
Abstract
The Eurasian beaver (Castor fiber) was an important member of Early and Mid-Holocene landscapes and animal communities in Northern Europe. Previous zooarchaeological research has established the alimentary roles of beavers for Mesolithic societies and their importance for fur procurement. In this paper, we develop an integrated biocultural approach to human-beaver interactions, examining the position of humans and beavers in Mesolithic and Early Neolithic multispecies systems. We contextualize beaver landscape agency in hydroactive environments with human behaviour, synthesizing currently available data on mammalian assemblages, ichtyofauna and beaver-related material culture across Northern Europe. This cross-cultural, diachronic analysis reveals previously overlooked facilitations of human behaviour by beaver practices and ecological legacies. We show that long-term trajectories of human-beaver cohabitation differed between northern European regions. While in Southern Scandinavia, human-beaver intersections witnessed major re-organizations during the Mid-Holocene, beavers retained a key role for human societies across Northeastern Europe throughout much of the Holocene and played an important part at the Mesolithic-Neolithic transition in the Dutch wetlands. Divergent pathways are also evidenced by Mesolithic beaver-related material culture, highlighting the cultural keystone status of Castor fiber in higher latitude European landscapes. We argue that this keystone status is grounded in the supply of human hunting, fishing, and gathering affordances by the animals, pointing to the generative commensality between Mesolithic foragers and their beaver neighbours. Taken together, our findings demonstrate the importance of the beaver in the making of Early and Mid-Holocene forager societies in Northern Europe and illustrate the fruitfulness of deploying an integrated multispecies approach.
Keywords
Introduction
Beavers hold deeply ambivalent positions in the Western imaginary. Attitudes have shifted between disdain and open demonization on the one hand and fervent, heartfelt celebration on the other (e.g. Hood, 2011; Liarsou, 2015; Poliquin, 2015). The Eurasian beaver (Castor fiber) is currently making a comeback across Eurasia due to successful reintroduction and conservation programmes, yet renewed tensions are particularly prevalent in countries where beaver populations are now thriving (Halley et al., 2021; Swinnen et al., 2017; Wróbel and Krysztofiak-Kaniewska, 2020). To a large extent, the polarized recognition of the beaver as a ‘friend’ or ‘foe’ is rooted in changing perceptions of the effects of beaver activity on landscapes and wider ecologies, and as to whether and how it interferes with human lifeworlds (Liarsou, 2013, 2020). On the one hand, beavers are increasingly celebrated as ‘nature’s architects’ (Crumley, 2015) who can make significant contributions to ongoing efforts of ecosystem restoration (Gorshkov et al., 1999; Law et al., 2017; Müller-Schwarze, 2011), rewilding (Gaywood, 2018; Gow, 2020; Liarsou, 2020; Willby et al., 2018) and climate change mitigation (Lorimer, 2020), and in turn require protection (Rosell and Campbell-Palmer, 2022). On the other hand, beaver engineering can also be destructive and disruptive to human property and infrastructure (Philip, 2022), provoking the culling of flourishing beaver populations in some areas (Jansman et al., 2016; Wróbel and Krysztofiak-Kaniewska, 2020).
While some of these tensions are unique to the contemporary period and more recent history, negotiations as to how humans and beaver should co-inhabit the landscape are not. Relations between humans and beavers indeed have a deep history (Coles, 2006; Hjørungdal, 2019a, 2019b; Liarsou, 2013) and both species have crossed paths under different ecological and historical conditions, and with varying consequences. Given the recent surge of attention on beavers’ conservationist capacities, and the hope they continue to spark in the Anthropocene (e.g. Woelfle-Erskine, 2019), it is thus informative and timely to revisit the archaeology of human-beaver relations, to interrogate the long-term dynamics and legacies of human-beaver co-living, and to ask what we can learn from the respective interspecies pasts that come into view in this way, as they have been argued to bear important implications for possible and imaginable multispecies futures (Živaljević, 2021).
Recentring coupled human-beaver prehistories aligns with current attempts in archaeology to develop more inclusive accounts of the past (Hill, 2021; Hussain, 2023b; Kay and Haughton, 2019; Pilaar, 2017) and to explore the varying contributions of nonhuman animals to human pursuits and projects (e.g. Armstrong Oma and Goldhahn, 2020; Brusgaard et al., 2019; Harris and Cipolla, 2017; Hill, 2013; Hussain, 2019; Løvschal, 2022; Mannermaa, 2013; Marciniak, 2020; Oma Armstrong, 2018; Overton, 2016; Russell, 2012; Sykes, 2015). Just as animal historians have proposed to employ a dedicated ‘animal lens’ (Specht, 2016) to disclose new perspectives, insights and understandings of the past, multispecies archaeologists are now beginning to draw attention to the often-underestimated involvement of animal others in the making of prehistory (Fredengren, 2021; Hamilakis and Overton, 2013; Kost and Hussain, 2019; Pilaar, 2017). As Eitler (2014) reminds us, however, the role of animals cannot simply be cast as invariant and hence as a priori given – that is, as a mere consequence of supposedly essential species-level qualities (see also Haraway, 2003; Howell, 2018). There is a pressing need to historicize animal behaviour and affect in order to make space for the animals themselves as historical agents and contexts of action (Hussain, 2023c). To qualify the involvement of animals in human material, social, and cognitive pasts arguably requires detailed contextual analysis of historically specific conditions of interspecies encounter, negotiation, and facilitation (Haraway, 2003, 2008; Tsing, 2012), and therefore depends on multi-stranded, synthetic investigations that couple conceptual and data-driven analyses in bold yet powerful ways.
The intersection between the Eurasian beaver and post-glacial human foragers in Northern Europe offers a privileged window into such multispecies prehistory, as beavers were among the pioneer species moving into the newly available glacier-freed environments of higher latitude Europe. Beavers have previously been hypothesized to have promoted the first Mesolithic hunter-gatherers occupying the same northern landscapes (Coles, 2006; Liarsou, 2020). Furthermore, beaver remains are prevalent in many Northern European Holocene faunal assemblages, suggesting extensive but likely variable interactions between humans and beavers in this time period (e.g. Enghoff, 2011; Schmölcke et al., 2017; Zeiler, 1987; Zhilin, 2014a). Liarsou (2020: 39) estimates that beavers were between 8 and 50 times more abundant in the European Mesolithic than human foragers, illustrating that beaver activity in the landscape was probably an important lifeworld context for human behaviour. This broader ecological and archaeological background invites the exploration of how human and beaver worlds were possibly co-configured, and to throw new light on forms of life that emerged at historically specific multispecies gatherings involving both beavers and humans. We thereby contribute to better mapping out the dynamics and diversity of human-beaver relations in the past and work towards a deep-historical baseline for discussing beaver relations and impacts in the present.
From an archaeological point of view, beavers frame a particularly interesting case of human-nonhuman intersection since they provide a paramount example of a species who acts upon and alters the ecology of its human co-inhabitants and in this way intercedes with human endeavours (Hjørungdal, 2019b; Hussain, 2023c; Riede, 2019). Another reason for focussing on the beaver is because previous research on the place of animal others in earlier prehistory has mostly privileged larger mammals – what Hjørungdal (2019b) has referred to as the ‘big hunter supremacy’ bias. Beavers and their remains, when discussed, are mainly treated as economic resources, in particular in relation to their valuable pelts (see Charles, 1997 and Overton, 2016 for a similar critiques). Little attention has been paid, by contrast, to the socio-ecological dimensions of human-beaver interfaces (but see esp. Hjørungdal, 2019a; Schmölcke et al., 2017). This is despite the fact that the engagement with animal bodies is often of key importance for human social and cosmological sustenance (cf. Eitler, 2014), especially in forager contexts where hunting constitutes a society-making practice (Hill, 2019; Hussain et al., 2022; Nadasdy, 2007).
In what follows, we first outline the archaeological background of human-beaver relations in the first half of the Holocene in Northern Europe, briefly deploy a suit of concepts derived from multispecies studies and geo-sociology to address the dynamics of integrated human-beaver systems in prehistory, and then present a synthetic analysis of beaver-related material culture and beaver-anchored ecosystem relations, including human fish-getting legacies, in the Mesolithic and Early Neolithic of the Netherlands (specifically, the Dutch wetlands), Southern Scandinavia (including Denmark, Southern Sweden, and Northern Germany), and Northeastern Europe (comprising the Baltic area and Western Russia). We focus on these three macro-regions as case studies where beaver remains are prevalent in faunal assemblages and where prehistoric landscapes were conducive to facilitating human-beaver interactions, thus providing sufficient data for a cross-cultural and diachronic analysis of human engagements with beaver others. We thereby provide new evidence and arguments for the role of the beaver as a ‘cultural keystone species’ (Jacques-Coper et al., 2019) in post-glacial Northern Europe, who facilitated the expansion and consolidation of human settlement, oriented key registers of human subsistence behaviour such as mammal hunting and fishing, and catalysed regionally divergent trajectories of cultural history across the vast wetland and boreal zones of the region.
Beavers in the Early and Mid-Holocene of Northern Europe
The deep prehistory of human-beaver interactions is largely framed by interglacial climate and landscape envelopes and there is only sporadic archaeological evidence for hominin interference with beaver affairs before the final stretch of the Pleistocene, although this may in part be a research bias (cf. Cuenca-Bescós et al., 2021; Lebreton et al., 2017). Notably, beavers might have played a currently underappreciated role in the sustenance and lifeworlds of at least some interglacial Neanderthals (Hérisson et al., 2015; Kindler, personal communication; Müller and Pasda, 2011). Brown et al. (2017) have drawn attention to a possible autoecological entanglement of beavers, eels, and horses underpinning mobiliary art-making in some European Late Upper Palaeolithic contexts, and beavers have been considered as potential high-value prey items for some Late Glacial, especially Allerød, foragers in Northern Europe (Baales and Street, 1996; Charles, 1997; Weber et al., 2011). Mills (2022: 391) has pointed out that beavers were likely important agents of driftwood procurement within the extensive catchment of the Terminal Pleistocene Channel system. In Northwestern Europe, however, beaver remains and traces of their activity become more frequent only in the Early Holocene, when beavers form part of the pioneer fauna re-occupying higher latitude Europe after the Younger Dryas climatic downturn (Coles, 2006).
The impact of beavers and their geohydrological shaping of Early Holocene landscapes is well-documented in Britain (Coles, 2001, 2006). Archaeological sites such as Star Carr (Milner et al., 2018b) and Thatcham (Coles and Orme, 1983: 95–102; Evans, 1975; Overton, personal communication; Wymer and King, 1962: 88) demonstrate that beaver activity comprising woodland modification, dam and channel building, and lodging precedes the earliest Mesolithic human settlement. Beaver ecosystem modification may have thus been a locational factor in the human re-occupation of Northern Europe (Coles, 2006). In the British Mesolithic, there is for example no evidence of beaver remains outside of their habitat (Coles, 2008). This may in part be a selection bias of archaeological excavation but could also suggest that human foragers engaged with beavers mainly where they lived – in beaver country. Beaver-framed locales and landscapes may have therefore constituted vibrant ‘contact zones’ where humans and beavers were drawn together (Hjørungdal, 2019b; sensu Haraway, 2008: 216). Coles (2000) has prominently argued that Mesolithic people were probably attracted by such beaver-infused localities because of the pre-procured and ready-made wood resources of various kind made available by beaver activity at these places.
There is also evidence for beaver presence at the lakeshore environment of Järingsholm 2 in Northern Skåne, Sweden, before humans settled there in the earlier Mesolithic (Kjällquist, 2005). Hjørungdal (2019a) has further drawn attention to the interesting observation that some Early Mesolithic dwellings currently interpreted as anthropogenic resemble collapsed beaver architecture – circular structures with a simple opening. It has been suggested that because beavers and Mesolithic people were critically exposed to one another’s rhythms and practices and in some cases might have literally inhabited them, they have likely exercised mutual influence and perhaps imitated each other (Coles, 2008; Hjørungdal, 2019b; Overton, 2018). Yet this nascent perspective on human-beaver relations in the Mesolithic of Northern Europe not only requires further qualification and contextualization, it also remains a fairly marginal perspective, as archaeologists overwhelmingly continue to picture the beaver as ‘good to eat’ and/or ‘good to use’, foregrounding the animals’ supplementary caloric value and role in early fur-getting economies (e.g. Price, 1991; Zhilin, 2014a, 2014b). This especially holds true for the interpretation of the abundant beaver remains recovered from continental Northern Europe. While the socio-ecological facets of human-beaver interactions have been comparatively well-studied for prehistoric Britain (Coles, 2006; Overton, 2014, 2016, 2018; Wells et al., 2000), the same can certainly not be said for continental Northern Europe, nor for higher-latitude Europe as a whole.
Osseous remains of beavers are a recurrent feature of the Mesolithic across continental Northern Europe, encountered from the Netherlands in the Northwest via Denmark, Sweden, and Northern Germany to the Baltic and Western Russia in the East (e.g. Groß, 2017; Lõugas et al., 2017; Price, 1985; Schmölcke and Nikulina, 2015; Zeiler, 1987, 1997; Zhilin, 2004). In the Northeastern Mesolithic, beaver remains in many cases quantitatively make up a substantial portion of the recovered faunal assemblages and are often only outnumbered by elk (Alces alces) bones (Lõugas et al., 2017; Zhilin, 2014a). Similarly, in the Dutch wetlands, beaver remains are frequent in faunal assemblages from the Late Mesolithic and Early Neolithic, together with other freshwater fauna, such as otter (Lutra lutra), wild boar (Sus scrofa), waterbirds and fish (Çakirlar et al., 2019; Lauwerier et al., 2005). In Southern Scandinavia, by contrast, beaver remains tend to be few in number and the handful of later Mesolithic assemblages with higher counts of beaver bones are typically interpreted as specialized hunting or trapping stations linked to delayed-return extraction systems (Rowley-Conwy, 1998). Enghoff (2011: 295) notes a geographical pattern in the Danish material, with beavers being considerably more common at Mesolithic sites on Zealand than on their counterparts on the Jutland peninsula, and Schmölcke et al. (2017) have suggested that beavers were of little economic relevance in this part of Europe given the generally small number of sites with significant beaver bone shares.
Zhilin (2020) has recently also drawn attention to the extensive record of organic tools made of beaver remains including mandible and incisor tools that form an integral part of the larger Mesolithic interface within the Eastern European forest zone. Similar beaver-procured tools have been reported from a few Mesolithic sites in Northern Germany (Schacht and Bogen, 2001; Schmölcke et al., 2017; Schuldt, 1961), the Netherlands (Coles and Kooijmans, 2001) and Denmark (Broholm, 1924: 133; Hatting, 1970), but previous work has paid surprisingly little attention to this beaver-related material culture and its possible significance (but see Hjørungdal, 2019a). Although ethnographic parallels have been invoked to interpret beaver-sourced tools, scholars have mainly highlighted the functionality of these objects and the capacity of Mesolithic people to make use and take advantage of the animal materials available to them. Hatting (1970: 126) for example symptomatically concludes that ‘it must [thus] be held with certainty that the idea of this kind of tool making was known in the Stone Age of Denmark’. In the Mesolithic of Northeastern Europe, beaver body-parts such as teeth and ankle bones sometimes also made their way into human burials (see e.g. Grünberg, 2013; Kashina et al., 2021; Mannermaa et al., 2021) yet, again, this material has so far hardly been considered and further contextualized, partly also because Mesolithic scholarship has focused on other, supposedly more symbolically potent nonhuman animals such as various bird and deer species, but also suids, and their role in Mesolithic identity construction (e.g. Bridault, 1992; Conneller, 2004, 2011; Kashina, 2005; Kashina and Zhulnikov, 2011; Lozovskaya, 2021; Mannermaa, 2013; Mannermaa et al., 2019; Price, 1985; Tilley, 2003; Zagorska et al., 2018).
In the extensive wetland and boreal zones associated with the earlier Holocene in Northern Europe, beaver remains appear in archaeological sites linked to the Preboreal, Boreal and early Atlantic chronozones, and generally span the whole period, from the earlier Mesolithic to the later Mesolithic and earliest Neolithic, even though some notable spatiotemporal patterns can be discerned. For the purposes of this paper, we distinguish between three chronocultural phases in Northern Europe (see Table 1 for an overview of the corresponding chronozones and time ranges). For the Baltic and Western Russia, we follow the literature (Hartz et al., 2010; Lõugas et al., 2017; Zhilin, 2020) and subdivide the period into an ‘Early Mesolithic’ phase including complexes such as Early Kunda and Butovo, a ‘Middle Mesolithic’ harbouring the later Kunda and Botovo phases, and a ‘Late Mesolithic/Early Neolithic’ consisting of the Mid-Holocene Early Comb Ware complex and featuring entities such as Narva and Valday. The same chronological scheme is applied to Northwestern Europe, where the Early Mesolithic comprises the earlier phase of the Maglemose complex as recently defined for Southern Scandinavia, including Northern Germany, and Britain (Milner et al., 2018a; Sørensen et al., 2018). The Middle Mesolithic is more-or-less synonymous with the later phase of Maglemose complex (Groß, 2017), and the Late Mesolithic/Early Neolithic accommodates the Dutch Swifterbant Culture (Dreshaj et al., 2023; Raemaekers and De Roever, 2010) and the Ertebølle complex (EBK) in Southern Scandinavia (Price, 2000, 2015).
Three-stage chrono-cultural scheme of the Northern European Mesolithic adopted in this study.

We collected a comprehensive sample of accessible faunal datasets from these periods. We recorded the Number of Identified Specimens (NISP) for fish and mammal species for all regions and periods, except for the Early and Middle Mesolithic of Southern Scandinavia where only Minimum Number of Individuals (MNI) was readily available (Supplementary Information M and F, available online). Both NISP and MNI introduce their own biases to the interpretation of the zooarchaeological record, as has been extensively discussed elsewhere (Domínguez-Rodrigo, 2012; Gifford-Gonzalez, 2018; Grayson, 1984; Lyman, 2008, 2019; Marshall and Pilgram, 1993). While comparing assemblages recorded by NISP with those recorded by MNI poses some limitations, we are primarily concerned with broad patterns in the (zoo)archaeological record and taxonomic abundances. The focus therefore lies less on absolute numbers and percentages and we instead concentrate on relative broad-scale similarities and differences, keeping these limitations in mind. It is nevertheless clear that both NISP and MNI can be severely impacted by taphonomy (Gifford-Gonzalez, 2018; Grayson, 1984; Lyman, 2008: 187–190), albeit Marshall and Pilgram (1993) have shown that they are in most cases equally sensitive to fragmentation and that MNI tends to be less representative only at very high levels of assemblage fragmentation.
For the Netherlands, Northeastern Europe and the earlier part of the Mesolithic in Southern Scandinavia, we recorded all readily available archaeological sites. For the Ertebølle complex, we had to make a selection of sites due to the sheer number of relevant archaeological instances. We selected only sites documenting >1000 NISP of mammals supplemented by sites with fish remains >1000 NISP, which assures a relatively representative sample, as suggested by Gron (2013).
This provides a total sample of 116 archaeological sites with faunal information (mammal and fish remains) encompassing modern day Denmark, Sweden, Northern Germany, the Netherlands, Poland, Latvia, Lithuania, Estonia and Western Russia (with the Urals delineating the study region in the east). We additionally review, synthesize, and re-contextualize relevant instances of beaver-related material culture published from across the study region.
Against this archaeological background of human-beaver engagement in the Northern Mesolithic, we attempt a new synthesis and offer a re-interpretation of the record based on multispecies thinking, stressing the importance of human-beaver entanglements as a history-making dynamic. Before doing so, we briefly outline our conceptual point of departure, deploying a ‘beaver lens’ and shifting attention to the assembly and long-term development of human-beaver systems in the past. This perspective, we argue, supplies the necessary conceptual resources to re-examine the potential formative but context-dependent role of beavers throughout the earlier part of the Holocene in Northern Europe.
Integrated human-beaver systems as a generative dynamic of European prehistory
Multispecies thinking challenges overly human-oriented approaches to the archaeological record (Boyd, 2017; Hamilakis and Overton, 2013; Hill, 2021; Kay and Haughton, 2019; Kost and Hussain, 2019), as the various worlds inhabited by past people are recognized as fundamentally co-constituted by a broad array of possible beings and entities, many of which are nonhuman (Harris and Cipolla, 2017; Pilaar, 2017). Although the contribution and influence of these variegated denizens of the past tends to differ vastly and is so highly context-dependent, it is their ‘becoming-with’ – to speak with Haraway (2003, 2008) – that calls for particular archaeological attention. The multispecies past in this way becomes a story of the diversity of life and ways of being, including the situated struggles and possibilities that emerge at the interstices and intersections of species. Rather than emphasizing capacities and concerns of individual beings and isolated actors, multispecies archaeologies highlight the creative potential for stability and change that emerges from the orchestration of life in all its heterogeneity. In Haraway’s (2016: 58) terms, the past is sympoietic, and hence lived and made ‘in company’. Tsing (2021) similarly maintains that ‘[s]taying alive – for every species – requires liveable collaborations’, necessarily entailing ‘working across difference’. The challenge for coming to terms with multispecies pasts, therefore, is to recognize and theorize difference across relevant actors and species while nonetheless not losing sight of their historical malleability and inter-relationality. Which species meet and under which conditions is crucial for what becomes possible – social, material or otherwise – and for what matters – both for people and their nonhuman interlocutors. This principle of ‘horizontality’, which is now increasingly recognized as a conceptual key across the environmental humanities (De Carvalho Cabral, 2021), symptomatically, also begins to feature more and more prominently in life-oriented accounts of natural evolution (Corning, 2005; Margulis, 1998; Walsh, 2015).
Working through beaver difference requires to foster a ‘beaver lens’ and to look at the past from the perspective of beaver practices and autoecologies. Beavers have been qualified as ‘nature’s architects’ (Crumley, 2015) because they build structures such as lodges and dams and engage in earth-working by digging canals and gullies. Their woodworking activities alter the dynamics of vegetational successions, thin out forest patches and create openings in the landscape. Beavers are potent ‘niche constructors’ and ‘ecosystem engineers’ (Brazier et al., 2021; Johnston, 2017), so that their continued presence and varied activities within a landscape quickly develop significant impact on the broader ecosystem. Beavers are agents of disturbance (Tape et al., 2018) and through their interference with hydrological and geomorphological systems foster dynamic wetlands (Crumley, 2015; Liarsou, 2015; Poliquin, 2015). Practices such as dam, lodge and canal-building change water regimes and create new habitats such as ponds of varying size and microecology as well as extended littoral zones, which support a diversity of aquatic and terrestrial life (Larsen et al., 2021), thus reconfiguring ecosystem structure and functioning in riparian landscapes across diverse spatiotemporal and ecological scales. By altering freshwater physical habitat, biotic composition and habitat connectivity (Macfarlane et al., 2017), this beaver-promoted regime change facilitates the ‘invasion’ of new species such as riverine plants, a range of invertebrates and various fish (Bunn and Arthington, 2002).
Notably, beaver-engineered freshwater habitats experience local shifts towards anaerobic biochemical cycling and nitrogen accumulation and encourage higher rates of sediment, nutrient and detritus trapping (eutrophication: Krylov, 2002; Naiman et al., 1986). The result is often an associated shift from lotic (moving water) to lentic (standing water) fish communities (Larsen et al., 2021) and the sustained promotion of species thriving under eutrophic conditions (O’Hare et al., 2018), including plants such as Nymphaeaceae (water lilies). As opposed to rivers, sustained beaver-activity in lakes does not tend to substantially change aquatic species composition but conserves lentic assemblages and generally increases habitat quality, probably incurring currently little known but likely extensive long-term legacy effects (Bashinskiy, 2020). Dynamic beaver-powered wetlands attract and promote waterbird breeding and diversity (e.g. Nummi and Hahtola, 2008; Nummi and Holopainen, 2014; Nummi and Poysa, 1997) and have been shown to increase localized mammal richness (cf. Gauvin et al., 2020; Wright et al., 2002). Beavers co-regulate biodiversity, often locally increasing it (Law et al., 2019; Stringer and Gaywood, 2016), and they typically complexify ecological networks and interactions where they are active in the landscape (Naiman et al., 1986). Fedyń et al. (2022) have shown that beaver-impacted habitats in temperate forest environments form hotspots of seasonal mammalian aggregation, especially for small and large carnivores but sometimes also deer. Beaver-modified landscapes therefore harbour resource patches of high potential foraging value and predictability.
Because of these disproportional ecological impacts of beaver practices measured by the overall abundance of the animals in the landscape, the beaver emerges as a ‘keystone species’ in riverine and boreal environments (Janiszewski et al., 2014). Beavers are a resident species yet can rapidly colonize new landscapes when these become available for them (Swinnen et al., 2017), and they also commonly abandon lodges when water and wood supply become insufficient. Because of this dynamic, beaver activity tends to inscribe itself into the life-history of its host landscapes and beaver effects can be charted through four broad stages of impact, from the initial flooding of river banks after colonization to the formation of legacy meadow complexes after site abandonment (Polvi and Wohl, 2012; Westbrook, 2021). Based on satellite time-series data from the Canadian Artic, Tape et al. (2018) have established that beaver colonization and its associated landscape transformations not only happen on the scale of merely decades, but also accelerate climate change as beaver landscape engineering promotes permafrost thawing, inter alia contributing to thermokarst formation. Beavers are thus likely a key agent of coupled climatic, environmental, and geomorphological change at the Pleistocene-Holocene transition in Northern Europe, where the retreating glaciers provided ideal, hydroactive habitat for incoming beaver populations, conditions that were subsequently reinforced by beaver activity.
The ecosystem impacts of the beaver not only frame the animal as a prominent and dynamic agent in the environment, potentially garnering special human attention, some of the above enlisted consequences of beaver behaviour enact profound ‘ecosystem services’ (sensu Balvanera et al., 2017) for foragers inhabiting the same landscapes. Beaver-propelled landscape dynamics cater alluring possibilities and affordances for human life, while human behaviour in turn may similarly shape the action-space of beavers – for the better or worse. This coming together, and possible coordination, of human and beaver practice in situated historical contexts as ‘becoming-with’ can be examined as a systemic articulation likely involving assembly, consolidation, and disintegration. Following Schroer (2022), we can explore such articulations from the perspective of a general ‘geosociology’. Sociality, in this view, emerges from the ‘geopraxis’ of all life as it contributes to the creation of serviceable living spaces, shared lifeworlds and efforts of world-building in general. All animals, accordingly, ‘organize, constitute and inform’ the world in their own way (Castoriadis, 2010: 156), and beavers are beings par excellence who directly act upon the geo- and biosphere and by means of the incurring consequences co-constitute the sociality of other species, including humans. Geopraxis, then, as performed by beavers, can become a life-service supporting the sustenance of humans in the landscape as well – a service then primarily conditional on two factors: (i) human practice must not undermine beaver ecosystem engineering, for example through overhunting; and (ii) human life must be predicated on practices and modes of occupation that can in principle benefit and take advantage of beaver practice or even rely/depend on it (see Liarsou, 2013, 2015 for similar arguments). By analysing and comparing different human-beaver systems in time and space, these issues can be scrutinized and addressed empirically and supported by data-driven analyses, yielding important insights for the possibilities and challenges of present and future human-beaver co-living.
Mustering a ‘beaver lens’ in this way allows to tackle the historicity and contingency of human-beaver systems. Importantly, the human acceptance and cultivation of beaver-provided life services can provide ‘impetus for new practices of multispecies hospitality and conviviality’ (Rigby, 2020: 110), and may thus ultimately lead to novel or unique but frequently tangible, archaeologically observable forms of beaver-related socio-material negotiation. As Eitler (2014) points out, animal materialities and materializations should never be approached as mere products or representations of their associated human-animal relationships but instead as partaking elements in the ongoing production of these relationships. Materializations such as prehistoric animal-rendering visual culture or animal-sourced tools are in this view misunderstood as a mere ‘output’ of either culture, cognition or ecology, or as representational forms (representationalist fallacy). These materializations rather help to generate and secure relations of ‘becoming-with’ and variously crystallize and/or synthesize ‘natural’ and ‘cultural’ conditions of specific human-animal arrangements. Haraway’s (2008: 216) notion of the ‘contact zone’, inspired by Pratt’s (2008) original human-oriented concept, aligns with this view, highlighting that historical subjects are never given, but always ‘constituted in and by their relations to each other’, which are in turn devised in terms of co-presence, intra-action, and interlocked understandings and practices, although typically within unbalanced power-relations. From such ‘contact zones’, then, historically specific forms and systems of ‘conviviality’ can emerge.
Conviviality here describes the ability of humans to interact creatively and autonomously with nonhumans in their environment and to satisfy their needs while thriving in the social company of these nonhuman others, thus framing prolific modes of multispecies cohabitation (Rigby, 2018, 2020; Straughan et al., 2022). Conviviality is not to be confused with living-in-harmony with nature and nonhuman others, however, and does therefore not re-introduce or revive the problematic notion of the ‘noble savage’. Conviviality is nevertheless often expressed in the adoption and cultivation of practices that enable and promote species co-living – frequently despite or because of tension and conflict – and these practices are often materially generative and foster heightened attentiveness to others’ presences, affordances, and life services. Such attentiveness can be directly investigated archaeologically, for example through the many ways animal materialities including bodies and material culture were handled by past people. Systems of conviviality are thereby expected to vary considerably in time and space, and not all human-animal systems promote long-term or even short-term conviviality. Convivial systems may even lay foundation to what is sometimes referred to as the ‘multispecies commons’ (Bresnihan, 2015; Centemeri, 2018; Haldrup et al., 2022; Satsuka, 2014) – a notion recently also mobilized by Woelfle-Erskine (2019) to frame human-beaver collaboration in the context of ecosystem restoration and stewardship. To what extent integrated human-beaver systems in the earlier part of the Holocene of Northern Europe may be qualified as an expression of deep-time conviviality and interspecies ‘commoning’ is an open but increasingly relevant question and will be empirically explored in the following.
A new synthesis of human-beaver intersections in the Northern Mesolithic
This section draws together the available evidence on (1) beaver-bearing mammalian faunal assemblages, (2) beaver-related material culture, and (3) fish assemblages (ichtyofauna) on a sub-continental scale and outlines macro-patterns in the study regions. This large-scale inventory of archaeological data from the Mesolithic and Early Neolithic of Northern Europe enables the subsequent discussion of beaver impacts on local ecologies, the possible provisioning of specific human foraging niches, and the cultural significance of beavers and their bodies and materialities. We pay particular attention to ichtyofaunal patterns and ecological successions because beavers, as discussed above, are known to engineer novel freshwater habitats and modulate broader dynamics within freshwater ecologies. The configuration of fish communities has so far also received comparable little attention when discussing the impacts and legacies of beaver geopraxis in the wider landscape and would arguably have a profound effect on forager lifeways as site assemblages in this region demonstrate the significant contribution of fish to human subsistence.
Macro-patterns in zooarchaeological beaver assemblages
In contrast to the Terminal Pleistocene of higher latitude Europe where beaver remains are rarely encountered in archaeological contexts (cf. Veil et al., 2012; Weber et al., 2011: 661), the Eurasian beaver becomes an important and recurrently encountered component of faunal assemblages from the Early and Mid-Holocene of Northern Europe (Figure 1 and Supplementary Information M, available online). We summarize the available zooarchaeological data in relation to the three delineated macro-regions, paying particular attention to changing beaver abundances and faunal co-associations.
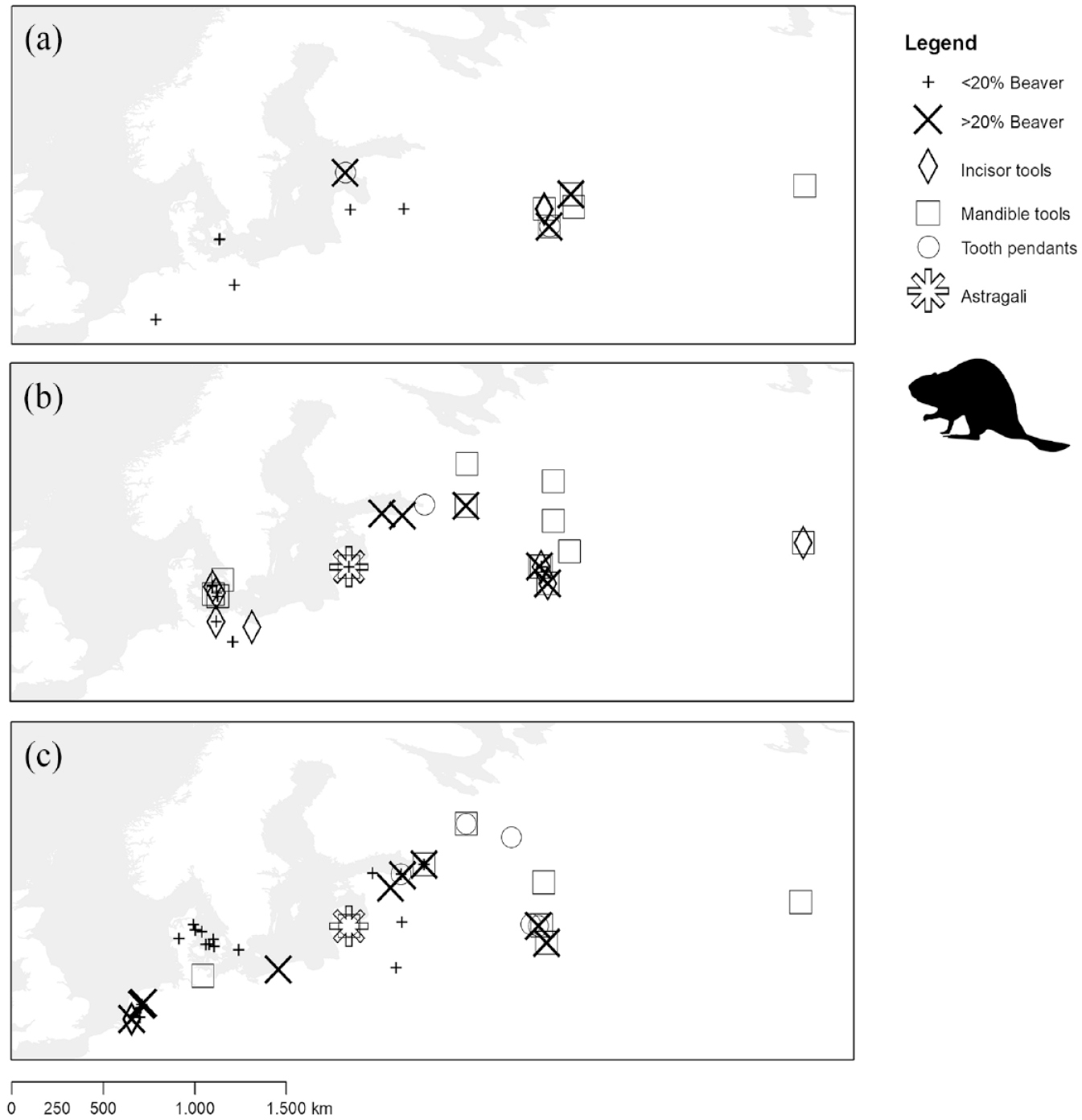
Overview of key sites from the Northern Mesolithic and associated beaver remains as well as beaver-related material culture: (a) Early Mesolithic; (b) Middle Mesolithic; (c) Late Mesolithic/Early Neolithic. Data provided in Supplementary Information 1, available online.
Northeastern Europe
The Early and Middle Mesolithic in the Baltic and the Russian West are represented by the Early Kunda, Veretje and earlier Butovo complexes (Damlien, 2016; Hartz et al., 2010; Manninen et al., 2021; Zhilin, 1996), while the Late Mesolithic/Early Mesolithic comprises Late Kunda, later Butovo and Janislawice as well as the regional variants of the emerging Early Comb Ware complex (Oras et al., 2017; Piezonka, 2021; Zvelebil, 1994). Site types and conditions of archaeological recovery vary greatly between and within regions. In the earlier Mesolithic, sites are mainly located above river and lake terraces or in extended wetlands and are often represented by small flint scatters or mixed deposits, while in the later part of the Mesolithic settlement systems become more structured and a duality between habitation and extraction sites as well as between coastal and inland occupations begins to take shape (Piezonka, 2021). For the Russian West, well-preserved faunal assemblages are mainly associated with the many bog and wetland sites of the Volga-Oka interfluve (Zhilin, 1996, 2004, 2007, 2014b), and the record may thus be biased towards specific localities and forager activities in the landscape.
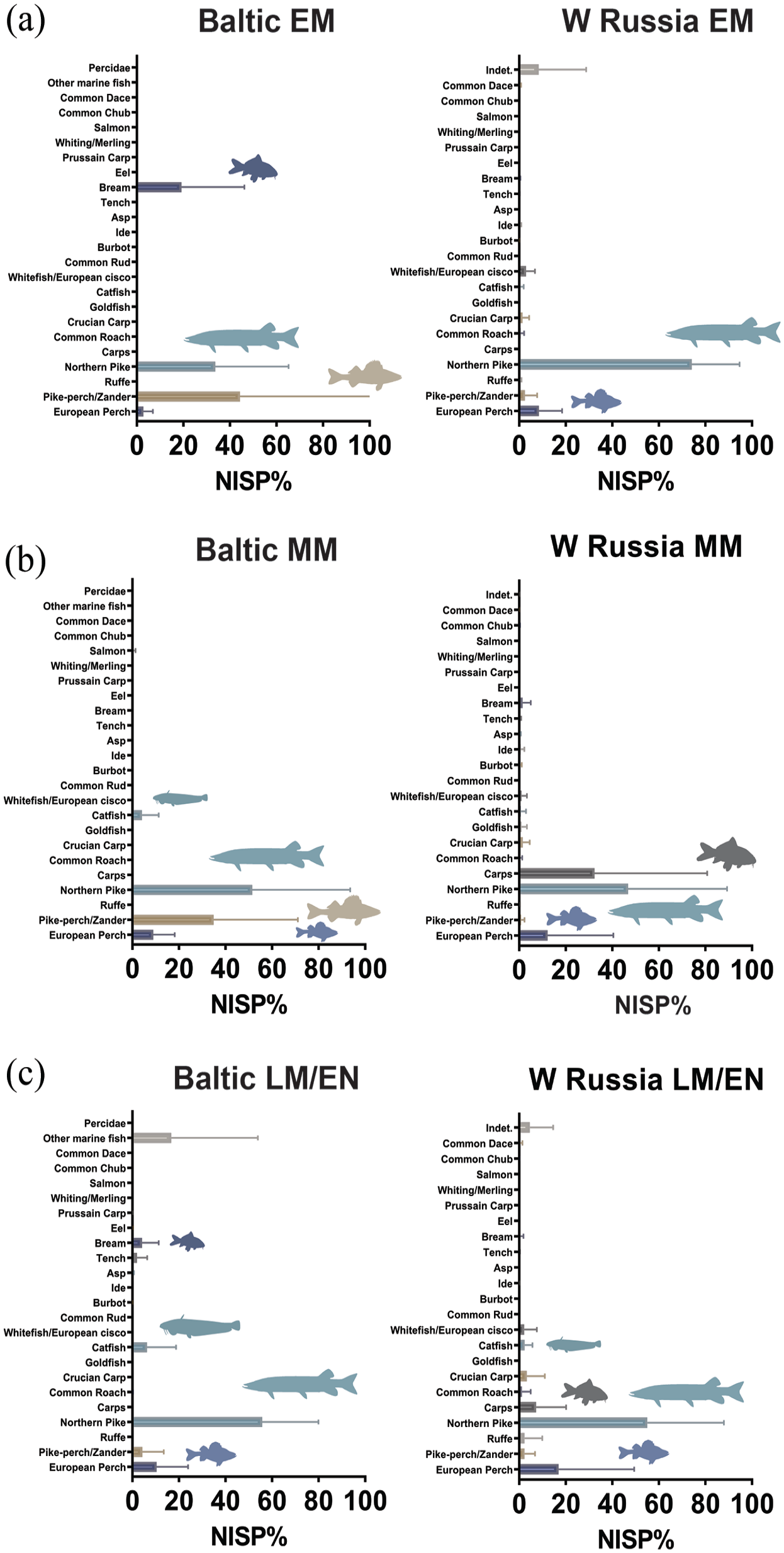
A directly dated beaver-gnawed piece of wood from the important Ivanovskoye peat bog dated to around 10,000 years ago (Zhilin, 2019) provides evidence for prolonged histories of human-beaver co-residence in the Russian Northwest. In the Early and Middle Mesolithic, beaver makes up between 5% and 47% of total mammalian NISP in the Baltic and between 19% and 36% in Western Russia, while the Late Mesolithic/Early Neolithic is characterized by beaver NISP frequencies between 3% and 61% in the Baltic and between 9% and 34% in Western Russia (Figure 2). In the Early Mesolithic of Western Russia and the Baltic, beaver is often the second-most frequent mammal after the elk, pointing to the beaver’s economic relevance and, possibly, elevated abundance in the ecosystem. Muskrats (Ondatra zibethicus) and martens (Martes sp.) are also an important faunal component in this spatiotemporal context.
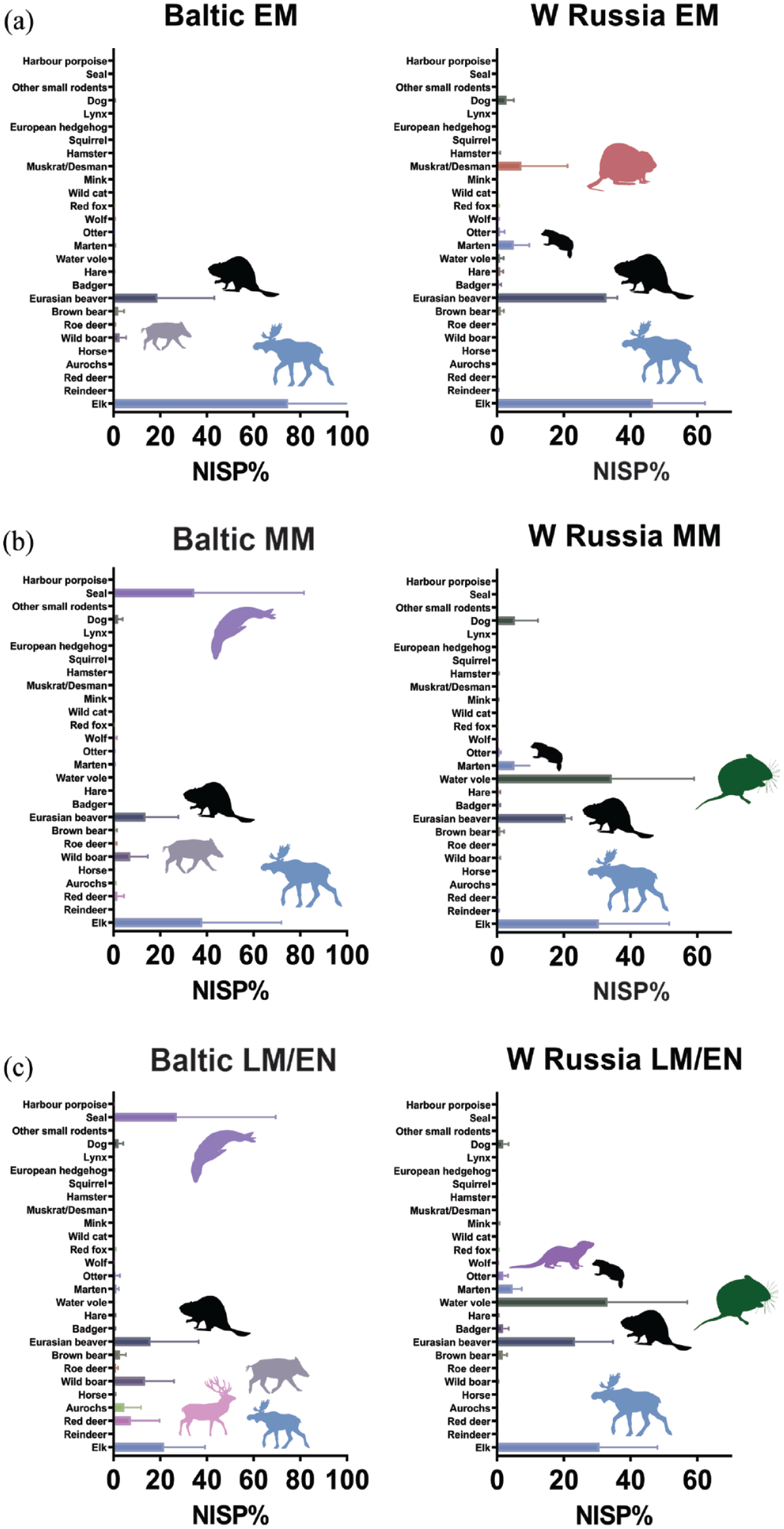
Number of identified mammal specimens (NISP%) from the Baltic countries and Western Russia from the Early to the beginning of the Mid-Holocene: (a) Early Mesolithic; (b) Middle Mesolithic; (c) Late Mesolithic/Early Neolithic). Horizontal bars represent median values with standard deviations. Animal silhouettes have been redrawn from https://www.phylopic.org/. Raw data are provided in Supplementary Information M Tables 1 and 2, available online.
From the Middle Mesolithic onwards, the importance of elk diminishes in the wider area, while wild boar and red deer (Cervus elaphus) become increasingly important in the Baltic and the water vole (Arvicola amphibius) in Western Russia. Interestingly, the increasing importance of water vole in the Northeastern inland Mesolithic, reaching up to 61% of recorded NISP at individual archaeological sites, corresponds to a general trend of decreasing faunal representation of the beaver during the Middle and Late Mesolithic of the region, and this may be related to long-term beaver-modulated changes in riverine-lakeland habitats, strongly promoting water vole populations on a local scale.
In the Baltic, beaver frequencies are overall less stable across archaeological sites and the pattern is more punctuated than in Western Russia: some sites harbour fairly low beaver abundances (c. 3–7% of NISP), while others exhibit increased beaver yields (c. >20–60%), pointing to functional differences in site formation and perhaps more diverse human-beaver interactions. The Late Mesolithic/Early Neolithic site of Dąbki 9 in Northern Poland, for example, has been argued to represent a seasonal special purpose locality mainly geared towards fur-extraction and possibly exchange (Schmölcke and Nikulina, 2015). It is notable that among the other species from Dąbki 9, the otter makes up c. 6% of the documented mammalian NISP, thus representing the third-most frequent animal in the whole faunal assemblage. Although otter remains are not generally frequent at beaver-bearing sites, they are often a common yet low-abundance feature. This association is conspicuous since otters and beavers are known for their sympatric relationships, as otters benefit from beaver-engineered and disturbed habitats, especially in riverine higher latitude woodland environments (LeBlanc et al., 2007; Reid, 1984; Tumlison et al., 1982).
The possible presence of muskrat (NISP = 28%) alongside beavers (NISP = 30%) in Early Mesolithic Stanovoye 4 at the Volga-Oka interfluve in Northwestern Russia (Zhilin, 2004) is notable and may similarly indicate that human foragers took advantage of the facilitative effects of beavers on their animal co-inhabitants (cf. Crego et al., 2016).
Northwestern Europe
The Early and Middle Mesolithic of Southern Scandinavia is characterized by the Maglemose complex (9500–6400 BC) and the earlier part of Kongemose (6800–5400 BC), known in particular from the many well-preserved bog sites across Denmark and Northern Germany. Two types of sites are known from the Early Mesolithic: so-called deposition sites and habitation sites (Sørensen et al., 2018). Well-preserved faunal material only derives from deposition sites in Southern Scandinavia, while in Northern Germany habitation sites also occur (Sørensen et al., 2018) – likely a consequence of site preservation. The respective lithic and faunal assemblages are often very small and fragmented, indicating short-term visits. The Middle Mesolithic sites from Southern Scandinavia as a whole are well-preserved, derive from wetland deposits, and have mostly been described as habitation sites (‘base camps’: Gramsch, 2000; Groß, 2017; Schuldt, 1961: 185), but we may generally miss the more ephemeral sites from the period. Early and Late Maglemose sites from Jutland and Eastern Denmark show patterned differences in preservation and depositional context (Blankholm, 1996; Nielsen, 2006). In the Netherlands, only a couple of archaeological sites with faunal remains are known from the Early and Middle Mesolithic. The zooarchaeological assemblages are small and beaver remains number only a handful, making the comparison of Northeastern Europe and Southern Scandinavia difficult for the Early Holocene.
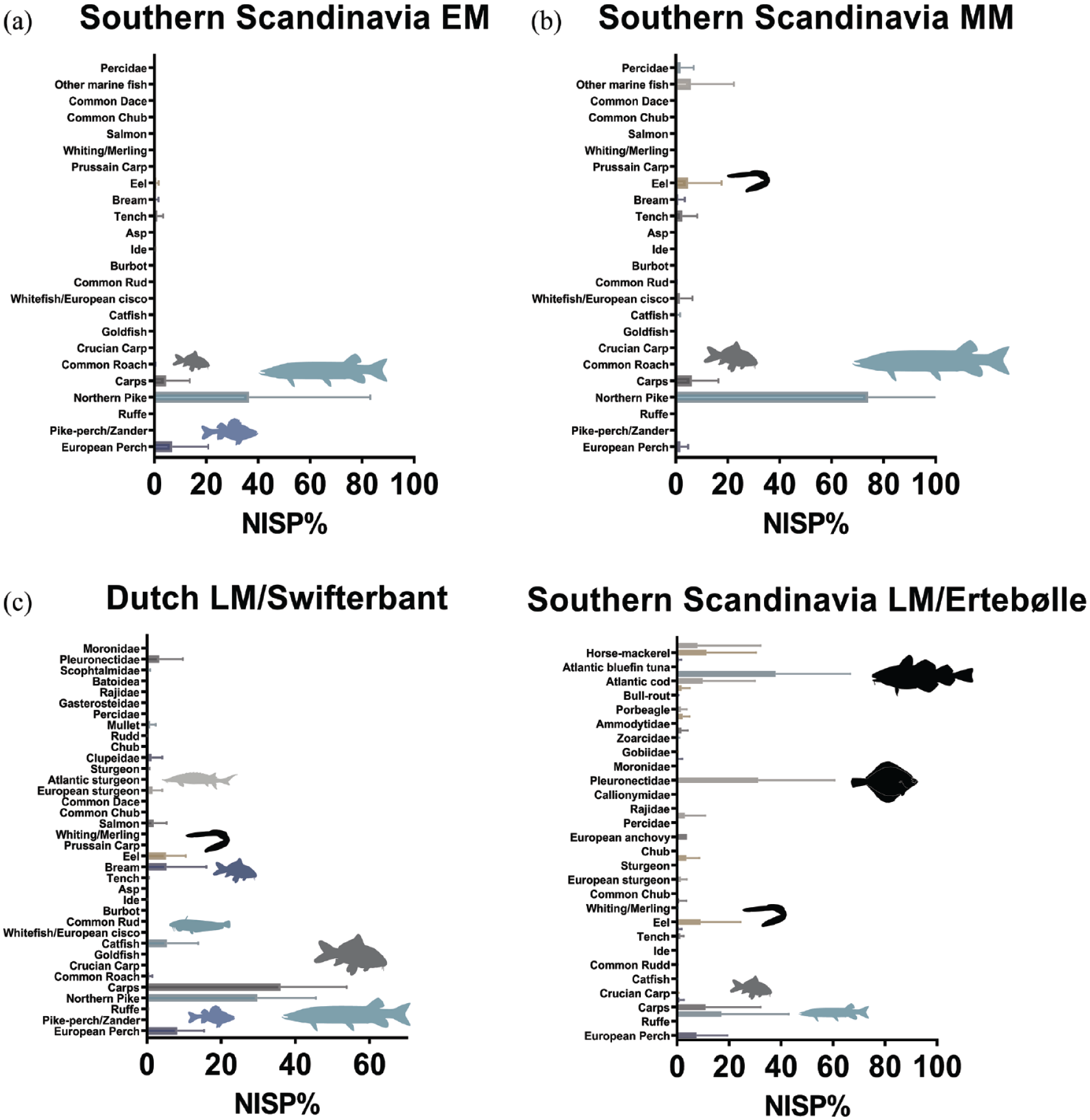
In Early Mesolithic Southern Scandinavia, beaver remains make up an average of 8% of total MNI, ranging between 5% and 14% (Figure 3). Most assemblages from this period are very small (<50 total MNI), making it difficult to draw firm conclusions from species compositions. At Sværdborg, where several excavations have yielded larger faunal assemblages, beavers are clearly present (MNI between 5% and 9%), as are other fur-bearing mammals such as otter (5–8%) and badger (Meles meles) (2–5%). In general, otters are relatively common in the Early Mesolithic of Southern Scandinavia (2–33% of MNI), supporting the observations made for Northeastern Europe. Overall, however, there is a greater emphasis on large fauna, such as wild boar and roe deer (Capreolus capreolus). Despite the noted differences in faunal preservation for habitation and deposition sites between Southern Scandinavia and Northern Germany, there is no apparent divergence in beaver MNI representation.
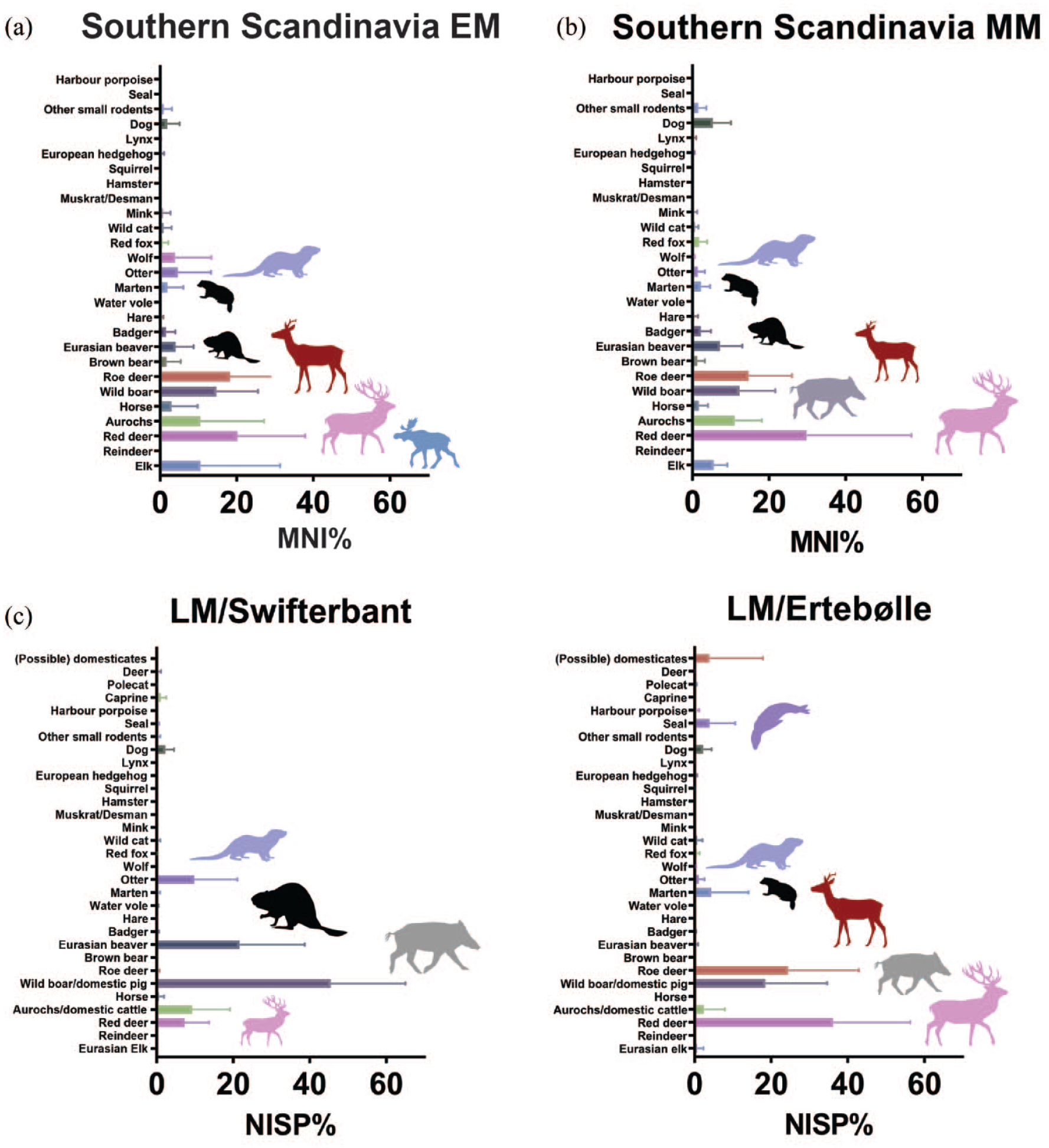
Number of identified mammal specimens (NISP%) or minimal number of individuals (MNI%) from Southern Scandinavia and the Dutch wetlands from the Early to the first part of the Mid-Holocene: (a) Early Mesolithic; (b) Middle Mesolithic; (c) Late Mesolithic/Early Neolithic. Note that data for the latter region is only available for the final phase and that MNI is used in (a) and (b). Horizontal bars represent median values with standard deviations. Animal silhouettes have been redrawn from https://www.phylopic.org/. Raw data are provided in Supplementary Information M Tables 3 and 4, available online.
In the Middle Mesolithic faunal assemblages from Southern Scandinavia, beaver remains continue to be present, but in consistently small numbers. They average c. 9% of total mammalian MNI, ranging between 2% and 18% respectively. Larger mammals are again found in greater numbers, but, as noted above, most of the respective assemblages are too small and fragmented to draw broader inter-site conclusions.
The Maglemose and Kongemose complexes are succeeded by the Ertebølle Culture (EBK; 5400–3950 BC), identified first and foremost by its famous coastal shell-midden sites. The EBK represents the end of the Mesolithic in Southern Scandinavia and many sites reflect transitionary phases to the Funnel Beaker Culture (TRB) marked by the appearance of domesticated animals and cereals (cf. e.g. Gron and Sørensen, 2018). EBK zooarchaeological assemblages derive from a variety of site contexts, such as bogs, submerged/waterlogged sites and shell middens, introducing systematic taphonomic and recovery biases (Gron and Robson, 2016). Beavers decrease to an average of only 0.4% of total NISP in EBK, with values ranging between 0% and 2% at individual sites (cf. Figure 3). Other fur-bearing animals, in particular marten and wild cat (Felis silvestris), make up a more substantial part of the faunal assemblages than beavers, while large wild fauna continue to be important. In EBK, marine mammal shares increase significantly in abundance and together with marine fish become a key part of the human diet (Rowley-Conwy, 1999).
In the Netherlands, the Late Mesolithic/Early Neolithic has produced a wealth of faunal data from Swifterbant Culture (ca. 5500–3400 BC) sites in the Dutch wetlands. The Swifterbant Culture has long been considered a transitionary archaeological complex at the forager-farmer interface, but recent evidence demonstrates that from at least 4250 BC on these communities had established agricultural practices while also relying on diverse natural resources (Brusgaard et al., forthcoming; Huisman and Raemaekers, 2014; Raemaekers et al., 2021). Swifterbant Culture sites are all habitation sites, some occupied seasonally and some on a year-around basis. In the Early Neolithic, Linearbandkeramik (LBK) farming communities also inhabited the southernmost zone of what is now the Netherlands, but faunal remains exist from only one site and are very few (eight total identified mammal NISP), so we have to restrict the discussion to the Swifterbant Culture in this region.
Beaver remains are altogether abundant at Swifterbant Culture sites, averaging c. 20% of total NISP, but the numbers are highly variable between sites, ranging between 0.4% and 49%. Beaver remains are predominant at the oldest sites, Hardinxveld-Giessendam Polderweg and De Bruin (5500–4250 BC) (34% and 49% of total NISP, respectively) and Schokland P14 (4900–3300 BC) (37%), where they rival or even outnumber larger ungulates such as wild boar and red deer. At the Hardinxveld-Giessendam sites, the total percentage of beaver NISP masks the notable increase in beavers over time, reaching frequencies of 83% at Polderweg and 51% at De Bruin in the final occupation phases, while the number of large fauna specimens decreases (Oversteegen et al., 2001; van Wijngaarden-Bakker et al., 2001). This is possibly a result of increasingly wet conditions due to sea level rise in the area – conditions in which beaver would have flourished and foragers may have become increasingly reliant on aquatic resources (Brusgaard et al., 2022a).
At the Swifterbant type sites S2, S3 and S4 (4300–4000 BC), where domesticated livestock are also present (Brusgaard et al., in press; Zeiler, 1997), beaver remains make up between 12% and 23% of total NISP. Beaver-gnawed willow branches were found at S3 (Casparie et al., 1977) and Prummel (2017) has recently highlighted that the Swifterbant environment would have been ideal for human exploitation due to beaver engineering. In contrast, at the archaeological sites Tiel, Hoge Vaart and Nieuwegein, beaver represents less than 5% of the total NISP, and there is more emphasis on large wild fauna. Otter remains fluctuate almost on par with beaver remains at each site, ranging between 29% at Hardinxveld-Giessendam Polderweg (where beavers are abundant) and <1% at Tiel (where beavers are few). The only exception to this pattern is Schokland where the NISP of beaver is high but only few otter remains were recovered.
From an environmental perspective, the prevalence of beaver remains at many of the Swifterbant Culture sites is not surprising considering their location in freshwater riparian landscapes. The Dutch data discussed here is generally biased towards such locations because faunal remains from archaeological sites on the sandy (drier) soils have not been preserved and no coastal sites are known, probably due to erosion (Vos, 2015). It is therefore presently unknown which wild fauna was exploited in these other regions and ecotones. While the presence of beavers at Mesolithic and Early Neolithic wetland sites is thus not unanticipated, the relative abundance of beaver remains points to some form of concentrated exploitation of these animals (and of otters) by Swifterbant communities. The main synchronic and diachronic trends in beaver remains appear to be negatively structured by the frequency of large prey animals such as cervids and wild boar, suggesting a strategic trade-off between either beaver and otter or the targeting of such larger species. There is, however, no evidence for a switch to other fur-bearing animals such as martens, as often observed in more recent periods, and which for example appears to be reflected in the faunal data from EBK hunting locales in Southern Scandinavia (see above).
Macro-patterns in beaver-related material culture
Beaver-related material culture has been recovered from across the Northern European wetland and boreal zones in the Early and Mid-Holocene (cf. Figure 1; Supplementary Information 1, available online). This material culture can be grouped into four primary groups: (i) incisor tools, (ii) mandible tools, (iii) tooth pendants, and (iv) other modified and unmodified but selected beaver bones not directly tied to food-getting or tooling endeavours, such as ankles. Incisor tools are made from the front teeth of beavers without their associated bone sockets, either by manipulating the teeth themselves or by simply using and/or subsequently re-sharping them. Mandible tools consist of completely or partially removed beaver mandibles including the front teeth, often modified and roughly shaped to facilitate effective instrumentalization. Tooth pendants are defined as beaver teeth that are either grooved or perforated for suspension.
Incisor and mandible tools
Incisor tools are found across the entire region, from the Early Mesolithic to the Late Mesolithic/Early Neolithic, whereas mandible tools are mainly known from the Baltic and the Russian West, where they are abundantly found at Mesolithic sites of all ages (Lozovskaya and Lozovski, 2015; Lozovskaya et al., 2017; Zhilin, 2001, 2020). For Russian contexts alone, Zhilin (2020) reports more than 1400 beaver mandible and incisor tools dated to the Mesolithic, mostly from the key wetland sites. Most of these objects represent prepared and/or used mandibles (n = 1388) and only a handful conform to modified frontal teeth (n = 34). The number of such objects varies greatly among archaeological sites, ranging from sites with only a small amount of such tools to archaeological sites such as Ozerki 5/IV and Veretje 1 bearing more than 100 objects. Mandible tools are least frequent in the Early Mesolithic and increase from the Middle to the Late Mesolithic/Early Neolithic (Zhilin, 2020). Incisor tools are currently unknown from the Early Mesolithic and most of these artefacts derive from Middle Mesolithic contexts within the region.
In Northwestern Europe, beaver-sourced tools from the Mesolithic are much less abundant but they have been reported in the literature (Figure 4 and Table 2). A single worked mandible has for example been described from the Early Mesolithic occupations of Star Carr in Britain (Knight et al., 2018), attributed to the Maglemose complex. Even though Star Carr lies outside of the study area, it showcases the early origin of beaver-related material culture also in northwest. In Southern Scandinavia and Northern Germany, mandible tools are presently only known from Maglemose contexts, including some of the classic Maglemose localities from Zealand in Eastern Denmark (Broholm, 1924; Hatting, 1970; Lautsen Lomborg, 2021). They mainly date to the Boreal/earliest Atlantic (‘Middle Mesolithic’ in the here-adopted terminology) and thus likely belong to the later part of the Early Holocene, even though this material should be radiocarbon dated to confirm this tentative placement. Beaver incisor tools, even though mostly represented by isolated pieces, were found at the Middle Mesolithic sites of Holmegård, Ørgård and Sværdborg in Denmark (Hatting, 1970; Lautsen Lomborg, 2021) and at Hohen Viecheln I (Schmölcke et al., 2017; Schuldt, 1961) and Rothenklempenow 17 (Schacht and Bogen, 2001) in Northern Germany. Beaver-sourced tools, both mandibles and incisors, were also found in a likely EBK context at the site of Heidemoor in the German Northeast (Ewersen, 2011), but this attribution similarly awaits future corroboration and is currently an isolated case.

Selected beaver-sourced tools from the Mesolithic of the Northwestern European wetland zone. 1–4: beaver incisor tools; 6–7: beaver mandible tools. 1: Hohen Viecheln (Germany), Middle Mesolithic (Schmölcke et al., 2017: Figure 6; photograph: H. Lübke, ZSBA Schleswig); 2: Hardinxveld-Giessendam Polderweg (The Netherlands), Late Mesolithic (Coles and Kooijmans, 2001: Figure 2); 3: Holmegård (Denmark), Middle Mesolithic (Hatting, 1970: Figure 10); 4: Øgårde (Denmark), Middle Mesolithic (Hatting, 1970: Figure 9b); 5: Spjellerup (Denmark), Middle Mesolithic? (Hatting, 1970: Figure 4); 6: Lynby (Denmark), Middle Mesolithic (Hatting, 1970: Figure 8b).
Overview of published beaver-related material culture from the Mesolithic and earliest Neolithic of continental Northwestern Europe.
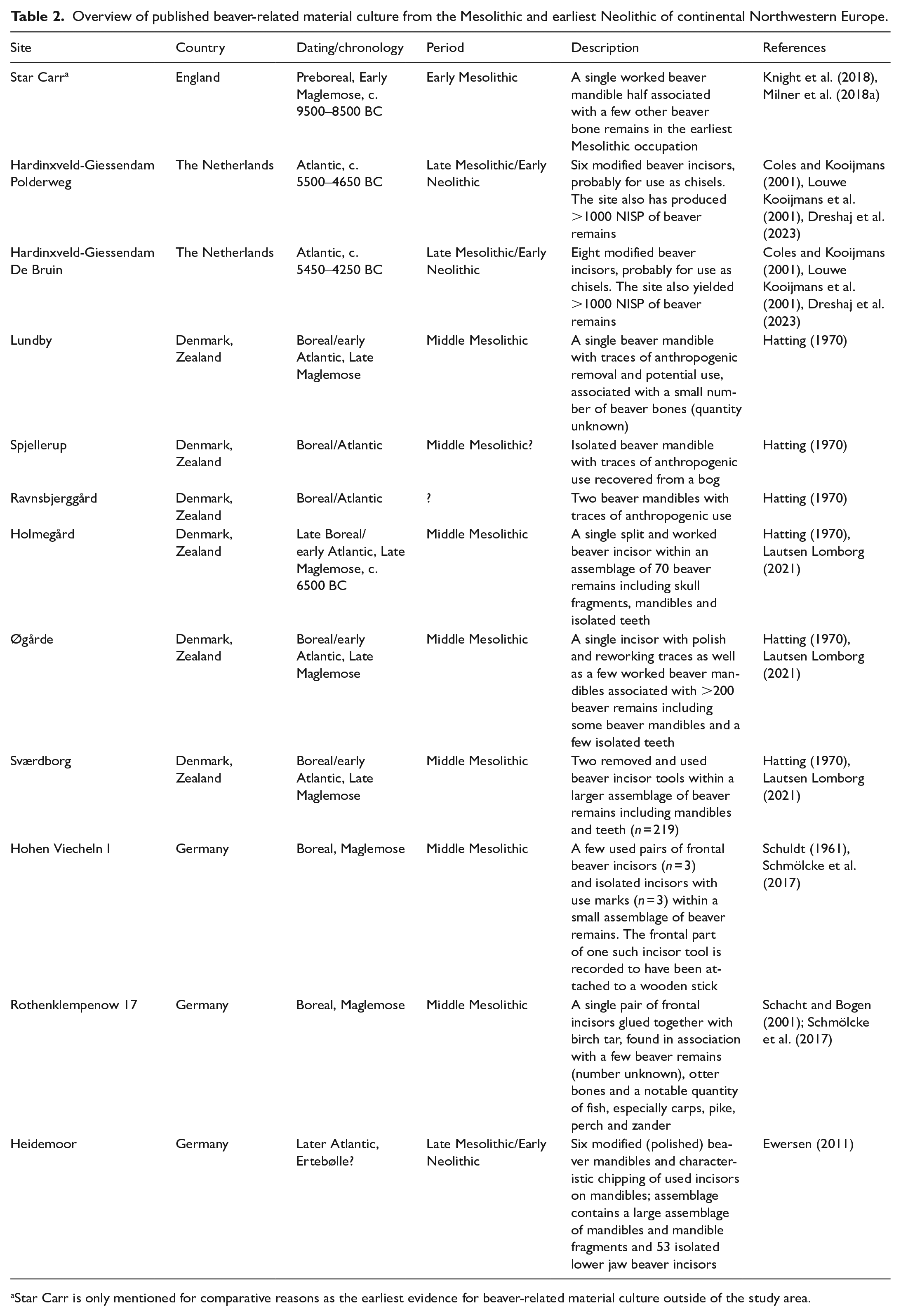
Star Carr is only mentioned for comparative reasons as the earliest evidence for beaver-related material culture outside of the study area.
No mandible tools have been found in the Netherlands. Only incisor tools are known and they derive from the two Late Mesolithic Hardinxveld-Giessendam sites (Coles and Kooijmans, 2001; Louwe Kooijmans et al., 2001, 2001). These tools bear a remarkable similarity to the incisor tools recovered from the Danish Middle Mesolithic sites. Esser et al., in preparation) further draw attention to the circumstance that at Tiel-Medel, while beaver mandibles have been recovered, only a very small number of beaver teeth occur in the assemblage, which is taphonomically unlikely, and may thus point to anthropogenic selection and filtering. It is for example possible that the missing beaver teeth have been removed for use or ornamentation elsewhere or were simply exported from the site but this hypothesis requires future empirical substantiation. That said, an analogous case is possibly framed by the Early Mesolithic site of Huseby Klev on the west coast of Sweden, where only beaver teeth and a single nearly complete mandible were found among the faunal remains and strontium isotope data points to a non-local origin of the material, contrasting with the rest of the zooarchaeological assemblage (Boethius et al., 2022).
The evidence from Northwestern Europe is extremely sparse, despite researchers being keenly aware of such finds and thus on the lookout for them (e.g. Esser et al., in preparation; Enghoff, 2011), suggesting this is most likely not due to researcher bias. The Northwestern earlier Mesolithic record thus appears to be structurally different from what is observed in the Northeast. Most relevant beaver-related material culture from the region either dates to the Middle Mesolithic or is associated with the earlier part of the Mid-Holocene, even though the precise dating of some of the objects remains problematic. It is worth noting that for the EBK specifically, tools made from animal bones are rare overall, so this difference may have less to do with the role of beavers in particular and more with the status of animal-related material culture in general. In Swifterbant Culture contexts, by contrast, a wide diversity of species appears to have been involved in tool production – from wild boar to swans to caprines (Aal and van Gent in preparation; Kranenburg and Prummel, 2020; Louwe Kooijmans et al., 2001) – suggesting little species-level discrimination. More detailed analysis, for example with regard to body part selectivity (see e.g. Hill, 2019 and Hussain et al., 2022 for the key importance of the latter), may be warranted, however.
The function of beaver-sourced tools has been discussed extensively in the literature (e.g. Coles, 2006; Lozovskaya et al., 2017; Schmölcke et al., 2017; Zhilin, 1997, 2020). At Middle Mesolithic Veretje 1, the first ever securely identified beaver mandible tool was reportedly still bound with a strip of bark, revealing its tool-character and the way it was instrumentalized (Oshibkina, 1983; Zhilin, 1997). Similar mandible tools attached to wooden handles are for example known from Indigenous people from Alaska who used them as scrapers, the molars functioning like a rasp (Osgood, 1940; Schmölcke et al., 2017). Following Zhilin (2014a, 2014b, 2020), Mesolithic instances of such beaver-sourced tools were mainly used as scrapers, knives, and chisels or as pressure flaking devices. Woodworking was an important task of many of these tools but some scraper-like mandible tools were apparently also used for bone-working (Zhilin, 2020). The utilization of these objects therefore largely mirrors the capacity of the respective body-parts in a living beaver, which has led Schmölcke et al. (2017: 8) to suggest that ‘perhaps by observing these animals prehistoric people got the notion that they have built-in woodworking tools’. We return to this point below and take it up again in the discussion.
The in-depth techno-functional analyses of these objects performed by Zhilin (2020) have further shown that beaver-sourced tools were not deployed ad hoc, as might perhaps hastily be inferred from a pragmatic copy-paste logic vis-à-vis beaver woodcutting practices. Instead, the chaîne opératoire of these objects is often surprisingly complex and demonstrates distinct stages of reworking (Zhilin, 2020), suggesting not only that many of these tools were probably in use for quite some time, but also that they were actively curated and thus generally cared for. We must assume that Mesolithic people generally had the capacity and means to acquire beaver bodies to replenish tool stocks if they had wished to, and the extended life-histories of beaver-sourced tools thus strongly suggest that the objects mattered to people, and keeping the same items in human systems for prolonged periods of time was a conscious concern. Furthermore, using and handling these tools would have brought beaver bodies to the centre of human ‘horizons of concern’ (sensu Bird-David, 2017), promoting understanding of and sympathy for the beaver, and thus bringing human and beaver perspectives closer together, with human and beaver bodies as well as perceptive and cognitive horizons literally merging, if only temporally.
Tooth pendants and other invested beaver objects
Beaver tooth pendants are relatively rare and currently confined to the eastern Boreal zone, where some examples have been reported from the Middle Mesolithic site of Ozerki 17 (Zhilin, 1996: 218), Late Mesolithic Okajomovo 5 and Nushpoli 11 at the Dubna River (Zhilin, 2007), Late Mesolithic Kubenino at the Onega River (Kashina et al., 2021), and >1200 beaver teeth in total were found in human burial contexts within the extensive Late Mesolithic/Early Neolithic hunter-gatherer cemetery of Oleniy Ostrov in what is today Karelia (Grünberg, 2013; Mannermaa and Rainio, 2020), making up about 20% of all animal tooth pendants originally published by Gurina (1956) for the site. Some of these were cut into plates, show macroscopic use-wear traces and bear notches and/or grooves, suggesting that they were worn extensively (Grünberg, 2013: 235; Mannermaa et al., 2019), perhaps by more than a single person. Beaver tooth pendants seem to be rare at Early Mesolithic sites but Zhilin (2014a) has recently reported several such objects from the Preboreal site of Ivanovskoye 7/IV (cf. Schmölcke et al., 2017: 5). Apart from the importance of the species origin for understanding these objects, Grünberg (2000) has suggested that beaver incisors were probably significant because of their unique orange-brown colouring. In addition, some of the beaver pendants were probably made or rather procured from former mandible or incisor tools (Zhilin, 2001, 2020: 10) and thus represent the final life-history stage of beaver-sourced material culture, so that placing these objects into human burials may be significant itself. In the Baltic, a small number of beaver tooth pendants were recovered from Early Mesolithic Pulli and the Early Neolithic site of Kudruküla in Estonia (Jonkus and Rannamäe, 2018), and there are likely more examples from this region that either await publication or escaped our literature survey (see esp. Macāne 2022).
Beaver astragali (ankle bones), perforated or not, have been found in greater numbers in human interments at the important Early-to-Late Mesolithic/Early Neolithic burial ground of Zvejnieki in Northern Latvia (Eriksson et al., 2003; Macāne 2022), where most of the animal bone pendants associated with the more than 100 buried human individuals represent beaver astragali (Zagorskis, 1987; cf. Grünberg, 2013: 237). The number of bone pendants at Zvejnieki generally increases from the Early to the Late Mesolithic/Early Neolithic. Interestingly, beaver tooth pendants seem to be associated mainly with female-sexed human bodies in Oleniy Ostrov (Gurina 1956; Fehner, 1963; O’Shea and Zvelebil, 1984: 10), while beaver astragali are predominantly tangled up with children or juvenile and adult males in Zvejnieki (Macāne 2022: 235), perhaps pointing to the differential involvement of beaver body parts in the construction of social persona in hunter-gatherer societies across Northeastern Europe. In Zvejnieki, this may be further supported by the circumstance that beaver astragali tend to be found in more extensively furnished human burials, are mainly linked to later burial activities (c. 6th millennium cal. BC) and appear to be spatially separated from burials containing beaver teeth (Macāne 2022). Some Mesolithic burials in Northeastern Europe, for example at Oleniy Ostrov, are also associated with other unmodified beaver bones such as ulnae (Grünberg, 2013), but it is presently difficult to establish how important, selective or wide-spread specific beaver bone-burial patterns and their linked cultural practices were.
We can thus generally observe a gradual diversification of beaver-related material culture and practices of materialization in the course of the Northeastern Mesolithic (Mannermaa et al., 2019). In this context, it is also notable that Zagorska et al. (2018) have recently re-interpreted a fragmented zoomorphic antler staff head from Late Mesolithic/Early Neolithic Zvejnieki as rendering either beaver or otter, linking the object to the distinct tradition of zoomorphic figurine-making which spans the Baltic and parts of Western Russia in the well-defined Middle Neolithic of this area at the end of the Mid-Holocene. This tradition features exceptional beaver and/or otter-like figurines, either plastically carved or rendered in bird’s-eye profile, for example from the settlement of Valma at Lake Võrtsjärv in Estonia (Jaanits, 1965), where an amber figurine from a female human burial is now also interpreted as a beaver (Ots, 2010).
In contrast, no ornaments and/or grave goods made from beaver remains have been documented in Northwestern Europe. This is despite the fact that pendants made from animal teeth are relatively common at Swifterbant Culture sites for example, both in settlement and burial contexts, including perforated teeth of cattle (Bos sp.), wild boar, pig (Sus domesticus), horse (Equus ferus), dog (Canis familiaris), and otter (Devriendt, 2008; Kranenburg and Prummel, 2020). These differences between eastern and western Northern Europe indicate divergent histories of human-beaver interaction and cohabitation, and may point to important differences in how beaver relations were negotiated, where the animals were assigned to in multispecies systems and, possibly, how significant they were for human livelihoods.
Two trajectories of beaver-related material culture
Two different trajectories of beaver-related material culture development can therefore be discerned. In the Northwest of continental Europe, beaver-related material culture appears to be mainly confined to the instrumentalization of beaver body parts as tools, documented in different areas at different time intervals. In the Baltic and in Western Russia, by contrast, beaver-related tool-making practices are joined by evidence for the use of beaver teeth as pendants and the special treatment of other beaver body parts, especially astragali. These large scale inter-regional differences are interesting, as they suggest, together with the faunal data reported above, that human-beaver exposition and cohabitation was more stable and consequential in the Northeast, developing its own distinct historical dynamic reflected in material culture diversification over time, while the significance of beavers in the Northwest may have been more situational and context-specific.
Ichtyofaunal patterns
Fish remains from the Northern Mesolithic provide important information on human landscape use and exploited aquatic ecologies, and may further disclose hitherto overlooked evidence for human-beaver cohabitation and encouragement in wetland and freshwater environments. Similar to the data on mammalian faunal compositions, the available evidence on human-procured fish during the Mesolithic indicates a broad distinction between ichtyofaunal assemblages in Northwestern Europe on the one hand and the Baltic and the Russian West on the other (Figures 5 and 6; Supplementary Information F, available online).
Number of identified fish specimens (NISP%) from the Baltic countries and Western Russia from the Early to the first part of the Mid-Holocene: (a) Early Mesolithic; (b) Middle Mesolithic; (c) Late Mesolithic/Early Neolithic. Horizontal bars represent median values with standard deviations. Animal silhouettes have been redrawn from https://www.phylopic.org/. Raw data are provided in Supplementary Information F Tables 1 and 2, available online.
Number of identified fish specimens (NISP%) from Southern Scandinavia and the Dutch wetlands from the Early to the first part of the Mid-Holocene: (a) Early Mesolithic; (b) Middle Mesolithic; (c) Late Mesolithic/Early Neolithic. Note that data for the Dutch wetlands is only available for the final phase. Horizontal bars represent median values with standard deviations. Animal silhouettes have been redrawn from https://www.phylopic.org/. Raw data are provided in Supplementary Information F Tables 3 and 4, available online.
Northeastern Europe
In Western Russia, northern pike (Esox lucius) dominates the ichtyofauna from the Early to the Late Mesolithic (mean NISP = c. 50–70%) but the relative importance of pike decreases slightly over time, while other large freshwater predators such as the European perch (Perca fluviatalis) become more frequent throughout the Middle and Late Mesolithic (Figure 5). Early Mesolithic assemblages tend to be more monospecific, focusing either on the pike or, in one case, on the predatory zander/pike-perch (Sander lucioperca: NISP = 95%). From the Middle Mesolithic onwards, the ichtyofauna not only becomes more diverse, it is also notably enriched in carps and ground-feeders in general. Wels catfish (Silurus glanis), occasionally found already in the Early Mesolithic, emerges as a regular although low-frequency component of Late Mesolithic/Early Neolithic assemblages. Importantly, the vast majority of represented species forms part of lentic communities (Figure 7) and some of the notable later Early Holocene and Mid-Holocene species suggest that eutrophic conditions must have been well-established in the landscape by then. The eel (Anguilla sp.) remains exceptionally rare even in the Late Mesolithic, when the Littorina Sea was already well developed (Kostecki, 2014) and eel has been shown to be generally present in Baltic waters (Enghoff and Ediger, 2016).
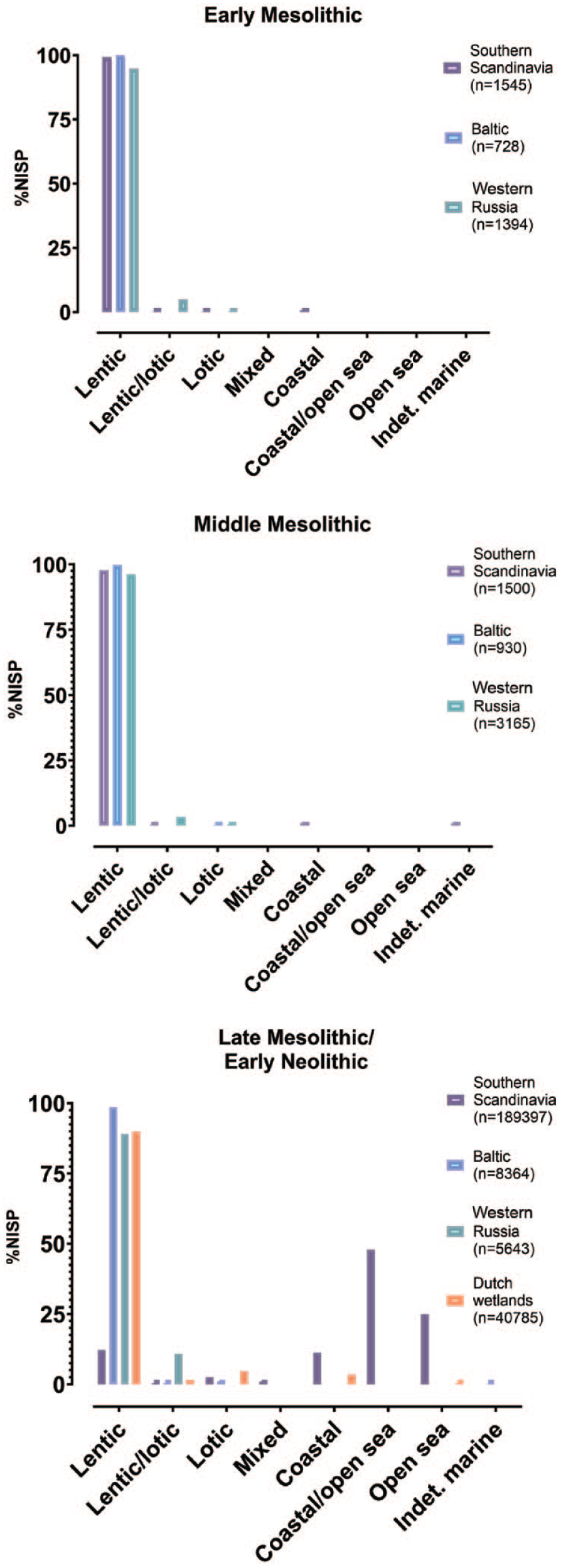
Marine versus freshwater fish ecologies within ichtyofaunal assemblages as represented by NISP shares in different regions and periods. Species classification and summary data table are provided in Supplementary Information 2, available online.
In the Baltic, the evidence is sparser and some patterns are repeated. In the Early and Middle Mesolithic, the ichtyofauna is dominated by both northern pike and zander/pike-perch, which together make up at least >60% and often >90% of recorded fish NISP. Yet in contrast to Western Russia, northern pike becomes more frequent in the Late Mesolithic/Early Neolithic, often at the expense of zander, and the Middle and Late Mesolithic are again characterized by a notable catfish input (up to NISP = 28% in the Late Mesolithic). Interestingly, Wels catfish seems to supplant carps in the course of the Mesolithic, but the dataset is not robust enough to securely establish this trend. Kõpu I in Estonia presents a rare example of notable marine fish input during the Late Mesolithic/Early Neolithic, but the fish assemblage is too small (NISP = 6) to extrapolate from this so far isolated context. Given that the site’s mammalian fauna is dominated by seal (Lõugas et al., 2017), the marine fish component is likely the result of specific foraging activities linked to targeted phocid hunting at the emerging Baltic Sea, and is thus not necessarily reflective of a systematic practice and/or well-established marine fishing economies (see esp. Boethius et al., 2017 for similarly cautionary arguments regarding another Baltic Early Holocene context from the Swedish island of Gotland).
Northwestern Europe
In Southern Scandinavia, the Early Holocene fish record strongly differs from its Mid-Holocene counterpart. Early and Middle Mesolithic fish assemblages from Northern Germany, Sweden, and Denmark are strongly dominated by northern pike (mean NISP ⩾ 80%), while carps are also common, especially in the Middle Mesolithic, and some pike-heavy assemblages feature a few catfish remains (Figure 6). These Early Holocene assemblages attributed to the Maglemose complex are exclusively freshwater-oriented. Bølling Sø in Jutland is the only site featuring marine fish, the catadromous (migrating down-river) European eel (Robson and Ritchie, 2019). Even though Mesolithic people in the area had undoubtedly access to the sea, they thus mostly exploited freshwater habitats and overwhelmingly focused on larger lentic fish, in particular predators and cyprinids, even though other species were clearly available. Future investigations may complement this picture, however, as most former Early Holocene coastal sites are submerged today (Astrup, 2018, 2020) and some early Mesolithic sites with evidence for marine exploitation exist, for example in Western Sweden (Boethius 2018). This being said, a similar emphasis on pike, perch (Perca fluviatilis), and carps has recently been established for the Early Mesolithic site complex at Lake Flixton in Northern England including the important occupations at Star Carr (Robson et al., 2018).
This picture changes in the Mid-Holocene and the ensuing EBK complex, which mostly spans the later Atlantic (cf. Enghoff, 1994; Enghoff et al., 2007). The focus is then shifted towards eel, cods (Gadidae), and flounder (Pleuronectidae), and thus to deep sea and/or transitional fish species (Ritchie, 2010). Whereas eels (NISP = 1–73%), cods (NISP = 2–74%) and flounders (NISP = 1–58%) compete for the top-ranking spot in the early Mid-Holocene fish record in Jutland, cods (NISP = 44–86%) dominate the record in Zealand, followed by flounders (NISP = 2–71%) and eels (NISP = 1–9%), who are much less important in this part of Mesolithic Denmark (Ritchie, 2010). Carps remain significant in some Danish Mid-Holocene contexts, notably on the Jutland peninsula, but overall there is a transition to marine fisheries. This change in human subsistence is broadly concomitant with emerging sea mammal and dedicated ocean-bound economies developing during the Atlantic chronozone (Price, 1985; Price et al., 2018; Rowley-Conwy, 1999).
The Mid-Holocene fish record from the Dutch wetlands is, like the Northeastern Mesolithic, also governed by lentic fish communities, albeit with more lotic and marine fish representation (cf. Figures 6 and 7). Together, northern pike and carps dominate the Late Mesolithic/Early Neolithic assemblages and it has been suggested that the migratory patterns of pike may have been critical in establishing the seasonal mobility of the communities using the Hardinxveld-Giessendam environments (Beerenhout, 2001a, 2001b). Pike becomes less frequent through time and carps show the reverse trend. At the Early Neolithic site Hoge Vaart A27, cyprinids contribute more than 69% of recorded fish NISP (Laarman, 2001). Pike is overall less abundant than in the Northeastern Mesolithic and there is a general shift from an initial focus on large predatory freshwater fish such as pike and European perch to an emphasis on carps and catfish, which can reach up to 13% of the ichtyofaunal NISP in the Early Neolithic (Kranenburg and Prummel, 2020). Salmonids are notably very rare. Interestingly, eel only becomes a factor at around 4000 cal. BC (c. 3–14% of NISP) and this parallels a notable increase in lentic fish, many of which also thrive in freshwater environments with reduced or locally disrupted riverine connectivity. The occasional occurrence of a small component of sturgeons (Acipenseridae) is also notable, a coastal species which is anadromous (migrates river-up). The marine fish component is otherwise negligible, even though people must have had direct access to the coast, so this is unlikely a question of availability alone. The Late Mesolithic/Early Neolithic evidence from the North European wetland zone thus clearly bespeaks of a unique and highly complex aquatic ecology at the mouth of the young Holocene River Rhine and the Swifterbant river system with different coexisting flow regimes. This complexity of riverine habitats within a vast, braided river network supporting varied levels of hydrological connectivity and eutrophication is at least in part the co-product of the activities of beaver populations in the region, who were probably present in the wider area from at least the Early Holocene onwards.
Discussion
The onset of the Holocene in Northern Europe may be framed as a ‘beaver event’ setting the scene for post-glacial human history above 50° latitude north. Beavers were among the pioneer species moving into the newly available high-latitude environments released by the retreating glaciers at the end of the Pleistocene. These novel environments with their rich and dynamic hydrologies, including many glacial legacy lakes, were not only suitable for beavers to establish themselves as prominent keystone agents in the north, these environments were also rapidly transformed by beaver geopraxis, promoting wetlands, riverine heterogeneity, distinct lake and pond-invested ecosystems, as well as unique patterns of plant and animal biodiversity across landscape scales. As Tape et al. (2018) have shown, beaver colonization of Arctic tundra landscapes can accelerate the thawing of permafrost and may thus quicken the transformation of periglacial into post-glacial ecosystems. At the onset of the Holocene in Northern Europe, beavers were not just widely present and recognizably important agents in the landscape (Liarsou, 2020) – documented well-before human foragers left any tangible traces in the archaeological record – they were probably co-responsible for shaping the kinds of environments that we today readily identify as earlier Holocene, from the extensive riverine wetlands of Northwestern Europe to the mosaic of lake-rich boreal and tundra environments in the Baltic and in Western Russia (Wohl, 2021). There is indeed much untapped potential here to conduct landscape-scale geoarchaeological research to better tease apart climatic, human, and beaver impacts contributing to the unique environmental profile of Holocene landscapes across the European north.
The combined archaeological and zooarchaeological evidence from the Northern Mesolithic reviewed and synthesized here (c. 9000–4000 cal. BC) opens up the interesting possibility that transformative beaver ecosystem agency provided an important framework and generative context for human foraging lifeways and ecocultural identities developing at the edge of former glaciers. The persistence of beaver remains through large parts of the Mesolithic across the study regions suggests that human-beaver intersections were likely more consequential and materially generative than previously recognized, and long-term developments in material culture and society appear to so have been critically interlaced with the changing dynamics of human-beaver systems. When deploying a dedicated ‘beaver lens’, the archaeological record and the recoverable spatiotemporal patterns in mammal remains, fish fauna and beaver-related material culture in the Mesolithic of Northern Europe drawn together here thus motivate re-evaluation and re-contextualization of beaver agency, ecological engineering and changing human historical registers.
As a whole, the available archaeological evidence points to notable differences in the long-term development of human-beaver systems between the Northwest and Northeast of Europe but also within the Northwestern study region. In the boreal and taiga zone of the Baltic and Western Russia, beaver remains form a substantial and stable component of the species composition throughout the Mesolithic. Beaver-related material culture is abundant and represents a significant and diversified corpus of beaver-related materialization, with beavers becoming increasingly important in ornamental and burial practices through time. In contrast, only a few beaver-sourced tools and no pendants are known from Southern Scandinavia. Here, beaver remains are far fewer than in the Northeast during the Early and the Middle Mesolithic, but they are a steady component of Maglemose and Kongemose assemblages. This changes significantly in the Late Mesolithic, when beavers proportionally all but disappear from the zooarchaeological record. In the Dutch Late Mesolithic and Early Neolithic, in opposition, beaver remains are abundant, rivalling Northeastern European beaver percentages. While there is notable variation between archaeological sites, beaver appears to have been an important component of Swifterbant Culture subsistence and remains so at wetland sites until well into the Dutch Neolithic (cf. Lauwerier et al., 2005). Yet only a few incisor tools have been found so far and none of the many animal tooth pendants are beaver-sourced. The lack of sites from the Early and Middle Mesolithic from the Dutch area makes it currently impossible to assess developments through time, however.
We suggest that the divergence of regional trajectories between and within Northwestern and Northeastern Europe is bound to the varying nature of human-beaver relationships, pertaining to modes of cohabitation and the shifting significance of beavers for human affairs. Querying these differences in human-beaver expositions in the Mesolithic of Northern Europe arguably requires unsettling current orthodoxy and methodologically inverting the direction of inquiry – to ask what beavers could do for human societies and how the animals would have framed human affairs, thus investigating possible life-services for human societies provided by beavers and their geopraxes. It is here that the other mammalian and ichtyofaunal evidence from the Mesolithic is of particular relevance (cf. Liarsou, 2013: 174, Liarsou, 2015, 2020: 38–42), authorizing renewed conversations on the supportive role of beavers for earlier Holocene human occupations of the northern wetland and boreal zones of continental Europe.
Mammal hunting affordances
A ‘beaver lens’ reveals interesting patterns in species co-associations and possible long-term dynamics in multispecies assemblages related to beaver agency. In Northeastern Europe, for example, the predominance of elk alongside beaver is noteworthy as beaver-elk ecosystem associations are reflective of incipient colonization scenarios where beaver-powered wetlands create a mosaic of standing water bodies and promote the growth of riparian or edge vegetation such as willow (Gibson and Olden, 2014), in turn attracting elk (Baker et al., 2005; Nummi et al., 2019; Ripple and Beschta, 2004). Beavers and elk also display a high degree of dietary overlap (Hoy et al., 2019). As previously argued by Petersen (2009: 46), beaver-elk interactions are often beneficial for human foragers and the beaver may have accordingly come into view as a ‘faunal regenerator’ because of this notable ecological interrelationship. Similar ecosystem dynamics are harder to trace in Northwestern Europe, in part because elk is much less frequent in faunal assemblages there. In Southern Scandinavia, for instance, elk altogether disappears from the zooarchaeological record in the course of the Mesolithic, even though, puzzlingly, the species remains culturally important (Bridault, 1992) – but these processes may in fact be interrelated. In the Dutch wetlands, elk is consistently present only in small numbers, despite wetlands being among the preferred habitats of the species (cf. Janík et al., 2021), opening up the possibility that Swifterbant Culture communities culturally opted not to hunt elk systematically. This scenario is not in principle inconceivable for the later Mesolithic of Southern Scandinavia as well.
A recent ecological study conducted in Denmark has shown that red deer and roe deer avoid water in areas with high beaver activity, and that these cervids tend to occur in higher numbers in areas with fewer beavers (Svanholm Pejstrup et al., 2023). This is noteworthy because the Dutch Mesolithic and Early Neolithic record contains few Cervidae and at the same time features high numbers of beavers – precisely the predicated pattern. In Northeastern Europe, the pattern is overall the same, and the few sites with abundant red deer (Zvidze and Zemaitiskes) yielded few beaver remains. In Southern Scandinavia, by contrast, red deer and roe deer are very common and, as noted earlier, beaver much less so than in the other regions. Svanholm Pejstrup et al. (2023) caution that human impact likely plays a formative role in these present-day observations on interspecies dynamics, so it remains an open question as to what extent such dynamics can be extrapolated back in time, but they are in any case worth considering and should further be explored in and tested against the archaeological record. It should at least be considered that beaver geopraxis may have been less prominent and consequential in Southern Scandinavia when compared to the Dutch wetlands and Northeastern Europe as a whole.
The faunal record of Northwestern Europe hosts a diversity of megafauna, including Cervidae, horse, wild boar and aurochs. Cervidae and horse indicate a more open wetland zone interspersed with deciduous and mixed woodland already very early in the Holocene. Over the course of the Mesolithic, these animals would have thus played a role in further opening up the dense alluvial forests through extensive grazing (e.g. Noe-Nygaard et al., 2005). In the Baltic, the documented increase of wild boar, red deer, and eventually aurochs may thereby have been encouraged by well-established openings at the edge of mature beaver habitats and the role of beaver hotspots as landscape attractors for these animals (Fedyń et al., 2022) – probably fuelled by early low-level human woodland impact and perhaps management (cf. Poska et al., 2004). This may point to an easily overlooked interplay – with notable feedback potential – between beaver woodland engineering and clearance on the one hand and ungulate suppression of woodland regeneration on the other hand – a dynamic possibly consequential for Northwestern Europe and its specific Late Mesolithic/Early Neolithic human histories.
Unlike deer, wild boar are strongly attracted to beaver-engineered landscapes and in particular aged beaver ponds provide ideal foraging and wallowing locales for them (Nitsche, 1997; Rosell et al., 2005). Their co-occurrence with beaver remains in the zooarchaeological record in all of the study regions is therefore interesting and probably influenced by such interspecies co-facilitation, in turn providing human foragers with the possibility to intercept and target boar in beaver country. As discussed earlier, wild boar forms an important element of subsistence across Mesolithic Northern Europe, for meat – and for teeth, tools and pendants. It is important to note that that wild boar appears to acquire a new role in human economies in the Late Mesolithic and Early Neolithic at least in some areas, probably due to local anthropogenic encouragement and/or incipient management strategies (Brusgaard et al., forthcoming; Magnell, 2006; Maring and Riede, 2019; Rosvold et al., 2010). This latter process may not be unrelated to the likely facilitation of wild boar in beaver landscapes, as humans foraging in these landscapes would also intersect and increasingly interact with wild boar, perhaps laying the foundation for subsequent management, taming, and domestication.
Some smaller mammals in the dataset may similarly signal beaver-related ecological promotion. In Western Russia, the Early Mesolithic is characterized by the importance of muskrat and marten, animals who have been argued to form an early colonizer assemblage within formative beaver landscapes (Crego et al., 2016; Nummi et al., 2019). The case of the muskrat may be especially instructive as this semi-aquatic rodent is known to parasitize beaver lodges (Mott et al., 2013), so that knowledge on beaver landscapes can promote muskrat foraging and generally supports integrative foraging strategies in Boreal ecologies (cf. Winterhalder, 1981; see below). The relationship between the beaver and the water vole in this region, as noted earlier, is of further interest in this context because of the common ecological association, and succession, of the two species, which is also an important concern of present-day ecological restoration projects (cf. Stringer and Gaywood, 2016). This latter interspecies relationship is perhaps also reflected in geoarchaeological findings from Grabow 15 in Northern Germany, where an early Atlantic beaver burrow system was documented in conjunction with a later, possibly Late-Holocene, water vole-gnawed piece of wood (Tolksdorf et al., 2017), supporting the idea that water vole is an indicator species of developed-matured beaver wetlands.
Otter is consistently present in Mesolithic assemblages in which beavers are also prevalent, probably because of the earlier outlined facilitation of otters through beaver geopraxis (LeBlanc et al., 2007; Reid, 1984; Tumlison et al., 1982). In the Netherlands, the exploitation of both beaver and otter has been shown to be a key feature of Swifterbant Culture sites, as well as Middle Neolithic sites attributed to the so-called Vlaardingen Culture (c. 3500–2600 BC) (Brinkkemper et al., 2011; Lauwerier et al., 2005; Zeiler, 1987). This pattern supports the observations from Northeastern Europe, where these two species tend to converge in the faunal record, and accordingly lends credence to the idea that integrated low-level beaver and otter foraging was often opportune and possibly sustainable. In Southern Scandinavia, marten, mink (Mustela sp.) and otter are also a recurrent but low-abundance faunal component in the Early and Middle Mesolithic, which may similarly be indicative of exploited foraging affordances within beaver habitats. It is not clear, however, whether the decrease in beaver remains documented in the Late Mesolithic of the region denotes a principal change in hunting practices and logics with regard to beavers, or instead signals a switch to other animals occurring in beaver landscapes at the expense of beavers, or both. For example, some sites feature hardly any beaver remains but have yielded abundant other fur-bearing animals like pine marten (Martes martes) and wild cat, who were probably targeted separately within specialized/dedicated fur-getting economies (Price, 1991; Richter, 2005; Richter and Noe-Nygaard, 2003; Trolle-Lassen, 1987). Otter in the EBK was likely targeted in the context of coastal and marine activities (e.g. Price et al., 2001), signalling a reorganization of foraging affordances and perhaps even their partial disintegration, and this may indeed indicate subtle but relevant changes in the role of beaver affordances in regulating larger foraging systems.
Taken together, these successional and interspecies faunal dynamics may point to the crucial significance of integrated foraging systems in the Mesolithic of Northern Europe, centred on animal resources directly accessible at hotspots of beaver activity or at sites of former beaver occupation (legacy sites). In Southern Scandinavia, this constellation may have fundamentally been reconfigured towards the end of the Mesolithic, while demonstrating substantive continuity in the other North European study regions. This human-beaver entanglement would have granted beaver landscapes special significance for human foragers and it is tempting to conceptualize such ‘beaver country’ in analogy to notions of Country perpetuated by Indigenous groups across Australia, emphasizing reciprocity and nourishment through human care (e.g. Urwin et al., 2022). Either way, these dynamics of human-beaver cohabitation would have drawn the beaver closer to the centre of forager lifeworlds and ‘horizons of concern’ (sensu Bird-David, 2017), fostering people’s attentiveness to the animals and their resource provisioning work as well as the attendant multispecies rhythms. Beavers, then, hold a double status as ecological and phenomenological cornerstones of Northern Mesolithic more-than-human landscape ‘dwelling’ (sensu Ingold, 2022). In this optic, the beaver, furthermore, quickly acquires the status of a collaborator – a feral partner – in the pursuit of human sustenance. The ecological keystone status of beavers in Northern Europe – their capacity to anchor and assemble animal ecologies and ecosystem processes – therefore arguably laid the foundations for the ecological facilitation of Mesolithic human foragers living in the same landscapes, who in turn integrated a large suite of beaver landscape affordances into their behavioural repertoire. As Kikvidze and Callaway (2009) point out, facilitation can be a powerful but often overlooked evolutionary factor, structuring multispecies communities and shaping long-term dynamics of history.
Fish-getting affordances
The possibility of beaver facilitation is equally supported by the Mesolithic fish record. The striking feature of the ichtyofaunal record from the earlier part of Holocene Northern Europe is its strong emphasis on larger predatory freshwater fish such as pike, zander and perch in conjunction with a growing importance of lentic bottom-dwellers over time, such as carps and Wels catfish. This accentuation is consistent with some known effects of beaver-fish interactions, while the observed regional variability similarly points to important differences in hydrological systems and aquatic ecologies across Northern Europe – especially the relative importance of larger lakes vis-à-vis riparian corridors – and the changing impacts of the beaver on these systems and flow regimes. Beaver activity generally fosters habitat heterogeneity over larger spatial scales by creating patches of lentic habitat within a corridor of lotic habitat, thus altering and framing new niches for both ‘stream species’ and ‘pond species’ (Collen and Gibson, 2000; Snodgrass and Meffe, 1999: 452). In warm water ecosystems, beaver ponding, eutrophication, and lentic shallow-water and edge-habitat engineering mainly promote potamodromous (i.e. freshwater-only) species such as pike and perch, who can over time outcompete and replace smaller bodied cyprinids (e.g. Gaywood, 2018; Pliūraitė and Kesminas, 2012; Rosell et al., 2005). Pike is found to benefit from beaver ecosystem modification especially when larger ponds or lakes are available (Collen and Gibson, 2000), while perches are attracted by beaver lodge debris (Gibson, 1969). As conditions within ponds become increasingly anaerobic, larger carps and species such as catfish increasingly benefit and typically grow in abundance. Wels catfish, once promoted, can further impact freshwater ecologies due to induced alterations of the trophic chain and physiochemical modifications of the water content (Rodriguez-Labajos et al., 2009), sometimes leading to the displacement and near-disappearance of other species such as the common carp (Cyprinus carpio) (Snodgrass and Meffe, 1998), whilst attendant algae growth may complicate even the spearing of larger fish such as pike.
In all study regions, the fish evidence often reveals a gradual re-configuration of lentic fish communities in the course of the Mesolithic – a successional sequence starting with an emphasis on pike and other large freshwater predators leading to increased attention to cyprinids and eventually catfish. This, then, is unlikely to be reflective only of human foraging preferences but probably also records a latent beaver legacy effect, as these changes are a consequence of consolidating and ageing beaver landscapes including maturing ponds, some of which are ultimately abandoned, offering distinct fishing opportunities. Fish-getting practices and human strategic decision-making during the Mesolithic were thus likely influenced, and dynamically modulated, by the specific fish-getting affordances emerging from long-term beaver activity in the hydroactive wetland and boreal environments of Northern Europe at the edge of former glaciers.
Beavers almost never fully disrupt riparian connectivity (Schlosser, 1995) but they can severely impede the capacity of river migrating species such as salmons and eels to traverse across riparian landscapes (cf. Kemp et al., 2012). Mitchell and Cunjak (2007) found that beaver dams in coastal rivers prevent upstream migration of salmonids and simultaneously, through competitive exclusion, increase fish diversity upstream. This dynamic may apply to, and in part explain, the ichtyofaunal patterns of the Late Mesolithic/Early Neolithic from the Dutch wetlands, where salmonids are conspicuously rare. This pattern indeed continues at Middle Neolithic wetland and coastal sites in the area, where seal and other marine mammals increase in importance, yet the ichtyofaunal emphasis remains on pike, sturgeon, eel and cyprinids (Lauwerier et al., 2005). This could suggest long-lived beaver legacies, hunting affordances, and the resulting enfolded cultural practices. A highly similar pattern emerges from the Middle Mesolithic of the Baltic, where beaver-supported lentic fish including pike, perch, and cyprinids remain the focal target of fish-getting practices, even though human foragers begin to engage in specialized seal hunting (Lõugas et al., 2017). The increasing importance of the sea does therefore, contrary to what might be assumed, not lead to a dramatic shift in ichtyofaunal acquisition patterns, and lotic fish continue to form only small portions of human-foraged fish. Consonant with this broader perspective, Boethius et al. (2017) have previously argued that freshwater fish derived from eutrophic lakes remained a key subsistence staple even in the context of the initial Early Holocene occupation of Gotland in the Baltic basin, while marine mammal hunting, especially the targeted pursuit of younger seals, was secondary and probably primarily oriented towards and motivated by raw material acquisition.
Salmonids and eel inputs to Mesolithic fish assemblages remain generally negligible, perhaps indicating some level of amensalism (cf. Arthur and Mitchell, 1989) between earlier Holocene beaver-powered environments and migratory, lotic fish requiring access to the open sea. This idea may be supported by the faunal evidence from Mesolithic Ireland, where the beaver is not part of the native mammalian fauna, while salmonids and eel are of key importance in the anthropogenic fish assemblages (Kelly, 2005; Warren, 2022), contrasting with the data from Mesolithic mainland Europe and Britain where these species are virtually absent (Robson and Ritchie, 2019; Robson et al., 2018; Zhilin, 2014a). This again suggests that beaver agency co-structured anthropogenic fish assemblages. Alternatively, or perhaps complementarily, lentic fish, especially larger predatory species and fast-growing carps, offered more attractive, more reliable and easier to access food resources due to their predictable association with beaver-fabricated ponds and wetlands.
In this context, a few comments on the evolution of fishing technologies in Northern Europe are useful. Sophisticated and curated fishing installations such as fish weirs, fences and/or proper fisheries only emerge in the course of the Mesolithic and typically date to the later part of the Middle Mesolithic or the Late Mesolithic/Early Neolithic in the study regions (e.g. Amkreutz, 2013; Fischer, 2007; Lozovski and Lozovskaya, 2016). In the Early Holocene, fish was thus probably often acquired via spearing and/or bowing, sometimes but not always in conjunction with angling, and these practices can easily take advantage of fish trapped in ponds, and the shallow water habitats engineered by beavers can greatly facilitate the spotting, spearing and/or bowing of larger freshwater fish such as pike. Eutrophic lakes and ponds have consequently been invoked as potent landscape attractors for Early Holocene foragers in Northern Europe (Boethius et al., 2017) and Welinder (1978) specifically suggested that overgrown lakes formed central elements of Maglemose adaptations. Advanced stages of eutrophication and pond maturation, as discussed above, may conversely complicate spearing and bowing and thus inspire new fish-getting strategies.
Contrasting with the Dutch and Northeastern European data, the Late Mesolithic and Early Neolithic record from Southern Scandinavia shows a clear break in ichtyofaunal species compositions (cf. Figure 6). EBK sites demonstrate an increased emphasis on both marine mammals and marine fish (cf. Figure 7). The emergence of coast-bound and increasingly marine economies in the EBK may have thus considerably weakened the probiotic effect of the beaver on human livelihoods and drawn human attention away from beaver habitats, thus simultaneously defusing, or at least re-configuring, the reliance on beaver-engineered, inland foraging affordances. To over-exaggerate, Early and Middle Mesolithic foragers in this region encountered the beaver as a key society-sustaining agent – as a nonhuman fishing aid or more-than-human fishing technology – whereas Late Mesolithic/Early Neolithic societies increasingly relied on their own fishing infrastructure and self-devised fish-getting technologies at coast-inland interfaces, yet perhaps nonetheless inspired by the transgenerational experience of beaver geopraxis. The nature and significance of a human-beaver ‘contact zone’ as envisioned by Hjørungdal (2019b; 2019a) for the Southern Scandinavian Mesolithic thus likely depended on the critical intersection of lived human and beaver geographies and, perhaps more importantly, the extent to which beaver habitats were routinely visited, and thus integrated into broader forager landscapes. In analogy to other documented, integrated human foraging strategies such as ‘garden hunting’ in the Americas (Guiry et al., 2021; Linares, 1976; Stahl, 2020), we may refer to this strategy as ‘pond hunting’ or ‘wetland foraging’. The development of coastal and open-water economies in the course of the Late Mesolithic in Southern Scandinavia would have contributed, then, to the disruption of these broader foraging schemes centred on the diverse resource opportunities in and close to beaver habitats.
Other multispecies affordances
Other indirect ecosystem effects of beaver geopraxis with benefits for hunter-gatherers include waterfowl encouragement and promotion (Brown et al., 1996; Nummi and Hahtola, 2008; Nummi and Holopainen, 2014) as well as beaver ‘gardening’. Waterfowl encouragement is a notable life service for human co-inhabitants as waterfowl was an important subsistence good for meat and feathers (e.g. Zhilin and Karhu, 2002) and this bird category is prevalent in the Mesolithic archaeozoological record of the study regions (e.g. Çakirlar et al., 2019; Lauwerier et al., 2005; Lõugas et al., 2017; Zhilin, 2014a). Water birds are also occasionally rendered in the visual art of the Mesolithic of Western Russia (e.g. Lozovskaya, 2021), coevally pointing to the prominence and potential abundance of these birds in earlier Holocene environments. Waterfowl presence draws in and sustains larger birds of prey such as ospreys and sea eagles and these species have often received special attention by Mesolithic foragers across Northern Europe (Amkreutz and Corbey, 2008; Hussain, 2023a; Mannermaa, 2013), again indexing the crucial role of beaver habitats in framing human forager life, perception, culture and possibly cosmology in the European north. Another example of the important and consequential role of beavers in modulating larger multispecies communities is provided by the pond turtle (Emys orbicularis), who is known to be strongly promoted by beaver wetland engineering and pond-making (Janiszewski et al., 2014) and is well-represented in the archaeology of the earlier Mesolithic of Northwestern Europe (Groß, 2017: 18). Pond turtles notably also make an appearance in the Dutch Late Mesolithic at the Hardinxveld-Giessendam sites (Oversteegen et al., 2001; van Wijngaarden-Bakker et al., 2001) and equally occur at some Southern Scandinavian Middle Mesolithic sites (Groß, 2017). Interestingly, these reptiles then seem to disappear again from the archaeological record during the Atlantic, and this may be linked to a shift in beaver preponderance, landscape impact and/or proximity to human habitation sites in some northern areas at the end of the study period.
In addition, beaver behaviour effectively conforms to a form of landscape gardening – encapsulated by the notion of a ‘beaver meadow complex’ – which promotes distinct plant community successions (Westbrook, 2021), and can notably encourage aquatic plants that grow and proliferate under eutrophic conditions. Water lilies (Nympahea) and water chesnuts (Trapa) both benefit from beaver-induced damming and pond formation (Benke et al., 1999; Kukuła and Bylak, 2017; Law et al., 2014) and the remains of both plants have been found in Northern Mesolithic sites as well as in human coprolites (e.g. Bakels et al., 2001; Kubiak-Martens and van der Linden, 2022; Price, 1991; Zvelebil, 1994). Beavers are known to strategically feed on white water lilies (Nymphaea alba) while promoting them ecologically (Law et al., 2014), and beaver habitats therefore not only signal the potential availability of high-value plant resources, Mesolithic people may have actually discovered this specific resource potential of wetland-lakeland ecosystems by observing and learning from beaver behaviour. Considering the possibility of beaver-directed mimicry as an important ecocultural practice may be fruitful in this context. Mimicry (sensu Bhabha, 1984; GoGwilt and Holm, 2018) describes the ability to imitate or re-enact a socially relevant other in asymmetric power-relations and may be evidenced by co-opting beaver body parts and using them in the same way as the animals, for example deploying beaver-mandibles for woodworking. Such appropriation of nonhuman capacities through material practices, in this optic, may then not only be an expression of relating to beavers as socially relevant others, but could reflect human attempts to literally assume a beaver gaze, as the beaver comes into view as a nonhuman guide and tutor of the north (see e.g. Stobiecka, 2022 for a general exposition of these latter notions), disclosing unique possibilities of navigating and using the landscape. This perspective brings us close to an understanding of Mesolithic beavers as a nonhuman landscaping technology, even though reducing beavers to purely instrumental roles would obviously undermine the gist of the here proposed argument.
All of this being said, beaver-shaped landscapes clearly invite particular foraging behaviours and offer exceptional possibilities for integrated food-getting strategies, but to exploit these efficiently requires intimate knowledge on beavers and their geopraxes. We thus propose that adapting to earlier Holocene environments in Northern Europe in many cases involved human adaptation to beaver behaviours and landscapes. The beaver, in line with Coles’s (2006) previous arguments for Britain, was thus likely a key agent in the Early Mesolithic (re-)occupation of high-latitude Europe as a whole, and ‘landscape learning’ (sensu Rockman, 2003) was promoted by attunement to and familiarization with beaver activity, including efforts to assume a beaver perspective, drawing the beaver into human affairs, at times merging human and beaver horizons, and thereby fostering human respect and care for beaver others, as ‘thinking with’ and ‘acting with’ the animals emerged as an important touchstone of Mesolithic forager life in different parts of Northern Europe.
Human-beaver cohabitation in the Early and Mid-Holocene
The Mesolithic data reviewed and synthesized here may thus be taken to suggest that exploiting the attractive foraging grounds curated by ongoing beaver ecosystem engineering was a central pillar of the earlier Holocene human occupation of Northern Europe, indicating that forager lifeways were at least in part predicated on beaver agency. Human-beaver cohabitation and its associated behavioural possibilities, in other words, emerged as an important precondition for human sustenance and livelihood within the vast wetland and boreal zones of the European north, only to be disrupted when some Late Mesolithic and Early Neolithic societies turned their attention to the sea (and perhaps to other foci of economic, cultural and cognitive concern). The Mesolithic/Early Neolithic of the Dutch wetlands, interestingly, shows notable similarities in overall beaver-related ecosystem relations with the Baltic and the Russian West, rather than Southern Scandinavia, as may initially be expected based on geographic proximity. This convergence may in part be a consequence of the rich delta landscapes in these regions, acting as biodiversity contraptions with extensive wetlands and catalysing beaver habitation as well as the attendant successional dynamics for human foragers (cf. e.g. Giosan et al., 2014; Richardson et al., 2021). Beavers have been shown to play key roles in the maintenance of these landscapes and the provisioning of associated deltaic wetland resources (Hutchings and Campbell, 2005). The cognitive and less tangible aspects of human-beaver cohabitation in these regions are more difficult to glean and open up a host of new questions. Compellingly, however, the here-adopted beaver perspective suggests that geographically close regions in the European Northwest appear to have embarked on divergent ecocultural trajectories, with notable differences in the place of beavers in multispecies systems.
In contrast to the Northwest, the European Northeast stands out in the richness and diversity of its beaver-related material culture. The productive co-habitation and cross-fertilization of humans and beavers in the Early and Mid-Holocene has thus arguably laid the foundation for an increasingly diverse repertoire of beaver-related material practices, understood here as a consequence of fostered human-beaver intimacies throughout the Mesolithic. Extended life-histories of beaver-sourced tools and the human care put into them (cf. Zhilin, 2020) as well as the association between the teeth and astragali of beavers with human bodies strongly suggests that beavers participated in the making of the human social world and became entangled, albeit differently across regions and time periods, with human bodies. The evidence is therefore consistent with the idea that beaver landscape significances became deeply sedimented into cultural memory systems as beavers and humans curated relatively stable and increasingly meaningful interspecies neighbourhoods.
The observation that beaver-related material culture appears in the archaeological record of Northwestern Europe less frequently than in the Baltic and Western Russia and generally tends to postdate the Early Mesolithic is thereby important, potentially showing that beaver knowledge took more time to crystallize in the respective human societies of the northwest, although the comparison may be hampered by the lack of sites from the Dutch Early and Middle Mesolithic. In all regions, however, there is some evidence for the co-optation of beaver capacities through the use and transformation of selected beaver body parts, which embody and exemplify the environmental agency of the animals.
Animal body-part selectivity is a common feature of forager zoo-materialities, often linked to broader concepts of trait fluidity and bodily transposition rooted in relational epistemologies and situated zooontologies emerging from lived interspecies intimacies (Hill, 2011, 2019; Hussain et al., 2022; McNiven, 2022). Beaver-related materializations linked to human instrumental, ornamental, and burial practices, from this perspective, may have helped to produce human-beaver co-sociality, pointing to the socio-historical efficacy and lived significance of beaver neighbourhoods for Mesolithic people. Beaver-related material culture can then be interpreted to reflect the recognition of beavers as ‘co-workers’ and as ‘community’ (sensu Welden, 2023), and thus as a symptomatic feature of what Bird-David (2017) has termed ‘plurispecies’ societies.
The systems-perspective on human-animal interactions pursued here places particular emphasis on the relational assembly, integration, consolidation and disintegration of humans, beavers, landscapes, and material cultures in the course of the Early and Mid-Holocene. It queries the changing ‘intra-actions’ (Barad, 2007; cf. Kirksey, 2015) within these systems, relationships such as conflict, tension, synergy, cross-pollination and possibly co-constitution. Figure 8 attempts to outline the central place of the beaver in these systems – in human world-making during the Mesolithic of Northern Europe as suggested by our analysis and broader discussion. In total, we argue that the beaver’s role as a potent ecological keystone species in the Early and Mid-Holocene of Northern Europe provided the larger context for a prehistory of human-beaver sympoiesis (sensu Haraway, 2016). The status of the beaver as a socially significant other in the Northern Mesolithic was thereby not given but made, and it emerged out of an extended but inter-regionally diverse history of human-beaver co-habitation in the north. Beaver remains and beaver-related material culture, in this view, trace the millennial-scale transformation of the beaver into a ‘cultural keystone species’ (Garibaldi and Turner, 2004; Jacques-Coper et al., 2019; Platten and Henfrey, 2009). As we have seen, the timing and trajectory of this process differs between Northwestern and Northeastern Europe and within Northwestern Europe, and articulates with other documented patterns in the archaeological record. The cultural keystone status of the beaver appears to be conserved in the Northeast and in the Dutch wetlands in the course of the Mesolithic, while human-beaver relationships in Southern Scandinavia appear to be transformed, and perhaps lose their former significance, as human practices are subject to dramatic changes and other animals such as the wild boar (e.g. Magnell, 2006; Maring and Riede, 2019) and marine mammals including killer whales (Andersen and Crabb, 1996) rise to economic, ecological and cultural prominence during the Mid-Holocene.
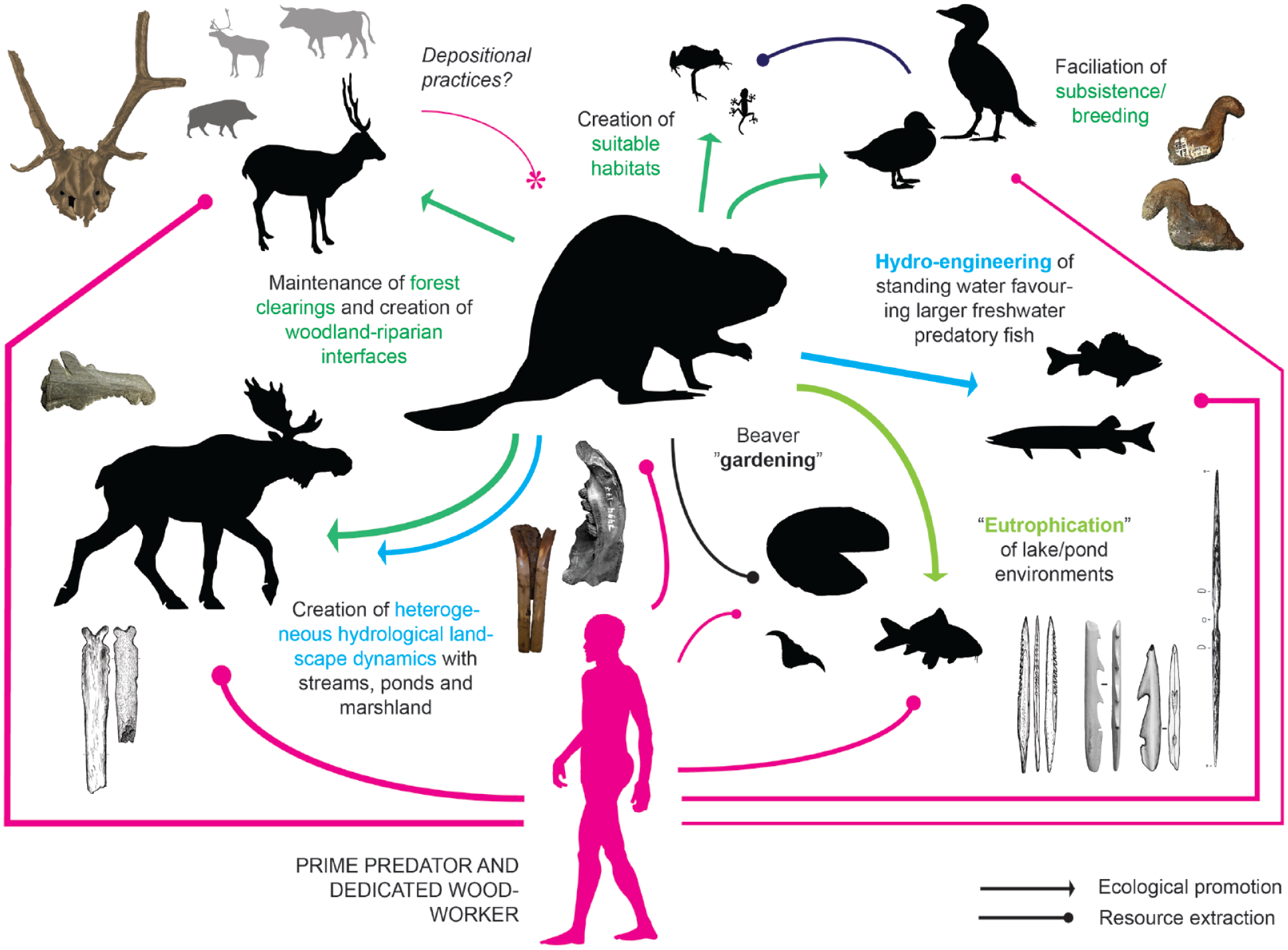
Tanglegram of human and beaver practice in the Northern Mesolithic. Beaver agency and geopraxis structure human behavioural systems and provide key affordances for subsistence (hunting, fishing, fowling and wetland gardening) as well as the production, curation and signification of material culture. Human-beaver co-living provides a key adaptive background for human forager life in the European North during the Early and Mid-Holocene. Human practices and materialities, in this view, cannot be properly understood outside of their attendant multispecies systems.
Outlook: The agriculturalist hypothesis
Based on historical data, Liarsou (2013, 2015) has provocatively suggested that the relationship between humans and beavers is re-tailored as humans introduce and invest into new landscape practices such as pastoralism and farming. There are several reasons for this general tendency and many of them have to do with human encroachment and/or destruction of beaver habitats. Increasing population pressure, intensification of economic activities in aquatic areas, deforestation, and cereal cultivation in sync with growing infrastructural and environmental fingerprints, including expanded riparian transportation, can have detrimental effects on the size and distribution of beaver populations, in addition to curtailing and fragmenting beaver habitat and mobility. Changing human relations to the landscape, especially claims to and early annexation or ‘propertization’ of particular places, can also provoke changes in the perception and conceptualization of beavers, frequently shifting human attitudes, as the attention is readily drawn to interactive tension and possible conflict. The beaver’s landscape-altering capacities are then easily cast as ‘destructive’. This is particularly the case when humans become perennial cultivators themselves since beavers may flood and/or severely damage fields and larger agricultural landscapes (Philip, 2022), thus changing the conditions and context of human-beaver interaction. Unsurprisingly, such dynamics form also a major source of present-day beaver-landowner tensions in reintroduction areas, such as Britain, the Netherlands, and Denmark (Coz and Young, 2020; Jansman et al., 2016; Naturstyrelsen, 2020) .
It is therefore possible that the arrival of agricultural life in Northern Europe marks an important turning point in human-beaver relations, and there is indeed evidence for substantial human population growth in this period (Shennan, 2013; Shennan and Edinborough, 2007), coupled with increasing evidence for aquacultural investment in the form of fishing infrastructure and extractive freshwater economies (Amkreutz, 2013; Beerenhout, 2001b; Price, 1985, 2000, 2015) as well as expanded riverine and oceanic transportation, at least in some areas such as Southern Scandinavia, and new systems of livestock management (Brusgaard et al., 2022b; Gron et al., 2016). In Southern Scandinavia including Northern Germany, the transition to agropastoral systems occurs around 4000 BC (Gron and Sørensen, 2018). A decrease of beaver representation within faunal assemblages can already be observed from the Middle to Late Mesolithic onwards, however, well before this poignant transition. In the Netherlands, sites with the first clear evidence for crop and animal management date to around 4250 BC (Brusgaard et al., in press; Dreshaj et al., 2023) and continue to yield relatively high percentages of beaver remains. While beaver abundances decline later in the Dutch Neolithic, this is the case for wild animals in general, in tandem with the increase of domesticated livestock (Çakirlar et al., 2019; Lauwerier et al., 2005). Wetland landscapes and natural resources remain generally important throughout the Dutch Neolithic and even the Bronze Age, with specialized sites continuing to be used for fur-animal extraction (Dusseldorp and Amkreutz, 2020; Zeiler, 1987).
At first glance, therefore, our analysis reveals no clear indications for a correlation between the transition to agricultural practices and human-beaver relations. Yet full-blown farming systems do not become established in the region before the later part of the Mid-Holocene and perhaps even later, so these changes may be delayed. It is also important to note that the expected developments in human-beaver relations may manifest themselves in different ways in the (zoo)archaeological records. Agricultural conditions may for example greatly favour the classification of beavers, together with other wildlife, as ‘pests’ (Liarsou, 2013: 177) and may thus foster concerns to remove them from human landscapes and lifeworlds, in turn motivating targeted ecological suppression or even overhunting. For example, agro-horticulturalists in Mexico carry out subsistence hunting of species that otherwise pose a threat to crops (Santos-Fita et al., 2012), which ironically occurs at the edges of human-shaped habitat which provides attractive habitat for these species. Other lines of evidence, such as harvesting profiles which can inform on the sustainability of hunting practices, may offer additional insights here in the future. Çakirlar et al. (2019) have for example concluded that there is currently no indication for beaver overhunting at Dutch Late Mesolithic sites as the corresponding harvesting profiles indicate mostly adult-oriented hunting, data which these authors interpret as being consistent with a stable source population. However, these profiles are based on a now outdated method of age-determination for beavers (Iregren and Stenflo, 1982) and thus need to be revisited. It is furthermore important to consider that forager hunting patterns may have been less influenced by utility-oriented decision-making predicated on universal, neoclassical notions of ‘rational choice’ than by situated animal ethologies and lived predator-prey responses, including so-called ‘ecologies of fear’ (Brown et al., 1999; Brusgaard et al., 2022a, 2022b; Holmern et al., 2006; Hussain, 2023c; Zanette and Clinchy, 2019). To account for such dynamics requires to acknowledge the ‘bounded rationality’ (Simon, 1957; Wheeler, 2020) of human behaviour, drawing attention to a possible role of diverse nonhuman agents in steering and stabilizing human foraging systems and their intrinsic rationalities. Beaver agency in Mesolithic hunting practices thus certainly demands further comparative investigation, which may in turn shed novel light on the influence of emerging agricultural systems and other landscape practices on beaver relations, behaviours and populations.
An interesting perspective for future work on later Holocene societies are the grazing affordances potentially emerging in aged beaver landscapes. Beaver landscapes where the animals’ activity has turned formerly flooded areas into meadows (Westbrook, 2021) may so provide localized feeding opportunities for human-accompanying livestock. Meadows can in this way act as ‘alluvial grasslands’ (Hejcman et al., 2013; see esp. Ritchie, 2017 for an illuminating historical account) and so serve as natural pastures (Liarsou, 2013: 175), thus offering land-use affordances which differ from the kind of beaver-supplied affordances for foragers without livestock (cf. Coles, 1992). Eriksson (2020) has thoroughly illustrated how meadows, once created, recruit novel human-animal entanglements and thus become central places or even focal points for people in the landscape, with important consequences for the assembly and dynamics of larger multispecies assemblages.
When contrasted with these possible later structural transformations in human-beaver systems, Mesolithic engagements with beavers can be described as ‘commensal’ (Liarsou, 2013: 178; see O’Connor, 2013 for a general account of commensalism) – with human foragers being commensal to beavers. This divergence suggests a changing status of beavers as ‘companion species’ in the sense of Haraway (2008) in the past, linked to important transformations in human lifeways and behavioural regimes. Contextualizing the available archaeological data for human-beaver interactions from the Early and Mid-Holocene of Northern Europe in this way helps to recognize that beavers may have offered different affordances and life services for different types of human societies, and that beaver landscapes, as a result, were likely unequally perceived, valued, and imagined because of these differences. While beaver agency in the earlier Mesolithic helped to anchor and spatially organize forager lifeways, and thus co-structured and fuelled human ‘taskscapes’ (sensu Ingold, 2022), beavers appear to have been gradually excluded from the centre of Southern Scandinavian lifeworlds in the course of the later Mesolithic, while mutually conducive relationships were seemingly maintained in the Dutch and Northeastern European Late Mesolithic. The latter thereby promoted large-scale diversification and promulgation of beaver-related material culture with continuities at least into the developed Neolithic.
In general, the documented dynamics clearly expose the fragility of northern systems of human-beaver conviviality that have evolved over millennia and that greatly depended on human practices allowing beavers to enter the realm of social significance and to garner human concern and care. These archaeological insights on the millennial-scale dynamics of multispecies systems involving beavers and humans – echoing Liarsou’s (2013, 2020) earlier arguments – are important sources of information for the ecological management and restoration of beavers in the present, as they demonstrate the inseparability of human lifeways and the functioning of human-animal systems that also feature beavers. Coupled archaeological and geological research can make highly significant contributions here as the vast majority of millennial-scale beaver landscape legacies remain undocumented, and this even though the cumulative ecosystem and geomorphological impacts of the hundreds of millions of beavers who once modified rivers and floodplains across the northern hemisphere can hardly be underestimated (cf. Wohl, 2021).
Conclusion
The archaeological evidence drawn together and re-contextualized in this paper from a dedicated multispecies perspective points to an important role of the beaver in the making of Early and Mid-Holocene forager societies in Northern Europe. The evidence is consistent with a role of beavers as important life-service providers for diverse human hunter-fisher-gatherers trying to establish themselves in the wetland and boreal-taiga zones of higher latitude Europe, and thereby showcases the distinct material generativity and vibrancy of human-beaver relations in this region. We have argued that beaver-related material culture is rooted in evolved modes of human-beaver cohabitation characterized by facilitation and mutuality, insofar as human life relied on and took advantage of beaver presence, and beavers, in turn, were acknowledged and belaboured as autonomous but significant social others, reflected in human material culture. We therefore suggest that much of human prehistory in the earlier part of the Holocene in Northern Europe can be re-framed as the result of developing human-beaver convivialities, with human practices drawing on, and increasingly acknowledging, a broader ‘beaver commons’ – that is, beaver-provisioned resources shared with human foragers to sustain them in the landscape. This not only exposes the multispecies constitution of the Northern Mesolithic, it also suggests that becoming-with beavers was a foundational condition of human life in the period. Beavers, in this view, contributed in distinct ways to Mesolithic developments across the European north, as embodied agents but also through co-shaping mammalian and ichtyofaunal assemblages. The latter also calls for more critical zooarchaeological attention to possible, and hitherto underrated (or unrecognized), animal ecological legacies in faunal datasets, as well as to the relationship between such legacies and other, for example taphonomic, biases.
Altogether, we have suggested that the archaeology of Early and Mid-Holocene continental Northern Europe reveals divergent co-occupational histories of humans and beavers, and that important re-negotiations as to the place of the beaver in larger multispecies systems of Southern Scandinavia may be linked to the emergence of coast-oriented lifeways and possible disruptions of human-beaver intersections incurred by agropastoral systems. These disruptions are not a historical necessity, however, but appear to be context-dependent, as the impact of agropastoralism on Northern European human-beaver relations likely varies greatly across different regions. We propose that these dynamics, many of which require further empirical investigation and qualification, thereby offer valuable information and insight for contemporary concerns of beaver restoration, rewilding, ecosystem management and biodiversity stewardship more broadly, as they contribute to a better understanding of the human dimensions and requirements of living together with beavers. The ‘beaver lens’ deployed in this paper thus not only provides a new perspective on the Northern Mesolithic and its ecocultural fabric by centring an archaeologically often underestimated animal, it also showcases the unique and important role that archaeology can play in elucidating the long-term, millennial-scale contributions of animals to human history by highlighting the conditions in which they can help securing human livelihoods.
Supplemental Material
sj-pdf-1-hol-10.1177_09596836231200444 – Supplemental material for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens
Supplemental material, sj-pdf-1-hol-10.1177_09596836231200444 for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens by Shumon T. Hussain and Nathalie Ø. Brusgaard in The Holocene
Supplemental Material
sj-pdf-2-hol-10.1177_09596836231200444 – Supplemental material for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens
Supplemental material, sj-pdf-2-hol-10.1177_09596836231200444 for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens by Shumon T. Hussain and Nathalie Ø. Brusgaard in The Holocene
Supplemental Material
sj-xlsx-3-hol-10.1177_09596836231200444 – Supplemental material for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens
Supplemental material, sj-xlsx-3-hol-10.1177_09596836231200444 for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens by Shumon T. Hussain and Nathalie Ø. Brusgaard in The Holocene
Supplemental Material
sj-xlsx-4-hol-10.1177_09596836231200444 – Supplemental material for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens
Supplemental material, sj-xlsx-4-hol-10.1177_09596836231200444 for Human-beaver cohabitation in the Early and Mid-Holocene of Northern Europe: Re-visiting Mesolithic material culture and ecology through a multispecies lens by Shumon T. Hussain and Nathalie Ø. Brusgaard in The Holocene
Footnotes
Acknowledgements
We thank the participants of the various academic conferences on which aspects of this paper were presented for their valuable feedback and critical comments. Kamilla Laustsen Lomborg helped with bibliographic research in an early stage of this project and provided some important input on the work presented here. Nick Overton, Ben Elliot, and Jörg Ewersen offered their expertise on and excitement about beavers. We thank the archaeological companies Archeoplan Eco, BAAC, and RAAP for providing unpublished data from recent excavations.
Author contributions
STH and NOB have contributed equally to this research.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: STH acknowledges funding received through the CLIOARCH CoG project from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement No. 817564). NØB was funded by a MSCA PF and her project ‘When Keystone Species Converge’ is funded by the European Union (grant agreement No. 101106017).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.