
Abstract
Pollen, plant macrofossils and charcoal analyses were used to study tree diversity, fire history and forest disturbance over the past c. 3500 years at three forest remnant sites in Vendsyssel, northern Denmark. All locations had a more diverse tree composition in the past including abundant Alnus, Betula, Corylus, Pinus, Quercus, Salix, Tilia and Ulmus. The changes in tree diversity through time can be attributed to a combination of factors including climate change, burning linked to shifting cultivation, grazing and felling. The balance between arboreal and non-arboreal pollen was already being influenced by human activities in the late Bronze Age c. 3000 years ago. The high pollen abundance values recorded for Tilia pre-2000 years ago are exceptional as compared to later periods at these sites. At one location, the transition from Tilia to Fagus indicated that Tilia prevailed until c. 1300 years ago. Subsequent periods of forest clearance, with charcoal and cereal cultivation, initially including Hordeum and subsequently also Secale, were recorded. There was pollen evidence for grazing followed by shrub regeneration including Calluna, Erica, Juniperus and herbaceous taxa, and following that, a forest recovery of mainly Fagus, Picea and Pinus. This recovery is also recorded in historical forest records from 1880 CE onwards, emphasising the dominant role of plantation schemes. Results are placed in a wider framework of other sites in Denmark and southern Scandinavia, which have also documented a reduction of tree diversity and forest cover over the same period. The evidence from the long-term record is used to draw conclusions to assist forest restoration programmes.
Keywords
Introduction
The composition and dynamics of potential natural forest is a crucial reference for the development of close-to-nature forest types and for adapting forest management practices to accommodate potential climate change impact. Increasing our understanding of long-term dynamic successional processes is also important for effective management of biodiversity, carbon stocks and forest cover as a significant component of the landscape (Willis et al., 2010).
Our understanding of past forest structures and developments generally relies on palaeoecological analyses for the long-term perspective and on historical records and documents for more recent developments. Combined with current knowledge of forest ecology, information derived from such sources provides a basis not only for the management of existing forests, but also for forest restoration and reforestation (Halme et al., 2013; Hennebelle et al., 2018).
Natural forest communities were cleared or modified in most of the temperate zone of Europe after expansion of human populations and the associated shift from hunter-gatherer societies to those depending on agriculture (Roberts et al., 2018). Abies, Carpinus, Fagus and Picea increased their European range more slowly than other major European tree taxa and their range expansions continued with the spread of agriculture in northern Europe (Giesecke et al., 2017). Fagus eventually came to be a leading dominant tree in the European temperate zone (Leuschner and Ellenberg, 2018), yet the palaeoecological record has repeatedly shown that the current dominance by Fagus is so closely associated with the dynamics of agricultural activities that the extent to which the status of Fagus-dominated forest is natural or anthropogenic is open to discussion (Behre, 1988; Björse and Bradshaw, 1998; Küster, 1997; Pott, 2000).
Fagus-dominated woodlands are widespread in Denmark and many are protected because of their biodiversity and presumed natural or semi-natural status (Nord-Larsen et al., 2023). In Denmark, forest cover and composition changed significantly over the past 4000 years, a change from almost complete forest cover in many parts of the country during the Bronze Age, to landscapes which by c. 1800 CE were dominated by cultivated areas, grassland, heathland or patchy woodland (Odgaard and Rasmussen, 2000). Subsequently, continued exploitation of Quercus forest, the ease of natural regeneration of Fagus as well as past priorities in silviculture, all largely supported by forest legislation introduced in the early 1800s CE and continued in present-day legal frameworks, have contributed to Fagus becoming the current dominant broadleaved tree species.
The northernmost limit of Fagus in Denmark coincides with the former Littorina sea shore in Vendsyssel, extending from the base of the Skaw Sand Spit at Frederikshavn northwest to Hirtshals (Figure 1). A number of natural or semi-natural forest remnants dominated by Fagus are located along this stretch, generally growing on marginal glacial till. Fagus stands are also located at similar and more northerly latitudes in Norway and Sweden (Bolte et al., 2007).
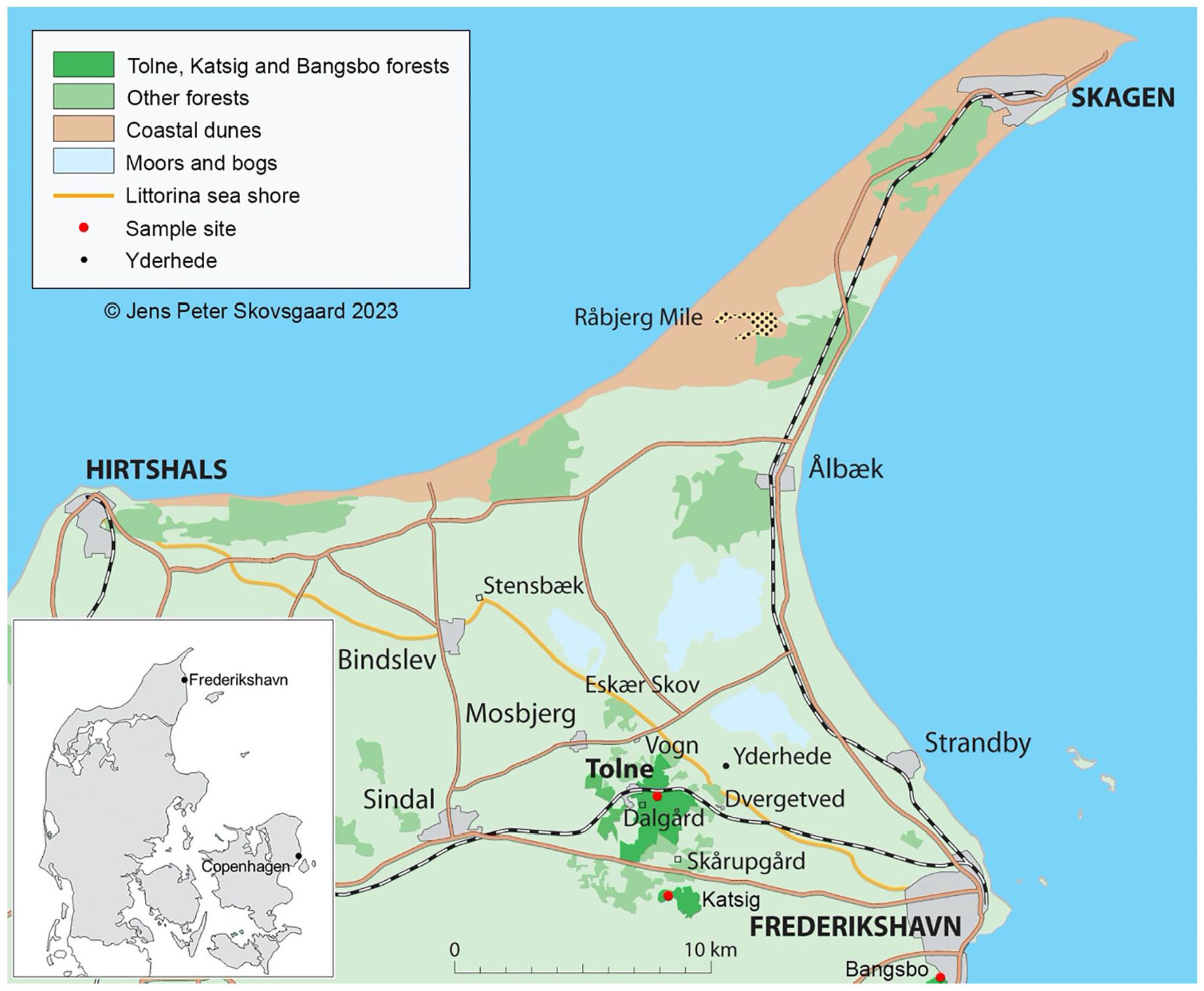
Study area in northern Denmark. Red dots indicate the three woodland sites at Tolne, Bangsbo and Katsig used in the present study; the black dot indicates the location of Yderhede analysed by Christensen and Nielsen (2008).
In this paper, we evaluate the hypothesis that the rise of Fagus to dominance at its northern limit in Denmark is a direct consequence of human mediated disturbance and the associated loss of tree diversity in the European temperate forest zone. We investigate the long-term dynamics of vegetation change in three woodlands located in Vendsyssel, northern Denmark and place the results in the broader context of palaeoecological research and current forest policy initiatives.
Material and methods
Fieldwork
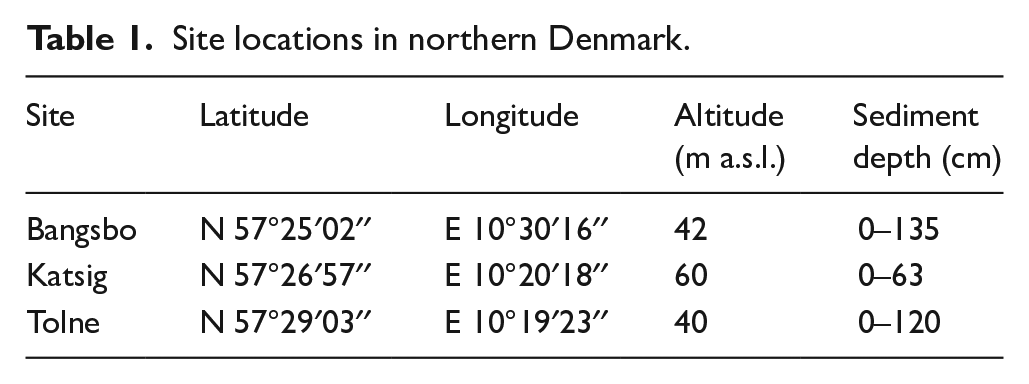
Three small, wet forest hollow sites, filled with peat and detritus, were chosen from within closed canopy woodlands which have mixed forest composition, currently dominated by Fagus (Figure 1). The sites were all less than 20 m in diameter under closed woodland canopies and were chosen to chiefly sample the local, stand-scale vegetation (Jacobson and Bradshaw, 1981). Sediments were collected with a Russian peat corer in May 2018 and November 2019. Site and sample details are shown in Table 1.
Site locations in northern Denmark.
Dating
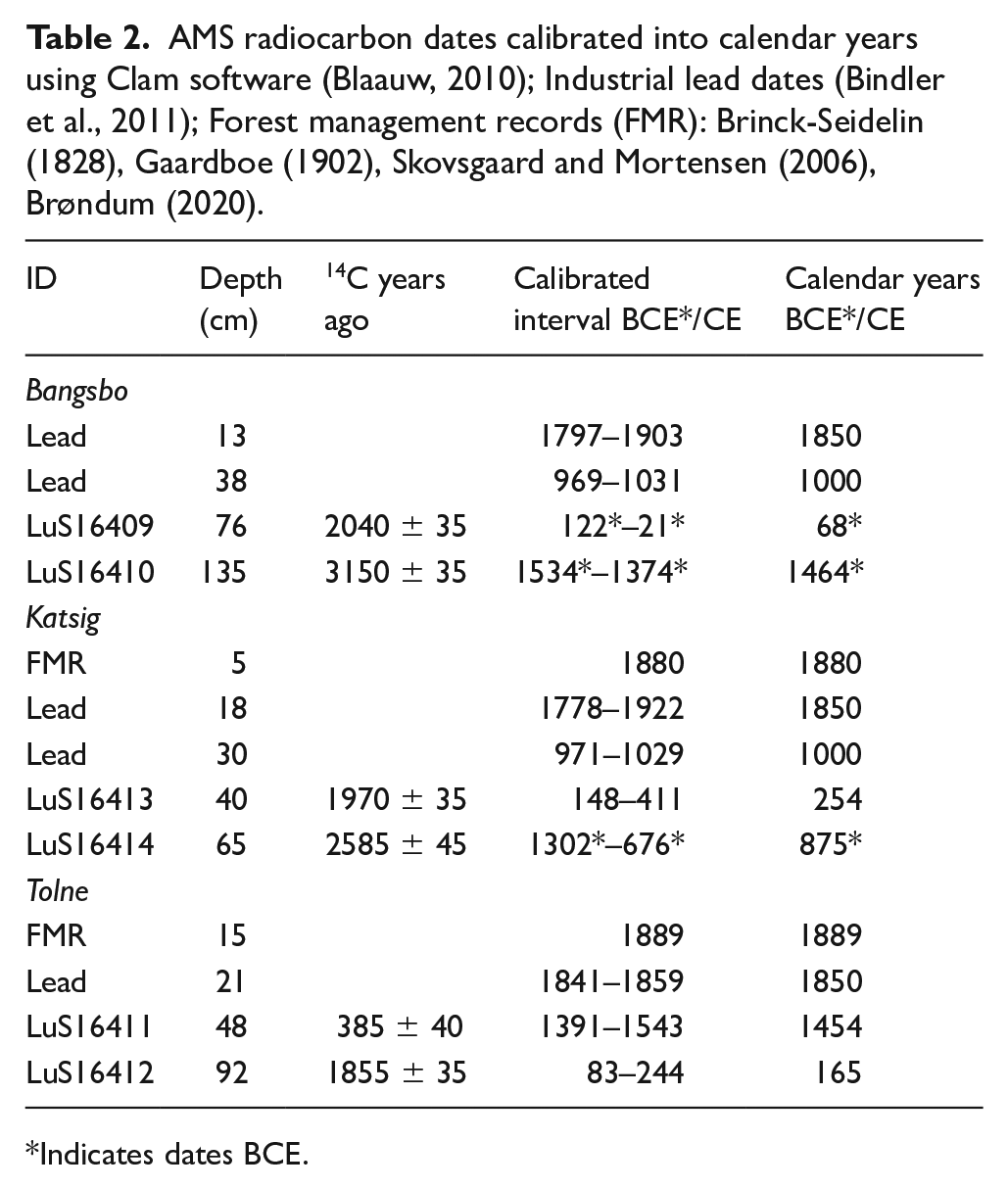
Each sediment core was µXRF-scanned (Olympus Delta ED-XRF), on a wet sediment basis, using a Geotek MSCL-XZ core scanner to obtain the industrial Pb record (Schillereff et al., 2016). This was used for dating purposes together with six bulk sediment samples (weight range 0.8–1.4 mg) from 1 cm horizons, for radiocarbon determination analysed at the AMS dating facility at Lund University (Table 2).
AMS radiocarbon dates calibrated into calendar years using Clam software (Blaauw, 2010); Industrial lead dates (Bindler et al., 2011); Forest management records (FMR): Brinck-Seidelin (1828), Gaardboe (1902), Skovsgaard and Mortensen (2006), Brøndum (2020).
Indicates dates BCE.
Palaeoecological methods
The sediment surface was scraped to remove any possible contamination and 1 cm3 samples were taken at between 2 and 5 cm intervals from a central point. Pollen and spores were prepared using standard methodology and identified using standard keys (Faegri and Iversen, 1989). A minimum of 300 upland terrestrial pollen grains was counted from each level. Pinus and Juniperus stomata were also counted on the pollen slides (Sweeney, 2004). Pollen percentage diagrams were drawn up using the TILIA software, Version 3.0.1 (Grimm, 2015). The pollen sum included trees, shrubs, dwarf shrubs and terrestrial herbs.
Residues collected during the sieving of the 1 cm3 samples (>100 µm) at each pollen level were used for plant macrofossil analyses. Macrofossils were counted and identified using Gina Hannon’s reference collection and Tomlinson (1985) (see Table 1, Supplemental Material). Samples were stored in a cold room at c. 4°C–5°C. The samples for macrocharcoal analysis were gently washed through a 250 µm sieve, stored in double distilled water, washed into petri dishes and viewed under a binocular microscope. Macrocharcoal was counted and entered into a spreadsheet alongside the pollen depths together with microcharcoal data collected from the pollen slides.
Results
Dating
Radiocarbon dates were calibrated to a calendar year timescale for each site. Age-depth relationships for each site were drawn up using Clam software (Blaauw, 2010) (See Figure A Supplemental Material). The dates for the last 1000 years were inferred from the atmospherically deposited Pb profiles recorded in the geochemistry analyses. Pb peak events were correlated with well-dated radiocarbon chronologies of Pb records from nearby lakes (Bindler et al., 2011). The planting dates of introduced conifer taxa (Abies, and Larix, Picea and re-introduced Pinus) from forest management records (FMR) were matched to pollen and macrofossil records from Katsig and Tolne (Brinck-Seidelin, 1828; Brøndum, 2020; Gaardboe, 1902; Skovsgaard and Mortensen, 2006). For Tolne, there were several pollen samples below the depth of the oldest radiocarbon date. Estimated dates for the samples below 92 cm were obtained by extrapolation using the Age/Depth model function and are consequently less secure.
Pollen, spores, plant macrofossils and charcoal
The pollen data from Bangsbo c. 3400−2000 years ago recorded a mixed woodland of Alnus, Betula, Corylus, Pinus, Quercus, Salix, Tilia and Ulmus (Figure 2) with Polypodiaceae (all monolete fern spores lacking an exosporium), Pteridium and diverse herbaceous taxa in the ground vegetation (Figure 2). Pinus pollen was consistently recorded and accompanied by stomata in the lowest part of the period, with charcoal and burnt, crumpled grains. Abundant macrocharcoal was recorded c. 2700 years ago coinciding with a peak in microcharcoal and macrofossils of Alnus, following which there was a 7–8 cm thick wood layer throughout the site and a decrease in Polypodiaceae spores. Major decreases in Tilia and Corylus pollen frequencies with increased Betula, Salix, Calluna, Erica, Cyperaceae, Poaceae, Polypodiaceae and open ground indicators were recorded during the period c. 2000–1100 years ago. This was followed by an increase in Juniperus, Quercus and Fagus pollen towards the end of the period.

Bangsbo. Pollen percentage diagram of (a) major trees, shrubs, Polypodiaceae, sum trees, shrubs and herbaceous taxa and (b) percentage diagram of herbaceous taxa and selected spores. White outlines indicate × 5 exaggeration.
The period c. 1100–200 years ago was characterised by increased values for Calluna, Erica and Cyperaceae with an increase in Betula and Fagus towards the end of the period and a decrease in Salix and Quercus. Juniperus, Alnus, Pinus and Poaceae remained stable. Abundant macrofossil bud bracts of Fagus were recorded (Table 1 Supplemental Material). Hordeum and Secale were continually recorded coincident with a peak of microcharcoal.
The period covering the last c. 200 years was characterised by increasing values of Alnus, Fagus, Picea, Pinus and Quercus pollen. Abies and Larix pollen were first recorded. There was a decrease in many herbaceous open ground indicators (e.g. Brassicaceae and Rumex) and dwarf shrub pollen (e.g. Juniperus and Calluna), while Cyperaceae, Poaceae and Polypodiaceae remained more or less stable (Figure 2). The last Hordeum pollen was from c. 100 years ago although there was a Secale cereale pollen record in the top sample.
At Katsig, the woodland c. 3000–2100 years ago consisted of Alnus, Betula, Corylus, Quercus, Tilia and Salix, with low pollen frequencies recorded for Carpinus, Fagus, Frangula, Fraxinus, Populus, Sorbus and Ulmus (Figure 3). The understorey vegetation contained heathland taxa and Polypodiaceae with a diverse herbaceous flora (Figure 3). Some microcharcoal was recorded.

Katsig. Pollen percentage diagram of (a) major trees, shrubs, Polypodiaceae, sum trees, shrubs and herbaceous taxa. (b) Percentage diagram of herbaceous taxa and selected spores. White outlines indicate × 5 exaggeration. Microcharcoal < 100 µm (cm2/cm3). Macrocharcoal > 250 µm (fragments/cm3). • represents minor types (values under 1%). + indicates presence of a macrofossil. The position of each determined date is indicated by *.
The period c. 2100–1400 years ago marked a transition period where Corylus and Tilia frequencies decreased (Figure 3). Alnus and Betula remained abundant woodland trees, accompanied by macrofossils, and the understorey remained that of Polypodiaceae with herbs and some heathland taxa. An increase in Calluna vulgaris pollen and in the diversity of the herbaceous flora was evident after an increase in the microcharcoal record and there was an increase from 30% to 50% of herbaceous taxa pollen. Hordeum cereal pollen was present (Figure 3). Fagus macrofossils were present from c. 1800 years ago.
During the period c. 1400–1000 years ago, Fagus pollen increased as Corylus and Tilia frequencies declined (Figure 3). Betula, Salix and Polypodiaceae varied little. A small peak in Calluna vulgaris pollen coincided with a peak in macrocharcoal, and sparse but continuous records for pollen of Hordeum and Secale were evident. Betula and Fagus were locally present as macrofossils while Alnus, Quercus and Tilia macrofossils were no longer recorded.
In the period c. 1000–200 years ago an increase in Sphagnum spores, Alnus, Calluna, Cyperaceae, Juniperus and Salix pollen was accompanied by a decrease in Fagus and an increase in microcharcoal (Figure 3). Corylus pollen decreased further while Tilia disappeared. Alnus and Quercus macrofossils were recorded c. 700 years ago.
The final period over the last c. 200 years was dominated by Alnus, Betula and Fagus pollen, supported by abundant macrofossils. Juniperus, heathland taxa and Polypodiaceae records remained stable while the herbaceous flora was less diverse. There was a gradual decrease in microcharcoal. Low frequencies of Carpinus, Frangula, Fraxinus, Picea, Sorbus and Ulmus pollen were observed with occasional Tilia. Pinus pollen were recorded in the most recent sediments.
At Tolne, during the period c. 2700−2000 years ago, the pollen record showed a forest of Alnus, Betula, Corylus, Pinus, Quercus, Salix, Tilia and Ulmus, with low pollen frequencies for Carpinus and Fraxinus (Figure 4). The ground vegetation comprised Juniperus with Polypodiaceae and Pteridium and there were high values of charcoal. The period c. 2000–1000 years ago was defined by a decline in macrocharcoal and, to a lesser extent, microcharcoal (Figure 4). An increase in the frequency of Fagus pollen was accompanied by macrofossil bracts in the sediments. Corylus, Tilia, Ulmus and Juniperus pollen and Polypodiaceae spore frequencies declined, while Quercus and herbaceous taxa increased (Figure 4). Frangula, Rhamnus, Sorbus and Sambucus pollen were recorded. Alnus and Betula pollen values remained the same and the understorey pollen spectra included more heathland plants with an increase in Calluna and Poaceae pollen and in the diversity of the herbaceous flora. The first cereal pollen, Hordeum, was recorded (Figure 4).

Tolne. Pollen percentage diagram of (a) major trees, shrubs, Polypodiaceae, sum trees, shrubs and herbaceous taxa. (b) Percentage diagram of herbaceous taxa and selected spores. White outlines indicate × 5 exaggeration. • represents minor types (values under 1%). Microcharcoal < 100 µm (cm2/cm3). Macrocharcoal > 250µm (fragments/cm3). + indicates presence of a macrofossil. The position of each determined date is indicated by *. Dates below 92 cm are extrapolated using the age/depth model and are consequently less secure (Figure A, Supplemental Material).
The period c. 1000–200 years ago showed a further decline in macrocharcoal while microcharcoal was variable. Tilia almost disappeared and there was an increase in Polypodiaceae, continuous Hordeum pollen and the first record of Secale cereale (Figure 4). Pollen from shrub and heathland taxa including Empetrum, Sambucus and Vaccinium persisted. Alnus, Betula, Corylus, Quercus, Salix and Ulmus pollen values remained stable while Juniperus increased. Herbaceous diversity remained high although there was a decline in Poaceae pollen and an increase in Cyperaceae.
During the last c. 200 years there was a decline in herbaceous taxa, cereal pollen types, Poaceae, Cyperaceae and heathland taxa, and an increase in forest tree pollen, mainly Fagus and Pinus with some Fraxinus, Salix and Sambucus (Figure 4). The first Picea pollen was recorded in this period. There was no macrocharcoal, and microcharcoal was much reduced. Abies and Larix pollen were recorded accompanied by needles. Alnus, Betula, Corylus, Tilia, Quercus and Ulmus were still recorded in the pollen spectra and herb diversity remained high.
Discussion
These are the first investigated forest remnant sites in Vendsyssel, the most northern part of Denmark. Our investigation covers the period from the late Bronze Age, when the landscape was chiefly transformed from a diverse temperate forest cover to more open conditions, until present. Christensen and Nielsen (2008) reported a 500-year record of pollen from a nearby coastal deposit at Yderhede dating to c. 7000 years of age (Figure 1). They described a rich temperate forest that comprised Alnus, Betula, Corylus, Fraxinus, Pinus, Populus, Quercus, Salix, Tilia and Ulmus. That forest possibly had continuity with the temperate forest described in our study from c. 3000 years ago.
Our data document temperate forest during the Bronze Age, followed by periods of more open conditions during the Iron Age and into the Middle Ages with burning, cereal pollen and heathland taxa, and a subsequent re-establishment of scattered woodland during the past c. 150 years (Figures 2–4). Fagus pollen is first recorded in Katsig c. 2500 years ago, in Tolne c. 2000 years ago and in Bangsbo c. 1600 years ago (Figure 5). Fagus in Katsig and Bangsbo had a slower development and populations peaked later than in Tolne. All three sites currently have local dominance of Fagus trees. The pollen records, strongly supported by the Katsig tree macrofossil records (Figure 3), show larger contributions from other tree taxa prior to the Iron Age. This suggests that before intensive anthropogenic activities, the local temperate forest was rather diverse, with intimate mixtures of species. A similar pattern has also been demonstrated by tree macrofossil records from Suserup Forest which is located on more fertile land in the southeast of Denmark (Hannon et al., 2000).
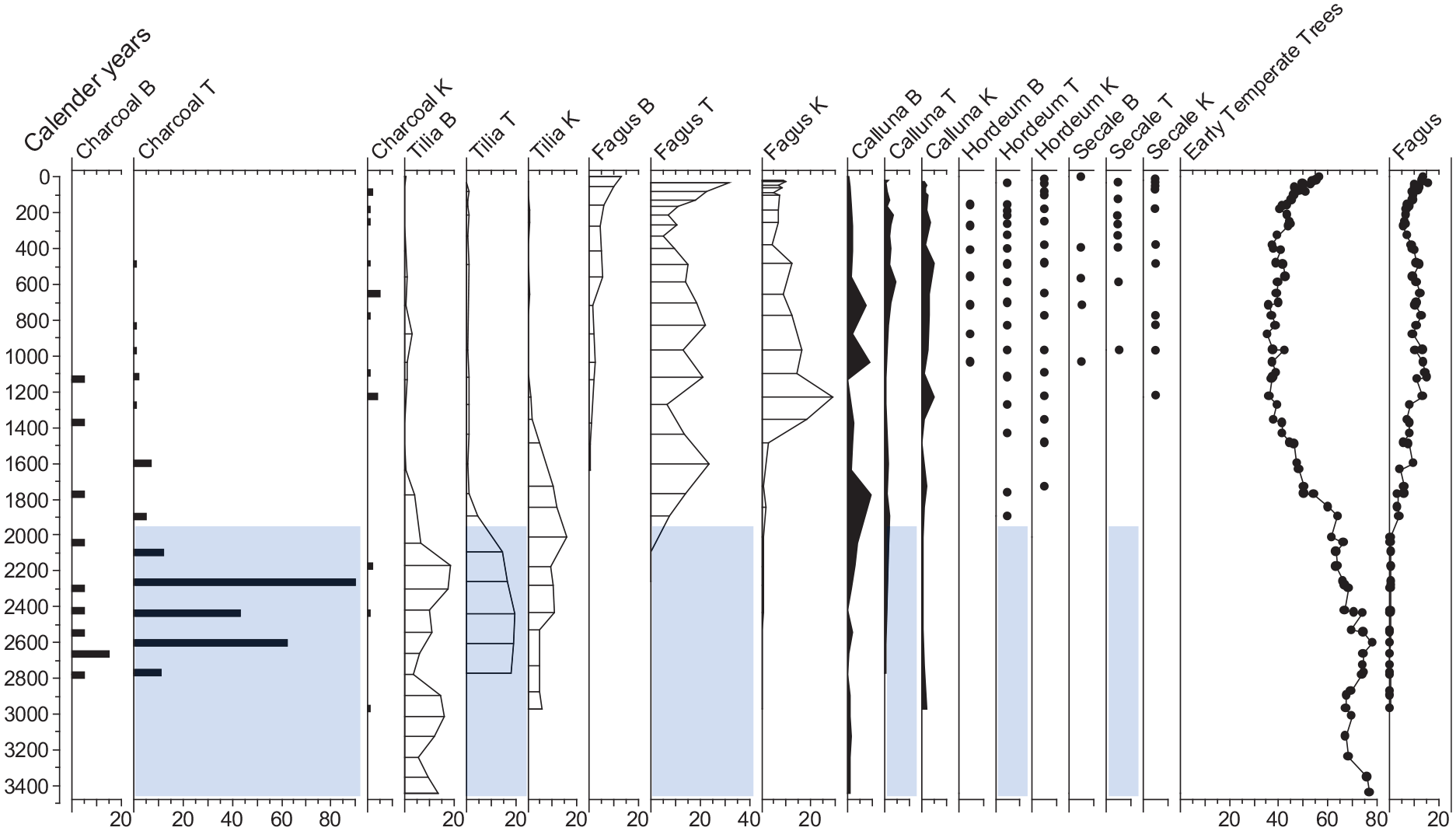
Summary percentage pollen diagram for Tilia, Fagus, Calluna, cereals and macrocharcoal (fragments/cm3 > 250µm). B = Bangsbo, T = Tolne, K = Katsig. Symbols (•) record presence. Temperate forest types include Corylus, Alnus, Tilia, Betula, Quercus, Pinus, Ulmus, Acer, Fraxinus. The shaded sections indicate where the Tolne data were dated by extrapolation of the age/depth model function and the dates are therefore less secure.
A well-dated pollen record from Lille Vildmose, approximately 100 km south of our study area, had a shorter but more detailed record of vegetation dynamics from c. 1300 years ago until present (Yeloff et al., 2007). At this site Alnus, Betula, Fagus and Quercus dominated the tree pollen record, but Tilia was already rare. The Lille Vildmose study recorded periods of forest clearance, cereal cultivation, grazing, temporary abandonment associated with the Black Death in the middle of the 1300s CE and planting of Picea, Pinus and Ulmus during the 1700s CE and 1800s CE, but did not record the earlier transition from rich temperate forest to open conditions or the arrival and initial establishment of Fagus.
Drivers of the transition from mixed deciduous forest to Fagus
The most significant change in the transition from a mixed deciduous forest community in Vendsyssel, northern Denmark, was the replacement of Tilia as a dominant tree by Fagus, with smaller adjustments in the importance of the other tree species from the genera Acer, Alnus, Betula, Corylus, Fraxinus, Quercus, Pinus and Ulmus. This change in forest composition is comparable with those that have been recorded at numerous locations in the temperate forest zone of Europe at various times (Björse and Bradshaw, 1998; Hultberg et al., 2017; Iversen, 1960; Pott, 2000; Turner, 1962).
Our hypothesis, that the rise of Fagus to dominance is a direct consequence of human mediated disturbance and associated reduction in tree diversity, can be evaluated by analysis of the range of potential drivers of this change in forest type at Bangsbo, Katsig and Tolne. These drivers include browsing/grazing, burning, climate change, agricultural cultivation, abandonment, plantation forestry and forest management.
With regard to browsing by ungulates, Tilia is generally preferred over Fagus (Pigott, 2020). Tilia has its recruitment significantly reduced by browsing (Kuijper et al., 2010), while Fagus has been demonstrated to be relatively unpalatable for red and roe deer (Borowski et al., 2021), even though Tilia has a greater capacity than Fagus to regrow vegetatively from the stump or when browsed (Harmer and Howe, 2003). An experimental study found that fire and herbivory were most detrimental to the survival and growth of Tilia in a five-species comparison (without Fagus) (Amsten et al., 2021). In addition, herbivory by cattle and some wild ungulates was more detrimental to Tilia than burning. These species-specific characteristics are consistent with the data presented here, where Tilia populations survived through a period with significant charcoal remains at Bangsbo and Tolne but disappeared from all sites during the first few centuries CE, when there is local pollen evidence for agriculture, including grazing/browsing by domestic livestock (Figures 2–5). The pollen indicators of local grazing/browsing by domestic stock as defined by Mazier et al. (2006) and recorded in this study, include Aster, Lactucaceae, Galium-type, Ranuculaceae, Stellaria-type and Potentilla-type.
Independent evidence for changing browsing pressure is rare prior to the historical period, although archaeological data indicate that settlement expanded steadily in northern Denmark, reaching a temporary climax in the centuries just before and in the beginning of the CE (Jensen, 2001, 2002, 2003, 2004). The related agricultural expansion primarily comprised a blend of domestic stock and cereal crop cultivation, the balance of which depended on soil and landscape characteristics (Nielsen and Odgaard, 2010), as well as on other factors (Jensen, 2003). The widespread but temporary collapse of European agricultural systems in the wake of the Black Death during the mid-1300s CE appears not to have reduced significantly the ungulate browsing pressure, as deer could browse the new shrub vegetation on abandoned land and benefit from decreased competition from domestic animals (Lagerås, 2015). Local historical records from Vendsyssel of domestic livestock grazing within the forest during the period 1683–2006 CE, combined with increasing deer populations (Skovsgaard and Mortensen, 2006), testify to the reduction in species diversity of mixed deciduous tree populations to the advantage of Fagus. This provides further support for the interpretation that grazing and browsing have most likely contributed to the transition from a Tilia-dominated mixed deciduous forest, to Fagus dominance.
Fire is probably a long-term intrinsic property of the temperate forest zone (Drobyshev et al., 2021), but its occurrence in recent millennia has been closely linked with human activities, including shifting cultivation and the maintenance of grazing lands (Bradshaw et al., 2010; Hannon et al., 2018). Both Fagus and Tilia are sensitive to burning, with Fagus being less tolerant of fire damage than Tilia (Bradshaw et al., 2010; Hannon et al., 2022; Kołaczek et al., 2020; Tinner et al., 2005). At the sites presented in this paper, Fagus rose to dominance when burning was gradually discontinued as indicated by low values of macrocharcoal at Bangsbo and Tolne. However, at Katsig macrocharcoal levels are low throughout (Figure 5), possibly reflecting that small forest hollows can be patchy recorders of low intensity fires (Higuera et al., 2005). A statistically significant relationship between Fagus pollen percentage values and macrocharcoal data, based on 18 stand scale pollen sites in southern Scandinavia, indicated that after establishment, Fagus was disadvantaged by local fire (Bradshaw and Lindbladh, 2005). The data from the present study are consistent with that conclusion, so changes in fire regime are a likely contributory factor to the transition from mixed deciduous forest with abundant Tilia to Fagus dominance.
The role of climate change in the rise to dominance of Fagus and the loss of Tilia dominance is complex, but likely to be influenced by changing water availability, late frosts and climate continentality (Bolte et al., 2007; Giesecke et al., 2007; Pigott, 2020). This topic has been explored using climate-driven ecosystem modelling and quantitative statistical analysis, both in southern Denmark (Cowling et al., 2001) and in southern Sweden (Hultberg et al., 2017). The ecosystem simulations in southern Denmark did not support the hypothesis that historical changes in climate were favourable for stand-scale dominance of Fagus. The simulations showed that following the coldest period of the Little Ice Age in Scandinavia between c. 1500 to 1800 CE, which must have severely restricted Tilia regeneration (Pigott, 2012, 2020), Tilia populations should have rapidly re-established their dominance in mixed deciduous woodland (Cowling et al., 2001). In the south of Sweden, the decline of Tilia was most likely driven more by human agricultural activity than climate change (Hultberg et al., 2017).
Iversen (1973) argued that Fagus needed natural or anthropogenic disturbance to spread, because most of Europe was already covered by dense forest within which it was difficult for invading trees to establish (Green, 1987). Subsequent work has indicated the type of anthropogenic disturbance that favoured the spread of Fagus, particularly in northern Europe (Behre, 1988; Bolte et al., 2007; Küster, 1997; Odgaard, 1994). In Denmark, periods of Fagus expansion have been associated with successions following abandonment of cultivated fields (Andersen et al., 1983; Odgaard, 1994). Charcoal pre-dating the arrival of Fagus testifies to the clearance of forest for pastoralism and/or agriculture. The local pollen records show peak Poaceae percentage pollen values together with macrofossils before any pollen evidence of cereal cultivation, indicating initial pastoralism (Figures 2–4). There is pollen evidence for subsequent cultivation of Hordeum and then Secale. Hordeum occurred together with Fagus during early agriculture. This, together with the occurrence of charcoal, indicates shifting cultivation and Fagus successions.
At Tolne and Katsig, the appearance of Hordeum coincided with the decline of Tilia, at Tolne from around 2000 years ago and at Katsig from around 1800 years ago (Figure 5). In contrast, at Bangsbo the decline of Tilia was accompanied by a gradual expansion of heathland, indicating grazing activities that prevailed until as late as around 1100 years ago.
At all three sites Secale first became prominent from around 1000 to 1100 years ago. This agrees with archaeological evidence that Secale spread in Denmark mainly in the wake of Viking activities (Helbæk, 1970). The nearest archaeological record of early Iron Age Hordeum in Vendsyssel is that of a granary stone cellar at the Overbygaard settlement, 75 km south of Tolne dating from c. 2000 years ago (Henriksen and Robinson, 1996; Jensen, 2003). A storage cellar of similar age is located at Dalgård (Figure 1) just 600–700 m southwest of our Tolne site (Kjærum, 1960) and several others have been found in the vicinity of our Bangsbo site (Haue, 2012). Their presence corroborates cultivation of grain at the time. Agricultural pressure increased at all three sites during the Middle Ages, but it was the Iron Age agricultural activities, most likely comprising both cattle grazing, burning and Hordeum cultivation that contributed to the transition from a Tilia-Quercus-Alnus-Corylus-Ulmus forest to the current dominance of Fagus. In summary, this analysis of the drivers of change corroborates our original hypothesis of human mediated disturbance and subsequent loss of tree diversity.
The historical records
Historical documents from Tolne, which record heathland dominated by Calluna vulgaris and Juniperus communis during the 1600s CE coincident with a decline in Fagus pollen (Figure 4), indicate overuse and partial destruction of the forest resource (Brinck-Seidelin, 1828; Skovsgaard and Mortensen, 2006). This exploitation was partially due to grazing within the forest by domestic livestock, a common practice when the first forest inventories were made for the cadastre of 1688 CE and continued in Tolne until the mid-1800s CE (Skovsgaard and Mortensen, 2006). Furthermore, during the AD 1600s CE foreign army troops roamed the area and are known to have cut or burned down forest (Klitgaard, 1938, 1939).
Historical records show that agricultural activities were reduced in Tolne during the last 200 years and a system of forest management was gradually introduced with a consequent increase in Fagus (Skovsgaard and Mortensen, 2006). Regular forest management was also introduced at Bangsbo and, with some delay, at Katsig. Next, Picea and Pinus were introduced into the area by extensive plantation practices (Brinck-Seidelin, 1828; Brøndum, 2020; Gaardboe, 1902; Skovsgaard and Mortensen, 2006). This process was furthered by including periodically subsidised afforestation of heathlands and marginal agricultural land from the late 1800s CE onwards.
Management applications
The European Union (EU) forest strategy for 2030 aims to contribute to ambitious greenhouse gas emission targets and the EU biodiversity strategy. The forest strategy includes strict protection of all primary and old growth forest and the establishment of legally binding nature restoration targets for forests (https://environment.ec.europa.eu/strategy/forest-strategy_en). Pollen, macrofossil and charcoal data can provide a context beyond the comparatively short observational, instrumental, and local records of forest composition, thus providing important insights into longer-term ecological dynamics under various climatic conditions (Brown et al., 2023). Palaeoecological data can contribute to the setting of appropriate targets for close-to-nature management systems and forest restoration programmes (e.g. Hennebelle et al., 2018).
All three sites presented in this paper are from forest conservation areas where protection of biodiversity and carbon storage are major management goals (Brøndum, 2020; Skovsgaard and Mortensen, 2006). All three sites record transitions from rich temperate forest, through a period of open or sparsely wooded vegetation cover, to current Fagus dominance (Figure 5). The open periods were most significant at Bangsbo and Tolne, while Katsig retained most continuity of tree cover. With regard to biodiversity, the pollen records from open periods indicated a greater diversity of herbaceous taxa associated with agricultural activities. These taxa are of reduced importance today and while they represent a significant cultural legacy, they are also indications of former periods of forest exploitation. Saproxylic insects, lichens, bryophytes, and fungi associated with the former rich deciduous forest were likely much reduced because of this agricultural phase, although their former abundance is hard to estimate using palaeoecological methods (Hannon et al., 2018). Referring to biodiversity, a strong argument can be made for restoration of species-rich deciduous forest, when Alnus, Corylus, Quercus, Tilia and Ulmus were the major forest trees (Hannon et al., 2018; Sundberg et al., 2019).
Restoring populations of native tree species is only appropriate if the combination of future climatic conditions, other site factors (including soil and terrain) and expected disturbance regimes are favourable, particularly with regard to browsing animals. Annual average temperature is forecast to increase by 2°C for northern Denmark and annual precipitation by 7% towards the end of this century (www.dmi.dk/klimaatlas). These changes will favour Fagus and Tilia as well as a range of other tree species. These include locally exotic species such as the Central European Acer pseudoplatanus (Hein et al., 2009) and the North American Prunus serotina (Nyssen et al., 2016). Both are abundantly present in the area (Brøndum, 2020; Skovsgaard and Hausager, 2011) and both are highly invasive and competitive. So far, they have not spread significantly into the conservation areas at Bangsbo, Katsig and Tolne, but they are considered a potential threat to these, and to other forest remnants with a composition of chiefly native tree species. With changed future climatic conditions it is appropriate to consider other tree species in addition to restoration of formerly abundant native species (Williams and Jackson, 2007).
Regarding carbon storage, managed plantation forests, particularly conifer plantations, can sequester carbon more rapidly than unmanaged forest conservation areas (Ameray et al., 2021; Skovsgaard et al., 2006). However, carbon storage values are often greater in the biomass of unmanaged old growth forests with very large trees (e.g. Nord-Larsen et al., 2019), although consideration of the total carbon storage capacity must also account for carbon storage in wood products derived from managed forest. Significant quantities of carbon that are released from managed forests following tree removal are retained in the litter, dead wood and soil of unmanaged forest (Naudts et al., 2016). Soil carbon stocks in old growth forest often exceed those in plantations (Liao et al., 2010), although this is not the case in Fagus dominated forests over 100 years of age in Denmark (Nord-Larsen et al., 2019). The Danish soil carbon figures under Fagus are comparable with those measured from managed Fagus sites in Germany (Mund, 2004), but lower than those quoted for unmanaged forest in Germany (Mund, 2004), Bialowieza, Poland (Matuszkiewicz et al., 2021), Lady Park Wood, UK (Hale et al., 2019) and temperate deciduous forest elsewhere (Scharlemann et al., 2014).
The Danish forests studied in this paper, together with Suserup Forest and elsewhere (e.g. Andersen, 1984; Hannon et al., 2000) show how tree diversity has significantly reduced through time related to human activities until this reduction was reversed by the introduction of a wider range of tree species and regular forest management in the 1700–1800s CE. Current data indicate that the carbon storage of biomass, deadwood and soil of species-rich unmanaged deciduous forest generally exceeds that of managed Fagus stands (Matuszkiewicz et al., 2021; Mund, 2004). The unmanaged seminatural deciduous woodland in Europe, with a diversity of tree species, can therefore be a major potential carbon sink that may have been previously underestimated (Allen et al., 2016; Liang et al., 2016).
Conclusions
Our original hypothesis, that the rise of Fagus to dominance at its northern limit in Denmark is a direct consequence of human mediated disturbance and subsequent loss of tree diversity in the European temperate forest one, is corroborated by our research results. Our results suggest that monospecific Fagus sylvatica stands or mixed stands dominated by Fagus sylvatica near its northern limits are an inappropriate reference for the ‘natural’ state of forests as there is strong evidence that these stands have benefitted from anthropogenic disturbance factors and management. How far into Central Europe this conclusion may apply is still under debate (Bolte et al., 2007).
All three sites in this study currently have a local dominance of Fagus. The pollen records from all sites indicate larger contributions from other trees prior to the Iron Age, suggesting that before intensive anthropogenic activities, the local temperate forest was rather diverse with intimate mixtures of species. How present vegetation communities have developed from earlier conditions and what likely future compositions may develop, is of value to managers in planning pragmatic and successful restoration measures. Our results have relevance for management because species-rich deciduous forest supports a number of EU forest policies.
Supplemental Material
sj-docx-1-hol-10.1177_09596836241236340 – Supplemental material for The history of Fagus sylvatica at its northern limit in Vendsyssel, Denmark
Supplemental material, sj-docx-1-hol-10.1177_09596836241236340 for The history of Fagus sylvatica at its northern limit in Vendsyssel, Denmark by Gina E Hannon, Richard HW Bradshaw, Richard C Chiverrell and Jens Peter Skovsgaard in The Holocene
Footnotes
Acknowledgements
We thank Frederikshavn Municipality and the board of directors of Tolne Forest for permission to sample sediments in their forests, Søren Hoff Brøndum of Frederikshavn Municipality for assistance with fieldwork and Michael Vinther of Vendsyssel Historiske Museum in Hjørring for consulting on archaeological issues. The comments of two reviewers improved the quality of the text.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We thank Frederikshavn Municipality, the University of Liverpool, the Swedish University of Agricultural Sciences and the Royal Physiographic Society of Lund for financial support.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.