands and other open spaces in response to anthropogenic activities (Figure 3b). In agreement, studies covering different biomes including meso- and submediterrenean vegetation have shown that open lands provide ideal habitats for a wide variety of herbaceous plants (Colombaroli and Tinner, 2013; Giesecke et al., 2014). Tree diversity, by contrast, remained relatively constant throughout the entire period 5100–3100 cal. BC (Supplemental Figure 4). This finding suggests a certain degree of forest resilience at local to extralocal scales (i.e. within the pollen-catchment of the site). Light-loving shrubs or small trees such as Corylus avellana and Juniperus t. expanded about a decade after land use peaked. Such pioneer species usually colonize free habitats after disturbance events such as fire, erosion, logging, grazing, and agriculture and represent one of the early stages of the ecological succession toward the forest climax (Pignatti et al., 2017). The expansion of Carpinus betulus followed that of C. avellana. This heliophilous and short-lived tree species rarely forms pure forests, and in the Italian lowlands it is often found with Q. robur or Castanea sativa, F. sylvatica, and C. avellana (Pignatti et al., 2017). At Lago di Mezzano, Carpinus betulus stands probably constituted a middle successional stage toward late successional beech-oak forest. Interestingly, the species is rather indifferent to changes in moisture availability, if compared to, for example, Fagus sylvatica and Quercus cerris (Pignatti et al., 2017).
At Lago di Mezzano, increasing in-lake productivity (TChl, Si/Ti) and hypolimnetic anoxia (high Bphe) during periods of forest closure was opposed to higher clastic input (Ti, R850/R900), oxic conditions (low Bphe), and decreasing productivity (TChl, Si/Ti) during forest opening phases (Figure 4). This finding likely reflects a rather strong link between in-lake and local upland environmental dynamics. Specifically, high forest cover likely increased wind shielding and soil stability, influencing lake biogeochemistry and mixing regime (Gassner et al., 2020; Lotter, 2001; Makri et al., 2020; Zander et al., 2021). However, the clear anti-correlation of in-lake productivity indicators with indicators of detrital input could also be the consequence of a dilution effect due to increased catchment erosion during forest opening phases. Our palaeoecological results, including the repeated findings of F. sylvatica macrofossils, suggest that oromediterranean beech forests grew near the lake. Indeed, HSI and the XRF data show a close connection with beech forest changes. For instance, high in-lake productivity (TChl; Figure 4) and low erosion (Ti and R850/R900; Figure 4) prevailed during periods of high beech forest cover (Figure 3a). On the other hand, mesomediterranean communities of Q. ilex, Pistacia, and Olea do not show such linkages. Therefore, on the basis of our multiproxy evidence, we hypothesize that F. sylvatica and other mesophilous temperate deciduous trees were growing on north-facing slopes and toward the bottom of the caldera (but outside the wetlands), where moisture and frost were likely more abundant and microclimatic conditions generally cooler. Oromediterranean and submediterranean species are more resistant to winter frost compared to Q. ilex and other evergreen broadleaved species, which probably grew on the warmer and drier south-facing slopes in and outside the Caldera of Latera.
Land use factors other than fire, grazing and browsing may have co-determined vegetation dynamics (Figures 5 and 6). For example, Fagus shows no correlation with microscopic charcoal from 5100 to 3100 cal. BC and a maximum negative correlation at lag 0 with LUP. This may reflect the use of its wood for construction purposes besides burning. F. sylvatica wood is valued for its hardness, resistance, and good workability, hence in Europe this tree species is today one of the most widely used in many fabrications such as furniture and cooking utensils, and for fire (San-Miguel-Ayanz et al., 2016). Indeed, in the Neolithic village La Marmotta on Lago di Bracciano beech wood was used for construction and navigation, although deciduous Quercus and Laurus nobilis wood was much more utilized (Beccaccioli et al., 2023; Fermé et al. 2021; Fugazzola Delpino and Tinazzi, 2010). Later on, during the Bronze Age at Lago di Mezzano, F. sylvatica was used along with other arboreal species for various lake-shore constructions as well (Sadori et al., 2004).
Effect of land use on forest vegetation in the wider regions of central and southern Italy
During the early and mid-Holocene, before the advent of agriculture, evergreen Quercus and Fagus sylvatica, together with several deciduous oak species, were predominant around Lago di Mezzano and other volcanic lakes in central Italy such as Lagaccione (Magri, 1999), Lago di Vico (Magri and Sadori, 1999), and Valle di Castiglione (Di Rita et al., 2013). The forests in the regions outside volcanic calderas were co-dominated by Abies alba instead of F. sylvatica (Colombaroli et al., 2007; Drescher-Schneider et al., 2007; Tinner et al., 2013). This difference in primeval central Italian lowland forests was likely caused by the higher heat and drought sensitivity of F. sylvatica (e.g. Ellenberg, 2009; Leuschner and Ellenberg, 2017), which allowed the species to thrive only in the rather cool and moist caldera environments.
Arable and pastoral farming spread in central Italy since the beginning of the Neolithic at about 6000 years BC (Malone, 2003). In the lowlands, lake shore villages established during the Neolithic (Mazzucco et al. 2022) and became widespread during the Bronze Age (e.g. Angelini et al., 2014; Angle et al., 2011; Sadori et al., 2004). Lake shore settlements influenced the local vegetation cover around the lakes, the water quality, as well as, for example, the erosional imprint in the lake sediments. In the uplands outside the calderas, human impact also increased (e.g. Colombaroli et al., 2008; Drescher-Schneider et al., 2007), ultimately leading to strong reductions in both mesophilous species A. alba and F. sylvatica all over central Italy. The strong negative human impact on lowland beech forests was also highlighted in a recent study by Buonincontri et al. (2023). However, in contrast to A. alba, which became rare also at higher altitudes in response to millennial-long human impact, F. sylvatica spread massively at higher altitudes in the Apennines (e.g. Allen et al., 2002; Branch, 2013; Cruise, 1990; Guido et al., 2013; Lowe, 1992; Morales-Molino et al., 2021; Vescovi et al., 2010; Watson, 1996). This difference is explained by the lower disturbance sensitivity of F. sylvatica if compared to A. alba. Although F. sylvatica also suffers from fires, its very high recruitment capacity allows it to recover and form increasingly pure beech stands (Ascoli et al., 2013, 2015; Maringer et al., 2016, 2020; Morales-Molino et al., 2021). Indeed, our high-resolution results show that beech forests always recovered, at least partially, after each land use phase (Figure 3a). F. sylvatica was common in southern Italy as well. Above the thermomediterranean, mesomediterranean and supramediterranean vegetation belts (Calò et al., 2012; Bisculm et al., 2012; Sadori and Narcisi, 2001; Tinner et al., 2009), F. sylvatica was dominant in the oromediterranean belt above c. 1200 m a.s.l., forming dense forests during the mid and late Holocene as south as Sicily (Tinner et al., 2016). At Lago Trifoglietti in southern Italy, F. sylvatica was co-dominant at high elevations (Joannin et al., 2012) during the entire Holocene. Very comparable to Lago di Mezzano, F. sylvatica also occurred in the caldera of Lago Grande di Monticchio (Allen et al., 2002). Still today the species co-dominates at low and intermediate altitudes in the caldera of Monte Vulture surrounding the Laghi di Monticchio (Spicciarelli et al., 2011), while it is absent in the submediterranean vegetation outside the caldera. To better understand the role of climate, land use and disturbance in the vegetation dynamics at the southern Italian locations, novel precisely dated, continuous, high-resolution multiproxy records are needed.
The role of climate
Palaeoecological, archaeological and modeling evidence suggests that in the past, human activities, vegetation dynamics and climatic conditions may have been closely linked to each other (e.g. Baum et al., 2020; Hafner and Schwörer, 2018; Lang et al., 2023; Maise, 1998; Rey et al., 2019a; Rösch and Lechterbeck, 2016; Tinner et al., 2003; Wirtz and Lemmen, 2003). At our site climate may have impacted the local wetland vegetation. During the human induced forest opening phase from 4950 to 4600 cal. BC, Alnus glutinosa, markedly decreased, whereas during the following land use phases it remained stable. Climate reconstructions inferred from tree rings, stalagmites, and ice core records suggest moderately drier and warmer conditions during the period 4950−4600 cal. BC (Figure 8; Bircher, 1982, 1986; Isola et al., 2019; Renner, 1982; Vinther et al., 2006), when A. glutinosa collapsed and other wetland plants such as Salix, Cyperaceae, Sparganium, and Pediastrum expanded (see Figures 3 and 8; Giraudi, 2004; Isola et al., 2019; Ramrath et al., 2000; Vinther et al., 2006). This finding suggests that the collapse of A. glutinosa may have been related to lake level changes (Sadori et al., 2004; Sadori, 2018). However, during other periods such as 4400−4200 cal. BC, when climate was even drier according to the Corchia cave mean anomaly index (Figure 8e; Isola et al., 2019), no such dependency between the Lago di Mezzano shore vegetation and climate existed, suggesting that climate may at most have exacerbated human impact on wetland vegetation. Alternatively, lake level decreases may have attracted people to settle on the open shores, promoting the mass disruption of alder carrs. Indeed, low Bphe values suggest oxic conditions and thus low lake level at Lago di Mezzano during this time (4950−4600 cal. BC, Figure 4).
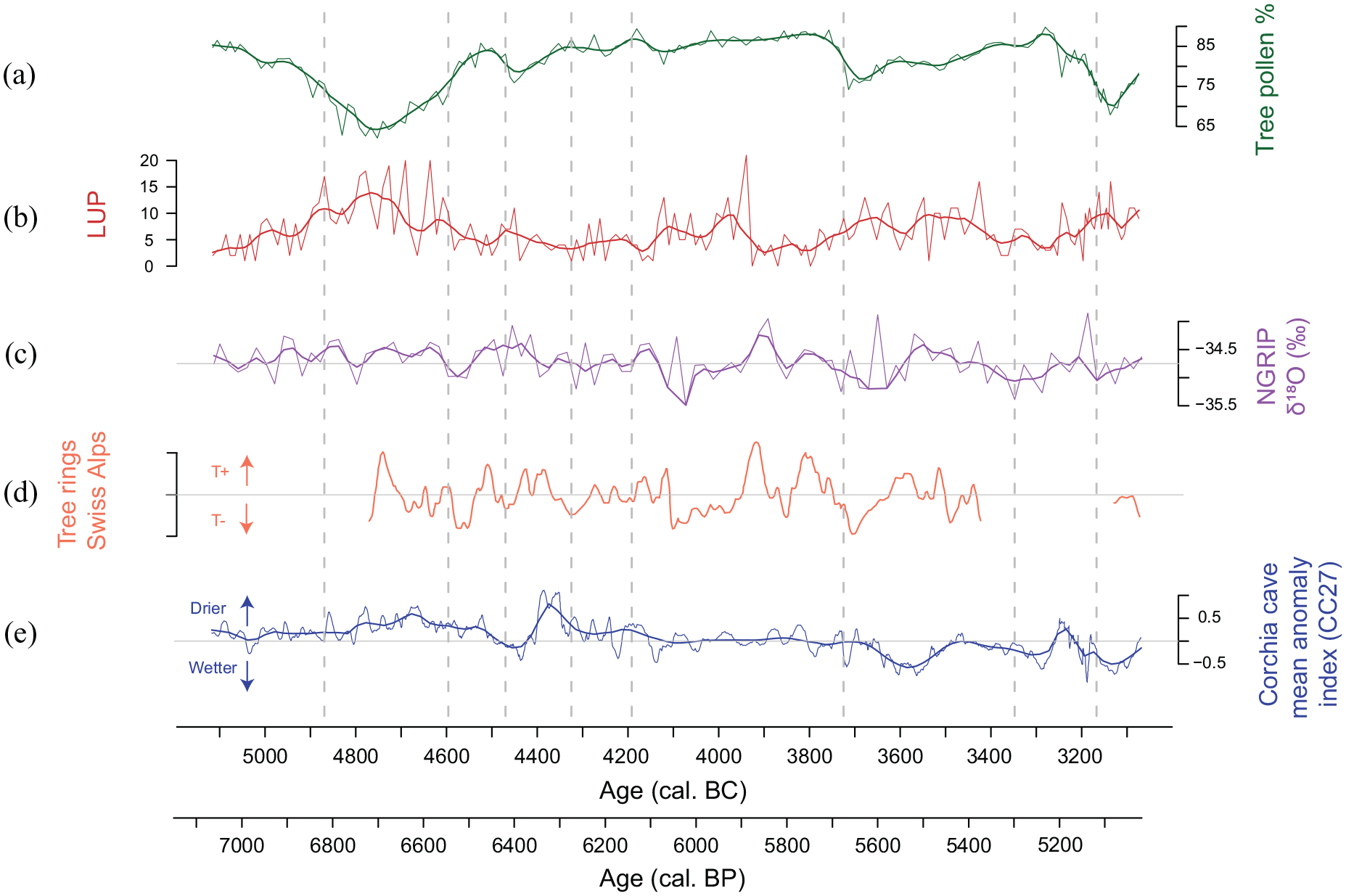
Comparison of tree pollen percentages and the agricultural land use probability index (LUP; Deza-Araujo et al., 2022) from Lago di Mezzano with independent climate reconstructions. (a) Tree pollen percentages with smoothing (loess = 0.05), (b) agricultural land use probability index (LUP) with smoothing (loess = 0.05), c) δ18O NGRIP (Vinther et al., 2006) with smoothing (loess = 0.05), (d) calibrated tree ring data from the Swiss Alps (Bircher, 1982, 1986; Renner, 1982), and (e) mean anomaly index from Corchia cave as an indicator of precipitation variability (Isola et al., 2019) with smoothing (loess = 0.05). Dashed vertical lines: local pollen assemblage zones (LPAZ).
The question to which extent climate may have triggered (contemporaneous) land use phases has been in the focus of archaeological (Gross-Klee and Maise, 1997) and palaeoecological (Tinner et al., 2003) studies for decades (e.g. Dark, 2006; Gajewski et al., 2006; Munoz et al., 2010; Tallavaara and Seppä, 2012; van Geel and Mauquoy, 2010; Woodbridge et al., 2014). Recently, a study based on the comparison of few continuously sampled, high-resolution, and chronologically precise records available from Southern and Central Europe demonstrated contemporaneous Neolithic land use phases south and north of the Alps (Rey et al., 2019a). The high temporal resolution (11.5 ± 2.7 years per sample) and the underlying chronological precision of our new record allow us to compare the dynamics observed at Lago di Mezzano with those observed at other sites with comparable resolution to verify if analogous land use phases extended to central Italy. Our comparison shows a certain analogy in the timing of land use phases (e.g. abundance of crops and weeds) and the structure of the vegetation successions (e.g. from pioneer to late successional species) between the regions of Latium and Ticino (Figure 7), which also extends north of the Alps. Such common phases may be explained by climate impacts on agricultural yields, promoting or reducing the carrying capacity. Thus, periods with favorable climatic conditions for crop production may have led to increased deforestations and burning because human population densities increased, while during periods with adverse climate, human population densities may have declined, leading to repeated afforestation pulses (Maise, 1998; Rey et al., 2019a; Tinner et al., 2005; Wirtz and Lemmen, 2003). Testing such hypotheses would require highly resolved local and regional pollen-independent palaeoclimatic evidence. In the absence of such records, continental-scale evidence might be used. Tree ring records from the Alps show a good agreement with NGRIP oxygen isotope records (Figure 8, Rey et al., 2019a), although the available mid-Holocene tree ring series are still fragmentary. The forest opening periods at Lago di Mezzano at 4950−4600, 4450−4300, 3750−3350, and 3200–3100 cal. BC seem to fall in rather warm periods (Figure 8), but some warm periods such as around 3900 cal. BC have no equivalent opening phases, neither at Lago di Mezzano, nor at Lago di Origlio. On the other hand, our data suggest that during the cold period from 4150 to 3900 cal. BC, although the forest remained closed, farming activities including forest grazing persisted in the Lago di Mezzano area. The Corchia cave record from within the region (Figure 8e; Isola et al., 2019) suggests that the opening phases at 4950−4600 and 4450−4300 cal. BC occurred under slightly drier climates, while the opening phases at 3750−3350 and 3200–3100 cal. BC occurred during slightly moister climates. Because the Corchia record mostly reflects moisture during the autumn and winter seasons but vegetation mostly responds to moisture changes during the growing season (spring to autumn) the comparison is only tentative. Taken together, the available palaeoclimatic evidence is elusive and insufficient to address the hypothesis of climate-driven land use phases in more detail. New pollen-independent and highly resolved palaeoclimatic records are thus needed to assess this issue more carefully.
Comparable land use phases occurred at Lago di Mezzano, Lago di Origlio, and Moossee (Figure 7). Independently from their exact timing, the forest vegetation responses to fire disturbance at the three study sites were similar, as trees declined sharply, thereby providing new habitats for shrubs and grasses (Figure 6). Trees and shrubs that were well represented in all three vegetation types such as Quercus sp., Fraxinus excelsior, and Corylus avellana had common successional patterns in response to fire disturbance at the three sites. This result is rather surprising as the three regions exhibit important vegetational and climatic differences. At all contiguously sampled sites used for comparison in this study, mixed oak and beech or silver fir forests dominated the original Neolithic vegetation, however, while mesomediterranean Quercus ilex was important in the Lago di Mezzano area, it was rare at Lago di Origlio (<1.5%) in Ticino, and absent at Moossee on the Swiss Plateau. Q. ilex is fire-adapted (Pignatti et al., 2017), as frequent fires are characteristic of the Mediterranean region, especially during hot and dry summers. In Ticino, at Lago di Origlio, the submediterranean Insubrian region exhibits far higher levels of summer humidity (with mean summer precipitation of c. 500 mm in southern Ticino versus c. 160 mm in Latium). Therefore, Insubrian (Lago di Origlio) fires have more biomass available, promoting fire intensity and severity (Tinner et al., 1999, 2005). Finally, in the temperate forests of Central Europe, high humidity levels and relatively cool temperatures result in lower fire frequencies (Adolf et al., 2018; Tinner et al., 2005). It is likely that such gradients were present during the entire Holocene and they may thus explain the low fire number (FN; Adolf et al., 2018) at Moossee, intermediate values at Lago di Mezzano, and highest values at Lago di Origlio (Supplemental Figure 3). Such long-term fire-weather and -fuel differences may have affected biodiversity responses to human impact, given that diversity increased in response to intermediate disturbance intensities at Lago di Mezzano and Moossee, whereas it decreased in response to high disturbance intensity at Lago di Origlio (Supplemental Figure 3 and Figure 6). Indeed, the Insubrian mixed oak-silver fir forests exhibited high diversity that was lost due to local extinction of important species in response to excessive burning and grazing that at the end extirpated these formerly important submediterranean forest communities (e.g. Henne et al., 2015; Martinelli et al., 2017; Tinner et al., 1999). Oak and beech forest communities also tended to become less diverse after repeated disturbance but to a far lesser degree (Morales-Molino et al., 2021; Rey et al., 2019a), while more generally, forest openings led to the formation of highly diverse open lands (Colombaroli and Tinner, 2013; Giesecke et al., 2014; Tinner and Ammann, 2005).
Conclusions
This study provides, for the first time, statistically significant evidence of land use practices, with a prominent use of fire, at Lago di Mezzano during the Neolithic. Forest recovery patterns after reduction of human activities at this site in central Italy were similar to those observed at Lago di Origlio, in the warm-temperate submediterranean forests of Southern Europe (Ticino) and to those of Moossee, under the rather cool-temperate conditions of Central Europe (Swiss Plateau). These similarities in vegetation dynamics occurred despite important differences in plant community composition and climate. Moreover, based on chronologically precise and continuous time series, our pioneering Mediterranean study suggests comparable and synchronous land use phases between Southern and Central European sites during the Neolithic. Our study demonstrates that high-resolution palaeoecological reconstructions are essential to better assess vegetation dynamics in response to human impact and climate change. To allow for more detailed spatio-temporal comparisons, further high-resolution and high-precision palaeorecords are needed in the Mediterranean realm.
Supplemental Material
sj-docx-1-hol-10.1177_09596836241247302 – Supplemental material for A novel, continuous high-resolution palaeoecological record from central Italy suggests comparable land-use dynamics in Southern and Central Europe during the Neolithic
Supplemental material, sj-docx-1-hol-10.1177_09596836241247302 for A novel, continuous high-resolution palaeoecological record from central Italy suggests comparable land-use dynamics in Southern and Central Europe during the Neolithic by Giorgia Beffa, Erika Gobet, Luc Hächler, Ilaria Isola, Marina A Morlock, Laura Sadori, Patrick Schläfli, Fabian Rey, Lieveke van Vugt, Hendrik Vogel, Paul D Zander, Giovanni Zanchetta, Martin Grosjean and Willy Tinner in The Holocene
Footnotes
Acknowledgements
We thank the Soprintendenza Archeologia, Belle Arti e Paesaggio per l'area metropolitana di Roma, la provincia di Viterbo e l'Etruria meridionale and the Comune di Valentano for granting us permission to take sediment cores from Lago di Mezzano. Willi Tanner, Erwin Tanner, Peter Ruprecht, Yunuén Temoltzin Loranca, Kathrin Ganz, Yannick Hunziker, and Carolina Senn are gratefully acknowledged for their help during the coring campaigns. We are grateful to Marco Conedera, Boris Vannière, and Christoph Schwörer for inspirational discussions, to César Morales-Molino for help with the pollen analysis, to André F. Lotter for help with core prallelization, and to Sönke Szidat for radiocarbon dating. We thank two anonymous reviewers for the constructive comments.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Swiss National Science Foundation (SNF 200020_182084).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.