
Abstract
By the time of the Mid-Holocene Ulmus pollen decline (UD) ca. 5100 14C bp (ca. 5900 cal. BP), the Neolithic was becoming well established in Britain and Ireland. The importance of cereal cultivation as part of the initial neolithization process in the British Isles is uncertain, as archaeological sites of the first Neolithic remain elusive. Palaeoecologists have recorded cereal-type pollen grains in peat deposits that pre-date the UD significantly, but as some wild grasses can produce pollen that closely resembles cereal pollen grains, these early pollen records are not trusted as evidence of cereal cultivation. Some of these wild grass taxa grow in coastal wetland environments, making cereal-type pollen from such locations particularly open to question. This study uses fine-resolution palynology through a sequence of coastal hydroseral deposits that contain no evidence of human activity, to look for the presence of wild grass pollen of cereal size and morphology. Our results show that while such grains are not recorded at 1 cm resolution, at contiguous 2 mm resolution sampling sporadic occurrences of large grass pollen of possible cereal-type, resembling Hordeum, were detected. Morphology suggests that these cereal-type grains are of wild grass origin, almost certainly Glyceria, but their presence suggests that high-resolution analyses of coastal zone sediments will often discover cereal-type grains. Great care must be taken in identifying cereal-type pollen in coastal palaeo-wetland sediments, and rigorous identification protocols should be applied. Where grains could still be of cultivated cereal-type, the presence of other disturbance indicators is an important factor in inferring their origin.
Introduction
The early Neolithic in the British Isles
The earliest Neolithic agricultural communities in Britain might well be archaeologically invisible, or at least difficult to distinguish from the last Mesolithic foragers, particularly if the initial introduction of agricultural practices, products and ideas was slow, piecemeal, selective and on a low scale (Zvelebil and Rowley-Conwy, 1986). Pioneer Neolithic immigrants to Britain might well have farmed within the forest rather than making large clearings (Edwards, 1993; Göransson, 1982). As was the case for a long period in continental Europe (Gron and Sørensen, 2018; Groß and Rothstein, 2023; Perrin, 2003; Ptáková et al., 2023), there could have been a period of overlap and contact of some centuries between the latest Mesolithic and the earliest Neolithic in the sixth millennium cal. BP (Griffiths, 2022; Gron et al., 2018; Warren, 2013; Zvelebil, 1994), although much shorter than in mainland Europe and with little genetic transfer (Mithen, 2022). ‘Terminal Mesolithic’ rod microlith sites in northern England, for example, date right up to the end the Mesolithic-Neolithic transition period and so overlap with Neolithic archaeological sites in the region (Albert et al., 2021; Albert and Innes, 2015; Griffiths, 2014; Spikins, 2002). The respective impacts of the late Mesolithic and the earliest Neolithic on the landscape might have been of a similar scale and difficult to distinguish, but a means of detecting the initial Neolithic might be through changes in human ecology and vegetation disturbance which, although probably subtle and spatially restricted, altered ecosystems (Welinder, 1983) and vegetation patterns (Caseldine and Fyfe, 2006; Woodbridge et al., 2014). Although methodological problems remain, palynological data (Edwards, 1988; Innes and Blackford, 2009, 2017) can potentially provide evidence for the start and nature of these vegetational and palaeoecological changes and it might be possible to interpret early Neolithic pollen data not only in terms of vegetation change at the site scale, but to extend that understanding to the landscape scale and so spatially link the pollen and archaeological records (Farrell et al., 2020).
The first cereal cultivation would be a signature of part of the neolithization process, whether early (Sheridan, 2010), through immigration of pioneer Neolithic settlers or the indigenous adoption of their techniques (Göransson, 1988), or at a later stage when genetic studies (Allentoft et al., 2022; Cassidy et al., 2015) show almost complete replacement of the hunter-gatherers by immigrants. Other Neolithic practices do seem to have been adopted by Late Mesolithic foragers in north Germany and Denmark (Hartz et al., 2002; Kalis et al., 2003; Price and Gebauer, 2005), so cereal adoption in some form is not out of the question. The recording of actual cereal pollen would be an indicator of cereal cultivation, but there are significant difficulties in distinguishing cereal pollen from pollen of some wild grass taxa (Andersen, 1979; Tweddle et al., 2005). The Mid-Holocene decline in Ulmus pollen frequencies (Elm Decline) at ca. 5200–500014C bp (ca. 6100–5900 cal. BP) is a major marker horizon on pollen diagrams in the British Isles and northwest Europe. It was multi-causal (Parker et al., 2002), with disease and climate possible factors (Tipping, 2010), and not a single event (Kearney and Gearey, 2020; Tipping 1995) but it is conventionally held (Bonsall et al., 2002; Kearney and Gearey, 2020; Whittle et al., 2011; Whittle and Cummings, 2007) to have occurred within the early Neolithic cultural period because of its age range and the presence of cereal and other agrarian indicators (Behre, 1981) in its pollen assemblage. By the time of the ‘traditional’ Elm Decline at around 5900 cal. BP in the southern Cumbrian region, which is the study area of this paper, an early farming economy (Richards et al., 2003; Schulting et al., 2004) had become established (Grosvenor et al., 2017; Pennington, 1970, 1997). Large grass pollen grains of cereal size (above 38 μm) and morphology (Andersen, 1979; Joly et al., 2007; Köhler and Lange, 1979; Küster, 1988; Tweddle et al., 2005) are not always recorded in Elm Decline assemblages (Kearney and Gearey, 2020) but those which do occur, both at and after the Elm Decline, can be considered of secure Neolithic age and so have often been routinely assumed to be indicative of actual cereal cultivation, although probably local and at small scale (e.g. Edwards, 1993; Ghilardi and O’Connell, 2013; Göransson, 1986; O’Connell et al., 2014).
Difficulties arise, however, when such grains are recorded before the Elm Decline, in contexts which much archaeological orthodoxy (Behre, 2007) would regard as Late Mesolithic and therefore by definition pre-agriculture. The consensual view has moved towards a relatively abrupt beginning of cereal cultivation in Britain around 4000 cal. BC (Brown, 2007; Rowley-Conwy, 2004, 2011; Rowley-Conwy et al., 2020), based mainly on improved radiocarbon data. It is still conceivable, however, that there was an earlier immigration of pioneer Neolithic groups (Sheridan, 2010) who brought cereals with them some centuries before this date, but have left little evidence. A few dates for Neolithic archaeology, in Ireland at Ballynagilly (Pilcher and Smith, 1979) and the Isle of Man at Billown (Darvill, 1999) are older than 5700 radiocarbon years ago and, if they are accurate age estimates, hint at such early arrival (although the Ballynagilly dates were obtained in the 1970s, prior to recent improvements in the technique, and are substantially earlier than other determinations from the same site). Such very early dates must be treated with caution and should perhaps be redated using more modern techniques, given their potential significance. Alternatively, later Mesolithic groups, presumably via trade or exchange networks (Göransson, 1988; Welinder, 1998), might have adopted cereal cultivation as an addition to their forager resource base (Tinner et al., 2007). This appears to have occurred in the Netherlands where carbonised macrofossil cereal grains have been found in levels with Mesolithic archaeology on a multi-period Mesolithic-Neolithic archaeological site (Meylemans et al., 2018), and perhaps elsewhere (Kalis et al., 2003). Contacts and exchange between farmers and adjacent hunter-gatherer societies were likely as farming spread across northwest Europe (Hartz et al., 2002), leading to elements of acculturation (Fairbairn, 2000).
Pre-Ulmus decline cereal pollen
There are now many published examples of pre-Elm Decline cereal-type pollen from the British Isles (e.g. Edwards, 1988, 1989; Edwards and Hirons, 1984; Groenman-van Waateringe, 1983; Innes et al., 2003a, 2003b; Stolze et al., 2012; Williams, 1985), several of which come from the western coastal areas of Britain and Ireland (Innes et al., 2003b). They are often recorded just as ‘Cerealia’ or ‘cereal-type’, which if correct would be evidence of pre-Elm Decline cultivation by a pioneer Neolithic or by foragers adopting new resources. Some examples are too old to be acceptable as cereals (e.g. Chambers et al., 1988; Day, 1991; Fossitt, 1996; Mighall et al., 2008; O’Connell, 1987; Tweddle et al., 2005) notwithstanding the contested report of very old cereal DNA on the south coast of England (Smith et al., 2015; Weiß et al., 2015). There is, however, a clustering of cereal-type identifications in Britain and Ireland (Innes et al., 2003b) dated between ca. 6000 and ca. 5700 radiocarbon years ago (e.g. Lynch, 1981; Ryan and Blackford, 2009; Williams, 1985; Wiltshire and Edwards, 1993), a plausible age range that might suggest a first pulse of Neolithic cereal cultivation (Williams, 1989). These early grains conform to the morphological criteria for cereal pollen, often for barley (Hordeum) but also sometimes wheat (Triticum), but there are several wild grass taxa that can produce grains which also fit these criteria, such as Agropyron, Glyceria, Elymus and Ammophila (Andersen, 1979; Beug, 2004; Dickson, 1988; Edwards, 1998). Detailed morphological work has made great progress in distinguishing cereal pollen from that of these wetland species (Albert and Innes, 2015, 2020; Küster, 1988; Tweddle et al., 2005), but it has not always been applied to Poaceae pollen, both pre- and post-Elm Decline. All of these wild grasses grow in coastal wetland hydroseral habitats, from saltmarsh to fen-carr and swamp (Rodwell, 1995, 2000), although perhaps Glyceria, primarily G. fluitans but also G. maxima, is the taxon most likely to produce cereal-type pollen (Andersen, 1979; Fitzpatrick, 1946). As several of the published examples of pre-Elm Decline cereal-type pollen in the British Isles (e.g. Cowell and Innes, 1994; Tooley, 1985; Waller et al., 1994) and also in northwest Europe (e.g. Joly and Visset, 2009; Kolstrup, 1988) occur in sediments which formed in Mid-Holocene coastal wetlands, the perimarine zone of Van der Woude (1983, 1985), these examples must be particularly insecure as evidence of cereal cultivation (Edwards et al., 2005; Waller and Grant, 2012) even if they occur within a clear phase of vegetation disturbance that might well represent human activity. The prevalence of large Poaceae pollen grains of cereal-type in Mid-Holocene coastal wetland sediments clearly requires further focused, detailed research, as their significance goes well beyond the individual site and species scale, with the potential to refute or support unorthodox models of the transition to the Neolithic.
The cereal-type pollen problem
As several wild grass types which produce cereal-type pollen are common in coastal wetlands, a question that must be considered is, instead of being so rare, why do large grass pollen grains of cereal-type not appear more often in pollen diagrams from coastal wetland sediments, particularly as most wild grasses are wind pollinated (Hall et al., 1993) unlike the poorly dispersed cereal pollen (Abraham and Kozáková, 2012; Behre and Kučan, 1986; Vuorela, 1973)? If Glyceria habitually produces pollen that is similar to Hordeum, for example, and is common in such wetlands (Boorman and Fuller, 1981; Lambert, 1947), the many pollen diagrams produced for the study of sea-level history and coastal change from sediments deposited in coastal hydroseral wetlands should record large grass pollen of Glyceria/cereal-type far more often than they do. In south Cumbria, for example, Birks (1982) recorded only two Glyceria grains in a pollen profile from a small hollow within Roudsea Wood. The same question applies to the other wild grass taxa that can also produce such pollen, and which are common in coastal wetlands. While the sampling intervals on some pollen diagrams designed to investigate coastal change are often coarse, reducing the chances of encountering large wild grass/cereal pollen grains in the pollen count, those concerned with reconstructing the vegetation history of the coastal zone are often highly detailed (e.g. Innes et al., 2005; Tooley, 1978; Waller et al., 1994, 1999). The general absence of such large wild grass/cereal-type grains in the coastal wetland pollen record suggests that they are either not commonly present or produce very little pollen. Perhaps more likely, however, is that they have not been commonly separated by palynologists from the general Poaceae count, and where this separation has been done on modern samples from fen communities (Waller et al., 2017) a consistent low presence of Glyceria has been observed. A hypothesis is, therefore, that Poaceae pollen grains of cereal size are not rare in coastal pollen sequences, but are under-recorded because of low pollen counting resolution or are unresolved during analysis. This does not mean, however, that Poaceae pollen of cereal-type size should be automatically assumed to have originated from wetland wild grasses rather than cereals, particularly if the cereal-type pollen occurs within an episode of vegetation disturbance with pollen indicators of clearance and agriculture (Williams, 1989), as coastal areas might well have been an attractive location for early cultivation, and also in later periods such as late Bronze Age and Iron Age cereal pollen at Deer Dyke Moss (Coombes et al., 2009), near to Roudsea Wood, and at Foulshaw Moss (Wimble et al., 2000). Although not exclusive to wetlands, prehistoric coastal settlement and exploitation was common in southern Cumbria (Hodgkinson et al., 2000) and adjacent areas (Middleton et al., 1995), with several important sites such as, for example, Ehenside Tarn (Walker, 1966), Eskmeals (Bonsall et al., 1994), Williamson’s Moss (Tipping, 1994), Little Hawes Water (Taylor et al., 1994).
In several studies focused, fine temporal resolution sampling (Green and Dolman, 1988; Turner and Peglar, 1988), has been undertaken to ensure that each counted level represents a very small time interval, perhaps only a few years, ideally with contiguous sampling ensuring that no part of the vegetation history is missed. The technique has provided a better understanding of vegetation changes during the Mesolithic-Neolithic transition (e.g. Garbett, 1981; Innes et al., 2013; Peglar, 1993; Scaife, 1988; Simmons et al., 1989; Sturludottir and Turner, 1985), including the discovery of cereal-type pollen not apparent at coarser resolution (Simmons and Innes, 1996). Most relevantly, this highly detailed scrutiny has been applied by Edwards and McIntosh (1988) and Edwards et al. (1986, 2005) to detect cereal-type grains, and this is the approach that is needed to assess the true prevalence of wild grass pollen of cereal size in sediments of any age, but particularly of the pre-Ulmus Decline millennium. This analysis should be undertaken in coastal wetland sediments which have no sign of human activity in their pollen diagrams, an undisturbed vegetation record in which cereal pollen should not be expected but wild grass pollen should logically occur. If such detailed analysis does not find such large wild grass grains, it could be that the occasional cereal-type pollen recorded in coastal sediment sequences might derive from cereal cultivation after all, perhaps at higher, drier locations within the wetland or at its edge, in situations similar to those described in studies of early Neolithic wetland agriculture in the Netherlands and elsewhere (Cappers and Raemaekers, 2008; Deforce et al., 2013; Out, 2009; Out and Verhoeven, 2014). In this paper we present the results of fine-resolution palynology of sediments formed under the full range of coastal wetland plant communities between ca. 6000 and 5700 14C years ago (ca. 7000–6500 cal. BP), a period when cereal-type pollen grains become recognised in the British and Irish pollen records in any numbers (Innes et al., 2003b). We address two research questions: are large Poaceae grains of wild grass/cereal-type size and morphology recorded more often in coastal sediments when these are subjected to contiguous fine-resolution sampling, and can existing identification protocols separate any such recorded grains into wild grass or cereal types.
The study area and site
The coastal wetland site chosen for fine resolution pollen analysis is in Roudsea Wood Valley, which lies in the southern part of the Roudsea Wood National Nature Reserve near the head of the Leven estuary in south-eastern Cumbria, one of the major rivers that drain into the northern part of Morecambe Bay (Figure 1). The valley is up to 100 m wide and is occupied today by freshwater reedswamp and fen-carr communities dominated by Alnus (alder). It is enclosed by steep carboniferous limestone ridges which today support the long-established, dense mixed deciduous woodland of Roudsea Wood. It is likely that this kind of species-rich woodland, which has been the subject of palynological research by Birks (1982), has occupied these steep ridges since the earlier postglacial, although perhaps modified by human activities at intervals in later prehistory and more recent times (Hodgkinson et al., 2000). There have been several palynological studies in the south-east Cumbria area (Coombes et al., 2009; Dickinson, 1973; Garbett, 1981; Oldfield, 1963; Smith, 1958; Wimble et al., 2000) although the time period of interest to this paper, leading up to the Ulmus Decline, is not always represented. Of most relevance to this study are the data from the raised bog of Ellerside Moss (Oldfield, 1963; Oldfield and Statham, 1963), which is very near to Roudsea Wood, particularly the fine resolution study of Garbett (1981). Birks’ pollen analyses at Roudsea Wood covered the Mid-Holocene period leading up to and including the Ulmus Decline, which was estimated to occur around 5400 radiocarbon bp (4300–4200 cal BP), early but not remarkably so as there are similar early Elm Decline dates from elsewhere in northern England (Bartley et al., 1976). As noted above, Kearney and Gearey (2020) point out that the Elm Decline was often a multi-phase event with an early, primary fall, as also noted by Tipping (1994) at Williamson’s Moss on the southwestern Cumbrian coast. Dates for the Elm Decline might well depend on which phase of the event is being dated, with the feature covering a couple of centuries, as elsewhere in northern England (Griffiths and Gearey, 2017). At his assumed Elm Decline level Birks recorded pollen of Plantago lanceolata and of Triticum-type, presumably reflecting early Neolithic agriculture in the nearby catchment. Before that level, however, Birks recorded unbroken forest cover in the wood, with no evidence of disturbance by humans, although this core was from a small hollow within the forest, and the pollen recruitment area might well have been limited (Bunting, 2008; Waller et al., 2005). Garbett’s fine resolution study found some weed pollen increases that could be slight evidence of disturbance in pre-Elm Decline levels, but no large (>38 µm) grass pollen grains, itself perhaps strange given the depositional environment.
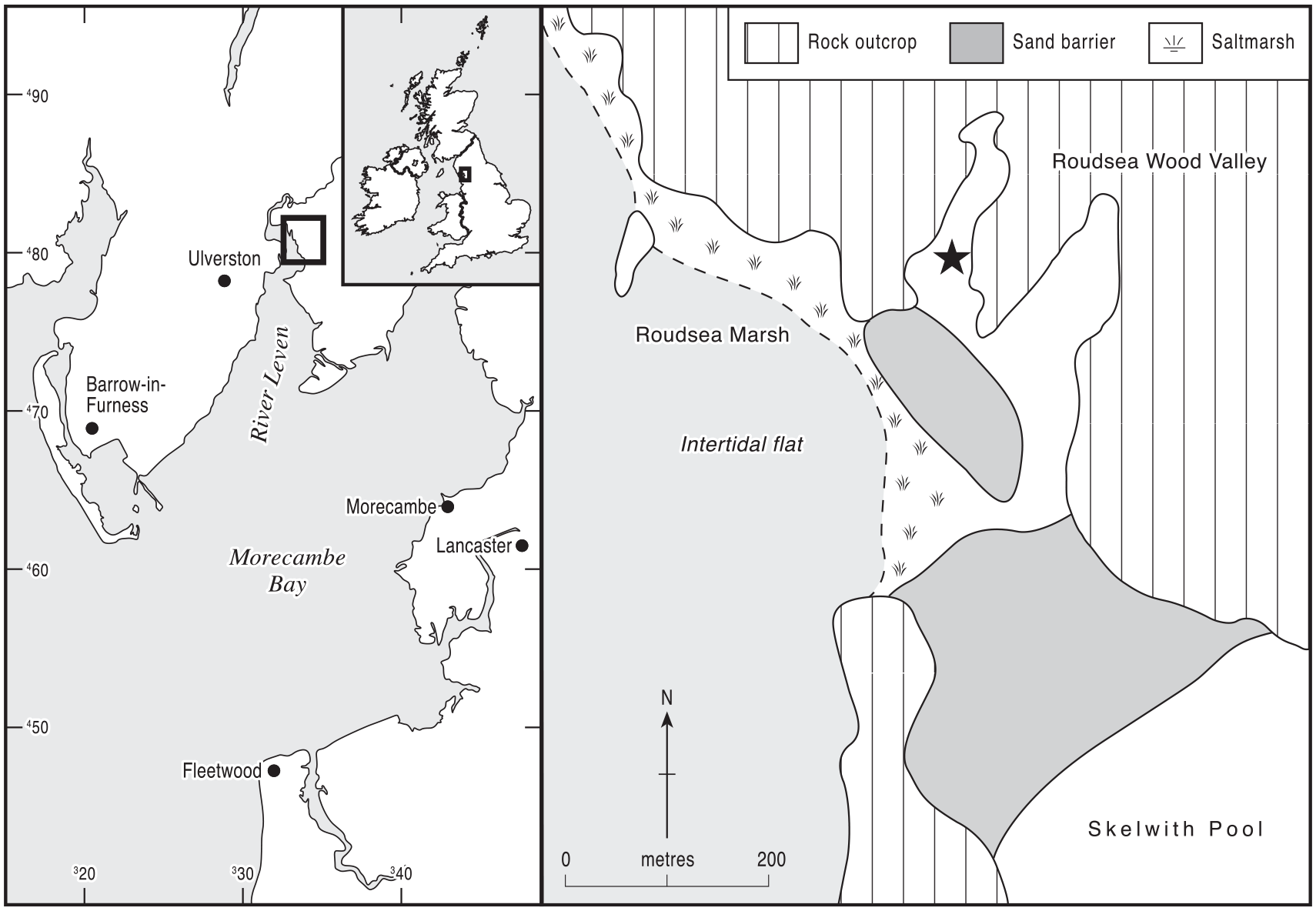
Location of the sampling site at core 33 (SD33198163), in the Roudsea Wood Valley in Cumbria, northern England. The site is shown by a black star.
As part of a study of sea-level history, Zong (1998) completed a transect of cores along Roudsea Wood Valley from the estuary’s coastal saltmarsh to the valley head, consistently finding 3–4 m of Mid- to Late-Holocene sediment. Lithostratigraphic and diatom analyses at core RW33 in the centre of the valley at UK grid reference SD33198163: lat. 54°13ʹ34ʺN, long. 3°1ʹ35ʺW (Tooley et al., 1997; Zong, 1998; Zong and Tooley, 1999) showed that its lower sediment fill comprised a basal freshwater peat overlain by marine silts and clays and so had been subject to inundation during the Mid-Holocene eustatic rise in sea level on this coast (Tooley, 1982; Zong, 1997; Zong and Tooley, 1996). The uppermost 1.5 m of sediments in the valley, however, were almost entirely organic and represented deposition within mainly freshwater environments, shown by pollen and sedimentary analyses to have been aquatic, reedswamp and fen-carr communities similar to those existing at the present time. Halophytic diatoms, in contrast to their dominance in the lower clastic deposits, were virtually absent in the upper organics, except for small peaks of polyhalobous types that probably reflect storm surge events (Tooley et al., 1997; Zong and Tooley, 1999). The switch from estuarine to freshwater deposition at RW33, shown by the replacement of polyhalobous and mesohalobous diatom species by oligohalobous species (Zong and Tooley, 1999), occurred at 1.56 m depth (+3.43 m O.D.) and was dated to 6030 ± 70 14C bp (Tooley et al., 1997) (7200–6725 cal BP). It probably occurred because of a slowing of sea-level rise at this time, increased organic sedimentation and the establishment of a sand barrier between the rock outcrops that enclose the valley. The creation of this barrier would have almost separated the valley from the estuary and the intertidal zone (Figure 1), as it does today, although it is clear from the pollen record at RW33 (Tooley et al., 1997) that saltmarsh communities must have been present close by, at least at the valley entrance, and there is a thin organic silt layer just above the highest counted pollen level that indicates a short-lived, final penetration of intertidal conditions to the site. Tooley et al. obtained further radiocarbon dates in the organic sequence above 6030 ± 70 14C bp (7200–6725 cal BP), and their pollen record continued up to 5740 ± 60 14C bp (6720–6450 cal BP).
The site of Roudsea Wood 33 therefore contains all of the elements required for testing the prevalence of large Poaceae pollen grains that might be identified as cereal type in coastal wetlands during the earlier stages of the Mesolithic-Neolithic transition: a series of three radiocarbon dates from c. 6000 to c. 5700 14C bp (7000–6500 cal BP) and evidence of the full range of coastal zone depositional environments from saltmarsh through reedswamp, fen and carr woodland to back-barrier lagoonal environments. Between them, these coastal hydroseral habitats (Walker, 1970) would have contained all of the wild Poaceae species (Rodwell, 1995, 2000) which can produce cereal-like pollen grains. The lack of any significant evidence for opening of the forest cover or human interference with the local vegetation during the pre-Elm Decline period (Birks, 1982; Garbett, 1981) suggests that any large Poaceae pollen found can probably be attributed to wild, non-cultivated grasses growing within the coastal and valley hydrosere.
Materials and methods
The part of the Roudsea Wood 33 peat profile between 120 and 170 cm was originally sub-sampled for pollen analysis at 1 cm intervals, and the results for selected taxa have been published previously by Tooley et al. (1997) and Zong and Tooley (1999), as core RW33A. The opportunity is taken in this paper to publish the full pollen diagram from RW33A (Figure 2). Most of the sediment from the radiocarbon-dated levels in the original core had been taken for the bulk peat radiometric analyses, and so a duplicate core which was collected at the same time as RW33A, termed RW33B, was used for the present paper. It was located very closely adjacent to core RW33A, only a few tens of centimetres from it. The two cores could therefore be laterally correlated exactly, confirmed by the matching depths of the contacts of their lithostratigraphic units which lie at the same altitudes relative to sea level and so form time-equivalence horizons. In this duplicate core RW33B, the peat between 120 and 136 cm was selected for fine resolution palynology and was sub-sampled at contiguous 2 mm intervals. This section of the core lay between the radiocarbon dates 5990 ± 70 (c.6845 ± 157 cal. BP) and 5740 ± 60 BP (c.6515 ± 145 cal. BP) and was chosen for close scrutiny because, as discussed above, many examples of pre-Ulmus Decline Poaceae pollen grains of cereal type have been dated elsewhere to that time interval (Innes et al., 2003b), particularly in the coastal lowlands around the Irish Sea.

Coarse resolution pollen diagram from core Roudsea Wood 33A calculated as percentages of total tree pollen, sampling at 1 cm intervals. Numbered lithostratigraphic units are described in Table 1 and shown in Figure 3, using the symbols and notation of Troels-Smith (1955). Radiocarbon dates and calibration age-ranges BP are shown.
Samples were prepared for palynological analysis using standard laboratory techniques, with alkali digestion, sieving at 180 µm, hydrofluoric acid and acetolysis (Moore et al., 1991). Pollen nomenclature follows Moore et al. (1991). Pollen residues were stained with safranin, mounted on microscope slides in silicone oil and counted at x400 magnification, with higher magnifications used for critical features. In both the coarse and fine resolution pollen diagrams a total tree pollen sum was used, as is appropriate for Mid-Holocene forested environments (Berglund and Ralska-Jasiewiczowa, 1986). Total land pollen counts, which included trees, shrubs and herbs, always exceeding 500 grains. Aquatics and spores are calculated as percentages of the tree pollen sum. Not counted at RW33A, microcharcoal particles were counted on the new RW33B analysis as a pollen/charcoal ratio (Robinson, 1984), with microcharcoal calculated as a percentage of the pollen sum. Particles that passed through the 180 μm pollen preparation sieve were regarded as microscopic and were recorded relative to the pollen count, with those around 30 μm in diameter regarded as the basic measurement unit (Innes and Simmons, 2000) as that is similar to the average size for pollen grains. Counts comprise multiples of that unit, with the size of individual fragments recorded relative to that unit so that a piece of 90 μm would be counted as three units. All pieces smaller than 30 μm were aggregated to produce countable data. Separate curves for different size ranges are not produced as fragmentation of particles will occur during the laboratory preparation (Clark, 1984).
For this new study, non-pollen palynomorphs (NPPs) were also counted on the pollen slides, as Clarke (1994) has shown that the great majority of fungal spores and other NPPs are largely unaffected by the preparation procedure. NPP analysis is restricted to the new, fine resolution diagram, and is also calculated as percentages of tree pollen. NPPs were identified using the illustrations and descriptions in manuals and in published papers (Miola, 2012; van Geel, 1978, 1986, 2001; van Geel and Aptroot, 2006). At least 100 NPPs were counted at every level, and those which could not be identified to taxon were recorded using the standard ‘Type’ numbering system, which employs the prefix ‘HdV’, of the Hugo de Vries Laboratory catalogue at the University of Amsterdam in The Netherlands. Known NPP taxa have their type number shown after their name on the diagrams and in their first mention in the text. The lithostratigraphy of the full core RW33A using the Troels-Smith (1955) system of notation was recorded in Zong and Tooley (1999), and the section of relevance to this paper is shown in Table 1. The microfossil diagrams were constructed using the TILIA programme of Grimm (1993, 2004). No new radiocarbon dates were needed for this new study, which relies upon the dates published by Zong (1998) from the identical duplicate core RW33A.
Lithostratigraphy of the section of the Roudsea Wood 33 core selected for palynology, divided into stratigraphic units.

Notation follows Troels-Smith (1955). A thin organic silt layer occurs between 117 and 120 cm, above the levels counted for pollen. The lithostratigraphy of the full core is recorded in Zong and Tooley (1999). Its ground altitude is +4.99 m O.D.
Results
Lithostratigraphy and radiocarbon dating
The lithology of the duplicate core RW33B is the same as that of the original core, being so very close to it, and it is divided into numbered lithostratigraphic units on Table 1. These unit numbers are shown on Figure 3, which is an environmental conspectus diagram, and on the pollen diagrams. The radiocarbon dates relevant to this paper are radiometric dates from Beta-Analytic, Miami on bulk peat samples, and their details are shown in Table 2, including measured radiocarbon age, calibrated age ranges at the 2δ level and mean calibrated age. They form a chronological series and are used to create an age-depth model curve (Reimer et al. 2020) for the original core RW33A, calculated using the classical age-depth model CLAM v.2.4.0 (Blaauw, 2010). A 4th order smooth-spline model was used, with 1000 model iterations and a goodness of fit of 4.38. Age-depth model data are shown in Supplemental File Table S1, available online. The modelled age-depth data can be applied to core RW33B as it is a duplicate with identical depths. Interpolated ages at the millimetre level can only be estimations and are derived from the modelled ages at the centimetre level.
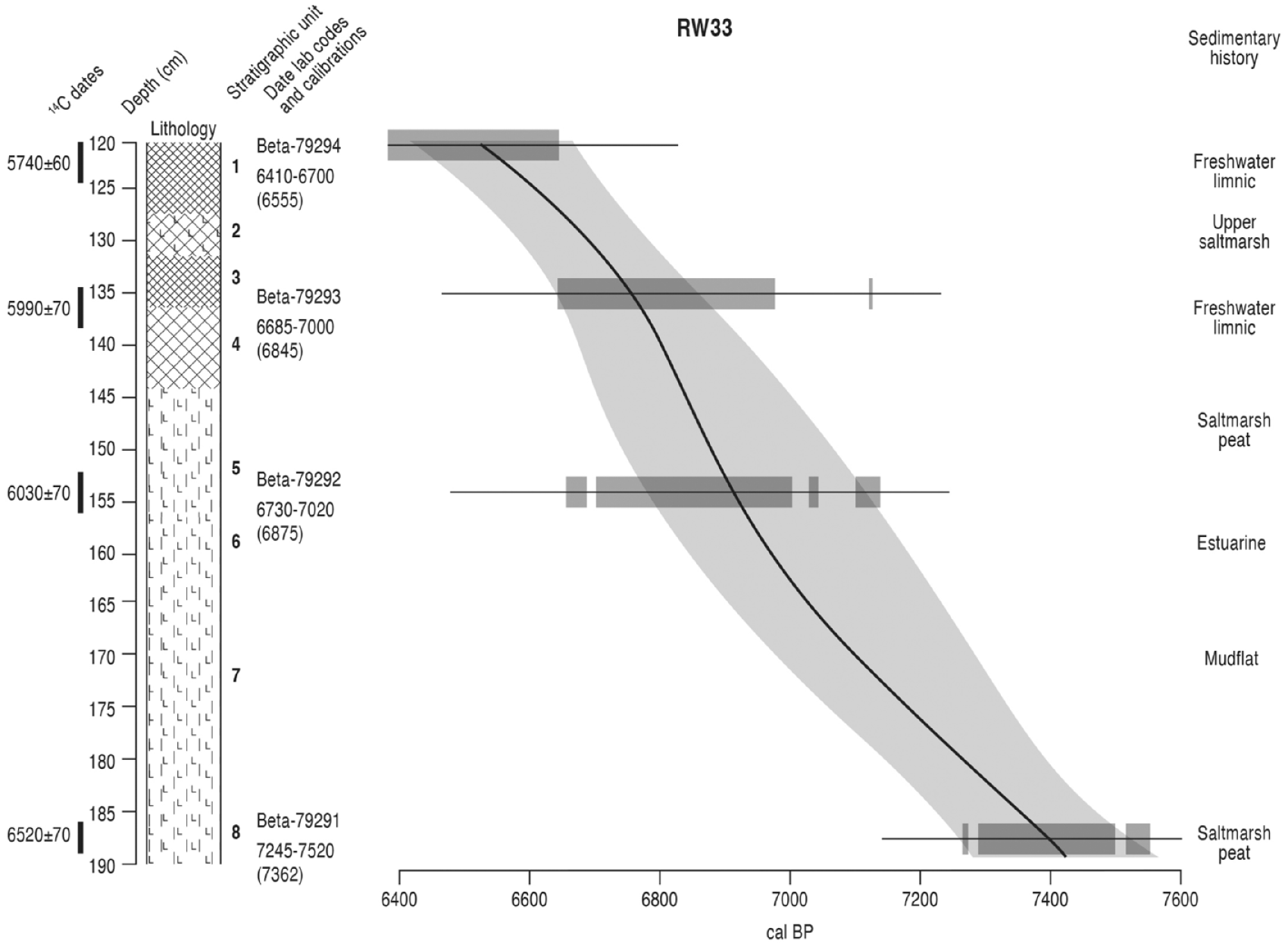
Conspectus of lithostratigraphy, Classical age-depth model, radiocarbon date information and sedimentary history at Roudsea Wood 33. Lithology symbols follow Troels-Smith (1955). The stratigraphic units are numbered, applied to the other diagrams and are described in Table 1 using the notation of Troels-Smith (1955). For each dated depth the calibrated age range (BP) is shown with the mean age in parentheses. Output from Clam v.2.4.0 (Blaauw, 2010), showing modelled age against depth. Age models use a 4th order smooth-spline model and IntCal20 calibration curve (Reimer et al., 2020). The shaded areas represent the 95% probability range confidence interval. Radiocarbon date details are shown in Table 2 and full age-model data are given in the Electronic Supplemental Material File S1.
Results of radiocarbon dating from Roudsea Wood 33, Cumbria (Tooley et al., 1997; Zong, 1998) that are used in this paper.
Palynology
The results of the palynological analyses at Roudsea Wood are shown in Figures 2, 4 and 5. The full, 1 cm resolution pollen diagram for Roudsea Wood 33A is shown in Figure 2, and is divided into eight local pollen assemblage zones which are described in Table 3, with their modelled age ranges cal. BP. The fine-resolution pollen diagram from Roudsea Wood 33B is shown in Figure 4 and extends between c. 6793 and c. 6550 cal. BP. It covers zones f, g and h from RW33A and is subdivided into seven zones, described in Table 4, which are also applied to the NPP diagram (Figure 5).
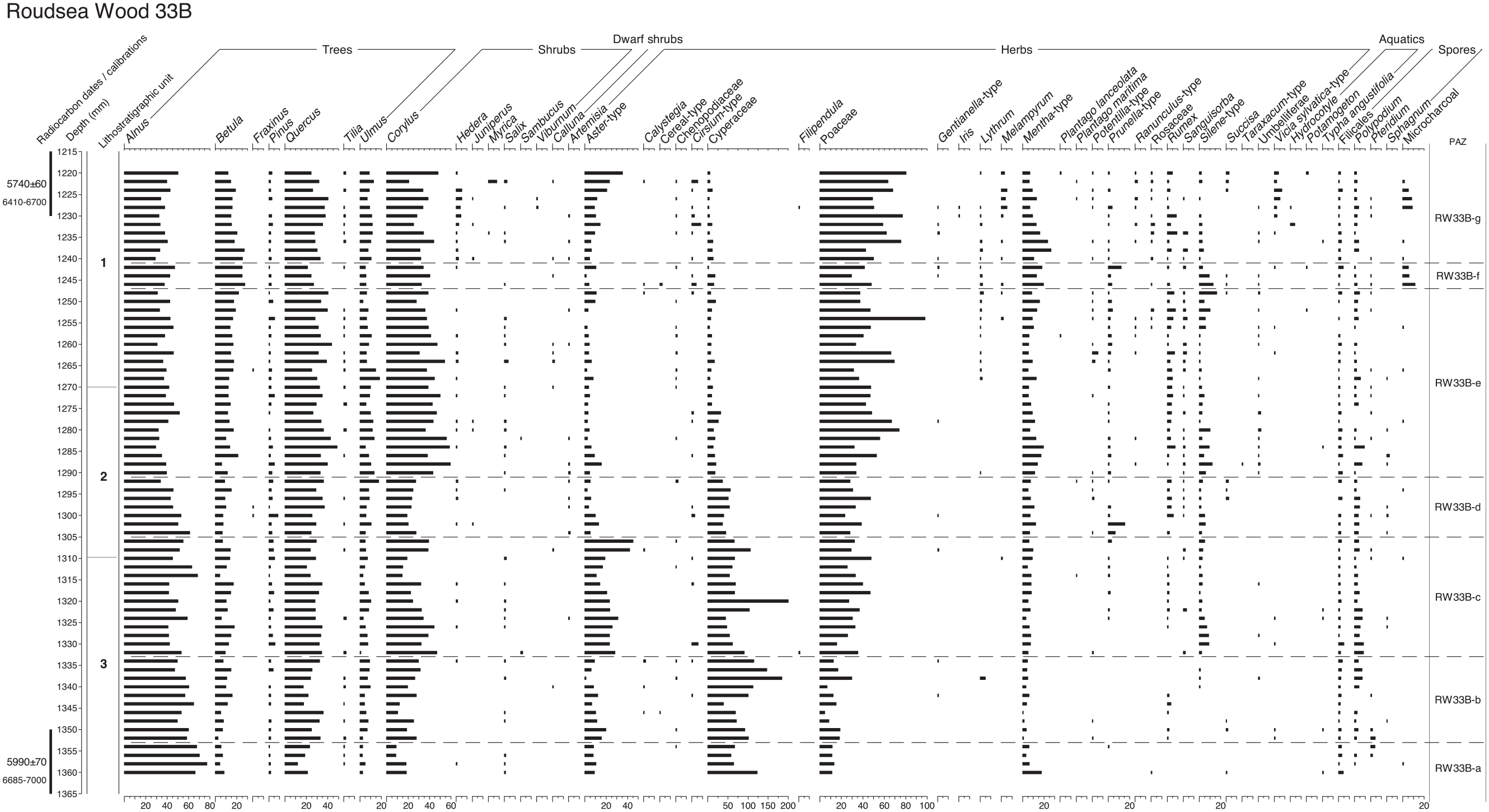
Fine resolution pollen, spore and micro-charcoal diagram through the Mesolithic-Neolithic transition levels at Roudsea Wood 33B, calculated as percentages of total tree pollen, sampling at contiguous 2 mm intervals. Numbered lithostratigraphic units are described in Table 1 and shown in Figure 3, using the symbols and notation of Troels-Smith (1955). Radiocarbon dates and calibration age-ranges BP are shown. This diagram covers zones f, g and h at RW33A (Figure 2).
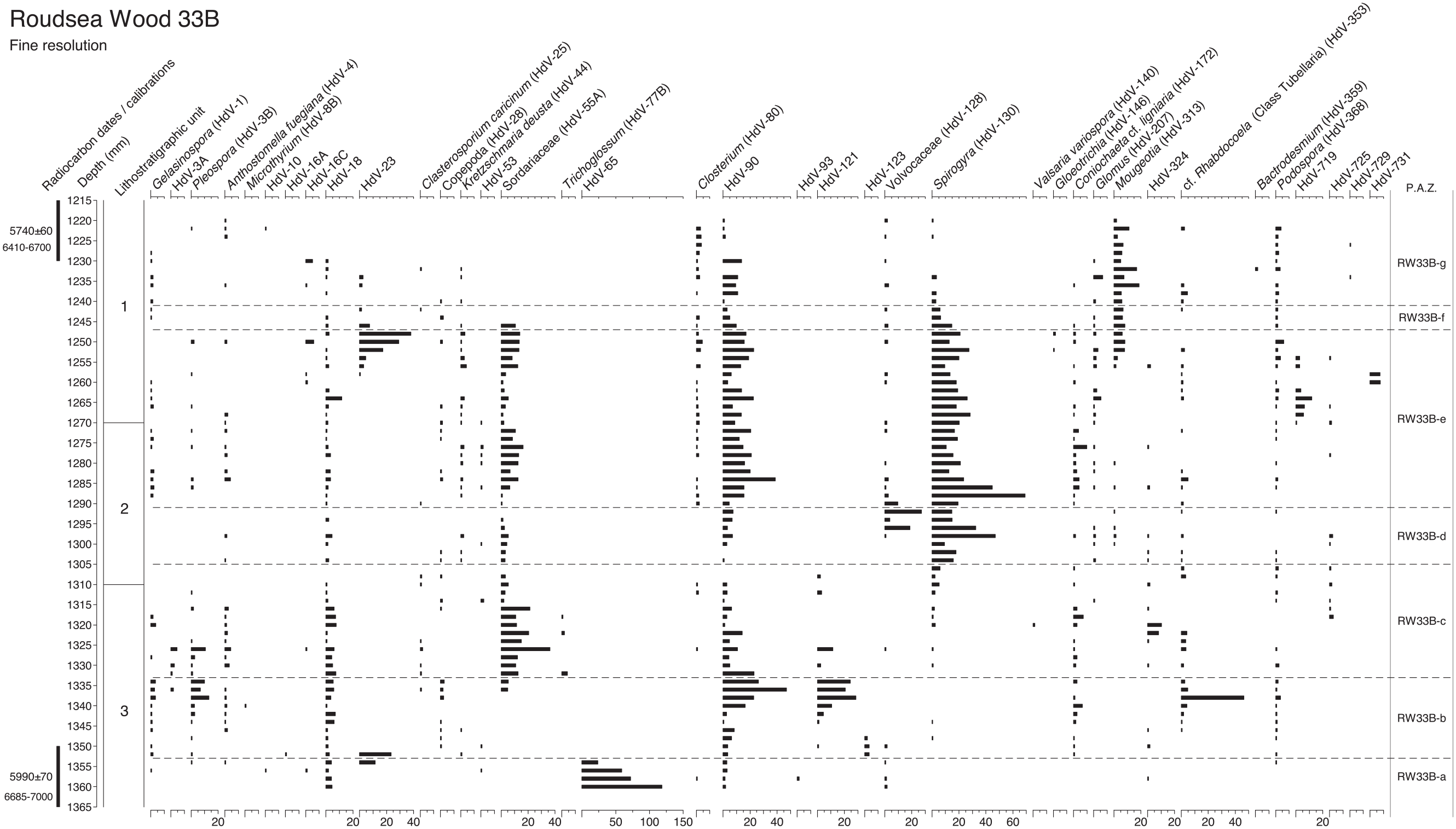
Fine resolution NPP diagram through the Mesolithic-Neolithic transition levels at Roudsea Wood 33B, calculated as percentages of total tree pollen, sampling at contiguous 2 mm intervals. Numbered lithostratigraphic units are described in Table 1 and shown in Figure 3, using the symbols and notation of Troels-Smith (1955). Radiocarbon dates and calibration age-ranges BP are shown. Zones are those used in Figure 4.
Pollen assemblage zone (PAZ) descriptions from Roudsea Wood 33A, including modelled ages from Supplemental File Table S1.


Although no pollen grains of cereal type were recorded in the full, 1 cm resolution pollen diagram at Roudsea Wood, at the fine resolution, contiguous 2 mm counted level four pollen grains were observed which exceeded 38 μm in length, the accepted size above which they should be investigated as being of possible cereal-type (Andersen, 1979; Küster, 1988; Tweddle et al., 2005), although Hordeum can often be below this size threshold. Albert and Innes (2020) have shown that the maximum long diameter of Glyceria maxima always lies below this 38 μm level, and so cannot be confused with cereals other than Hordeum. The Roudsea Wood grains all lay between 38 and 41 μm, and so are not G. maxima. Their other details are shown in Table 5. Close scrutiny of these grains, based upon the criteria established in the previous publications and tested in detail for Hordeum and Glyceria by Albert and Innes (2020), shows that all four grains fall into the Glyceria fluitans category rather than G. maxima or any cereal taxon. Grain long diameter alone is insufficient to distinguish G. fluitans from cereals as individual grains can be of either greater or lesser length than the 38 μm threshold. The maximum diameter of annuli and pores, and the gradient of the annulus rise, is of most importance for identification (Albert and Innes, 2020) and while the annulus diameter for Hordeum is between 8 and 10 μm, any greater diameter excludes Hordeum. As the fossil grains at Roudsea Wood all have annuli equal to or greater than 12 μm (12, 15, 15 and 17 μm), they cannot be Hordeum. The Triticum/Avena group grains have pore diameters greater than 4 μm whereas the fossil grains’ pore diameters do not exceed 4 μm (all 3 or 4) and so are disqualified from that cereal grouping. The cereal-size Poaceae from Roudsea Wood must therefore be assigned to a wild grass origin rather than to any cultivated grass (cereal) type, almost certainly Glyceria because of their annulus diameter and to G. fluitans because of the annulus and pore data and flat annulus rises, as indicated by Albert and Innes (2020).
Details of the four cereal-type pollen recorded in the Roudsea Wood fine resolution data (Figure 4).
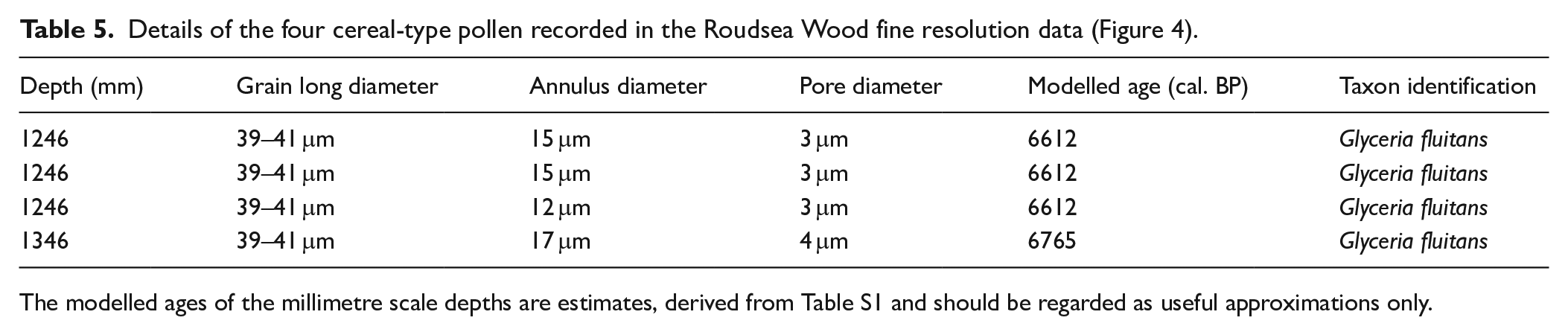
The modelled ages of the millimetre scale depths are estimates, derived from Table S1 and should be regarded as useful approximations only.
Discussion
Vegetation history at Roudsea Wood
The modelled ages of the main core from Roudsea Wood show that the vegetation record extends between approximately 7100 and 6500 cal. BP, firmly within the Mid-Holocene mixed-oak forest maximum between the rise of Alnus pollen frequencies and the decline of Ulmus values that define the start and end of that period of English Holocene vegetation history (Birks, 1989; Smith and Pilcher, 1973). The pollen assemblages in Table 3 are dominated by Quercus and Alnus with lesser Ulmus and Corylus, and so conform with the general pre-Ulmus decline woodland pollen record, with a relatively closed-canopy deciduous tree cover. In south-east Cumbria at this time, however, the several available pollen diagrams indicate a high degree of diversity, the proportions of the components of the forest differing considerably according to environmental factors, even within the Roudsea Wood area. High Fraxinus and Tilia percentages are recorded inland in the area on more calcareous soils by Oldfield and Statham (1963), Dickinson (1973) and Pigott and Huntley (1980), and Birks (1982) on limestone within the Roudsea Wood forest itself. In contrast, those two calcicole trees are barely represented in the Roudsea Wood 33A pollen diagram, perhaps because of its coastal valley location on organic soils and a dominantly local tree pollen source area. Fen oakwoods at the landward wetland fringes might well have contributed much of the tree pollen rain, with Quercus dominant during the earlier part of the record (Figure 3), together with Corylus which will represent hazel woods on the drier limestone slopes around the valley. The less well transported pollen of any Tilia and Fraxinus trees growing on these slopes might well have been filtered out by the screening oak and alder woods around the wetland.
The decline of oak and hazel at about 141 cm depth at the start of zone RW33-e (modelled as 6830 cal. BP) and the expansion of Alnus frequencies will mark hydrological changes within the mire and at its margins, with a switch from a reedswamp wetland to a sedge fen and alder swamp-carr environment at around the same time as Poaceae declines to be replaced by Cyperaceae. The high Poaceae frequencies through most of the diagram can be referred to Phragmites, and thus to reedswamp, because of the consistent presence of Phragmites macrofossils in the sediment column. This might account for the very low numbers of Glyceria pollen grains (cf. cereal-type) recorded in the pollen diagrams (none in Figure 2 and only four in Figure 4), as Phragmites will outcompete and suppress Glyceria (Buttery et al., 1965; Buttery and Lambert, 1965; Lambert, 1947), forcing Glyceria to marginal locations against the fringing Salix and Alnus carr, which it is unable to penetrate (Westlake, 1966). Glyceria grains (Table 5) occur mainly near the top of Figure 4 when Poaceae dominance was reduced, and increased sedimentation reduced water levels.
It is clear that the vegetation history recorded at Roudsea Wood was governed greatly by local hydrological changes which altered wetland plant communities, including the depth of freshwater systems as well as the introduction and withdrawal of marine/estuarine influence by secular movements in sea level, as shown by diatom data (Zong, 1997, 1998) but also by probable storm surge events (Tooley et al., 1997; Zong and Tooley, 1999). The reciprocal nature of Quercus and Alnus curves in coastal wetlands has long been recognised (Godwin and Clifford, 1938) as reflecting fluctuations in the proportions of carr and fenwood at the landward margins of the coastal freshwater hydrosere (Binney et al., 2005). Freshwater depth changes and fluctuation between reedswamp, fen and deeper water habitats are apparent in Figure 2, with Typha peaks in zones RW33-b and RW33-d recording pool formation, interleaved with interludes of greater estuarine influence as shown by saltmarsh taxa such as Chenopodiaceae, Aster-type and Plantago maritima. In Figures 4 and 5 in the fine resolution levels for the later part of the pollen record it is clear that a very complex suite of wetland habitats existed in close proximity, with the coring site located on the estuarine/perimarine ecotone. Variations in the frequency curves for the aquatic algal spores of Volvocaceae (HdV-128), Mougeotia (HdV-313) and Spirogyra (HdV-130) support this evidence of high freshwater tables, but fungal spores including Pleospora (HdV-3B), Sordariaceae (HdV-55) and Kretzschmaria deusta (HdV-44), which are not transported far from their point of origin, indicate terrestrial vegetation close to the site. The fluctuating curves for saltmarsh taxa, particularly Aster-type, during the same period as the freshwater algae indicate a spatially diverse palimpsest of communities, perhaps with saltmarsh creeks penetrating reedswamp, fen and carr communities. The different behaviours of the Cyperaceae and Poaceae curves between Figures 2 and 4, in closely adjacent sequences, is a good example of the spatial complexity of the wetland vegetation across very small distances and the importance of very local plant cover.
Central to the investigations of this paper, however, is the record of disturbance or open dry ground, which is virtually absent from the centimetre-resolution diagram (Figure 2), which is the reason this profile was chosen for closer scrutiny. There are no dryland weeds and all herbs can be referred to wetland types within their genus. The sporadic Pteridium curve probably represents growth along the margin of the fenwood. The 2 mm resolution diagram, however, having five times as many levels across the upper part of the profile, has some evidence of vegetation disturbance in these levels that was not apparent at the coarser sampling interval. For most of the diagram there are no indications of disturbance, microcharcoal being hardly present, similar to the findings of Garbett (1981) with only very occasional small fragments. However in zone RW33B-f Quercus falls sharply, replaced by peaks of Alnus and Betula, and three cereal-type pollen grains occur at the start of the zone at 1246 mm depth, at a level where microcharcoal frequencies are significant for the first time, although still in moderate values. This level approximates to c.6612 cal. BP, derived from the calibrations at the cm level in Table S1. There are no ruderal herbs, although there are peaks in Cirsium-type and Silene-type, herbs which could be species of disturbed woodland habitats although they could as easily be members of the coastal wetland community. This phase could be interpreted as small-scale burning within the local oakwoods, with cereal cultivation at the start then regeneration through alder and birch until restoration of oak dominance. Table 5, however, shows that all three grains are identifiable as Glyceria fluitans rather than Hordeum, the possible cereal type. The appearance of the Glyceria pollen might have been aided by improved transmission to the site as fire opened local shrubby wet woodland and reduced its pollen filtering effect. There was a small burning episode in the oak fenwood, but it was not associated with cereal cultivation.
Of the taxa that are often used as disturbance indicators, there are increased Melampyrum percentages in zone RW33B-g between 1228 and 1224 mm. This period approximates to 6581–6574 cal. BP, interpolated from Table S1. Although this rise in Melampyrum could as easily represent changes in mire hydrology (Moore et al., 1986), the presence of microcharcoal in these levels suggests burning, to which Melampyrum responds (Delarze et al., 1992; Innes et al., 2013; Simmons and Innes, 1996), and as the microcharcoal particles are large the fire might have been close to the site. Tree pollen frequencies are unaffected however and there are no ruderal weeds in the assemblage. The small burning episode might well have been within local fen-reedswamp vegetation, as recorded at the margins of other wetland sites in northern England (Law, 1998). It is impossible to tell whether the burning was caused by natural or human ignition, although natural fire in wetland vegetation is rare and Mesolithic people might well have been responsible for starting fire at wetland margins as part of their land use strategy (Innes et al., 2010, 2013; Simmons, 1996).
There are no indicators of woodland opening in the rest of the pollen record, with only two grains of Plantago lanceolata, a dryland indicator of open ground, in the whole diagram, neither of which coincide with the large Poaceae (cereal type) pollen grains or the burning events. It is clear from Figure 4 that while there was some small-scale burning in the area around the Roudsea Wood core on two occasions, estimated from the cm modelling (Table S1) as occurring around c.6580 and c.6612, in only one of the episodes is there a disturbance context within which to place the cereal-type grains. It might seem reasonable to ascribe the burning events to human agency, as natural fire within wetland environments during the climatically damp Mid-Holocene in northern England (Hughes et al., 2000) seems unlikely, but that remains speculation.
Hordeum-type pollen grains in pre-Elm decline sediments
Very early examples of Cereal-type pollen have been recorded in northern Europe, often as Hordeum or just as Cerealia, mostly based on size alone, and these have been regarded as almost certainly originating from wild grasses (Clark et al., 1989; Hörnberg et al., 2006; Kalis et al., 2003). Similarly, several records of pre-Elm Decline pollen grains in the British Isles have been dismissed as spurious as they occur well before 6000 radiocarbon years bp (7000 cal BP), including Hordeum type referred to Glyceria (Brown et al., 2014), and so well before any realistic introduction of cereal cultivation from Europe, whether by trade or migration (O’Connell, 1987). This presents a dilemma of interpretation. If the same types of evidence are interpreted inconsistently due to the inferred age of the sediment, the interpretation of the later phases must also be brought into doubt. The interpretation and identification becomes subjective, and depending on expectation, rather than being objectively classified due to the characteristics of the grains in question. Those large grass grains recorded as occurring after approximately 6000 radiocarbon bp (7000 cal BP), however, in both Europe and the British Isles, do deserve greater scrutiny, especially those that have been recorded as of Hordeum type, because the possibility of proximity to Neolithic influence is much higher.
Ideally, objective criteria can be used to rule out or in the possibility of cultivated Poaceae being present in any horizon, but this can only be applied from now on, and not to previous studies which are up to 50 years old. These are of greater relevance to this paper as the large grass grains recorded at Roudsea Wood were all of Hordeum type when first recorded. As Hordeum was one of the most common types of cereal cultivated in the British early Neolithic (Bishop et al., 2013), closer attention is warranted. Two particular time periods are of interest. The first is after about 5400 radiocarbon bp (6300 cal BP), as some Elm Decline dates occur at this early time (Parker et al., 2002), particularly in the lowlands (e.g. Bartley et al., 1976) but also at altitude (Bartley, 1975) and the earliest Neolithic archaeology begins at about this date (Hedges et al., 1994), unless early dates at Ballynagilly are accepted. The pollen grains identified by size and morphology (Albert and Innes, 2020) as Hordeum at Esklets on the North York Moors (Albert and Innes, 2015), for example, fall into this time frame and so despite being almost half a millennium before their local upland Elm Decline might possibly be acceptable, as they are contemporary with the earliest dates for some lowland Elm Declines, during the early stages of the Mesolithic-Neolithic transition in northern England. Other Hordeum type grains occur in this time period, centuries before their local Elm Declines, as in near-coastal fen peat at Machrie Moor (Robinson and Dickson, 1982) in western Scotland.
More contentious are the cereal pollen grains identified as of Hordeum-type in the earlier period when cereal-type grains begin to be recorded in pollen diagrams in northern and western Europe, including the British Isles, between 6000 and 5700 radiocarbon bp (7000–6500 cal BP) (Innes et al., 2003b). Lynch (1981) recorded Hordeum type grains at Cashelkeelty in southwest Ireland. Some studies have applied morphology criteria to cereal-type grains, primarily Hordeum, from the beginning of this period in Britain (Albert et al., 2021) and northern Europe (Alenius et al., 2017; Poska and Saarse, 2006; Wieckowska et al., 2012) and concluded that they do represent Hordeum rather than any wild grass, presumably procured by hunter-gatherers during an ‘availability phase’ (Zvelebil and Rowley-Conwy, 1984) before the advent of the Neolithic, or by small groups of pioneer settlers with at least a partly ‘Neolithic’ resource base. As all of these studies separated Hordeum from Glyceria on morphological criteria, it seems that such records from this early time period must be accepted, or at least given serious consideration, even in these more peripheral areas of Europe.
Conclusions
This study confirms the expectation that increasing the sampling resolution of the pollen analysis, either by increasing the pollen count or, as at Roudsea Wood, greatly reducing the interval between counted levels, increases the chances of encountering rare pollen types which are poorly transported or produced in low numbers. Contiguous fine resolution samples at millimetre scales will ensure that no pollen data will be missed. Some of these pollen types will include taxa which might yield important ecological information or indicate a cultural presence. Such fine temporal resolution sampling, in which no temporal gaps are left in the pollen stratigraphy, must always improve the chances of recovering diagnostic information. In the case of pollen sequences from coastal wetland sediments, it is very probable that fine resolution analyses and high pollen counts will produce large Poaceae grains that need evaluation as either of wild grass or cereal origin.
The earliest large Poaceae (Hordeum-type) grain from Roudsea Wood occurs at 1346 mm depth, in levels without any other indications of vegetation disturbance. This find suggests that pollen assemblage composition and thus the plant community context of the grains is an important second indicator of their likely taxonomy, after their physical characteristics, but before timescale. ‘Real’ cereal pollen grains, from either cultivated, transported or self-seeded cereal plants, given their poor transport ability, are unlikely to occur in deposits with no supporting microfossil evidence of vegetation disturbance or human activity, and so these earlier grains could be expected to be almost certainly of wild grass origin, as morphological data then showed them to be. The later cereal-type grains at 1246 mm depth, in contrast, occur within a phase with indications of disturbance, which might encourage their identification as more likely to be of cereal (Hordeum) type. That close scrutiny of quantified characteristics has shown them to originate from a wild grass taxon demonstrates that associated vegetation disturbance, although perhaps making it more likely, is not a reliable indicator that cereal-size Poaceae are actually cereals, and hence the interpretation of previously published examples where the pore and annular diameters were not measured could be problematic.
Deposits formed within the coastal hydrosere are good habitats for the type of wild grasses that can produce pollen grains easily confused with cereal types. Where they are still available, pollen grains recorded as Cerealia or cereal-type in previously published diagrams, and especially those from coastal locations, should be re-examined and re-evaluated where possible, particularly with a view to separating Glyceria in particular from other coastal grasses and possible cereals. This would only be possible where grains were mounted in silicone fluid and well preserved, rather than in glycerine jelly in which grains might well have swollen over time.
More rigorous criteria need to be applied for the identification of all fossil cereal-type large Poaceae grains, in general but particularly in coastal wetlands, in all cultural periods and not just for the Mesolithic-Neolithic Transition when cereal cultivation might be considered to have been less likely. There is much evidence that prehistoric agricultural societies, from the Early Neolithic onwards, occupied and made economic use of wetlands (Coles and Lawson, 1987; Lillie and Ellis, 2007; Menotti, 2012). While nutrient and resource poor acidic mires and bogs were less utilised, lowland wetlands such as lake edges, reedswamps and fens, as at Roudsea Wood, were particularly favoured because of their higher nutrient status and consequently greater wild plant and animal resources. Their exploitation by agriculturalists probably included cereal cultivation in favourable drier locations within the wetland, such as islands, wetland edges and dry peat surfaces, as occurred in Dutch deltaic wetlands (Cappers and Raemaekers, 2008; Deforce et al., 2013; Louwe Kooijmans, 1987, 1993; Out, 2009; Out and Verhoeven, 2014) and in many British examples, as discussed by Van de Noort and O’Sullivan (2006), such as the many small islands that rise above the wetland surface in the Somerset Levels (Coles et al., 1973). Despite the several wild grass taxa, and particularly Glyceria, that produce large pollen grains in such environments, therefore, it should not be assumed that any large grass grains encountered in pollen analyses of coastal wetland sequences will not be of cereal type. Nor should it be presumed that such large Poaceae grains are more likely to be of cereals during later prehistoric cultures which used cereal cultivation more extensively. Each identification of such potential cereal grains should be made on its own merits using the available, accepted measurement criteria.
The new pollen identification protocols (Albert and Innes, 2020) used in this paper are an important step forward in the assessment of large cereal-type Poaceae grains, and will be a useful tool for eliminating as cereals many large Poaceae grains at present being recorded as ‘cereal-type’, particularly in contexts where alternative wild grass taxa are common, such as coastal wetlands. Using pollen grains that pass the above protocols alone as an indicator of cereal cultivation in coastal wetlands continues to carry a degree of uncertainty, however, particularly as to the location and scale of the cultural activity and sites (Farrell et al., 2020) which were linked to the cultivation. It might be that, in the absence of cereal macrofossils, pollen indications of early Neolithic cereal cultivation in coastal wetlands must remain preliminary, until used in combination with new diagnostic techniques, such as, for example, sedaDNA (e.g. Hudson et al., 2022, 2023).
Supplemental Material
sj-doc-1-hol-10.1177_09596836231219461 – Supplemental material for Testing the presence of cereal-type pollen grains in coastal pre-Elm Decline peat deposits: Fine-resolution palynology at Roudsea Wood, Cumbria, UK
Supplemental material, sj-doc-1-hol-10.1177_09596836231219461 for Testing the presence of cereal-type pollen grains in coastal pre-Elm Decline peat deposits: Fine-resolution palynology at Roudsea Wood, Cumbria, UK by James Innes, Mairead Rutherford, Peter Ryan, Peter Rowley-Conwy and Jeff Blackford in The Holocene
Footnotes
Acknowledgements
We are grateful to Chris Orton of the Cartographic Unit, Geography Department, Durham University for the figures. We are also grateful to Professor Yongqiang Zong and the late Professor Michael Tooley for making available their cores from Roudsea Wood 33.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We are very grateful to the Leverhulme Trust for funding the research under project LF/001/28P.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.