
Abstract
Recent zonal steppes of Eastern Europe and South-West Asia have their origin in the glacial steppe with its specific continental climate. The westernmost edge of the steppe belt in Central Europe has been climatically shifted from a continental course during the Last Glacial to a rather oceanic one during the Holocene. Steppe enclaves could survive only locally in the driest parts of this area, but as we have confirmed, early arrival of Neolithic agriculture played a significant role in their preservation as well. While pollen analyses can provide the main pattern of landscape development, mollusc successions provide more information about landscape details we need for a better understanding of a steppe transformation towards the recent agricultural ecosystems. Based on the study of 18 mollusc successions in two neighbouring lowlands in the chernozems and alluvial areas of Central and North Bohemia, we described a postglacial development of these two climatically different agricultural landscapes. Based on mollusc evidence, we illustrated the impoverishment of forest communities and the continual occurrence of open habitats throughout the forest climatic optimum of the Holocene. Since the Neolithic colonisation, several erosion events were observed in many profiles documented by a lack of preserved fossil molluscs and layer redeposition which are in excellent agreement with the reconstruction of the fluvial sedimentation and vegetation in the Elbe River floodplain and with the continual settlement of this area. Because we have evidence of a fully developed forest fauna in the Eem interglacial from the Polabí lowland we know that the impoverishment of forest communities and the occurrence of open habitats during the interglacial forest climatic optimum is characteristic only of the Holocene.
Introduction
The so-called steppe question, which means the survival of zonal steppe habitats in the western outputs of the Euro-Siberian steppe belt during the Holocene forest optimum (Gradmann, 1906), has been the focus in many studies mostly based on recent vegetation (e.g. Chytrý, 2012; Chytrý et al., 2007; Korneck, 1974; Mahn, 1965), pollen succession evidence (Pokorný et al., 2015), and mollusc succession (Juřičková et al., 2013a). There is no doubt that the recent zonal steppes of Eastern Europe and South-West Asia have their origin in the glacial steppe defined by the continental climate course with low annual precipitations and large temperature differences between summer and winter (Tarasov et al., 2000). However, the western end of this steppe belt in Central Europe has been climatically shifted from a continental course during the Last Glacial to rather oceanic during the Holocene. This is because the steppes could survive only locally, in the driest parts of this area, usually affected by the rain shadows of mountains. The annual precipitation of these areas is recently around 500 mm. In addition to the extreme climate, these steppe enclaves could be kept open by large herbivores grazing (Vera, 2000; but Birks, 2005), thus the first farmers probably changed open chernozem areas to agricultural landscapes (Dreslerová and Pokorný, 2004). But there is not enough evidence for either.
We focus here on two neighbouring areas – the drier Polabí and more humid Pojizeří lowlands in the Czech Republic, where agricultural land use appeared very early (for review see Pavlů, 2007). These lowlands are built partly by chernozems situated on glacial loess sediments and partly by alluvial sediments. Today, it is an agricultural area with an absolute minimum of natural habitats, so it is difficult to get an idea of the natural appearance of this landscape. It is clear, that broadleaf forests did not cover this entire area before the onset of Neolithic settlement and open habitats survived the Middle Holocene Forest optimum (Juřičková et al., 2013a; Pokorný et al., 2015). Abraham et al. (2016) estimated the range of open areas in the Polabí Lowland at 9%. Moreover, the undergrowth vegetation of the local forest was probably very impoverished, as evidenced by fragmented woodland mollusc communities, which lack the most demanding forest species as evidenced in the surrounding Lower Ohře Lowland (Juřičková et al., 2013a). The hilly landscapes surrounding the lowlands, such as the volcanic area of the České Středohoří Mts., also show a reduced woodland malacofauna affected probably by human impact in forests (Juřičková et al., 2013b). On the other hand, in the eastern more humid part of the chernozem lowlands (see Figure 1), the canopy forests with fully developed forest malacofauna were detected but only as a fragment of alluvium sediments dated to the Bronze Age (3200–3700 cal yr BP), the rest have undergone erosion (Břízová and Juřičková, 2011). A fully developed forest fauna also existed in the lower Poohří (Kovanda et al., 2005) and Polabí (Ložek and Šibrava, 1968) regions during the late Eem interglacial.
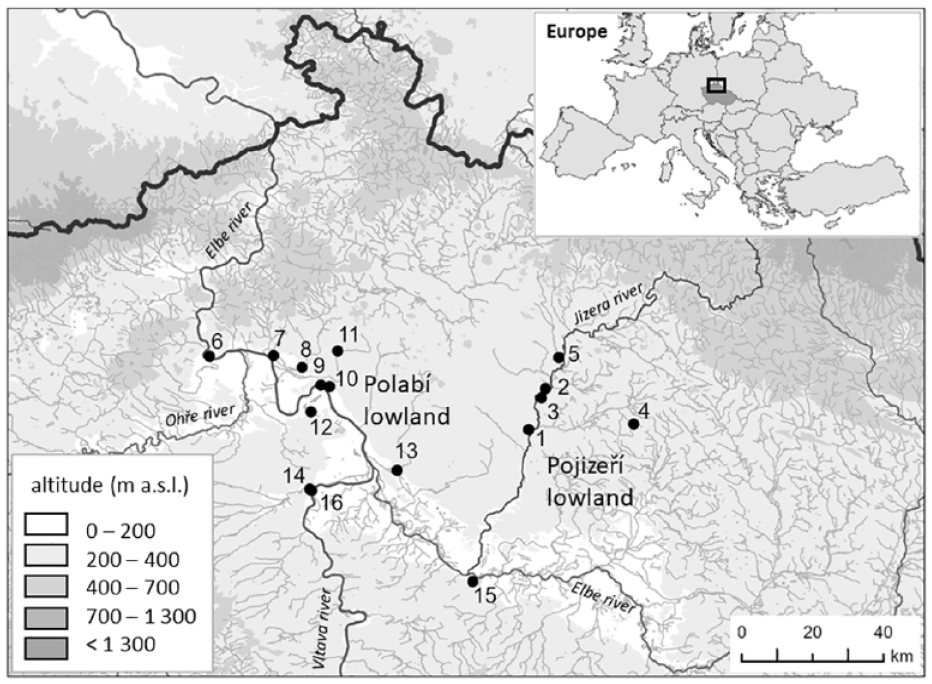
Location of the study area in Europe and Czechia, showing both lowlands and their Holocene profiles (black dots). Pojizeří lowland: 1 – Choboty (50.40416N, 14.87305E), 2 – Zvířetice (50.46916N, 14.90553E), 3 – Josefův Důl (50.45457N, 14.90553E), 4 – Vlčí Pole (50.41305N, 15.13861E), 5 – Ptýrov (50.51866N, 14.94933E); Polabí lowland: 6 – Žernoseky (50.52030N, 14.06519E), 7 – Křešice (50.52173N, 14.22723E), 8 – Vrutice (50.50305N, 14.29944E), 9 – Štětí, Zdymadlo (50.47499N, 14.34666E), 10 – Štětí, Pod Hošťákem (50.47241N, 14.36856E), 11 – Velký Hubenov (50.52888N, 14.38916E), 12 – Bechlín (50.43249N, 14.32194E), 13 – Mělnická Vrutice (50.33917N, 14.53961E), 14 – Vepřek, Lindov (50.30963N, 14.31904E), 15 – Čelákovice (50.16166N, 14.73111E), 16 – Vepřek, Na Hebroně (50.30694N, 14.32277E).
The Late-Holocene erosion has also been recorded in the alluvium of other Central European rivers being attributed both to the widespread influence of agriculture that keep the country open and frequently flooded (Crombé et al., 2019; Kappler et al., 2018; Kołodyńska-Gawrysiak et al., 2018; Kühn et al., 2017; Ward et al., 2009; Wolf and Faust, 2013). Several periods of Holocene erosion were detected directly in the Polabí (Jílek et al., 1995) which caused difficulties in the reconstruction of both settlement development (Dreslerová et al., 2019) and vegetation reconstruction (Petr and Novák, 2014), so the evidence of Holocene development of these lowlands’ areas became scattered.
To complete the picture of the Late Glacial and Holocene development of an important ancient agricultural area, we asked (i) how open habitats have changed from natural Late Glacial steppes to open habitats affected by agriculture, (ii) if the climatic differences of the Polabí and Pojizeří lowlands were reflected in the proportion of open habitats in these areas, and (iii) when and to what extent were the local alluvia affected by erosion.
Material and methods
Site characteristics of Polabí and Pojizeří lowlands
Pojizeří lowland
All five profiles are situated in the area of the Pojizeří lowland in Northern Bohemia. Its geological bedrock is mainly created by more or less calcareous marine sediments (marlstones, sandstones, claystones, siltstones) of the Bohemian Cretaceous Basin and quaternary fluvial deposits of the rivers Elbe and Jizera. Due to its higher altitude ranging from 200 m in the south to 300 to 350 m in the north where it gradually turns into a hilly area, it is more humid than the Polabí lowland. It is characterised by a relatively warm climate with average annual temperatures fluctuating around 9°C in the south and 8°C in the north of the lowland. There is a distinctive north-south climate gradient with an average annual precipitation ranging from 550 mm at the southern edge of the area to 650 mm in the north (e.g. Tolasz et al., 2007). In terms of vegetation, the Pojizeří lowland lies in an area characterised as a thermophyticum (Kaplan, 2012). This lowland area is heavily used for agriculture. Arable lands predominate here and forest monocultures are grown on a smaller area. Currently, there are only a few natural habitats, an important part of which is concentrated in the Jizera river floodplain.
Five mollusc successions were studied in the Pojizeří lowland (Figure 1); four were newly analysed and one (Ptýrov) was already preliminary published by Ložek (1988). Four sites with fossiliferous sediments are situated in the Jizera River valley (Ptýrov, Josefův Důl, Zvířetice, and Choboty) and one site (Vlčí Pole) lies app. 16 km east of them on the left tributary of the Jizera River nearby Dolní Bousov.
The mollusc succession Ptýrov nearby Mnichovo Hradiště is situated at the edge of the foot slope above the Jizera river floodplain. It was excavated in slope deposits of the app. 30–40 m high calcareous sandstone rocks also known as the Ptýrov Cliffs. The sedimentary sequence is created by loamy calcareous sands and screes of 421 cm thickness. There is currently a secondary floodplain forest surrounding the site. This previously published mollusc succession (Ložek, 1988) was supplemented here with detailed mollusc analysis and radiocarbon dating. The Josefův Důl profile is situated nearby Kosmonosy. This tufa deposit excavation of 120 cm thickness on the marlstone bedrock lies at the slope of the valley app. 60 m above the Jizera River. The youngest upper part of the sequence is formed by calcareous loams, the lower part is tufaceous. There is currently a beech wood around the site. The Zvířetice profile nearby Bakov nad Jizerou, is situated at the foot slope of the Zvířetice Hill with Castle ruins of the same name, app. 20 m above the Jizera River. This 340 cm deep profile was excavated in colluvial deposits of calcareous sandstone screes and loams. There is currently mixed forest covering the site and its surroundings. The alluvial profile Choboty nearby Mladá Boleslav has been exposed directly to the Jizera River floodplain. There are fossiliferous fluvial sediments of 200 cm thickness in the bedrock of calcareous sandstones. Sand-rich loams create the upper part of this fluvial sedimentary sequence and the lower part is rich in loamy sands. At present, the surroundings of the profile are built up and/or covered by mixed shrubland. The last mollusc succession Vlčí Pole nearby Dolní Bousov is situated in the plain beside the Jizera River floodplain, app. 20 km east of the Choboty site. These 70 cm thick tufa deposits in the marlstone bedrock lie at the southern edge of a village Vlčí Pole, on one of the smallest left tributaries of the Jizera River. Topsoil here consists of a calcareous humus-rich loam followed by tufaceous layers with clay, weathering product of Turonian marl at the bottom. For detailed lithology and layers description see Figures 1–5 in Supplemental Material 1 (all Supplemental Material, available online).
Polabí lowland
All 13 mollusc profiles are situated in the area of the Polabí lowland which is warmer and less humid than the Pojizeří lowland which adjoins at the eastern edge of Central Bohemia. Its altitude is ranging from 140 m at the western edge of this area (nearby Žernoseky) to 200 m in the east (nearby Čelákovice). The geological bedrock consists also of marine sediments of the Bohemian Cretaceous Basin (marlstones, calcareous sandstones), but several types of fluvial sediments (gravels, sands) of the Elbe river and aeolian sediments as loesses and loess loams on which the most fertile chernozems were formed are predominated here. This whole region is therefore heavily used for agriculture, with arable land and settlement predominating. This is due to favourable climatic conditions with an average annual temperature of 9°C–10°C and average annual precipitation of 500–550 mm (e.g. Tolasz et al., 2007). This region is thus one of the warmest and the most fertile in Bohemia. In terms of vegetation, the Polabí lowland lies in an area characterised as a thermophyticum (Kaplan, 2012).
Thirteen mollusc successions were studied in the Polabí lowland (Figure 1) of which one (Bechlín) was excavated in 2011 and then analysed. The other 12 successions were previously published by Ložek (see below) but without radiocarbon dating and a detailed analysis.
The small Bechlín mollusc succession of only 76 cm thickness was excavated in the alluvium near the small brook spring at the cretaceous weathering. Upper layers are formed by calcareous loams where the fourth layer indicates the edge of the foam pile that is no longer there. The bottom of the sedimentary sequence is without fossil molluscs and consists of clays, weathering products of Turonian marlstones. At present, the location is covered by a floodplain forest.
The Vrutice profile (Ložek, 2006a) is situated at the foot of a south slope in the shallow valley of the Úštěcký brook close to the Vrutice village at the foothills of the České Středohoří Mts. Its surroundings are dominated by fields with patches of natural xerothermic grassland. The depositional sequence was exposed in a small loam pit in the talus below the local railway. The almost 300 cm sedimentary sequence consists of two different groups of strata: basal layers 8–6 are composed of a loess-like matrix, whereas the upper layers 5–1 are dominated by debris from weathering products of Cretaceous marlstones. Not far from here, near the town of Štětí there were four profiles researched – Štětí pod Hošťákem with three pits A, BI, BII, and Štětí Zdymadlo (Ložek, 1991; Ložek and Šibrava, 1968; Smolíková and Ložek, 1978) all situated in the chernozem area. All four successions are situated on the right bank of the Elbe River and created by floodplain and colluvial deposits with complicated stratigraphy (for more details see Supplemental Material 1, available online). The Štětí Zdymadlo profile includes almost 5 m of deposits as humic carbonate chernozems and loess-like soils developed on eolic glacial sediments of this area. The Štětí pod Hošťákem – pit A is a several metres deep profile with only one fossiliferous layer formed by calcareous loess with the last glacial mollusc fauna. All of three pits – A, BI and BII lie on the river Elbe terrace. Pit BI includes a long sedimentary sequence of 413 cm thickness with 10 distinguishable fossiliferous layers consisting of calcareous loams with the last glacial loess at the bottom. Pit BII is also 4 m deep profile but consists only of five fossiliferous layers in its upper part formed by humic clays and loams. The Velký Hubenov profile (Smolíková and Ložek, 1973) was found at the boundary of the chernozem and brown soils area. It is located 8 km north of Štětí profiles at a foothill nearby a small Obrtka River. There are dominated fossiliferous calcareous loams across the upper sedimentary sequence, but the lower three layers are formed by clayey loams without fossil material and the bottom is composed of loess and rhythmically bedded loamy calcareous sand with last glacial mollusc fauna. The other two profiles are situated nearby Vepřek village. Vepřek-Lindov lies in the Bakovský brook valley (Ložek, 1995) and includes 208 cm of fossiliferous sediments. It is created by a Holocene terrace at the right brook bank consisting of calcareous loamy and sandy sediments with fossil molluscs. The non-fossiliferous two layers at the bottom are formed by alluvial stream gravels replaced from the older Vltava terraces and by the bedrock layer of partially weathered Turonian marlstones. The second profile Vepřek-Na Hebroně (Ložek, 2006b) is an archaeological site with prehistoric settlement (Sklenář and Lutovský, 1993). Unfortunately, the detailed lithostratigraphy of the site was not preserved due to complicated archaeological work. A peat bog (recently drained) with fragments of pottery (16th–17th centuries; layer 1), rich in phytopalaeolontological material (Kneblová, 1956), and many bones and mollusc shells were found nearby Křešice town close the right bank of the Elbe River east of Litoměřice (Ložek, 1956). Peat deposits of calcareous clays and phonolite debris at the bottom were laid in shallow depressions between sandy sediments of a valley terrace and preserved three fossiliferous layers of 318 cm thickness with mollusc material. The bottom of this profile is created by sand without any palaeontological material. There was investigated also another Žernoseky profile near the right slope of the Elbe valley consisting of fluvial gravels and fluvial and aeolian loamy deposits of the Late Glacial terrace accumulation (Ibrava, 1972; Ložek, 1989; Ložek and Šibrava, 1982) of 322 cm thickness. Across the entire sequence, a fossil mollusc material was preserved with the exception of the lowest two layers at the bottom formed by non-fossiliferous fluvial sands. Holocene malacofauna was also found near Čelákovice town (Ložek, 1986) approx. 20 m above the Elbe river floodplain. No molluscs were found in the upper four layers of non-calcareous clays, only the lowest layer of Mid-Holocene calcareous clays was fossiliferous in the depth of 76–101 cm. Finally, the thick complex of freshwater lake marl deposits was researched near Mělnická Vrutice town. There was surveyed series of eight test holes of which the first by Obora was the richest and processed in detail (Ložek, 1952). The sedimentary sequence was 270 cm thick and consists of calcareous loams in the upper two layers followed by freshwater chalky clays to the bottom. For detailed lithology and layers description see Figures 6–15 in Supplemental Material 1, available online.
Sampling method
All mollusc successions in the Polabí lowland were obtained between 1950 and 1994 by Vojen Ložek with only one exception of Bechlín researched in 2011 by Lucie Juřičková. In the Pojizeří lowland, sites were also researched in the 1980s by Vojen Ložek. All the mollusc successions were sampled by standard methods (Ložek, 1964) – 8 dm3 of space discrete samples of the sediment were taken from the central part of each lithologically distinguishable layer (see Supplemental Material 1, available online) within 80 cm wide excavation pits because the macroscopically distinguishable changes in lithology mean the crucial changes of habitat characteristics and thus mollusc assemblages. Therefore, there are used much more modern and accurate methods nowadays, at the time of sampling those were advanced methods that could be supported lithologically and often also archaeologically or palaeolontologically. We realise that those sampling methods at present are not optimal for reconstructing very fine environmental changes because the samples could be taken from central parts of the sometimes very thick layers including fossil records of a long sedimentary period. For that reason, we cannot reconstruct so fine environmental variations as enabled by current modern sampling methods which are taken samples from more fine layers of about 5 to 10 cm of thickness (e.g. Granai et al., 2020; Hájek et al., 2016). But we certainly can reconstruct the main changes in environment and biodiversity in the Holocene and Late Glacial periods.
After terrain sampling, mollusc shells were extracted from the sediments by a combination of floating and sieving. After careful drying, each sample was disaggregated in water and then hydrogen peroxide. Floating snails were repeatedly decanted into a 0.5 mm sieve and dried in laboratory conditions. Afterwards, the sediment was dried and sorted by sieving. Shells were systematically removed from the sediment and examined under a binocular microscope.
Ecological groups were used sensu (Juřičková et al., 2014; Ložek, 1964). The nomenclature follows Horsák et al. (2023). Mollusc diagrams expressed absolute and relative proportions of the total number of species (MSI and MSS malacospectra) in separated layers. Classification of Holocene was used sensu (Walker et al., 2012).
Radiocarbon dating
Radiocarbon data analyses were predominantly performed in the Center for Applied Isotope Studies of the University of Georgia, USA, and several of them were dated in the Poznan Radiocarbon Laboratory, Poland, or in the Isotoptech Zrt. in Debrecen, Hungary. Mollusc shells were measured by the AMS (Accelerator Mass Spectrometry) method and calibrated for variable initial 14C concentration using the OxCal v4.1 calibration programme (Bronk Ramsey, 2009). Mollusc shells can contain so-called dead carbon, which can lead to an overestimation of their age (Goodfriend and Stipp, 1983). To minimise this problem, we used an amalgam of all shell fragments that showed no signs of mineralisation (Rech et al., 2011). We know from experience that dating from such material corresponds to data from other standard materials (unpublished data). But we didn’t have them available because we mostly worked with samples taken in the 1980s. The lithology was further used as another proxy (for detailed descriptions and radiocarbon data see Supplemental Material 1, available online) to control the development of particular sites, which is important in such dynamic environments as floodplains.
Results
Successions of fossil mollusc assemblages in the Polabí
Although most of the mollusc successions in the Polabí lowland have been published before, none of the profiles was radiocarbon dated, which turned out to significantly distort their interpretation. Just by looking at the malacodiagrams (Figure 2), we can see that this area was affected by erosion, apparently in several waves. We can recognise the Late Glacial preserved sediments, part of the Middle Holocene sediments, and the best-preserved Late-Holocene sediments. Radiocarbon dating of the model profile Vepřek-Lindov reveals some disturbances and sediments redeposition but probably only in the Late-Holocene sediments, where the superposition of layers is not chronological.
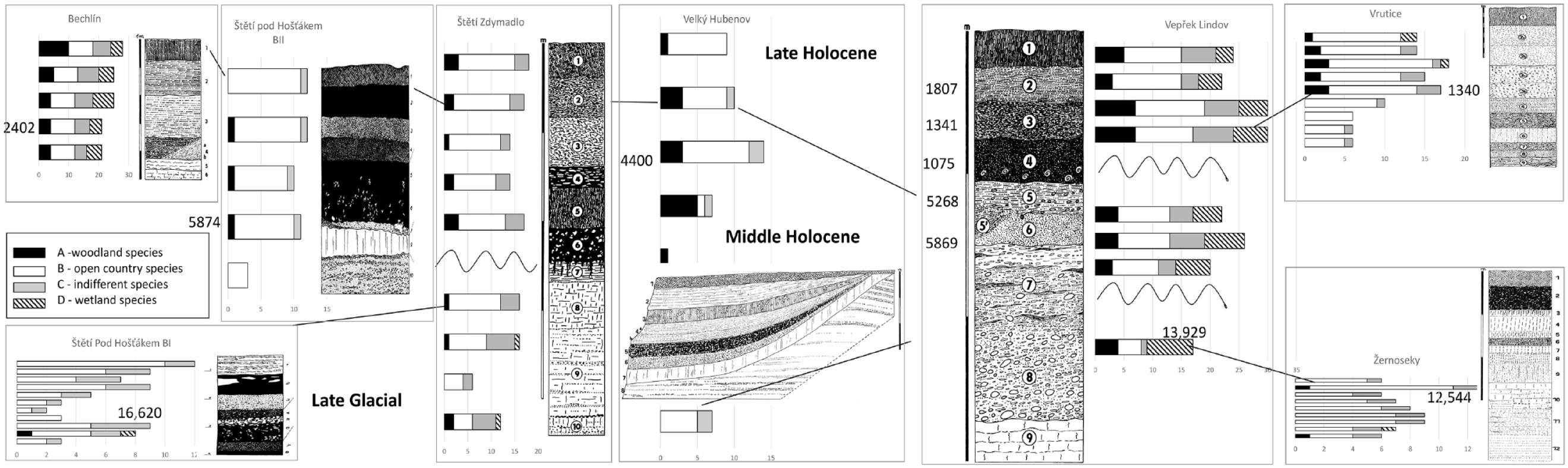
Comparison of the total number of mollusc species (MSS malacospectra) in particular profile layers of nine mollusc successions from the Polabí lowland with their lithology. Ecological groups sensu Juřičková et al. (2014) On the right side of MSS malacospecra are radiocarbon data for particular dated layers cal yr BP.
We can see the predominance of the open-country assemblages in all molluscan successions in the Polabí lowland. Altogether, 94 species were recorded in the 13 mollusc successions, but only 19 of them are forest species whose presence in all profiles is 14%. There are not many open country species (18 spp.) but they are presented in all profiles in 33%. The most common are aquatic species (35 spp.) which are presented in profiles similar to open-country species (Figure 2, for species details, see Supplemental Material 2, available online).
Three profiles contain layers of the Late Glacial age (Štětí-Pod Hošťákem BI – 16,620 cal yr BP, Vepřek-Lindov – 13,929 cal yr BP, and Žernoseky – 12,544 cal yr BP), which mollusc assemblages consist of the open country and indifferent species. The steppe successions started with assemblages of a cold glacial steppe with index species Pupilla loessica and Vallonia tenuilabris that changed during the Holocene climatic optimum to a more thermophilous steppe with Granaria frumentum and even Truncatellina claustralis.
Vepřek-Lindov is a model precise radiocarbon dated profile. The base of the profile is built by alluvial gravel replaced by older river terraces (layer 8) and laying on the Turonian marlstones (layer 9). This bedrock did not contain any shells. The oldest layer (7) with preserved snails is of the Late Glacial age (13,929 cal yr BP) and contains only fragments of shells except for Vallonia enniensis and V. pulchella, indicating an open mineral-rich habitat. The Early Holocene sediments were eroded. Despite layers 5 and 6 being of the Forest Optimum age (5268–5869 cal yr BP) they contain only a few common woodland species. Most species are common open country elements and sensitive wetland species such as Vertigo antivertigo, Pseudotrichia rubiginosa, and Euconulus alderi. Another erosion is related to the older part of the Late-Holocene sediment. Next sediments are dated approximately to the Bronze Age but based on radiocarbon dating that did not fit the superposition of layers, they are re-deposited. Again, only a few common woodland species occurred there, but besides the high occurrence of common open country species, important thermophilous steppe elements appeared such as Caucasotachea vindobonensis, Granaria frumentum, and Xerolenta obvia. Again, a high proportion of sensitive wetland species was found.
Only a minimum of sensitive woodland species in very low abundances (Platyla polita, Sphyradium doliolum, Discus ruderatus) occurred in entire successions in other profiles, clearly indicating the poor character of forests malacofauna in the Polabí lowland. On the other hand, common open country species (ecogroup 5) occurred in all mollusc successions from the Late Glacial to the recent. But sensitive steppe species gradually appeared in the Holocene thermal optimum. Well-preserved wetlands with sensitive species are rare. The best-preserved wetland assemblage (Vertigo moulinsiana, V. genesii, V. antivertigo, Euconulus alderi, Pseudotrichia rubiginosa) was revealed in Mělnická Vrutice (see Supplemental Material 2, available online).
Successions of fossil mollusc assemblages in the Pojizeří
Altogether, 75 mollusc species were found in the fossil records in the Pojizeří lowland, belonging to all ecological groups sensu (Juřičková et al., 2014; Ložek, 1964), including 32 woodland species, presence of woodland species in all profiles is 48%. Open country species (10 spp.) are presented in all profiles only by 16% (for detail see Figure 3 and Supplemental Material 2, available online). The five compared profiles covered probably only the Middle and previously Late-Holocene period, with the exception of the one Late Glacial old layer in the Ptýrov profile. The Early Holocene sediments were eroded (Figure 3). According to radiocarbon data (Figure 3 and Supplemental Material 1, 2, available online), even the Late-Holocene sediments were not found in chronological superposition and were probably repeatedly redeposited, which is visible in the Ptýrov and Choboty profiles. On the contrary, Josefův Důl profile seems to be undisturbed. We have no radiocarbon data for Vlčí Pole and Zvířetice profiles.
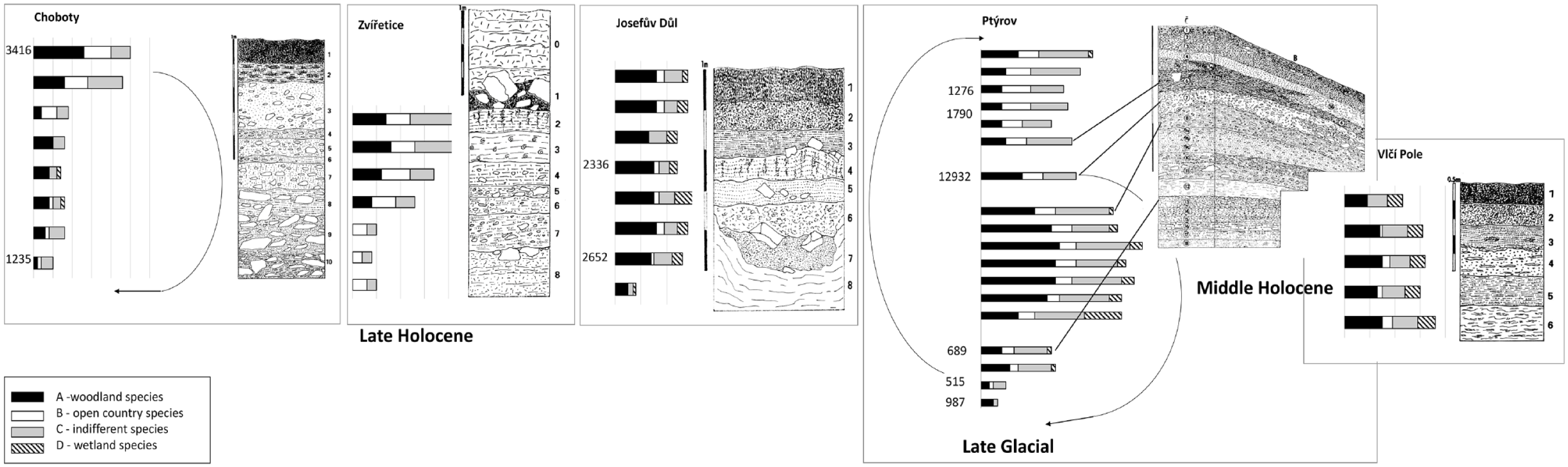
Comparison of the total number of mollusc species (MSS malacospectra) in particular profile layers of five mollusc successions from the Pojizeří lowland with their lithology. Ecological groups sensu Juřičková et al. (2014). On the left side of MSS malacospecra are radiocarbon data for particular dated layers cal yr BP.
Although badly disturbed, the absence of the Early Holocene index mollusc species in Ptýrov profile and radiocarbon data indicate previously Late-Holocene origin. While layers 17 and 18 are completely free of molluscs, only fragments of molluscs were found in layers 13–16. But some vertebrate remains were preserved including Rattus rattus in layer 16, together with further thermophilous open-country vertebrate species in layers 15–16 (i.e. Eliomys quercinus, Microtus arvalis, and Talpa europaea) (Ložek, 1988). The entire lower layer formation was apparently exposed to weathering, which either completely or partially destroyed the shells. Woodland and indifferent species predominate in layers 8–12, and open-country species occur there in lower abundance. It is possible that these layers are of the Late-Holocene age, as inferred the ongoing presence Helicigona lapicida, an index species of this period, but it might be older. The significant proportion of wetland species indicated both – high plants on riverbanks (Succinea putris), as well as short-stemmed wetlands (Vertigo angustior). The layers 4–7 are probably badly disturbed, as is visible in lithology (Figure 5 in Supplemental Material 1, available online) and mollusc assemblages contain previously open country species, while indifferent and non-demanded woodland species are less represented. The wetland and aquatic species are practically absent. Layers 1–3 seem to be undisturbed and contain a mosaic of open country, indifferent, and non-demanded woodland mollusc assemblages. The common occurrence of Clausilia rugosa parvula throughout the entire profile is of particular interest and will be commented on below.
In the Josefův Důl profile, only scattered fossil material was preserved in the lowermost layer 8. However, the lithology indicates marked changes in the type of sedimentation (calcareous clay) in comparison with overlying radiocarbon-dated tufa layer 7 (2625 cal yr BP). Then, fully developed woodland malacocoenoses occurred to layer 4 (2336 cal yr BP). In layers 1–3 cold tolerant Discus ruderatus disappeared and thermophilous D. perspectivus appeared even in layer 1. Otherwise, woodland communities continue to dominate (Table 1 in Supplemental Material 2, available online). The modern immigrant Oxychilus cellarius, and typically floodplain forest species Perforatella bidentata or synanthropic species of nutrient-rich habitats with nitrophilous vegetation Trochulus hispidus also appeared only in sub-recent.
Common open-country and woodland/open-country species dominated the entire profile at Zvířetice (Table 2 in Supplemental Material 2, available online). Several woodland species appeared only sub-recently (layers 1–2), but woodland fauna is reduced. The age of this profile is unknown.
The undated Vlčí Pole profile covered probably part of the climatic optimum of the Holocene because many demanded woodland species occurred there.
The mollusc succession of Choboty (Table 3 in Supplemental Material 2, available online) differs from the others because its specific type of sedimentation above the loess series in which some wedges of a Holocene fossil material were settled. However, radiocarbon data again do not indicate the chronological superposition of the layers, so the sediment is probably redeposited.
Discussion
Continuity of steppe habitats in lowlands and forest fauna impoverishment
The Polabí lowland Late Glacial and Holocene environment has been reconstructed previously based on pollen analyses (Břízová, 1998; Dreslerová and Pokorný, 2004; Dreslerová et al., 2004; Petr and Novák, 2014; Petr et al., 2014) and scattered mollusc findings (Ložek, 1952; Ložek and Šibrava, 1982). On the contrary, we have almost no earlier evidence of the Holocene environment of the Pojizeří lowland, so the mollusc succession described in this study represents the first view of the development of this area. We can therefore only compare our data with previous reconstructions of the Holocene vegetation in the Polabí. Such a comparison shows how reconstructions of landscape details, based on molluscan successions, can complement pollen analyses, which usually show more general landscape characteristics being unable to capture such details.
The Late Glacial mollusc fauna of the Polabí is characterised by species typical on chernozem soils with a deep humus horizon especially Helicopsis striata and Chondrula tridens, but also other common species of loess steppes that are unable to live in light forests such as Pupilla triplicata, P. muscorum, and Vallonia pulchella (Supplementary 2 – Žernoseky, Štětí – Pod Hošťákem BI, and probably Štětí – Zdymadlo – layers 6–9 and Mělnická Vrutice – layers 5–11). The Early Holocene layers of all molluscan profiles were probably eroded. The pollen analyses obtained from bogs less susceptible to erosion characterised the Early Holocene in the middle Polabí lowland by the dominant role of pine (Pinus sylvestris) initially combined with birch (Betula). Novák and Sádlo (2005) assume that the Late Glacial herbaceous vegetation of lighted pine forests can survive in this area for a long time. But the above-mentioned open-country mollusc communities could not survive in such an environment, therefore we assume that some steppes patches must have appeared here at the same time. Novák and Sádlo (2005) also assumed that the immediate vicinity of the Elbe river was soon overgrown by thick willow meadows. But some areas of open wetlands must have persisted here, as evidenced by species such as Vertigo angustior, V. antivertigo, Vallonia enniensis (Supplementary 2 – Vepřek-Lindov, Vepřek-Na Hebroně, Křešice). As a dominant vegetation type on loess sediments of the Middle Holocene were reconstructed mixed oak woodlands (Břízová, 1998). With the onset of the Holocene climatic optimum, mixed oak woodlands spread previously in the river floodplains and pushed pine forests (Novák and Sádlo, 2005) and steppe patches (our data) onto drier parts of river terraces. Petr and Novák (2014) described the gradual migration of Ulmus, Corylus, Quercus, Tilia, and Fraxinus since 8500 BP in the Middle Polabí lowland. Such a forest type would be ideal for the development of rich forest mollusc communities, but none have been found. However, it could also be that large-scale erosion events (discussed below) destroyed the relevant sediments. In any case, we have no dated mollusc communities from the first half of the Middle Holocene. Novák and Sádlo (2005) reconstructed the differentiation of the mixed oak forests into forest communities of roughly current type: acid oak, oak, fir-oak, and beech forests during the Late-Holocene. Because we have a number of radiocarbon-dated mollusc communities from the second half of the Middle Holocene and the Late-Holocene, it shows us a completely different picture of the landscape here. While forest species are minimally represented in this period, open landscape communities are predominant. From this evidence, we cannot conclude that almost no forests occurred there. Such forests could, due to grazing pressure, raking of litter, logging, and other specific forms of management (e.g. Pavlů, 2007) gradually degrade up to cultural forests with very fragmented or almost no mollusc fauna.
Although most of the cited palaeobotanical articles admit the possibility of the occurrence of open habitats in the Polabí lowland, their share of landscape mosaic was mentioned only vaguely. Until Abraham et al. (2016) modelled about 9% of the open habitats (Poaceae) in the Polabí lowland during the Holocene climatic optimum. However, an analysis of the mollusc successions of both the Polabí and the Pojizeří areas shows that the proportion was probably larger.
Similar impoverishment of forest fauna as we documented in the Pojizeří and especially in the Polabí lowlands was found in the mollusc successions in the nearby lower Poohří (Juřičková et al., 2013a) but the proportion of the open-country species in the Polabí surpassed this area. It is therefore clear that as in the lower Poohří (Juřičková et al., 2013a), we have a documented continuity of steppe communities in these lowlands from the Late Glacial across the Holocene. This seems to be a different landscape development trajectory than that described in the Seina Basin in France (Granai and Limondin-Lozouet, 2018), where the transformation of the landscape into an open one takes place in three phases characterised by increasing human influence but demanding steppe or forest elements never occur here, so comparing both mollusc successions is difficult.
Both, in the lower Poohří, and the Polabí we have evidence of a fully developed forest fauna in the Eem interglacial including forest species missing in all the Holocene successions in the Poohří, Polabí, and Pojizeří lowlands (Petasina unidentata, Vitrea diaphana, Cochlodina orthostoma) (Kovanda et al., 2005; Ložek and Šibrava, 1968). Thus, the impoverishment of forest communities and the occurrence of open habitats during the interglacial forest climatic optimum is characteristic only of the Holocene. It can be explained by a combination of the drier Holocene climate in comparison with the Eem (Stebich et al., 2020) and early agriculture impact (Dreslerová, 1995; Dreslerová et al., 2004; Dreslerová and Pokorný, 2004; Dreslerová et al., 2019) amplifying frequent erosion events (Jílek et al., 1995).
Erosions in agricultural lowlands
Most studied profiles both in the Polabí and Pojizeří are badly affected by erosion and redeposition of sedimented material in river alluvia. Many authors show large erosion events in the floodplains of Central European rivers alternating with periods of sediment accumulation (Crombé et al., 2019; Jäger, 1969; Kappler et al., 2018; Kołodyńska-Gawrysiak et al., 2018; Kühn et al., 2017; Ward et al., 2009; Wolf and Faust, 2013). However, these periods do not seem to be synchronous, being area (Kołodyńska-Gawrysiak et al., 2018) or even site-specific (Wolf and Faust, 2013). Nevertheless, many studies emphasise the start of the important erosion periods around 5000–4000 BP. Radiocarbon-dated sediments have been preserved from this period in the Polabí (Štětí pod Hošťákem BII, Velký Hubenov, Vepřek-Lindov) while none was preserved in Pojizeří indicating different erosion history of both area. Most of the aforementioned works consider the most dramatic erosion events that occurred during the past 1000 years. The redeposition of sediments at the Ptýrov locality, when young sediments from the first millennium are covered by older ones. Also, the redeposition in Vepřek-Lindov dated to this period or a little bit earlier confirms this dramatic period. Therefore, we do not have direct evidence on other localities.
In any case, deforested areas are much more prone to erosion than forested ones (Kappler et al., 2018; Kołodyńska-Gawrysiak, 2019; Kołodyńska-Gawrysiak et al., 2018). Ward et al. (2009) showed erosion up to three times higher in a deforested than in a forest-covered landscape. The role of humans as erosion mediators clearly depend on their activities in the landscape. Crombé et al. (2019) bring strong evidence against an anthropogenic origin of Early Holocene erosion. Since Neolithic colonisation, the role of humans in the acceleration of erosion seems to be indisputable. The formative human impact considered by Kühn et al. (2017) is representative of around 9400 km2 of Central European dry loess landscapes.
A similar situation we can observe in the Polabí lowland. The late-glacial or even older radiocarbon-dated layers with molluscs were preserved in four sites (maybe more) in the Polabí. While mosaic of woodland, open country, and wetland habitats are evidenced from mollusc assemblages in Vepřek-Lindov, only steppe habitats occurred in Štětí pod Hošťákem, Velký Hubenov, and Žeroseky sites. The sedimentation continued after probably early Holocene erosion events in Vepřek-Lindov and Velký Hubenov sites, but only old layers were preserved in the two localities left. Jílek et al. (1995) bring the reconstruction of the Holocene fluvial sediments directly in the Polabí lowland. They suggest several abrupt changes in temperature and precipitation during the Holocene in this area. These changes caused periods of hydrological disequilibrium that resulted in the formation of two Holocene terraces, and sedimentation of four layers with tree trunks that correspond to climatic catastrophes. During 10,000–9500 cal yr BP they described the erosion of older sandy gravels and accumulation of sediments of the upper floodplain level; then 9500–9000 cal yr BP old layer contained tree trunks that evidenced dramatic erosion events. We have no directly dated layers with molluscs from these periods, so these layers were probably destroyed by erosion. Around 8000–6900 cal yr BP authors described disruption of the graded profile, fluvial erosion, and redeposition of older sediments including tree trunks. Again, no dated sediment with molluscs was preserved from this period.
Around 5500 cal yr BP the river system reaches an equilibrium and begins to meander and deposit organic material in oxbow lakes. Directly radiocarbon-dated layers were preserved from this quiet period from two sites. We reconstructed there both open habitats only (Štětí-pod Hošťákem BII) and a mosaic of open habitats, degraded forests, and wetlands (Vepřek-Lindov). The onset of the Neolithic settlement was recorded there (Dreslerová, 1995; Dreslerová et al., 2004). Since 3700 cal yr BP Jílek et al. (1995) found again the layer with tree trunks, destruction of river equilibrium, and then sudden renewal of sedimentation of gravels and sands when fluvial sands and gravels overlied the previous surface. We have later radiocarbon data proving the redeposition of layers with mollusc assemblages. For the Late-Holocene period, the redeposited layers seem to be typical in the Polabí lowland. The continual settlement was recorded there (Dreslerová, 1995; Dreslerová et al., 2004).
The comparison of the reconstruction of the fluvial sediments of the river Elbe and the mollusc successions is therefore in excellent agreement, in contrast to the above-mentioned works from the wider Central European area, and proves the dramatic development of the local landscape. Therefore, the absence of sediments with molluscs of the Early Holocene and older parts of the Middle Holocene in our profiles can be well explained by these erosional events. We believe that the open character of the Polabí lowland, clearly evidenced by the mollusc communities, makes this area prone to erosion for most of the Holocene.
Change of the steppe mollusc fauna
Although the Polabí open habitats have continuity from the Late Glacial to the present, their mollusc fauna composition has changed. Certain differences exist between the Late Glacial and Middle/Late-Holocene mollusc communities (the Early Holocene mollusc were eroded) (Figures 2 and 3 and Supplemental Material 2, available online). Common open country species (Vallonia costata, V. pulchella, Vertigo pygmaea, Pupilla muscorum, Truncatellina cylindrica) together with index species of soils with deep humus horizon Helicopsis striata, Chondrula tridens (Ložek, 1964, 2001) occurred there since the Late Glacial. On the contrary, the more demanding thermophilic steppe fauna begins to spread later in two periods. Caucasotachea vindobonensis and Granaria frumentum appeared around 5800 cal yr BP, while Oxychilus inopinatus and Xerolenta obvia seem to be young immigrants in Central Bohemia appearing around 1340 cal yr BP. Both of the latter species indicate disturbed open habitats in Central Europe, that is, more intensive agriculture, so their spread is probably related to this. The exact route, from which these southern European species spread, is not yet known. The glacial refugia of these four thermophilic species are still unknown, with the exception of C. vindobonensis, which originates from the Western Balkans. From there it gradually spread to Central Europe, and we have its oldest occurrence here from the Slovenian Karst from the very beginning of the Middle Holocene (Korábek et al., 2023). In Polabí, where this species has its northwestern temperature-dependent limit of distribution, it is strictly bound to the steppe, although it inhabits a wider range of biotopes further southwest. To the colder and wetter Pojizeří C. vindobonensis gradually disappeared.
The Quaternary mollusc fauna of this area is characterised by the occurrence Clausilia rugosa parvula. This taxon is recently associated with damp limestones and argillite rocks being an ecological specialist. But it was also considered a regional loess steppe element (Ložek, 2001; Moine, 2014). So, this subspecies occurred both in the glacial and interglacial context of the western part of the Central European chernozem area switching its ecology between glacials and interglacials which is quite unique. It seems that during interglacials it could live on calcareous rocks at medium to higher altitudes (220–1100 m) (Hájek et al., 2016; Juřičková et al., 2019, 2020; Moutelíková, 2015; Pavézková, 2019, Pojizeří in this study), while during cold glacial phases, it moves to loess steppes at lower altitudes (170–500 m) (Ložek, 1956, 1965) or exceptionally survives in small karst refugia (Juřičková et al., 2019). During the Holocene, this species no longer occurred on the drier Polabí, where it had lived for two previous glacials (Ložek and Šibrava, 1968) but survives scattered in the Pojizeří recently. Changes in Quaternary and recent distribution of this species indicate the change of climatic gradient between the drier Polabí and more humid Pojizeří.
Mollusc fauna of the Polabí and Pojizeří differs in frequency of demanded steppe and wetland species
If we compare the proportion of open country species in mollusc assemblages of the Polabí and Pojizeří, the answer to our second question is clear. The drier Polabí and more humid Pojizeří lowlands differ significantly (Table 1). While in the Polabí only 14% of forest species were recorded in mollusc assemblages, in surrounding areas the proportion of forest species during the Holocene was 51% in České Středohoří (Juřičková et al., 2013a), 42% in Český ráj (Juřičková et al., 2020), and 48% in the Pojizeří (this paper).
Comparison of mollusc assemblages, species frequencies in particular periods and proportion of their ecological groups in Polabí and Pojizeří lowlands.
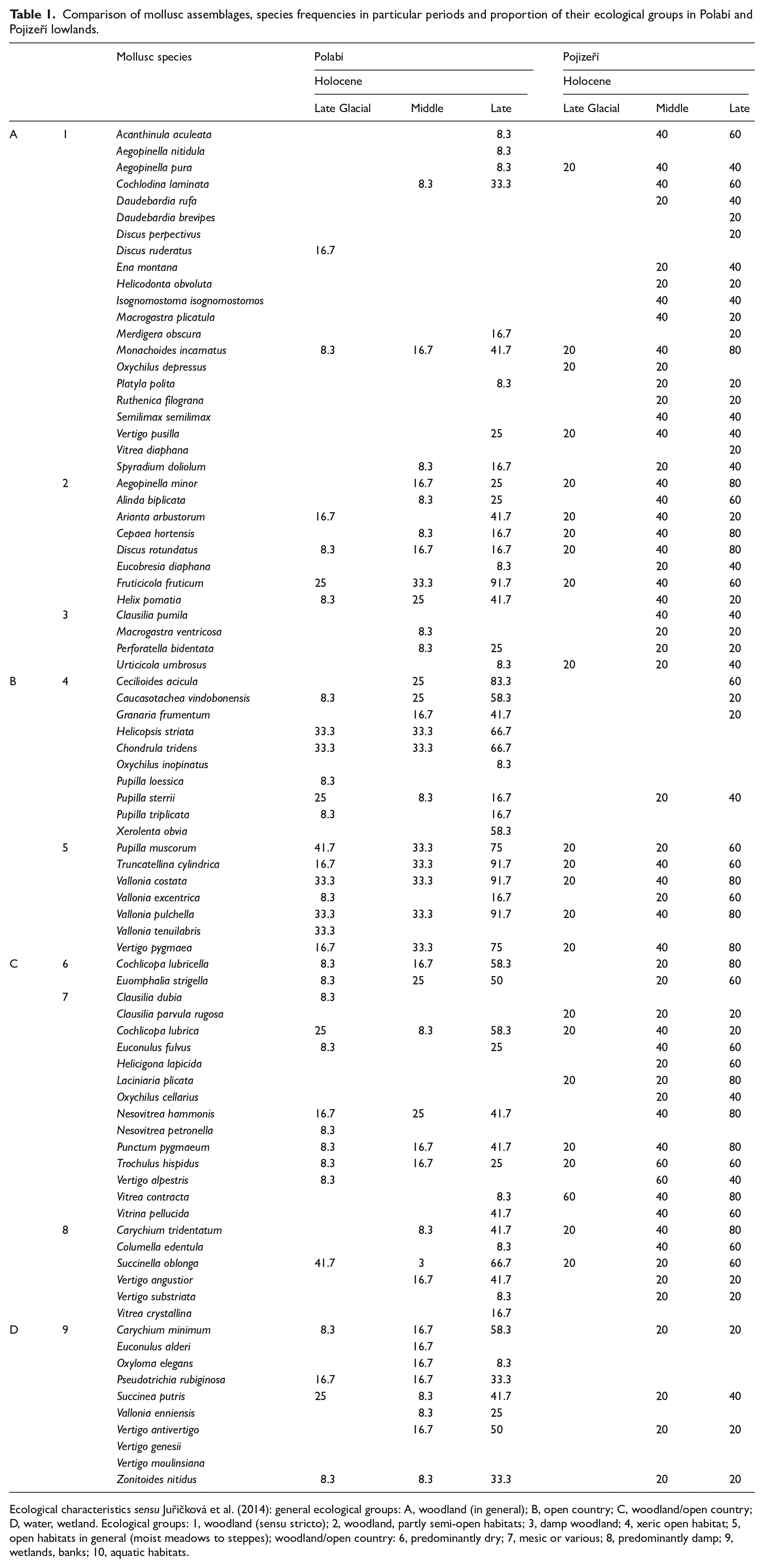
Ecological characteristics sensu Juřičková et al. (2014): general ecological groups: A, woodland (in general); B, open country; C, woodland/open country; D, water, wetland. Ecological groups: 1, woodland (sensu stricto); 2, woodland, partly semi-open habitats; 3, damp woodland; 4, xeric open habitat; 5, open habitats in general (moist meadows to steppes); woodland/open country: 6, predominantly dry; 7, mesic or various; 8, predominantly damp; 9, wetlands, banks; 10, aquatic habitats.
If we look in more detail at the representation of individual ecological groups in both areas (Table 1), we find that the lower frequency of forest species in the Polabí is mainly due to strictly forest species of ecogoup 1, especially epigeic ones. It is possible that the ranking of litter in combination with frequent floods are the main causes of this impoverishment. We do not see a difference in frequencies of the woodland/semi-opened species between both areas, but the moisture-loving forest species clearly predominate in the Pojizeří region. In Table 1 differences are shown also in the frequencies of open-country species. While undemanding species of open landscape occur more or less equally often in both areas, demanding steppe species are rare in the Pojizeří, while in the Polabí they are common since the Late Glacial. It follows that in both areas open habitats were probably continuously preserved, but only those in the drier Polabí had a steppe character. We see similar differences in wetland species. While generally moisture-loving species are more or less equally represented in both areas, demanding wetland species are much more common in the Polabí. It seems that the higher situated and wetter Pojizeří probably consists of a landscape mosaic of open and wooded habitats of varying humidity throughout the Holocene, but more demanding species did not appear here at all or only sporadically in the Late-Holocene. On the contrary, the Polabí landscape, which is practically destroyed by intensive agriculture today, was a species-rich mosaic of relict steppes and wetlands throughout the Holocene, probably with continuity into the glacial period, while fully developed forest fauna probably never occurred here.
Supplemental Material
sj-pdf-1-hol-10.1177_09596836231200441 – Supplemental material for Holocene transformation of natural steppe into an agricultural landscape in the Polabí and Pojizeří Lowlands, Czech Republic, based on mollusc evidence
Supplemental material, sj-pdf-1-hol-10.1177_09596836231200441 for Holocene transformation of natural steppe into an agricultural landscape in the Polabí and Pojizeří Lowlands, Czech Republic, based on mollusc evidence by Jitka Horáčková, Štěpánka Podroužková and Lucie Juřičková in The Holocene
Supplemental Material
sj-xlsx-2-hol-10.1177_09596836231200441 – Supplemental material for Holocene transformation of natural steppe into an agricultural landscape in the Polabí and Pojizeří Lowlands, Czech Republic, based on mollusc evidence
Supplemental material, sj-xlsx-2-hol-10.1177_09596836231200441 for Holocene transformation of natural steppe into an agricultural landscape in the Polabí and Pojizeří Lowlands, Czech Republic, based on mollusc evidence by Jitka Horáčková, Štěpánka Podroužková and Lucie Juřičková in The Holocene
Footnotes
Acknowledgements
This paper is dedicated to our teacher and one of the founders of Quaternary malacology, Vojen Ložek, who unfortunately passed away before preparing of the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Czech Science Foundation supported the study by grant No. GA23-05132S.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.