
Abstract
Human impacts on lakes can lead to nutrient release, increased algal productivity and consequently low oxygen concentrations in deepwater environments. Here we report the unexpected and contrasting finding of a pronounced shift to chironomid and invertebrate assemblages suggesting oligotrophication and oxygen increase in Lac de Champex, a Swiss mountain lake, due to human activities during the Mediaeval period. Chironomid assemblages in the lower part of the lake sediment record show changes that agree with known climatic shifts during the Lateglacial period and early Holocene and a progressively stronger influence of peat development, and possibly humic conditions, from ca. 8500 calibrated 14C years BP (cal. BP) onwards. This resulted in low chironomid influx and assemblages with high abundances of Procladius and Chaoborus ca. 2000–3000 cal. BP. These assemblages resemble those found in relatively nutrient-rich, oxygen-poor lakes and may have developed due to natural, long-term development to more nutrient-rich and possibly humic conditions, and from the Bronze Age (ca. 4000 cal. BP) onwards, due to human activities such as pasturing. Ca. 1000 cal. BP, an abrupt increase in sediment accumulation, chironomid influx and distinct changes in the chironomid assemblage composition were observed, coinciding with evidence for local human activities from pollen data. A major transition in chironomid and invertebrate assemblages followed ca. 500 cal. BP with the disappearance of Chaoborus, a decrease in Procladius, and the increase or appearance of several chironomid taxa, including several rheophilous groups. We conclude that Late-Holocene human impact led to re-oligotrophication and increased oxygen availability for invertebrates in the lake. This reversal of the long-term Holocene trend was possibly promoted by increased erosion from the Mediaeval period onwards and from ca. 500 cal. BP likely by the construction of an irrigation canal (bisse), leading to inflow of clear, cool water which changed the lake’s hydrological characteristics.
Keywords
Introduction
Lakes and other freshwater bodies are of major importance, both ecologically as reservoirs of biodiversity and in terms of their socio-economic values (e.g. for drinking water, fishery, irrigation, hydroelectric power and recreational areas; Dudgeon et al., 2006; Grizzetti et al., 2016). Human activities have modified many freshwater ecosystems and led to major increases in nutrient concentrations, modifications of shorelines and flow corrections, and other changes in hydrology (e.g. Dokulil, 2014; Jeppesen et al., 2015; Smith and Schindler, 2009), in some cases well before the start of systematic lake monitoring in recent decades. The peak of human-induced eutrophication was registered in many central Europe lowland lakes in the second half of the 20th century. Thereafter, major efforts were undertaken to reduce the negative impacts of human land use and nutrient release on lake water quality and to restore lake ecosystems towards a less impacted state, which was often considered to be represented by lower nutrient concentrations and higher in-lake oxygen concentrations than observed when remediation measures were initiated. These measures have led to a reversal of the anthropogenic eutrophication trend in many European lowland lakes in the course of the late 20th and early 21st century (e.g. Lotter, 1998; Vonlanthen et al., 2012). However, the longer-term eutrophication and oxygenation history of lakes can be complex and for many lake types and regions is presently poorly understood. For example, in Switzerland the first significant human impacts on freshwater ecosystems are reported as early as the Neolithic period for lowland lakes (i.e. >5000 years ago; e.g. Rey et al., 2019; Tóth et al., 2019) and the Bronze Age for mountain lakes (i.e. ca. 4000 years ago; e.g. Heiri and Lotter, 2003), with the few available detailed studies indicating discrete centennial-scale phases of water quality impacts and recovery. Furthermore, on multi-centennial to millennial time scales natural processes such as long-term temperature changes, basin infilling, soil and catchment maturation as well as climate-induced changes in erosive input and catchment vegetation are major influences on the nutrient and oxygen status of lakes. For example, vegetation change and peatland development may influence the transport of particulate or dissolved organic matter into aquatic ecosystems (e.g. Rose et al., 2015; Rosén and Hammarlund, 2007; Solomon et al., 2015), which in turn can affect decompositional processes, water transparency and therefore also deepwater oxygen concentrations in lakes. These processes may lead to changes in trophic state and oxygen availability, for example, more nutrient-rich and oxygen-poor conditions in stratified lakes, even in the absence of human activity. Although a wide consensus appears to exist that the natural pre-impacted state of lakes in Central Europe and the Alpine region was associated with lower nutrient concentrations and higher deepwater oxygen concentrations than observed in the 20th and 21st century, very few datasets are available that actually demonstrate the conditions that existed before early or industrial-era eutrophication.
Small and remote lakes, such as mountain lakes at higher elevations, are particularly underexplored in respect to their long-term eutrophication history and regarding changes in deepwater oxygen availability. This may be because they are typically less frequently surveyed than larger lakes in more populated and urban regions, and also since human impact on such lakes may be associated as much with land use changes before the instrumental period as with population growth and remediation measures during the industrial period and the 20th century. For example, for mountain lakes in the Alps only very limited palaeoecological information on long-term ecosystem development and human impact is available. Some studies have shown that human activities such as pasturing can lead to eutrophication and abrupt decreases in oxygen concentrations in small lakes, in some cases well before systematic water chemistry measurement campaigns in the 20th century, with potentially adverse effects on deepwater benthic organisms in these ecosystems (e.g. Belle et al., 2016; Hausmann et al., 2002; Heiri and Lotter, 2003).
Palaeolimnological studies can provide longer-term assessments of ecosystem change and the effects of environmental pollution on lakes than available biomonitoring and analytical water chemistry data. For example, such studies, based on diatom assemblages or other palaeoecological indicators sensitive to nutrient or oxygen change, have revealed that in many Central European lowland lakes the most recent eutrophication trend was initiated in the early or mid 20th century, and that lake restoration measures have led to a recovery of aquatic assemblage composition in many instances (e.g. AquaPlus, 2004; Hürlimann et al., 1999; Lotter, 1998). Chironomid remains in lake sediments can also be used to track past changes in oxygenation and nutrient status of lakes (e.g. Verbruggen et al., 2011). Larvae of Chironomidae and other aquatic midges such as Chaoboridae represent an important component of macroinvertebrate populations of lakes and can be found in a wide range of aquatic environments (Andersen et al., 2013; Armitage et al., 1995; Saether, 1979). Chironomid larvae are of particular interest as bioindicators, as many chironomid taxa are sensitive to environmental variables and tolerate a narrow range of ecological conditions (Walker, 2006). The chitinous head capsule of the larvae preserve in sediments for thousands of years, making it possible to reconstruct past environmental variables (Bolland et al., 2021; Brooks, 2006; Walker, 1987). Chironomid fossils have been widely used to reconstruct past temperatures (Barley et al., 2006; Brooks, 2006; Heiri et al., 2011) but are also sensitive to changes in the trophic state and particularly changes in deepwater oxygen availability in lakes (Brodersen and Quinlan, 2006; Saether, 1979; Wiederholm and Eriksson, 1979). Other invertebrate remains, often analysed together with chironomids, can provide valuable supporting information for interpreting chironomid records (Courtney-Mustaphi et al., 2023; Greenwood et al., 2006; Kenward, 1976). For example, the abundance of Chaoborus remains (Diptera, phantom midges) shows a correlation with lake bottom oxygen concentrations (Ursenbacher et al., 2020), whereas simuliid remains (Diptera, blackflies) can provide information about the fluvial conditions (Currie and Walker, 1992).
In this study, we analysed larval chironomid head capsules and other available aquatic invertebrate remains in the sediments of a subalpine lake, Lac de Champex, covering an interval from the Lateglacial period (ca. 13,000 year ago) to the present. Lac de Champex (1466 m a.s.l) is a mountain lake situated in the Valais canton of Switzerland, where archaeological findings attest the presence of human settlements from the Early Mesolithic to the present (Curdy, 2007). The region is of particular interest, since its hydrology has been strongly influenced by the construction of canals (called bisses) for irrigation purposes from the 14th to 15th century (650–450 cal. BP) onwards (Reynard, 2001). These constructions influenced the ecology of streams due to water re-routeing and water abstraction, but also potentially affected the ecological conditions of small lakes receiving this diverted water, such as Lac de Champex that receives water by the bisse from the adjacent Arpette valley. The aim of our study is to investigate the long-term aquatic ecosystem development of Lac de Champex based on chironomid and other invertebrate remains and the effect of human induced disturbances on the lake ecosystem. Particularly we are interested in assessing whether hydrological modifications intensifying in the region from 500 cal. BP onwards (Reynard, 2001) potentially impacted the lake. In addition to hydrological changes, increased erosional input by landscape opening, increased nutrients due to grazing activities (e.g. Hausmann et al., 2002; Heiri and Lotter, 2003), and fish introduction (e.g. Millet et al., 2009; Pechlaner, 1966) are all human-induced disturbances that could potentially have influenced the state of the lake during the Late-Holocene.
Study area
Lac de Champex is a small subalpine lake situated 15 km south of Martigny in the inner Swiss Alps (Valais, Switzerland) (Figure 1). It lies on a granitic bedrock at an elevation of 1466 m a.s.l. The lake has a surface area of 0.11 km2 and presently a maximal depth of ca. 6 m (Lugon, 1927). Based on the closest weather station in Montana, ca. 40 km from Lac de Champex and at 1427 m asl, the average annual temperature at Lac de Champex is ca. 5.6°C, the highest monthly average temperature is in July at ca. 14.4°C and the lowest monthly average temperature in January at ca. −2.1° (MeteoSwiss, 2021; Rey et al., 2022). Annual precipitation has been estimated to 786 mm at Orsières (929 m asl) ca. 2 km from the lake (MeteoSwiss, 2021; Rey et al., 2022). The lake is surrounded by a closed forest (Picea abies, Pinus sylvestris, Larix decidua and sparse Betula pendula) and the catchment is in large parts very steep, except in its western part, which is covered by typical bog vegetation. Champex was a mayen (pasture at intermediate altitude) until the rapid development of the village Champex-Lac due to increasing tourism from the late 19th century onwards. Today, the village extends along the lake shore in the south, east and north of the lake (Figure 1). Water from the Arpette Valley reaches Lac de Champex through the bisse which enters the lake on the western side (Lugon, 1927).
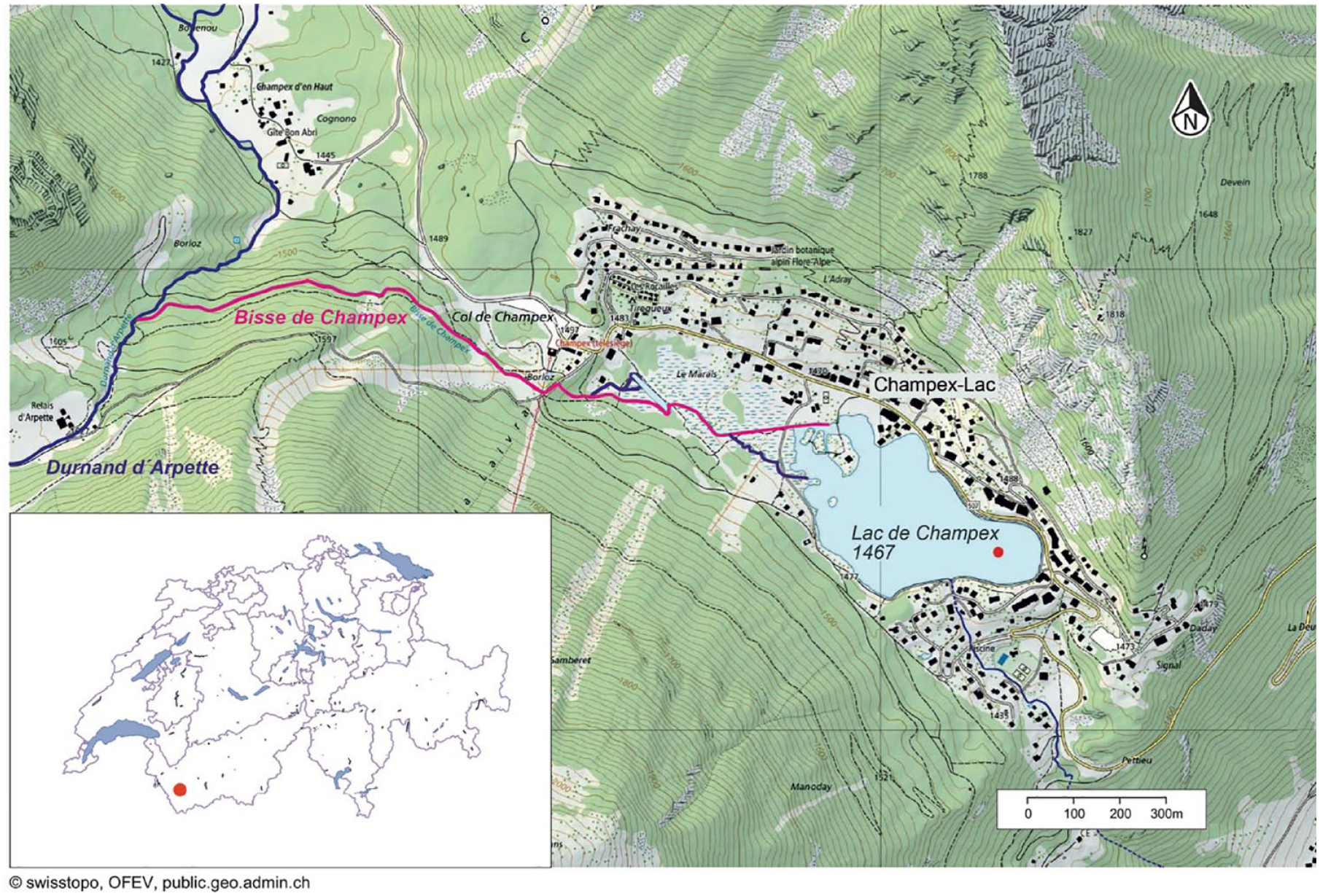
Map of Lac de Champex showing watercourses (dark blue lines) and the bisse (magenta line). The red dot indicates the coring site. The inset shows an outline map of Switzerland, with the red circle indicating the location of Lac de Champex (© swisstopo, OFEV, public.geo.admin.ch).
Methods
Coring, dating and loss on ignition
In October 2015, two parallel cores were taken from the lake sediment with a UWITEC-piston corer (diameter 60 mm). The two parallel cores, one from 0 to 600 cm, and the other from 50 to 650 cm, were correlated based on lithological features to result in a master-core with a length of 612.5 cm. Additionally, the uppermost 20 cm were cored with a Kayak gravity corer. The cores were taken from the eastern part of the lake (coordinates: 46°01′39.025″N/7°07′07.943″E), at a water depth of 5.2 m. Sediment sampling and core correlation are described in detail in Rey et al. (2022). These authors also provide a detailed reconstruction of the vegetation and fire history of the region based on pollen, spore, stomata, macrofossil and charcoal analyses of the sediments of the lake.
Terrestrial plant remains were collected from 15 different depths along the core and sent for dating to the Laboratory for the Analysis of Radiocarbon with AMS (LARA) at the University of Bern. To obtain the calibrated dates from the radiocarbon age estimates, the programme CALIB 6.1.0 was used (Stuiver and Reimer, 1993) and calibrated ages are presented as calibrated 14C years BP (cal. BP). The radiocarbon dates and the age depth model used to date the sediment sequence are described in detail in Rey et al. (2022).
Loss on ignition (LOI) at 550°C, as an estimate of the organic matter content of the sediments, was measured following Heiri et al. (2001). LOI was only analysed in the uppermost 420 cm of the sediment section.
Invertebrate analysis
A total of 47 samples were taken at intervals of 2.5–15 cm along the 612.5 cm master-core. Sample volume varied from 0.5 to 6 cm3. From 200 to 435 cm, the samples were treated with 10% KOH for 6 h at room temperature to facilitate the sieving, which was performed with a 100 μm sieve. For the remaining samples no pretreatment with KOH was needed.
Chironomid head capsules and other invertebrate remains (mostly aquatic insect mandibles), were picked out under a stereomicroscope (Leica, Germany) and mounted on microscope slides in Euparal® after drying. Then, the remains were identified using a microscope at 100–400× magnification. Taxonomic keys from Brooks et al. (2007) and Andersen et al. (2013) were used for the identification of the Chironomidae. We mainly followed the taxonomic resolution described in Brooks et al. (2007), but the types that were difficult to separate with sufficient confidence were combined together at a lower taxonomic resolution. For the same reason, the different taxa of Pentaneurini were not distinguished. Other invertebrate remains were identified by using the photo and slide collection of the Geoecology group (University of Basel, Department of Environmental Science), Courtney-Mustaphi et al. (2023) as well as descriptions in Vandekerkhove et al. (2004) for ephippia, Francis (2001) for statoblasts and Sweetman and Smol (2006) for Chaoboridae.
For chironomid analysis we were aiming for a minimum of 50 chironomid head capsules (hc) per sample (Heiri and Lotter, 2001). Enumeration of hc followed standard procedures described in Brooks et al. (2007). When the chironomid concentration was deemed too high, a modified two-chamber subsampler (modified from Meyer, 1990) was used to separate samples into two equal portions before picking out the remains (following Heiri et al., 2003).
Numerical analysis
Chironomid influx was estimated by first calculating chironomid concentrations by dividing counts (as number of larval hc) by the analysed volume of sediment (as cm3) and multiplying this value by the sediment accumulation rate (in cm/a). We also calculated the ratio between Chaoborus and chironomid remains, as this has been shown to be sensitive to changes in late summer deepwater oxygen concentrations in stratified lakes (Quinlan and Smol, 2010a; Ursenbacher et al., 2020), with high values (i.e. high concentrations of chaoborids) representing conditions with low hypolimnetic oxygen concentrations. The ratio is presented after log (x + 1) transformation as the data are strongly right-skewed and the transformation aids in seeing long-term variations of this ratio in our record. Chironomid assemblage zones were identified by using optimal sum of squares partitioning (Birks and Gordon, 1985), and the statistically significant boundaries were determined using the broken-stick approach (Bennett, 1996). Finally, to assess whether chironomid and invertebrate assemblage changes were indicative for changes in nutrient and/or oxygen concentrations, fossil samples from Lac de Champex were plotted passively in Detrended Canonical Correspondence Analyses (DCCAs; Ter Braak, 1986) of surface sediment datasets representing the distribution of chironomids and chitinous invertebrate remains in the modern environment. Chironomid assemblage percentage data from Lac de Champex were plotted passively in DCCAs based on the surface sediment datasets from Verbruggen et al. (2011) and Heiri and Lotter (2010). These represent the two available datasets that include sites from Central Europe and describe the relationship between subfossil chironomid assemblages in lake surface sediments and total phosphorus (TP), the main plant nutrient that determines lake productivity in the region (e.g. Lotter et al., 1998). Log-transformed TP concentrations measured at these sites (as presented in Bigler et al., 2006; Heiri and Lotter, 2005; Verbruggen et al., 2011) were used as sole constraining environmental variable for these DCCAs. As a consequence, variations of passively plotted samples along DCCA axis 1 represent changes in fossil chironomid assemblages at Lac de Champex typical for variations in TP values in the modern environment. The Verbruggen et al. (2011) dataset covers 28 lakes ranging from Swedish Lapland to Central Italy over a wide TP range. In contrast to the Heiri and Lotter (2010) data, this dataset does not include major elevational differences but covers several regions with different biogeographic and geological settings within Europe (Scandinavia, northern, central and southern Germany, northern, central and southern Italy). TP in this dataset was closely and negatively correlated with late summer deepwater oxygen concentrations. The Heiri and Lotter (2010) dataset consists of samples from 113 lakes. In contrast with the Verbruggen et al. (2011) data these are all located in one region, the Swiss Alps, and they cover a large elevational range of 418–2815 m asl. Several water chemistry variables (e.g. TN, alkalinity, DOC) correlate strongly and positively with TP in this dataset (Bigler et al., 2006; Lotter et al., 1998). However, deepwater oxygen has not specifically been measured in this campaign. Invertebrate percentage data, including overall chironomid abundances but also other invertebrate remains such as Chaoborus mandibles, ephippia of daphniids, bryozoan statoblasts, and oribatid mites, were plotted passively in a DCCA of modern assemblage data from 36 Swiss lakes covering an elevational gradient of 403–2065 m asl (Ursenbacher et al., 2020). Ursenbacher et al. (2020) showed that variations in percentages of invertebrate remains in surface sediments from these lakes were closely related to late-summer deepwater oxygen concentrations (botO2). To mainly capture the assemblage variations typical for changing deepwater oxygen conditions this DCCA was constrained by log-transformed botO2 only, with variations in DCCA axis 1 scores for the passive samples expected to capture variations in invertebrate assemblages typical for changing oxygen conditions. All ordinations were calculated on square root transformed percentage data. If necessary, DCCA axes were transformed by multiplying by −1 and subtracting the minimum value so that high DCCA axis 1 scores represent conditions in modern lakes with high TP and low botO2 values. All the statistical analyses were performed using RStudio (R Core Team, 2016), except the DCCAs that were done with the programme Canoco 5 (Ter Braak and Smilauer, 1997). Optimal partitioning was performed in R with support by Dr. Christoph Schwörer (Institute of Plant Sciences, University of Bern).
Results
Lithology and chronology
The majority of the Lac de Champex core is composed of gyttja, except at its lower part, where it consisted of clay (463–500 cm) and organic-rich silt (438–463 cm). Between 99 and 130 cm, the gyttja is enriched with peat. The organic matter content (LOI) of the sediment ranges between ca. 40% and 60% for most of the gyttja section of the record (Figure 2). Above ca. 180 cm sediment depth, organic matter content starts to decrease to reach values around 20% in the uppermost sediment layers. This decrease is interrupted by higher values characterizing the peat layer 99–130 cm. As described in Rey et al. (2022) the age-depth relationship developed for Lac de Champex shows a more or less constant sediment accumulation rate (ca. 50 year/cm) until 193 cm, when the rate increases considerably (ca. 5 year/cm). The two outliers in the age-depth model were from the peaty layer and obviously older than other dates just below this section, and therefore are interpreted as being old inwashed material brought into the lake together with the peat.
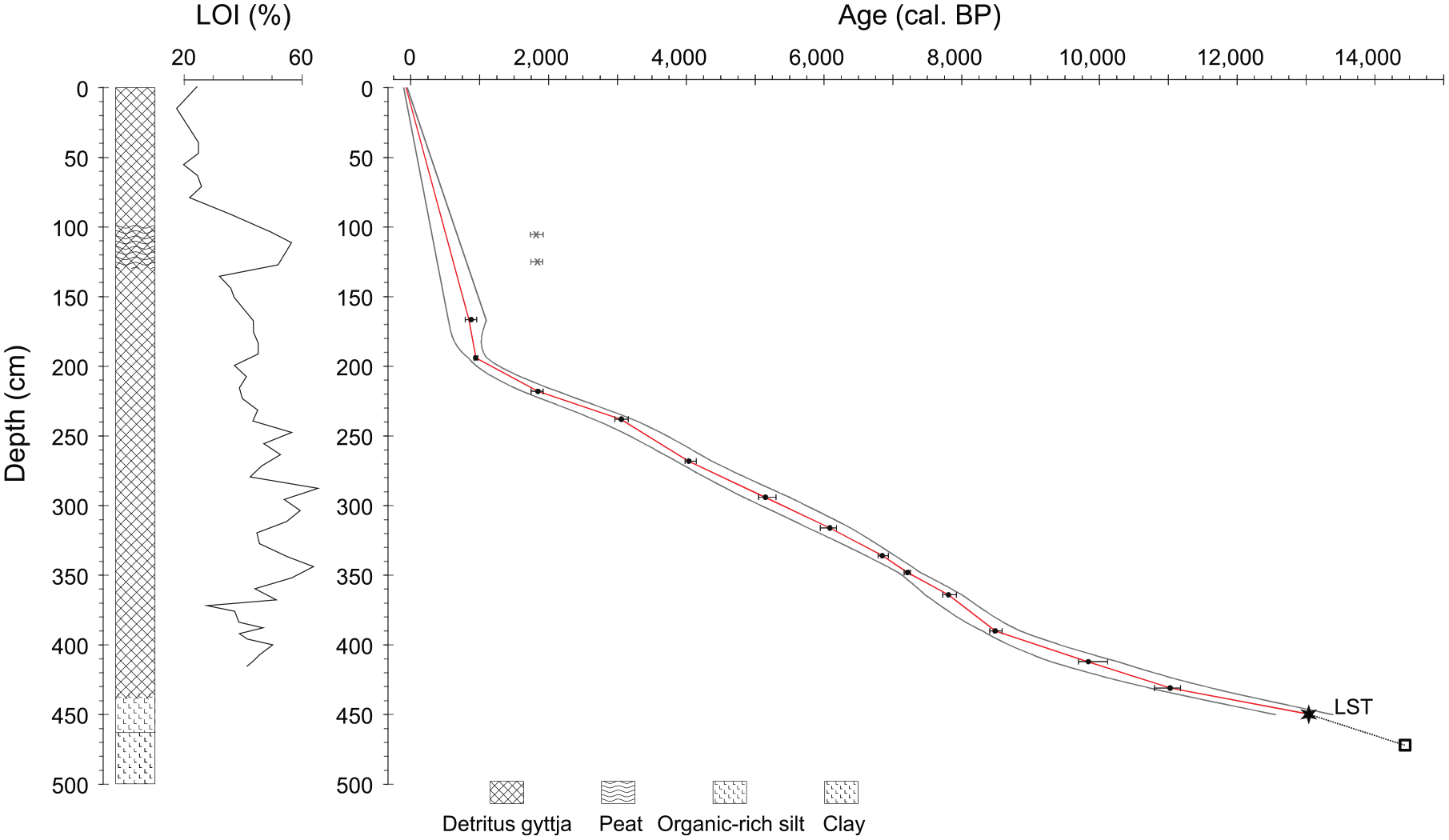
Age-depth model (from Rey et al., 2022), lithology and LOI of the Lac de Champex record. Interpolated ages (red line) are shown together with an uncertainty envelope calculated with a Generalized Additive Model (GAM; Heegaard et al., 2005). Points indicate calibrated 14C ages of terrestrial macrofossils and their 95% age confidence intervals, except for the two oldest points corresponding to the Laacher See tephra (LST) and a pollen-stratigraphic date (see Rey et al., 2022 for details). The uppermost two 14C dates are from a reworked peat layer and not included in the age-depth model.
Chironomid analysis
A total of 50 taxa were identified in the 47 samples, with an average of 86 head capsules per sample. Considerable variations of chironomid concentration were observed, from 587 to 1.5 hc/cm3. Due to the low concentrations in some parts of the record, five samples had between 35 and 40 identified head capsules and three samples less than 35. One further sample (390 cm) was excluded because of too low concentration (1.5 hc/cm3). However, since major changes in chironomid assemblages were well represented in the record, these low-count samples were not merged with adjacent ones to maintain the record at the maximum temporal resolution. Significant zone boundaries were detected at 448.75, 436.25, 422.5, 390, 335, 97 and 67.5 cm depth using the broken-stick approach (Bennett, 1996). Only four of these boundaries (448.75, 436.25, 390 and 97 cm) are used for discussing the results and shown in the diagram representing the chironomid results (Figure 3), since they represent the most relevant changes in chironomid assemblage composition.
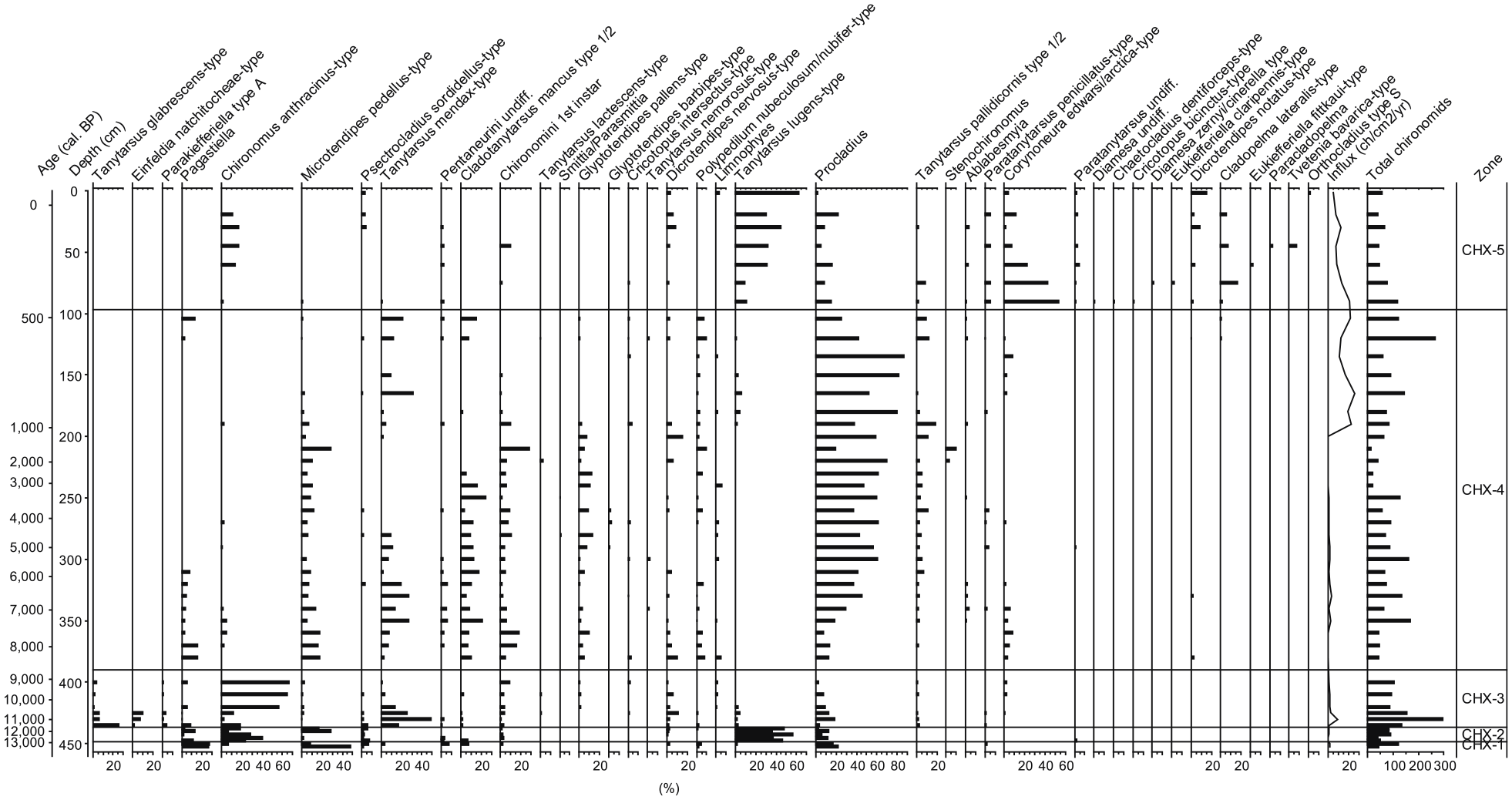
Chironomid percentage diagram. Identified taxa are shown as percentages of total number of head capsules (hc). Selected statistically significant zones are indicated with horizontal lines. Some individual rare taxa are not displayed.
Chironomid assemblage zone 1 (CHX-1, 452.5–448.75 cm, ca. 13,200–12,950 cal. BP)
This zone is the oldest of the sequence and includes only two samples. It is characterized by a dominance of Procladius, Pagastiella and Microtendipes pedellus-type. The almost total absence of individuals from the Tanytarsini subfamily is also distinctive for this period.
Chironomid assemblage zone 2 (CHX-2, 448.75–436.25 cm, ca. 12,950–11,600 cal. BP)
This period is dominated by Tanytarsus lugens-type and Chironomus anthracinus-type, which increase distinctly in this zone. Procladius remains at relatively constant abundances whereas Pagastiella and Microtendipes decrease particularly in the middle of this zone.
Chironomid assemblage zone 3 (CHX-3, 436.25–390 cm, ca. 11,600–8500 cal. BP)
C. anthracinus-type remains high in abundance and is overall the dominant taxon of CHX-3. This zone is also marked by the appearance of Tanytarsus mendax-type and Tanytarsus glabrescens-type, with peaks at depths of 430 and 435 cm, respectively. Both taxa become less abundant towards the end of the zone, but populations of T. mendax-type recover in the following zone (CHX-4), whereas T. glabrescens-type disappears completely and never recovers in the record. Procladius is present within this zone at decreasing intermediate to low abundances.
Chironomid assemblage zone 4 (CHX-4, 390–97 cm, ca. 8500–500 cal. BP)
CHX-4 is the zone of the longest duration. This zone is distinguished by the appearance in higher abundance of Cladotanytarsus mancus-type, Polypedilum nubeculosum/nubifer-type, Glyptotendipes pallens-type, and by a major reduction of C. anthracinus-type from values >60% to values below 5%. Procladius starts to increase at the beginning of the period and quickly stabilizes at high percentages to become the dominant taxon. In this zone, Pagastiella, T. mendax-type, C. mancus-type and Corynoneura edwarsi-type populations seem to disappear one after the other. In addition, the head capsule influx reaches its lowest level from 240 to 200 cm. At the end of CHX-4 (from ca. 1000 cal. BP onwards), these taxa all recover, and at the same time, head capsules influx and concentration increase drastically. Simultaneously, low percentages of T. lugens-type are observed again, for the first time after an absence of more than 9000 years.
Chironomid assemblage zone 5 (CHX-5, 97–0 cm, ca. 500 cal. BP–present)
The most dramatic shift in the chironomid composition takes place at the beginning of this zone. This period starts with a massive abundance increase of Corynoneura edwardsi-type (more than 50%), although percentages decrease again with time. T. lugens-type reaches more than 10% at the beginning of CHX-5, for the first time after the population’s collapse at the end of CHX-2, which took place more than 11,000 years before. T. lugens-type shows an increasing trend during this period and reaches 58% of the head capsules in the uppermost sample. Procladius decreases significantly, and T. mendax-type disappears completely. CHX-5 is also characterized by the reappearance of C. anthracinus-type at intermediate abundance, after only small and sparse appearances for more than 8000 years. This period shows a higher diversity, especially among the subfamily Orthocladinae, with some rare taxa present only once in the record, such as Chaetocladius dentiforceps-type, Orthocladius-type S, Psectrocladius septentrionalis-type, Tvetenia bavarica-type or Eukiefferiella fittkaui-type.
Non-chironomid remains
In addition to the chironomid head capsules, other invertebrate remains were found in the sediments of Lac de Champex (Figure 4). Most of them were mandibles of arthropods, namely Amphipoda, Ephemeroptera, Trichoptera, Sialis and Chaoborus, the latter being the most abundant group. Chaoborus mandibles first appear at the beginning of the third chironomid assemblage zone (CHX-3), and are consistently present until the beginning of the last zone (CHX-5) when they disappeared again. On the other hand, Simuliidae mouthparts and Plumatella statoblasts were found only when Chaoborus mandibles were absent, in the most recent and oldest part of the record, respectively.
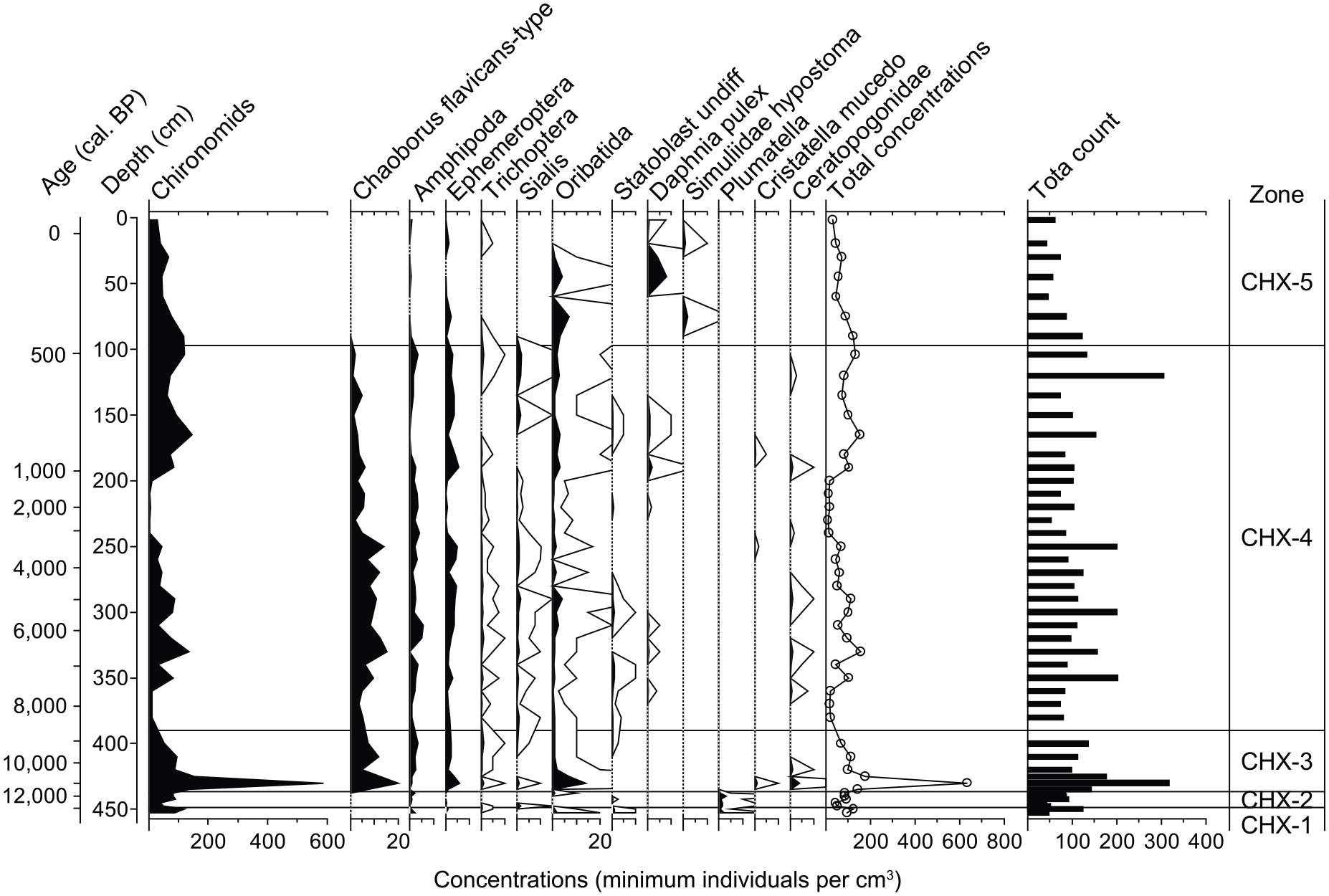
Invertebrate concentration diagram. For comparison the statistically significant boundaries of the chironomid assemblages shown in Figure 3 are also indicated (CHX-1 to 5). White outline graphs show 10× exaggeration for rare taxa.
Numerical analyses
DCCAs of modern chironomid assemblage data with TP as the only constraining variable, and the fossil data added passively, were used to examine whether chironomid assemblage composition in Lac de Champex changed between assemblages representing different levels of TP in the modern environments. The results consistently show that the chironomid assemblage composition in the lowest part of the record, representing the Late Glacial period, is typical of modern surface sediment samples with low to intermediate TP concentrations. This was the case both when the pan-European dataset of Verbruggen et al. (2011) and the Alpine dataset of Heiri and Lotter (2010) formed the basis of these inferences (Figure 5d). DCCA axis 1 values increase in the early Holocene and remain high with an increasing trend towards the Late-Holocene. From ca. 1000 cal. BP onwards values decrease abruptly to reach exceptionally low values in sediments representing the past ca. 500 years. This indicates that chironomid assemblages in the youngest sediments of Lac de Champex are typical for lakes with distinctly lower TP values than those in the mid-Holocene with high DCCA axis 1 values. A similar pattern is apparent when the Lac de Champex invertebrate data are added passively to a DCCA of the modern invertebrate assemblage data from the Alpine region of Ursenbacher et al. (2020, Figure 5e). Lateglacial samples have low DCCA axis 1 values, indicative of high-oxygen lakes in the modern environment. Values increase abruptly in the early Holocene followed by a more gradual, long-term increase in the mid- to Late-Holocene until 2000 cal. BP, suggesting increasing deepwater anoxia in the lake. An abrupt reversal of this trend and decreasing values are then apparent in the past ca. 1000 years. Lowest values are observed in the past ca. 500 years, indicating that invertebrate assemblages in this part of the record are typical for oxygen-rich deepwater environments. These results are supported by the chaoborid to chironomid ratio (Figure 5f) that shows relatively high values from the early Holocene to ca. 1000 cal. BP, typical for lakes with anoxic deepwater conditions. Maximum values were observed between 3000 and 1500 cal. BP, followed by a major decrease between ca. 1500 and 1000 cal. BP and absence of chaoborids for the past ca. 500 years.
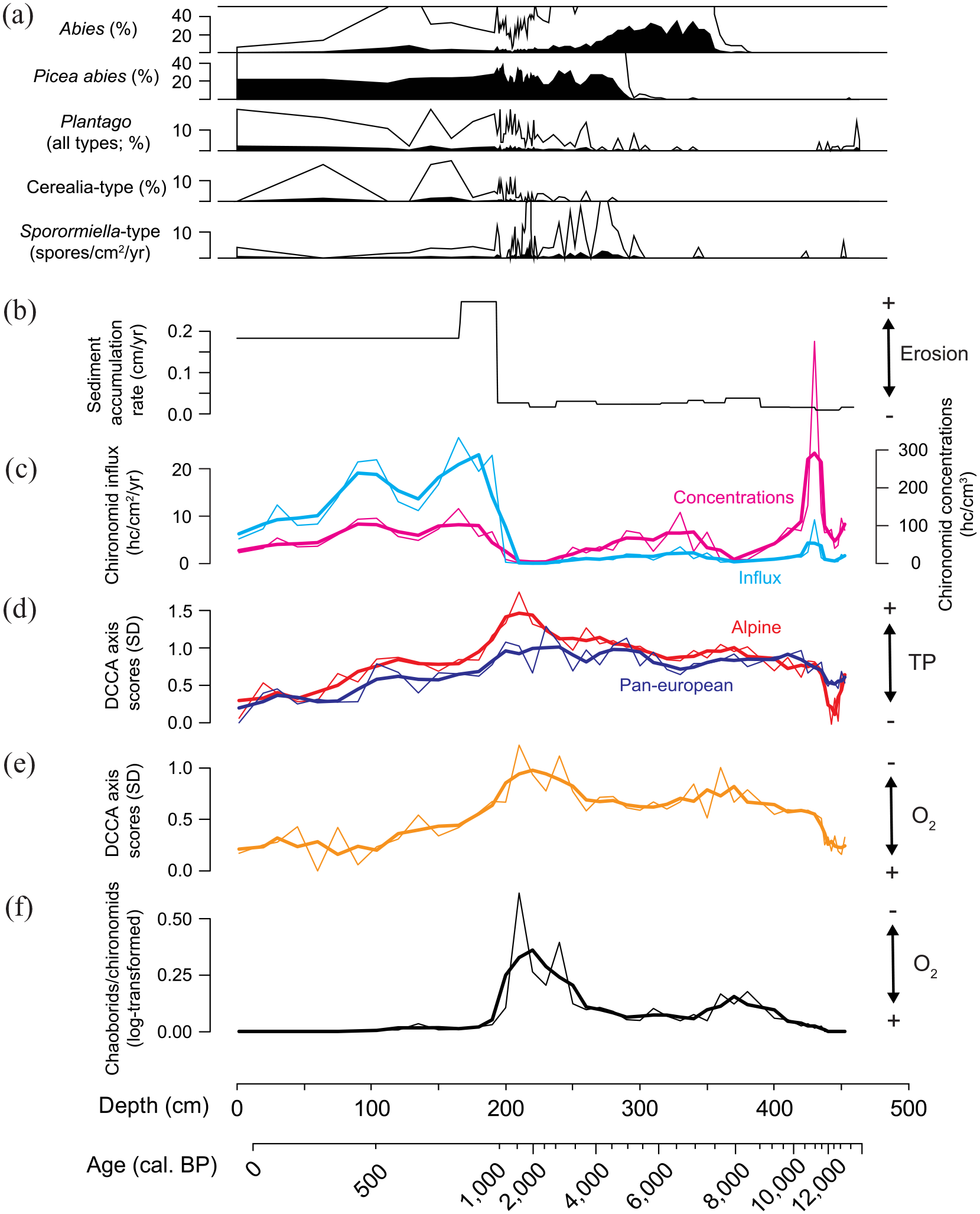
Selected pollen types for the Lac de Champex record together with chironomid influx and concentration data, sediment accumulation rates and numerical reconstructions of TP and oxygen concentration changes in the lake. (a) Pollen percentages of key conifer species (Abies, Picea abies) expanding in the mid-Holocene together with examples of pollen of grazing indicating (Plantago) and cultivated (Cerealia) plants as well as influx (as spores/cm2/year) of dung-indicating fungi (Sporormiella-type; from Rey et al., 2022). (b) Sediment accumulation rates. (c) Chironomid influx and concentrations. (d) Axis 1 scores of a DCCA of chironomid assemblages from lakes across Europe (Verbruggen et al., 2011) and the Swiss Alpine region (Heiri and Lotter, 2010) with TP as only constraining variable and the Lac de Champex chironomid data added passively. (e) Axis 1 scores of a DCCA of invertebrate assemblage data from small Swiss lakes (Ursenbacher et al., 2020) with deepwater oxygen concentration as only constraining variable and Lac de Champex invertebrate data added passively. (f) Log (x + 1)-transformed ratio between chaoborid and chironomid remains in the sediments. Arrows next to (d–f) indicate whether Lac de Champex samples are more typical for high/low TP and oxygen concentrations based on these analyses.
Discussion
Major shifts are apparent in the chironomid and invertebrate record of Lac de Champex, indicating pronounced changes in within-lake environments. Initial changes at ca. 12,950–11,650 cal. BP coincide with and mainly represent the climatic transitions of the Late Glacial period, particularly the warming at the transition from the Younger Dryas cold period to the early Holocene (Heiri et al., 2014). Several assemblage changes are also apparent during the Holocene, for example at ca. 9000–8500 cal. BP, when the abundance of Chironomus anthracinus-type decreased in the record and at ca. 3000 cal. BP when chironomid influx and concentration values decreased and Cladotanytarsus disappeared. Further changes are apparent at ca. 1000 cal. BP when Tanytarsus mendax-type and Tanytarsus lugens-type reappeared in the record after an absence of thousands of years, Microtendipes and Glyptotendipes decreased distinctly and chironomid concentration and influx values suggest a distinct increase in overall chironomid abundance in the lake. At about this time the chaoborid to chironomid ratio also showed a major decrease in the record. At ca. 500 cal. BP, finally, Corynoneura edwardsi-type increased abruptly together with a number of taxa that were largely absent from the lake or not as abundant since the early Holocene. Taxa that illustrate this last compositional shift are Cladopelma lateralis-type, Tanytarsus lugens-type, Chironomus anthracinus-type, Dicrotendipes notatus-type and Paratanytarsus penicillatus-type. This later transition also coincided with the final disappearance of Chaoborus, a taxon that was abundant in the lake for thousands of years. Variations in chironomid and invertebrate assemblage changes are captured in the DCCAs that strongly suggest that many of these changes are related to changing nutrient and oxygen availability in the lake, particularly those in the past 1000 years. DCCA axis 1 values based on analyses of the chironomid and invertebrate data are strongly and highly significantly correlated (Pearson correlation 0.67–0.81, p < 10−6), indicating that chironomid and invertebrate assemblage change occurred in parallel and represented a similar trajectory of change. In contrast to zonation procedures and broken stick-based assessments of significance, which assess whether major changes occur between individual, adjacent samples in a record, DCCA can also capture longer-term, progressive changes in assemblages in palaeoecological datasets. For Lac de Champex, variations in DCCA axis 1 values, together with chironomid influx and concentrations as well as the chaoborid to chironomid ratio, suggest a progressive transition to assemblage states more typical for lakes with anoxic bottom waters and relatively high nutrient conditions in the course of the Holocene. This culminates between ca. 3000 and 1000 cal. BP in a period with high DCCA values, very low chironomid influx in the lake and the highest Chaoborus abundances. This period was followed by a reversal of this trend as DCCA axis 1 values and the chaoborid to chironomid ratio decreased and chironomid influx values increased again from ca. 1000 cal. BP onwards. This indicates that assemblage states in these youngest intervals of the Lac de Champex record were again typical for lakes with more oxygen-rich bottom water and lower nutrient concentrations in the modern environment.
Early environmental and climatic influence on the invertebrate assemblages
Early changes in the chironomid record coincide with major shifts in climate and vegetation at Lac de Champex, that are also clearly represented in the pollen record from the lake (Rey et al., 2022). Samples in the lowest sediments (CHX-1), originating from the Allerød interstadial period, are dominated by taxa typical for mountain lakes, such as Microtendipes and Chironomus anthracinus-type (Heiri and Lotter, 2010), but also contain some taxa with a very wide temperature distribution, such as Polypedilum nubeculosum-type and Procladius (Heiri et al., 2011). During the Younger Dryas cold period, the last cold interval of the last ice age, Tanytarsus lugens-type, a taxon typical of cool and relatively nutrient-poor (oligo-mesotrophic) environments (Heiri et al., 2011; Lotter et al., 1998; Saether, 1979) expanded in the lake and some of the formerly abundant taxa (e.g. Polypedilum nubeculosum-type, Pagastiella, Microtendipes pedellus-type) disappeared or decreased in abundance (CHX-2). Based on both the ecology of the occurring chironomid taxa and the DCCA results, Lac de Champex was a cooler environment with lower nutrient but higher oxygen concentrations than before or after this episode. At the transition to the Holocene (CHX-2/CHX-3), a period in which summer temperatures increased by several degrees within ca. 100 years (Heiri et al., 2015), Tanytarsus lugens-type decreased abruptly and new chironomid taxa typical for warmer and more nutrient-rich environments expanded, such T. mendax-type and T. glabrescens-type (see e.g. Heiri et al., 2011). Simultaneously, remains of Chaoborus flavicans-type appeared for the first time (CHX-3, Figure 4). This taxon can benefit from reduced oxygen concentrations in lakes, as it can evade predators by escaping to hypoxic water layers or sediments (Voss and Mumm, 1999). Increasing nutrient concentrations and decreasing deepwater oxygen availability are also indicated by the DCCA analyses which consistently indicate that chironomid and invertebrate assemblages at Lac de Champex became more similar to those presently found in nutrient-rich and oxygen-poor lakes.
The taxonomic composition of chironomids and other aquatic invertebrates at Lac de Champex remained relatively stable in the early Holocene with some minor oscillations. However, at ca. 9000–8500 cal. BP a major shift in chironomid assemblages occurred when Procladius started to increase to become the dominant taxon, while Chironomus anthracinus-type strongly decreased in abundance. Around this period (from ca. 8200 cal. BP onwards), silver fir (Abies alba) expanded in the region and locally at Lac de Champex (Figure 5). The expansion of Abies alba is usually interpreted as being the consequence of an initial early to mid-Holocene climatic shift to more oceanic climate conditions, specifically with less late frost occurrence and increased summer moisture availability (Berthel et al., 2012; Rey et al., 2013; Tinner and Lotter, 2001). Other studies of Swiss mountain lakes have reported changing chironomid community compositions as coinciding with the establishment of Abies alba (Heiri et al., 2003; Zweifel, 2012). In these cases, the expansion of Abies alba was accompanied by a decline in the sediment accumulation rates together with an increase in the sediment organic content and these changes were suggested to be possible drivers for the observed changes in chironomid composition. Since many chironomid larvae colonize the sediments themselves, and feed on organic matter deposited at the lake bottom, the organic matter content of sediments can be expected to influence chironomid assemblages. Furthermore, the organic carbon contents of the sediments will also have an effect on the amount of oxygen available for sediment-dwelling chironomids. However, at Lac de Champex, the chironomid assemblages changed before Abies alba arrived at the lake (Figures 3 and 5) and major changes in the lithology and sediment composition are not observed in this interval (Figure 2). The expansion of Abies alba therefore cannot explain the observed changes in chironomid assemblages in Lake Champex ca. 9000–8500 cal. BP.
Based on observed vegetation changes, the early to Mid-Holocene transition is generally considered to be associated with a shift to moister climatic conditions in Central Europe and the Alpine region (e.g. Herzschuh et al., 2023; Rey et al., 2022; Tinner and Lotter, 2001). Moister conditions would have likely promoted the growth of peat around the lake. Lac de Champex presently has a large peat area in the western part of the catchment (Figure 1), and peat coverage may have been even more expansive in the past. Peat mosses generate humic and fulvic acids, which, in extreme situations, in dystrophic (or humic) lakes, colour the lake water dark brown (Hansen, 1962). Dissolved organic substances can have pronounced impact on lakes at relatively low concentrations, by reducing the water transparency (Molot and Dillon, 1997) and decreasing deepwater oxygen concentrations (Couture et al., 2015). Lower water transparency results in a reduced and shallower photic zone in lakes with elevated dissolved organic carbon (DOC) concentrations (Brothers et al., 2014) and consequently also lower algal growth and reduced oxygen production, whereas the oxidation of dissolved organic substances increases oxygen demand in the water column (Couture et al., 2015). Expansion of peat around Lac de Champex would have been associated with such changes in lake water chemistry, which may have contributed to the chironomid assemblage shift starting at ca. 8500 cal. BP. Lower water transparency and oxygen concentrations may also have favoured Chaoborus in the lake, as Chaoborus has been reported to benefit from dystrophic conditions in small lakes (Quinlan and Smol, 2010b) with reduced visibility reducing the predation pressure on these meroplanktonic invertebrates. Other alpine lakes studied for chironomid remains in Switzerland are mainly situated on carbonate bedrock (e.g. Heiri et al., 2003; Heiri and Lotter, 2003; Zweifel, 2012). It has been suggested that in these geological settings humic acid and DOC concentrations in lakes may be reduced (Sepp et al., 2019). Lac de Champex’ catchment is on non-calcareous granitic bedrock and this may have promoted the transition to a lake with higher DOC concentrations, leading to a more oxygen-limited environment. This interpretation is supported by the DCCA analysis of the invertebrate assemblages as well as increasing values of the chaoborid to chironomid ratio, which both suggest a moderately strong trend towards lower oxygen availability for benthic invertebrates in this part of the record (Figure 5). That the lake was characterized by a tendency to low oxygen concentrations above the deepwater sediments is also supported by the presence of larvulae (first instars) of Chironomini in most of the record preceding 1000 cal. BP, as these have been observed to be more abundant in sediments of lakes with low oxygen values (Quinlan and Smol, 2001). Chironomini larvulae show particularly high values ca. 8100–7800 cal. BP and ca. 1500 cal. BP when the DCCA based on invertebrate remains and the chaoborid to chironomid ratio suggest strongly depleted deepwater oxygen concentrations in Lac de Champex.
First human influence on the terrestrial environment
Pollen and macrofossil data suggest that Abies alba started to decline from around 4600 cal. BP onwards. Subsequently it was gradually replaced by European spruce (Picea abies) and green alder (Alnus viridis; Rey et al., 2022). In the Western Alps this transition was caused by early human activities such as clearcutting, burning and pasturing in the subalpine vegetation zone (Markgraf, 1970; Rey et al., 2013, 2022; Garcés-Pastor et al., 2022; Schwörer et al., 2015). No major shift in the chironomid percentage composition occurred at the same period. However, later, around 3000 cal. BP, the overall influx and concentrations of chironomid head capsules decreased (Figures 3 and 5), suggesting lower deepwater abundances of chironomid larvae. The concentrations of ephemeropterans also decreased strongly (Figure 4) whereas the remains of other invertebrate taxa such as Chaoborus and Amphipoda remained relatively constant. Pollen data suggest that just before this period, Plantago lanceolata, a grazing indicator (Ejarque et al., 2011), established (Rey et al., 2022). In addition, Sporormiella, a dung spore, which shows the presence of excrement in the catchment (Raper and Bush, 2009), appeared earlier in the record and persisted during this period. Overall these changes suggest increased grazing during the Bronze Age at Lac de Champex, likely leading to a further decrease in oxygen in the deeper sections of the lake. In a system where oxygen concentration is already critical, even relatively moderate nutrient release through grazing and pasturing could exacerbate the situation (Feuchtmayr et al., 2009), thus making the habitat unfavourable even for resistant chironomid groups like Procladius and resulting in an overall decrease in chironomid concentration and influx values. In contrast, Chaoborus, adapted to anoxic environments, and lacustrine amphipods, which in Swiss lakes dominate in near-shore sections of lakes, remained less affected. This interpretation is supported by the DCCA results and the chaoborid to chironomid ratio which suggest that 3000–1800 cal. BP was the most oxygen-depleted period in the record and a time interval characterized by chironomid assemblages typical for relatively nutrient-rich lakes.
Opening of the landscape, re-oligotrophication and higher oxygen availability
From ca. 1000 cal. BP (193 cm) onwards, sediment accumulation increased massively, with sediment accumulation rates estimated to be almost 10-fold higher than during the rest of the Holocene (Figures 2 and 5). Similar trends with increasing Late-Holocene sediment accumulation rates are described in numerous studies in which human activity was documented to open the landscape for agriculture and/or grazing, hence making the catchment more erodible (Edwards and Whittington, 2001; Giguet-Covex et al., 2011). Typically increased sedimentation rates lead to lower chironomid concentrations in lake sediments, due to high sediment accumulation rates and dilution of fossils by eroded material. However, at Lac de Champex, this change coincided with a major increase in chironomid concentrations and chironomid influx values (Figures 3 and 5). This implies that conditions became much more favourable for the chironomid larvae in the deepwater sections of the lake. A similar increase is observed for the Ephemeroptera mandibles also suggesting an increase in mayfly populations. Chironomini first instar larvae, regularly found in older parts of the sequence, decreased in abundance and were only found irregularly in younger sediment samples. This trend is in line with reduced oxygen stress for deepwater chironomid assemblages and ephemeropterans in the lake, which is also confirmed by the DCCAs as well as the chaoborid to chironomid ratio, which indicate that chironomid and invertebrate assemblages became more typical for nutrient-poorer and oxygen-richer lakes from ca. 1000 cal. BP onwards (Figure 5). The reappearance of T. lugens-type, even at low abundance, also supports this interpretation, as the taxon is adapted to oligo- to mesotrophic and usually relatively oxygen-rich conditions (Saether, 1979), although the presence of Chaoborus remains still suggests a tendency to anoxia. This reoligotrophication trend, associated with enhanced oxygen availability in the lake, is unexpected, since enhanced erosional inputs are generally considered to promote eutrophication, by bringing more phosphorus and other plant nutrients into lakes (Carpenter, 2005; Rekolainen et al., 2006). However, significant changes in erosion and sedimentation rates will also impact the substrate within a lake, possibly reducing the redissolution of phosphorus from the sediments and the time available for organic matter decomposition before burial. This potentially decouples the sediment organic matter from the water column and reduces the uptake of oxygen at the lake bottom due to decomposition of organic material. Irrespective of the exact cause, the trend towards chironomid and invertebrate assemblages typical for more oligotrophic and oxygen-rich environments at Lac de Champex clearly coincided with the human-induced increase in sedimentation rates. Furthermore, it is consistently supported by DCCAs of chironomids and invertebrates constrained by TP and deepwater oxygen concentrations, respectively, and by increasing chironomid abundances and decreasing ratios between chaoborid and chironomid remains.
The last 500 years: A major shift to nutrient-poorer and oxygen-rich environments
The most drastic shift in chironomid assemblage composition in Lac de Champex took place ca. 500 years ago (start of CHX-5). The trend towards more nutrient-poor conditions that began with increasing sediment accumulation rates at ca. 1000 cal. BP accelerated again ca. 500 cal. BP (Figure 5d). The oligotrophic taxon T. lugens-type, that reached only low abundances during the entire Holocene, increased to abundances comparable to those attained under the cold climatic conditions of the Younger Dryas (Figure 3). Moreover, remains of several other taxa typical of running waters (e.g. Diamesa, Chaetocladius, Orthocladius, Eukiefferiella) (Heiri et al., 2011; Saether, 1979) were observed during this period, although at relatively low abundances. Finally, the complete disappearance of the Chaoborus mandibles and a slight decrease in DCCA axis 1 scores of the analysis based on invertebrate remains (Figures 4 and 5) also suggests a more oxygenated lake (Ursenbacher et al., 2020; Voss and Mumm, 1999), although the disappearance of Chaoborus may also have been promoted by less humic conditions and clearer water. On the other hand, the presence of Chironomus anthracinus-type, usually associated with higher nutrient availability (Saether, 1979) may seem contradictory. But high nutrient content does not necessarily mean low oxygen availability, and Corynoneura edwarsi-type and Tanytarsus lugens-type may have populated shallower sections of the lake whereas Chironomus anthracinus-type only colonized the deepest sections where oxygen may still have been limiting.
Lakes in the Valais region experienced major changes in hydrology in this period, as the economic and agricultural evolution led to the need to irrigate fields and pastures. For this reason bisses (irrigation canals) were widely built in the canton (Reynard, 2001). Unfortunately, the age of the Champex Bisse is not known, but most likely it dated from around this period, as most of the other bisses from the area. For instance, the bisse of Orsières, the closest village to Champex-Lac, dates from 1471 CE (479 cal. BP; Reynard, 2001), closely coinciding with the age of the youngest change in chironomid and invertebrate assemblages in the Lac de Champex record. In agreement, no younger changes in the sediment and invertebrate record occurred that could be linked to major limnological and hydrological changes. Such changes would be expected in a small lake such as Lac de Champex when a new irrigation canal is developed that brings significant amounts of cold water from higher altitudes to the lake and vastly expands its hydrological catchment.
Interestingly, peat became mixed with the gyttja sediments ca. 25 cm below this major change (99–130 cm; Figure 2), whereas peat deposition stopped at the depth that chironomid assemblages changed abruptly (104–90 cm). On the basis of the lithology it is extremely unlikely that the lake dried out completely, which is also confirmed by the consistent presence of aquatic chironomid and other invertebrate remains in the peaty sediment sections. The peat must therefore have been brought into the lake from catchment, perhaps related to the construction or re-modification of the bisse taking water from the Durnand (the river flowing in the Val d’Arpette) into Lac de Champex (Figure 1). Indeed, this channel flows through the peat bog on the western side of the lake and the transition zone before reaching the lake itself.
It is likely that increased inflow by the bisse which presently feeds Lac de Champex may also have changed other limnological conditions in the lake. For example, peat mosses generate humic and fulvic acids, which reduce the water transparency (Molot and Dillon, 1997), particularly in lakes on siliceous bedrock such as Lac de Champex. With the newly built bisse, which brings water from higher up the mountain, the direct influence of peaty runoff from the catchment area will have been weakened, leading to higher water transparency of the lake and reducing oxygen stress due to high DOC values. Increased inflow and shorter water residence time due to the channel would have reduced nutrient concentrations, increased transparency, and the input of cold and dense water from higher elevations during summer stratification would have led to higher oxygen concentrations at the lake bottom. Increased occurrences of Simuliidae larvae, a group that can only survive in running water (Currie and Walker, 1992) as well as stream chironomids such as Chaetocladius, Diamesa, Eukieferiella and Tvetenia (Andersen et al., 2013) also support the interpretation of increased running water input during the past 500 years.
Conclusions
Our chironomid and invertebrate analyses demonstrate the complex long-term development of within-lake environments of Lac de Champex, firstly driven by natural processes such as climatic change, vegetation and catchment development, and from the late Bronze Age onwards increasingly by human activities. For millennia this development was towards chironomid and invertebrate assemblages more characteristic of more nutrient-rich and oxygen-depleted systems. This is in line with other studies that suggest that climatic changes, soil formation processes, vegetation change and changes in lake bathymetry as lake basins fill up may lead to long-term changes in lacustrine environments such as variations in nutrient availability and oxygenation (Fritz and Anderson, 2013; Law et al., 2015). At Lac de Champex these changes were apparently at least partly related to more humid conditions developing since the Mid-Holocene, which may have promoted paludification in the catchment and changed humic acid and DOC concentrations in the lake water, leading to aquatic insect assemblages dominated by anoxia-tolerant midge larvae of Procladius und Chaoborus. Early human activities such as pasturing from the Bronze Age onwards apparently enhanced this trend, possibly by promoting nutrient release from the catchment. This is again in agreement with earlier studies in Switzerland, which showed that prehistoric human activities can promote local (e.g. Tóth et al., 2019) or even lake-wide (e.g. Haas et al., 2019; Heiri and Lotter, 2003) eutrophication and anoxia. Such shifts occurred from the Neolithic onwards in lowland lakes and as early as the early Bronze Age in lakes at high altitudes. However, our finding that preindustrial human impacts such as increasing erosion and hydrological changes such as the establishment of irrigation canals can lead to rapid re-oxygenation and re-oligotrophication is, to our knowledge, new and has not been observed before. In our case, it is difficult to differentiate in detail the relative importance of changes in oxygen availability, DOC concentrations or lake productivity in driving the observed assemblage changes. However, the different indicators and numerical analyses consistently indicate that chironomid and invertebrate assemblages from 500 to 1000 cal. BP onwards were typical for more nutrient-poor, transparent and oxygen-rich conditions than those preserved in older sediments. It is unclear how common such re-oxygenation and re-oligotrophication trends are for Alpine lakes, and we suspect that they will mainly be limited to relatively arid regions, such as the Central Alps, where irrigation practices were implemented well before the industrial era (e.g. Reynard, 2001).
Our findings are relevant for assessments of the expected natural background state of Alpine mountain lakes before human intervention. Earlier studies have shown that prehistoric and historic changes in land use, and particularly pasturing, may have eutrophied such mountain lakes as early as centuries to millenia ago (e.g. Hausmann et al., 2002; Heiri and Lotter, 2003), with a complex history of eutrophication and re-oligotrophication observed for some lakes. As a consequence it is clear that even apparently pristine remote mountain lakes may presently feature higher nutrient concentrations and more anoxic bottom waters than they would have in the absence of human intervention. However, our study from Lac de Champex demonstrates that the opposite may also be true, that in some instances modern lake ecosystem states may be characterized by chironomid and invertebrate assemblages characteristic of more oxygen-rich and nutrient-depleted conditions than would be the case under present climate without human interventions in the past. For example, it is clear that the present benthic invertebrate assemblage state at Lac de Champex, and therefore also the present limnological conditions of the lake, have developed in a period with major human modifications of the landscape and local hydrology. They are therefore a poor analogue for the natural background state that was characteristic for much of the Holocene. Indeed, the last time that Tanytarsus lugens-type, the most dominant chironomid taxon in the youngest sediments of our record, dominated assemblages in Lac de Champex was during the Younger Dryas cold period 12,000 years ago, when environmental conditions were very different from today. We therefore suggest that next to induced eutrophication and anoxia that may have occurred as early as thousands of years ago (e.g. Heiri and Lotter, 2003; Tóth et al., 2019) as well as early introductions of fish that strongly affect some mountain lake foodwebs and have been reported as much as 500 years ago in the Alps (e.g. Pechlaner, 1966), reoligotrophication and reoxygenation of lakes due to hydrological and erosional changes should be considered as a further human activity that may have preconditioned and affected apparently pristine mountain lakes, in some situations well before the industrial era.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836231211821 – Supplemental material for Human impact leads to unexpected oligotrophication and deepwater oxygen increase in a Swiss mountain lake
Supplemental material, sj-xlsx-1-hol-10.1177_09596836231211821 for Human impact leads to unexpected oligotrophication and deepwater oxygen increase in a Swiss mountain lake by Nina Perret-Gentil, Fabian Rey, Erika Gobet, Willy Tinner and Oliver Heiri in The Holocene
Supplemental Material
sj-xlsx-2-hol-10.1177_09596836231211821 – Supplemental material for Human impact leads to unexpected oligotrophication and deepwater oxygen increase in a Swiss mountain lake
Supplemental material, sj-xlsx-2-hol-10.1177_09596836231211821 for Human impact leads to unexpected oligotrophication and deepwater oxygen increase in a Swiss mountain lake by Nina Perret-Gentil, Fabian Rey, Erika Gobet, Willy Tinner and Oliver Heiri in The Holocene
Footnotes
Acknowledgements
We would like to thank Willi Tanner, Carole Adolf and Moritz Gubler for assistance during fieldwork. We thank Romain Andenmatten for support and valuable discussions on the environmental history of the Lac de Champex region, Christoph Schwörer for assistance with calculating the zonation of the record, and editor Vivienne Jones and two anonymous reviewers for valuable comments on earlier versions of this manuscript. This work has been supported by the association ‘Recherches Archéologiques du Mur d’Hannibal’ (RAMHA) and the village Liddes.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.