
Abstract
Migration is a frequent phenomenon in human history. Previous studies mainly used migration as a simple process to account for any cultural changes observed in migrant communities. Recent studies, however, have recognized that migration is embedded in both environmental and social contexts, thus making it necessary to study the consequence of migration on a case-by-case basis. To better understand the changes associated with migrational processes, this case study investigates the subsistence pattern of a Neolithic site (5300-4700 cal. BP), Liujiazhai, in Northwestern Sichuan Highland by employing microbotanical residue analysis on pottery vessels. Our results on millet phytolith contribute to the overall picture of millet agriculture in Yangshao and Majiayao migrant communities and enrich our understanding of how varying crop patterns in Neolithic western China are likely a consequence of migration. In addition, we suggest that Liujiazhai migrants adapted to the high-altitude environment by utilizing more local wild plant resources. This study shows that, although Yangshao migrants were still connected to their homeland in terms of material culture, relocating to the challenging environment in NW Sichuan requires adaptive strategies that diverged the Yangshao migrants, including Liujiazhai, from their home culture. Hence, this study exemplifies how migration is an agent of change.
Keywords
Introduction
Migration is an essential part of human history. From the dispersal of Neolithic cultures to the diaspora of ethnic communities, migration is such an omnipresent phenomenon in the archeological record Defined as “one-way residential relocation to a different ‘environment’ by at least one individual,” migration is more than the physical movement of people, but it entails a profound change evident in biology, socio-culture, and linguistics (Cabana and Clark, 2011). Thus, many have argued for a more rigorous framework to explore the explanatory power of migration and better understand cultural change as its consequence (Adams et al., 1978; Anthony, 1990, 1997; Burmeister, 2000; Champion, 1990). Studies since then have not only recognized that migration is an inherent social strategy (Harper et al., 2019; Hofmann, 2020; Leppard, 2014), but have also found that the impact of migration is multifold – in many cases, migration leads to adaptive changes in subsistence, agriculture, and technologies, and stimulates a stronger expression of identity, whereas in other cases it produces little impact (Clark, 2011; Conolly et al., 2008; Cruz Berrocal, 2012; Ivanova et al., 2018; Kreuz et al., 2005; Mazzucco et al., 2020). Burmeister (2000) proposed that migrants often form a new cultural identity that is distinct of both the home culture and the local culture. Nevertheless, Silliman (2005) cautioned that archeological studies shouldn’t take material culture as the direct representation of cultural identity, adding complexity to migration studies. Without doubt, migration is an agent of change, but the scope and extent of change is context specific. This thinking has been adopted by many postprocessual archeologists who argue for a case-by-case examination on migration because any changes in migrants’ culture are influenced by specific social and environmental contexts that they encounter during migration (Cabana, 2011).
Despite recent progress on migration studies, van Dommelen (2014) reckoned that there still exists a strong concern with finding the “hard” evidence of large population migration, and that “an archaeological understanding of migration as a ‘multilayered process’ is practically non-existent” (p. 479). To better articulate the consequence and the various dimensions of migration, we present a case study of a Neolithic site, Liujiazhai (5300–4700 cal. BP), located in the Northwest Sichuan highland in China. By employing microbotanical residue analysis on pottery vessels, we investigate how agriculture intersected with new landscape and led to changes in crop pattern and adaptations in subsistence strategies. Such study showcases how migrant society evolved in a new environmental setting as they moved further away from their homeland, also adding more information to our understanding of human-environment interaction in the Tibetan Plateau.
Archeological background
The Northwest Sichuan Highland is situated on the eastern margin of the Tibetan Plateau. The initial peopling of this extreme environment has received much attention as to understand the capacity of human adaption as well as the history of the Sino-Tibetan family (Aldenderfer, 2011; d’Alpoim Guedes and Aldenderfer, 2020; Lu, 2023). Upper Paleolithic sites (Piluo, Fulin, Shizishan, Yanyundong, etc.) found in western Sichuan show that a foraging population had already existed before the Neolithic (Shi, 2004; Sichuan Provincial Institute of Cultural Relics and Archaeology, 2021a; Sichuan Provincial Institute of Cultural Relics and Archaeology and School of Archaeology and Museology in Peking University, 2022). However, demography in western Sichuan during the Neolithic is much more complex. Both phylogenic and linguistic studies suggest that the modern-day Tibetans are rather a mix of a local population and a migrant population, possibly the Yangshao culture from the Yellow River valley, dating to 7800–4200 BP (He et al., 2021; Ning et al., 2020; Zhang et al., 2018; Zhao et al., 2009). In addition, genetic studies have found that low-altitude populations (the migrants) acquired adaptive alleles from the local highlanders, attesting to potential interactions between the two groups (He and Ding, 2013; Jeong et al., 2014).
The interactions between farming communities and the local foraging population still require more research, and the geographical location of NW Sichuan Highland offers a great opportunity to conduct such an investigation. So far, many middle and late Neolithic sites have been found in NW Sichuan. Among those sites, the Liujiazhai site (AMS date 5300–4700 cal. BP) is located in the upper reaches of the Dadu River, at an elevation of approximately 2630 m above sea level (Figure 1) (Sichuan Provincial Institute of Cultural Relics and Archaeology, 2021b; Sichuan Institute herein). The macrobotantical record at Liujiazhai consists mostly of broomcorn millet (Panicum miliaceum) and foxtail millet (Setaria italica), as well as chenopods and Asteraceae seeds (Sichuan Provincial Institute of Cultural Relics and Archaeology, 2022). While domesticated plants are present, animal remains only have a limited number of pig bones (wild or domesticated unclear) and are mainly from wild animals (Sichuan Institute, 2022).
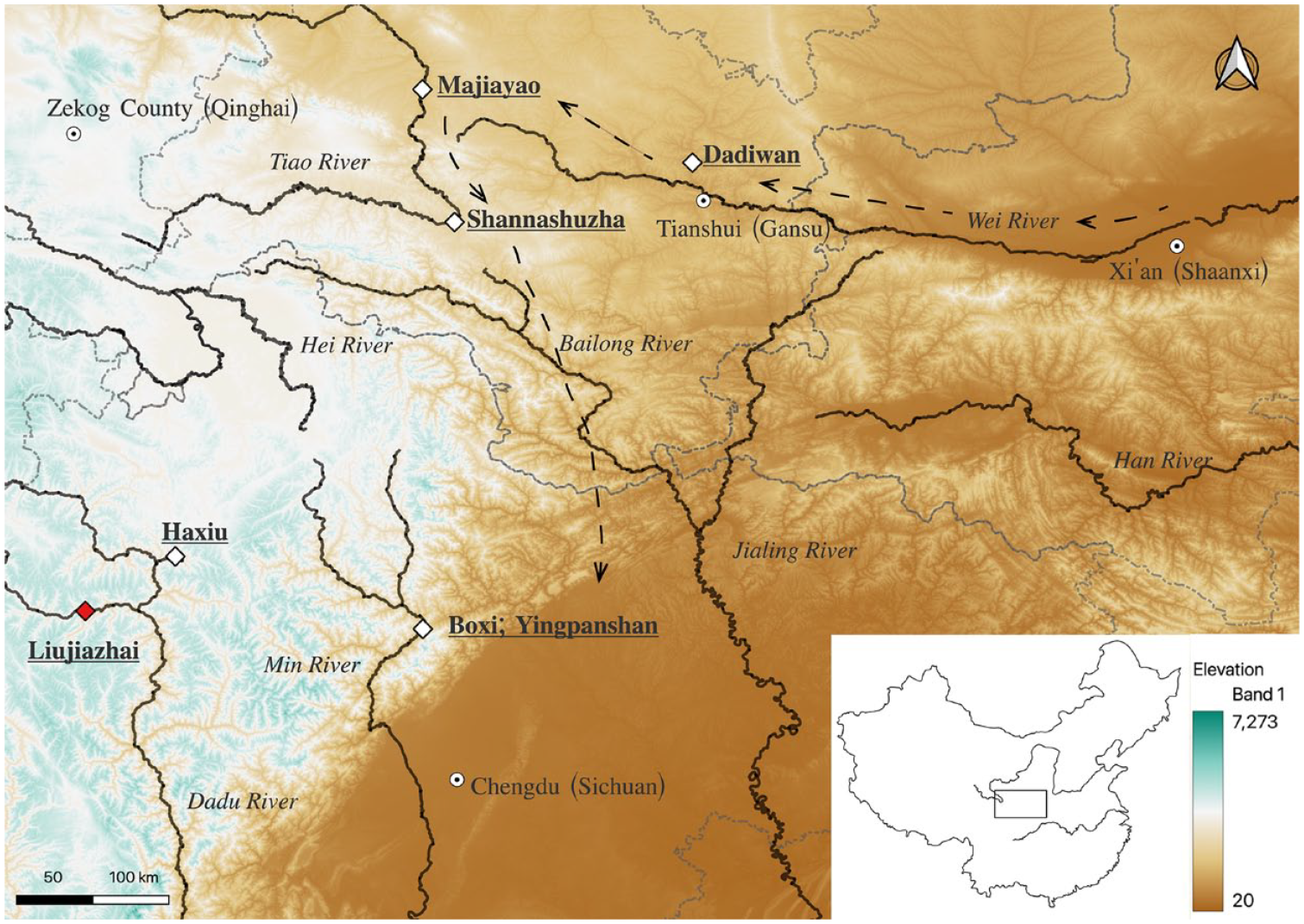
Map showing the locations of sites mentioned in this study and major rivers. Gray dashed lines mark the boundary of Qinghai, Sichuan, Gansu, and Shaanxi provinces. Arrows show possible migration routes suggested by Zhu (2019).
Migration background
It is proposed that the Neolithic migrants in NW Sichuan came from the Yangshao culture that flourished in the Yellow River valley beginning 7000 cal. BP. After broomcorn and foxtail millet were domesticated in northern China around 10,000 and 8000 cal. BP respectively (Lu et al., 2009b), Yangshao culture adopted dryland millet farming since the early phase (7000–6000 cal. BP) and involved millets in their cultural practices such as fermenting millets in amphoras to make alcoholic beverages (Liu, 2021; Liu et al, 2020). Red and painted pottery are distinct characteristics of Yangshao pottery style, and it became a symbol of cultural identity when Yangshao communities migrated out of the Yellow River valley. Starting 6000 cal. BP, the Yangshao migrants moved westward, settling in where is now Gansu and Qinghai, and formed new cultural variants such as the Shilingxia transitional phrase (5900–5200 cal. BP) and the Majiayao culture (5300–4500 cal. BP) (Ding 2010). Meanwhile, another wave of migration saw the emergence of Yangshao cultural sites in NW Sichuan (Figure 1), including Boxi and Yingpanshan (5300–4600 cal. BP) in the upper reaches of Min river, and Haxiu (5500–4700 cal. BP) and Liujiazhai (5300–4700 cal. BP) in the upper reaches of Dadu River (He, 2015; Jiang and Chen, 2003; Zhao and Chen, 2011). Though the radiocarbon date is not available for the Boxi site, it is dated to the middle Yangshao period, or the Miaodigou phase (6000– 4700 BP), based on its material culture (Jiang and Chen, 2003). Painted pottery and amphoras are ubiquitous in the material assemblages from these sites, although the majority are not painted pottery (He, 2015; Jiang and Chen, 2003; Ren, 2015). The provenance of those painted pottery vessels received much debate as it pertains to the development of pottery traditions, and potentially cultural identity, of the Yangshao and Majiayao migrants. Chemical analysis suggests that painted potteries in NW Sichuan were produced in Gansu and Qinghai, possibly acquired via trade, whereas the unpainted ones were made locally (Cui et al., 2011; Hung et al., 2010). Chen (2009), Ren et al. (2013), and Ren and Chen (2022), on the other hand, argued that vessel forms and decoration styles of some painted pottery from NW Sichuan are not found in Gansu and Qinghai, thus a local production of painted pottery remains likely. While the origin of painted pottery is open to future research, a cultural tie between the Yangshao/Majiayao culture and the NW Sichuan sites is still evident. Both macrobotanical record and starch residues show that the Yangshao/Majiayao migrants at Boxi, Haxiu, and Yingpanshan continued practicing millet cultivation (Liu et al., 2022; Sichuan Institute, 2022; Zhao and Chen, 2011). In addition, residue analyses on pottery samples from Boxi and Haxiu suggest they retained cultural traditions such as alcoholic fermentation and drinking rituals (Liu et al., 2022).
Environmental background in NW Sichuan
The Holocene amelioration provided foraging communities with the conditions to transition toward sedentism and agriculture. However, periodic climatic variations still occurred in the Holocene. A humid and warm climate from the early Holocene allowed forests to emerge and dominate the landscape in NW Sichuan. This trend continued until the Mid-Holocene optimum ended around 6000 cal. BP (An et al., 2004; Ran and Feng, 2013). According to paleoclimatic reconstruction, the Mid-Holocene precipitation in NW Sichuan was about 500–800 mm, and the average temperature was 2°C higher than today, categorizing a semi-arid environment (Wen et al., 2017; Zhou et al., 2021). Beginning in 5500–5300 cal. BP, the East Asia Summer Monsoon weakened, causing a series of hydrological recessions and declined seasonal precipitation in both Yellow River Valley and western China (An et al., 2004; Chen et al., 2015a; Huang et al., 2019; Jarvis, 1993; Zhang et al., 2023). This cold and dry period persisted until 4800 cal. BP when the climate became wetter again (Wen et al., 2017). The warm and wet Mid-Holocene period supported the Yangshao culture to expand, which is evidenced by the increasing settlement size (Gong, 2003). As a result, population growth and the subsequent population-resource imbalance could have played a role in motivating migration (Wu et al., 2018).
Materials and method
Field sampling
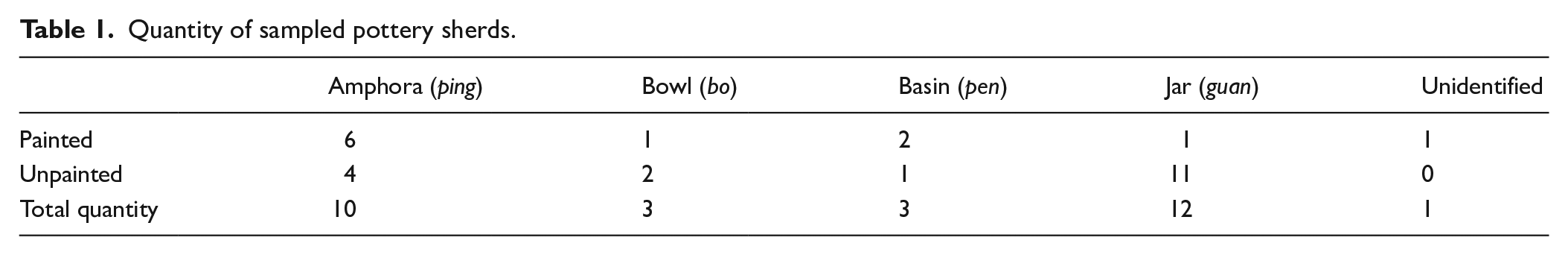
The field sampling took place at Sanxingdui Workstation in Sichuan, 2019, where all artifacts for this study were curated and stored (Figures 2 and 3). The vessel types and decoration styles of the 29 sherd samples are listed in Table 1 (full information of sampled sherds is in Supplemental Table S1, available online). We sampled 10 amphora sherds to determine if they had previously contained fermented beverages as it is a common practice in the Yangshao culture. To prevent contamination, we used new and sterile plastic bags, test tubes, pipettes, toothbrushes, and razor blades during the sampling process. First, all pottery samples were brushed and washed in running water to remove surface dust and particles. For small pottery sherds, we sonicated them in an ultrasonic bath, while larger sherds were brushed using an ultrasonic toothbrush. We placed each small sherd in a new polyvinyl bag with approximately 15 mL of distilled water and then placed each bag into the ultrasonic bath for 6 min. For larger sherds, we gently brushed the interior surface using an ultrasonic toothbrush while simultaneously adding distilled water using a pipette to rinse the surface. We used a new ultrasonic toothbrush and a new pipette for each sherd. After the ultrasonic bath or toothbrushing, each residue liquid sample was transferred to a new 15 mL test tube and then stored in a sealed plastic bag before laboratory analysis. In addition, we collected sediment samples from the exterior surfaces of Pot 1, Pot 5, Pot 10, and Pot 19 by using a clean razor blade, which served as control samples to compare with the residue samples obtained from the interior surface of these pots.

Examples of sampled pottery sherds. (a) Pot 25, bottom of an unpainted basin. (b) Pot 4, rim of a painted amphora. (c) Pot 22, bottom of an unpainted jar with cord mark. (d) Pot 10, rim of an unpainted amphora. (e) Pot 9: body sherd of a painted bowl. (f) Pot 8, bottom of a painted basin. (g) Pot 24, rim of an unpainted amphora. (h) Pot 13, rim of an unpainted jar with cord mark. (i) Pot 19, bottom of an unpainted jar with cord mark.

Examples of pottery types at Liujiazhai. (a) Jar with applique (2011TSE12:26). (b) Amphora (2011H116:107). (c) Basin (2011H116:106). (d) Bowl (2012 G1:34). (a–c) are from the 2011 Excavation Report (Sichuan Provincial Institute of Cultural Relics and Archaeology, 2021b) and (d) is from the 2012 Excavation Report (Sichuan Provincial Institute of Cultural Relics and Archaeology, 2022).
Quantity of sampled pottery sherds.
Laboratory processing
The processing method followed the established method, as detailed in the Supporting Information, available online.
Starch and phytolith identification
All starch granules were counted and recorded. For phytolith, the entire slide is counted only when phytolith is fewer than 300. We identified starch and phytolith taxa by comparing them to our reference collection of more than 1000 economically important plant specimens from Asia and other published literature (Ge et al., 2018; Huan et al., 2015; Lu et al., 2009a; Zhang et al., 2011). Starch granules with cooking or heating damages were identified by comparing with the published literature (Babot, 2003; Henry et al., 2009; Wang et al., 2016). We followed the International Code for Phytolith Nomenclature 2.0 to describe phytolith morphotypes (Neumann et al., 2019).
Results
Starch residue analysis
In total, we found 351 starch granules, consisting of 272 identified starch granules and 79 unidentified ones (UNID), from 26 out of the 29 samples (Supplemental Table S2, available online; Figure 4). The identified starches (77.5% of the total count) are divided into eight types and detailed in the following (for modern starch reference and identification, see Supplemental Figure S1, available online). The UNID starch granules (22.5% of the total count) consist of gelatinized starch (n = 11), starches with unidentifiable damages (n = 19), unidentified starch (n = 44), and unidentified compound starches (n = 5). It is known that starch granules are subject to morphological alterations either caused by cooking or enzymatic damages. Gelatinization is the result of the heating process such as cooking, and it is characterized by swelling, distortion, and partial or complete loss of extinction cross (Williams et al., 1982). On the other hand, starches with unidentifiable damages show signs of cracks and pitting that result from various food processing and taphonomic processes (Takaya et al., 1978).
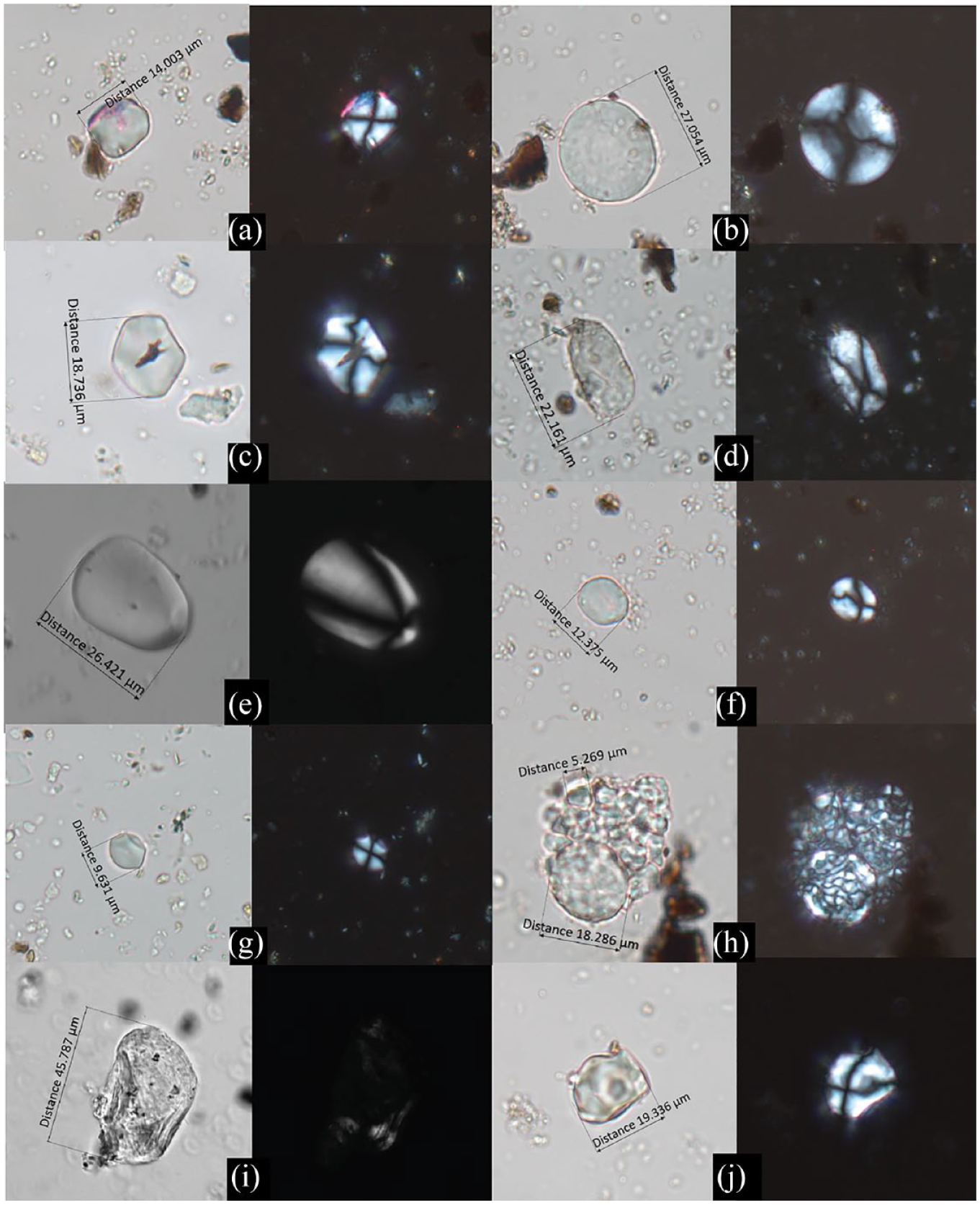
Types of starch granules identified in this study. (a) Panicoideae. (b) Triticeae. (c) Job’s tears (Coix lacryma-jobi). (d) Unidentifiable USO. (e) Zingiberaceae (under DIC view). (f) Fern root. (g) Pueraria sp. (kudzu vine root). (h) Euryale ferox (foxnut). (i) Gelatinized. (j) UNID.
Type 1: Panicoideae, n = 108, size range: 5
69–16.839 μm; 30.8% of the total count; ubiquity = 0.66.
Type 1 starch granules are mostly spherical or polyhedral in shape, with hila located in the center and straight arms of extinction cross. Foxtail millet (Setaria italica) and broomcorn millet (Panicum miliaceum) most likely contribute to this type, with sizes ranging from 11.21 to 16.75 μm and 5.92 to 13 μm in our reference collection, respectively. The majority of Panicoideae starches (n = 65) are found in jars, with some are found in amphoras (n = 18) and bowls and basins (n = 18). Most of Type 1 starches (n = 83) are found in pure-clay cooking (jars) and serving vessels (bowls and basins), thus it is mainly associated with cooking and serving activities. Type 1 is the most abundant and ubiquitous type, which is consistent with the macrobotanical record of Liujizhai and millet cultivation in Yangshao culture.
Type 2: Triticeae, n = 69, size range: 17
704–31.3 μm; 19.7% of the total count; ubiquity = 0.62
Type 2 granules are lenticular or discoid in shape with a flat surface, with centric hila and visible lamella on some large granules, consistent with the Triticeae tribe. When the flat side of the granule is under observation, the extinction cross takes on a “+” shape, but when observed from the side, it becomes more like an “x.” The size of modern Triticeae granules can vary from 2.46 μm to larger than 20 μm. Agropyron, Elymus, and Leymus, which are commonly found in arid regions in China (Chen et al., 2013), are potential candidates for this type. Another possible source is Secale sp. (rye), as charred Secale sp. has been discovered at Yingpanshan (Zhao and Chen, 2011). However, due to the lack of more diagnostic features, the starch granules grouped here are referred to as Triticeae. The majority of Triticeae granules (n = 35) are found in jars with Panicoideae, while some (n = 18) are found in bowls and basins.
Type 3: Job’s tears (Coix lacryma-jobi), n = 47, size range: 17
759–25.322 μm; 13.4% of the total count; ubiquity = 0.48
Type 3 granules are spherical or polyhedral in shape, which are similar to Type 1 but show several distinctive features based on Liu et al. (2014)’s criteria for Job’s tears starch, which includes three features: (1) larger size, (2) higher eccentricity ratio of hilum, and (3) z-shape curved arms of extinction cross. Job’s tears typically grow in wetlands, distinguishing them from millets, which grow in different habitats. Although the history of Job’s tears cultivation or domestication remains unclear, its exploitation is evident in many nearby sites, including Haxiu, Boxi, Dadiwan, and the late Neolithic Baodun site in Sichuan (Guedes et al., 2013; Liu et al., 2022).
Type 4: Unidentifiable underground storage organs (USO), n = 39, size range: 8
415–15.45 μm; 11.4% of the total count; ubiquity = 0.48
Type 4 granules are round or elongate oval in shape with eccentric hila and bent arms of extinction cross, which are consistent with starches from USOs. USOs include tubers, roots, rhizomes, and bulbs, and their starch granules exhibit similar morphology but lack more diagnostic features for further identification. The relatively high ubiquity of USOs makes them an important food source at Liujiazhai, in addition to the previous three types.
Type 5: Zingiberaceae, n = 2, size range: 27
19–33.53 μm; 0.6% of the total count; ubiquity = 0.07.
Type 5 starch granules have elongate oval or nearly triangular shape, and it is generally larger than many other types. The hila of extinction cross is highly eccentric and is located extremely close to the edge. They show clear relief under the differential interference contrast (DIC) view, which is similar to the criterion set by Torrence and Barton (2006: Plate 27). In our modern reference collection, young ginger roots (Zingiber sp.) tend to have fan-shaped granules whereas mature ones have elongate granules. The starch granules grouped here generally match the description of ginger roots. The exploitation of ginger roots is evident at many sites in the Wei River valley (Liu, 2021; Wang et al., 2019), and it has long been used as medicine and food ingredients in China (Wang and Wang, 2005).
Type 6: Fern root, n = 2, size range: 11
545–30.071 μm; 0.6% of the total count; ubiquity = 0.07.
Type 6 starch granules exhibit various shapes, including round, semi-spherical, elongate oval, and facetted. The hila can be both centric and eccentric, and the extinction cross sometimes has bent or zig-zag arms. This type shows a high resemblance to the root of the eagle fern (Pteridium aquilinum) in our modern reference collection. Many fern species are distributed in Sichuan today, and their roots and leaves are frequently used as food and in medicine (Li, 1994). In addition, eagle fern is highly rich in proteins, carbohydrates, fats, and minerals (Qi et al., 2015). Thus ferns are great options for exploitation.
Type 7: Pueraria sp
(kudzu vine root), n = 2, size range: 9.631–15.105 μm; 0.6% of the total count; ubiquity = 0.07.
Type 7 starch granules are mostly facetted, with some sub-round or oval in shape. The hila is usually centric, and the extinction cross sometimes has uneven thickness with thicker arms toward the peripheries. This type is consistent with the morphological characteristics of kudzu vine granules in our modern reference collection. Kudzu produces edible starchy roots, and its fibrous vines have been used to make textiles and paper in ancient China. It is also a traditional medicine to treat fever and diarrhea (Wong et al., 2011).
Type 8: Euryale ferox (foxnut), n = 2, size range: 2
723–5.29 μm; 0.6% of the total count; ubiquity = 0.03.
Type 8 starch granules usually appear in a compound structure with a round or oval shape, while the individual starch granules are more polygonal. The hilum of the individual granule is mostly centric. This type is most consistent with the starch granules found in the roots of foxnut (Euryale ferox) in our modern reference. Foxnut is a wetland species that grows in ponds and lakes. It has also been identified at Boxi although the abundance and ubiquity are higher there (Liu et al., 2022, Supporting Information, available online).
In addition, the residue samples also revealed 11 gelatinized starch granules, most of which (n = 8) were from pure-clay vessels (jars, bowls, and basins) that were likely used for serving food. To conclude, cereals from Type 1 Panicoideae and Type 2 Triticeae account for most of the starch remains in Liujiazhai. In addition to these upland crops and grasses, wetland resources such as Job’s tears were also utilized, and to a lesser extent, foxnuts. USOs and other less ubiquitous wild plants such as fern roots and kudzu indicate reliance on local resources other than millets. The similarities of the starch remains found in Liujiazhai, Haxiu, and Boxi imply the exploitation of a similar set of local resources (Table 2). It is worth pointing out that Liujiazhai doesn’t have rice remains, unlike the other two sites. Whether this discrepancy implies different cultivation or subsistence strategies, or contrasting cultural practices, calls for further research.
Comparing starch residues found in the Liujiazhai, Haxiu, and Boxi sites.
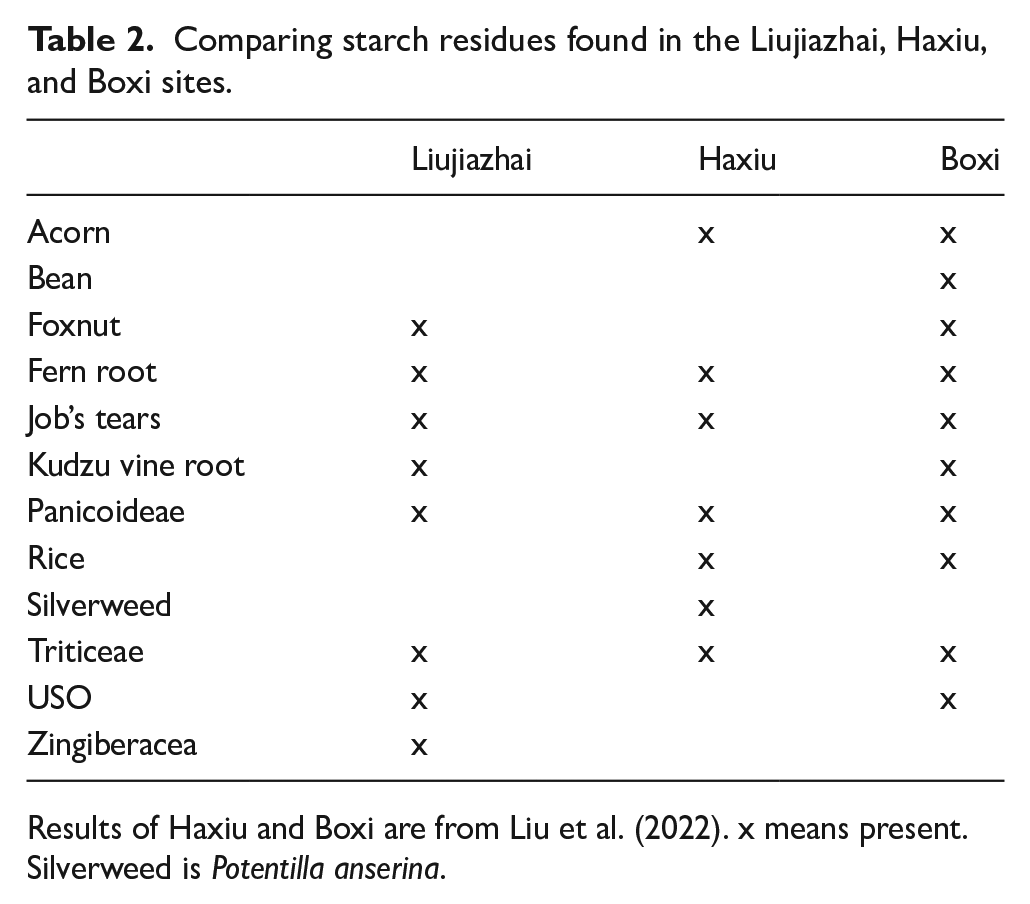
Results of Haxiu and Boxi are from Liu et al. (2022). x means present. Silverweed is Potentilla anserina.
Phytolith results
We identified a total of 640 phytoliths, including 167 multicell and 473 single cell phytoliths (Supplemental Table S3, available online; Figure 5). Among the multicell phytoliths, 31 were identified as
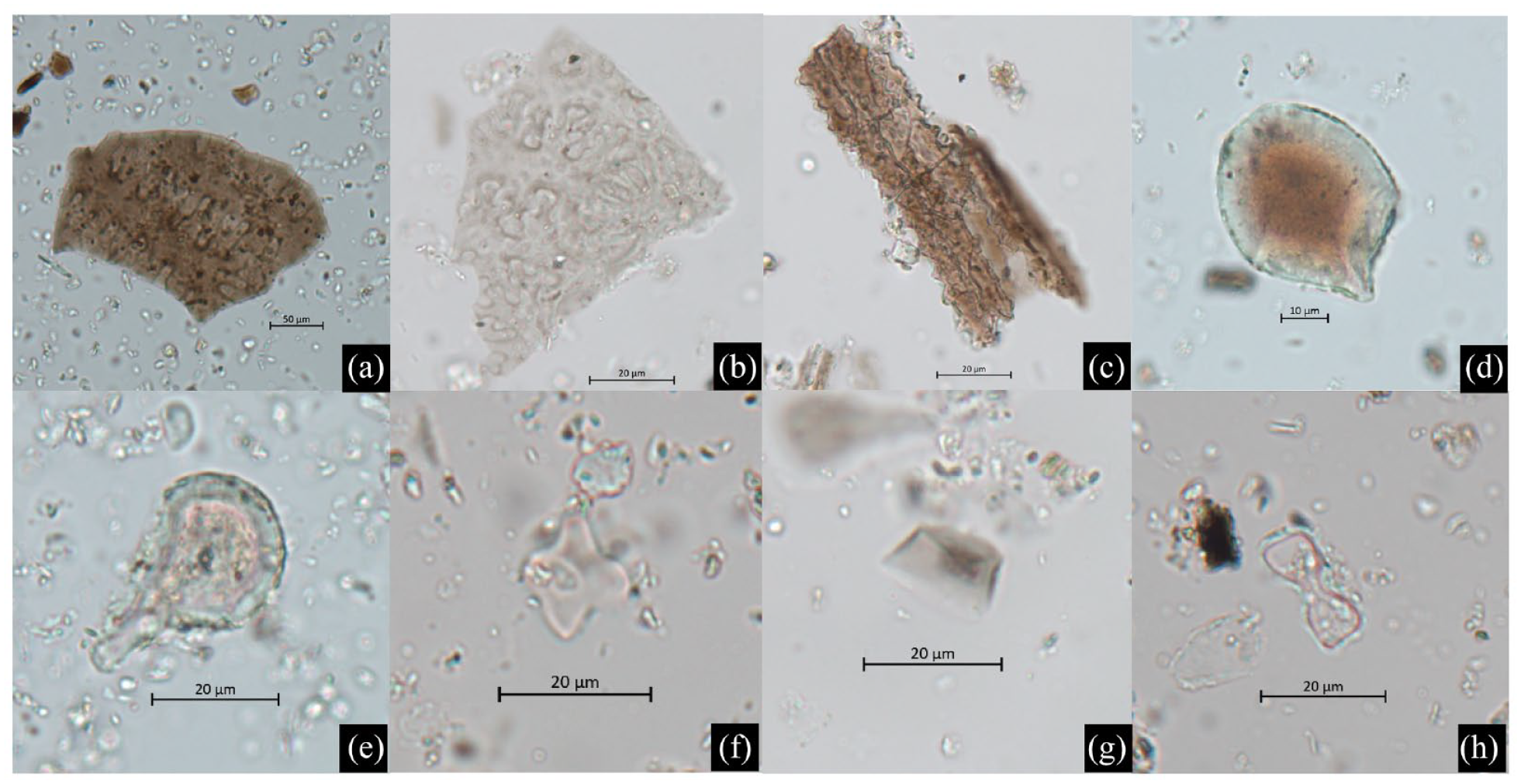
(a)
The most abundant single cell phytolith was rondel (n = 218) from Poaceae, with a significant presence in Pot 17 (n = 186) and low numbers in other samples. Hair cell was the second most abundant form (n = 114), commonly found in plants but not indicative of taxa. We also found phytolith types indicative of Panicoideae plants such as bilobate (n = 40), polylobate (n = 1), and cross (n = 10), which were consistent with the starch remains and suggesting that millets were the source of these phytoliths. In addition, Phragmites bulliform (n = 7) was also found, suggesting a minor presence of wetland resources. Other morphotypes include common bulliform (Poaceae; n = 34), trapeziform (likely Pooideae; n = 4), and elongates (grass husk, inflorescence, or leaves; n = 15).
Overall, the phytolith remains all come from Poaceae, including millets and other wild grasses. They are consistent with the starch remains and suggest additional use of wetland plants such as Phragmites. One difference between Liujiazhai and other sites is that
Control samples
We analyzed four control samples, which were extracted from the sediments adhering to the exterior surfaces of sherds. These samples showed an absence of starch granules, and only two samples revealed a few phytolith. Pot 5 control had three phytolith short cells (common bulliform, trapeziform, and Elongate irregular) and Pot 10 control had only one common bulliform. We determined that the minor presence of these phytoliths in the control samples suggests that the residue samples from the interior surfaces are associated with culinary practices rather than environmental contamination.
Discussion
Overall subsistence, millet agricultural, and local adaptation
Both starch and phytolith remains suggest that millets and local resources (such as fern roots and other USOs) characterize the subsistence pattern at Liujiazhai. The macrobotanical record of Liujiazhai shows exploitation of Chenopodium sp. and Brassica sp., as well as other weeds such as Digitaria sp. and Setaria sp. Thus, the overall subsistence at Liujiazhai has two main components: (1) millet cultivation, which is practiced in all Yangshao and Majiayao sites and (2) foraging local herbaceous plants and starchy tubers as a means of adapting to a new environment.
Both foxtail millet and broomcorn millet played significant roles as crops in the Yangshao culture of the Central Plain. While broomcorn millet was more dominant in the early periods, a shift in the crop pattern occurred around 5500 BP. Foxtail millet began to increase in quantity and eventually replaced broomcorn millet as the dominant crop (Bestel et al., 2018; Chen et al., 2020; He et al., 2022; Jia et al., 2013; Wang et al., 2018). This change can be attributed to three factors: population growth, environmental change, and the physiological characteristics of millets. Population likely expanded in the late Yangshao period (around 5500–5000 BP) as settlement size and density all increased compared to previous periods (Bi et al., 2013; Gong, 2003). Meanwhile, the weakened East Asia Summer Monsoon by 5500 BP resulted in reduced hydrological conditions and created an arid period (Chen et al., 2015a; Huang et al., 2000). The combined effect of population growth and declined hydrology likely motivated Yangshao communities to increase crop yields. Foxtail millet, as experimental studies have shown, is more productive and suffers less yield loss under water stress (Matsuura et al., 2012; Nematpour et al., 2019; Tadele, 2016). Therefore, the higher productivity of foxtail millet likely led to its intensification during a time of population growth and declining precipitation (Yang et al., 2022; Zhong et al., 2020).
However, millet farming among Yangshao migrants in western China show divergent patterns. In contrast to the general trend of increasing foxtail millet dominance in the Yangshao homeland, crop patterns in NW Sichuan, Gansu, and Qinghai show more local variations. On one hand, foxtail millet is more abundant than broomcorn millet at Yingpanshan, Shannashuzha, Dadiwan (Phase IV, Late Yangshao), and sites in the Northeastern Tibetan Plateau, as suggested by the macrobotanical record (Chen et al., 2015b; Gansu Provincial Institute of Cultural Relics and Archaeology, 2006; Hu, 2015; Zhao and Chen, 2011). On the other hand, broomcorn millet still outnumbers foxtail millet in the macrobotanical remains from the Lijiatai site (ca. 5400 cal. BP) and the Andaqiha site (ca. 5300 cal. BP) in Qinghai (Jia, 2012). Similarly, phytolith residue analysis on Haxiu, Boxi, and Dadiwan potteries (Phase III and IV, Middle to Late Yangshao) indicates that broomcorn millet is more abundant and more ubiquitous (Liu et al., 2022; Zhao and Liu, 2021). Our own phytolith analysis found an equal contribution of broomcorn (
This variation is likely resulted from two factors: local conditions at each migrant site and the physiological differences between broomcorn and foxtail millet. Firstly, as migration involves relocating to different environments, Yangshao migrants would counter varying local conditions that are significantly different from their homeland when they moved west and southwards to Gansu, Qinghai, and NW Sichuan. Elevation increases from Gansu to Qinghai and NW Sichuan as the latter two are located at the edges of the Tibetan Plateau (see Figure 1). Consequently, precipitation decreases with higher elevation. Moreover, numerous mountain ranges and basins in these three areas led to local microclimates (i.e. thermohydrology) and multiple vegetation types (i.e. dessert-steppe, forest, scrub, and meadow) (Chen et al., 1994; Li and Yang, 1989; Peng et al., 1989). Secondly, although foxtail millet has higher yields, broomcorn millet requires fewer nutrients and has a shorter growth cycle (Hunt et al., 2011). Thus, migrant communities could have developed their own crop patterns based on the suitability of each millet type to their specific local conditions. It is important to note that migration acts as an agent of change. When a small population migrants out of the home population, they adapt to the local conditions of their new settlement by altering subsistence patterns according to the adaptivity of their domesticates (see Ivanova et al., 2018; Kreuz et al., 2005). Therefore, the observed variation in crop patterns among Yangshao migrant communities, including Liujiazhai, shows how millet agriculture became diversified as a result of migration.
Furthermore, we propose that Liujiazhai migrants heavily exploited local wild resources, including plant roots (Starch Type 4–7) and wild Triticeae grasses, due to risky agricultural production and relative lack of food resources. d’Alpoim Guedes et al. (2015) argued that millet cultivation, especially broomcorn millet, would be extremely challenging in the Tibetan Plateau based on Growing Degree Days (GDD) modeling. Besides, the estimated Mid-Holocene mean annual precipitation (MAT) in the NW Sichuan highland is 500–800 mm, similar to the present-day MAT of 600–800 mm (Li and Yang, 1989; Zhou et al., 2011, 2021). This level of precipitation would not be ideal for millet cultivation since millet agriculture is nearly absent in most places in NW Sichuan today (Li and Yang, 1989). Thus, crop yields were likely very low. In addition to the difficulties in agricultural production, Liujiazhai had fewer resources to exploit compared to nearby sites. Macrobotanical analysis at the nearby Yingpanshan site reported a greater variety of plant taxa, including rye (Secale sp.), shiso mint (Perilla frutescens), wild grape (Vitis sp.), and woody and shrub species like Chinese Yew (Taxus chinensis), Chinese sumac (Rhus chinensis), peach (Prunus persica), and seaberry (Hippophae ehamnoides) (Zhao and Chen, 2011). In addition, faunal analysis of Yingpanshan and Liujiazhai shows that Yingpanshan consumed more domesticated pigs whereas Liujiazhai migrants relied more on wild animals (Luo et al., 2018). These differences in accessing plant and animal resources, coupled with low crop yields, suggest a challenging food environment in the mountainous highlands where Liujiazhai migrants exploited more wild resources to supplement their subsistence. Since the migrants likely interacted with the local highlanders as supported by genetic evidence (He and Ding, 2013; Jeong et al., 2014), we suggest that Liujiazhai migrants, like other migrant communities at Boxi and Haxiu (see Liu et al., 2022), integrated with the indigenous groups and embraced their subsistence practices with local plants and animals.
Another issue we need to address here is the absence of fermented beverages at Liujiazhai, considering evidence of fermentation has been observed at Haxiu and Boxi sites. This absence of evidence raises questions about potential changes in food-related practices. Liu et al. (2022) inferred that fermentation was practiced at Boxi and Haxiu based on the presence of rice starch granules, yeast, and the Monsacus (a type of fungi) qu starter, all of which are ingredients commonly used in fermentation. However, our study did not find such evidence in the 10 amphora sherds we sampled. It is possible that the absence of rice and Monascus may be due to insufficient sampling rather than a lack of fermented beverages at Liujiazhai. To draw conclusions on the continuity of this cultural tradition, more research on Neolithic NW Sichuan sites is required. The results of the current study have not yet reached a finalized statement about the practice of fermentation by the migrant communities when they settled in NW Sichuan.
Conclusion
In this study, starch and phytolith analyses were employed to investigate the subsistence economy of Liujiazhai migrants in NW Sichuan and explore the consequences of migration. Our findings indicate that the Majiayao migrants in Liujiazhai continued millet cultivation, similar to the Yangshao culture. Additionally, we propose that the Liujiazhai migrants adopted a different subsistence economy compared to their home culture, as evidenced by divergent crop patterns and the inclusion of local wild plants. These findings suggest active adaptation to the challenging landscape in NW Sichuan and potential integration with the local foraging population. Thus, our study exemplifies how migrating to a different microenvironment can lead to changes in agriculture and subsistence patterns.
While this study primarily focuses on subsistence, we acknowledge the need for further research to explore the cultural identity of migrant communities in NW Sichuan. As discussed earlier, material culture alone cannot serve as a direct representation of cultural identity. Indeed, our study reveals that the Liujiazhai migrants adopted a subsistence strategy diverged that from the Yangshao/Majiayao culture, despite their use of Majiayao-style pottery. Therefore, additional lines of evidence are necessary to determine the cultural identity of Liujiazhai and other migrant communities in NW Sichuan. This comprehensive approach will not only enhance our understanding of Sino-Tibetan history but also provide insights into the multifaceted dimensions of migration.
Supplemental Material
sj-docx-1-hol-10.1177_09596836231200438 – Supplemental material for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP)
Supplemental material, sj-docx-1-hol-10.1177_09596836231200438 for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP) by Yiyi Tang, Jiajing Wang, Li Liu and Wei Chen in The Holocene
Supplemental Material
sj-jpg-5-hol-10.1177_09596836231200438 – Supplemental material for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP)
Supplemental material, sj-jpg-5-hol-10.1177_09596836231200438 for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP) by Yiyi Tang, Jiajing Wang, Li Liu and Wei Chen in The Holocene
Supplemental Material
sj-xlsx-2-hol-10.1177_09596836231200438 – Supplemental material for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP)
Supplemental material, sj-xlsx-2-hol-10.1177_09596836231200438 for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP) by Yiyi Tang, Jiajing Wang, Li Liu and Wei Chen in The Holocene
Supplemental Material
sj-xlsx-3-hol-10.1177_09596836231200438 – Supplemental material for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP)
Supplemental material, sj-xlsx-3-hol-10.1177_09596836231200438 for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP) by Yiyi Tang, Jiajing Wang, Li Liu and Wei Chen in The Holocene
Supplemental Material
sj-xlsx-4-hol-10.1177_09596836231200438 – Supplemental material for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP)
Supplemental material, sj-xlsx-4-hol-10.1177_09596836231200438 for Local adaptation and subsistence strategy of Yangshao migrants in Northwestern Sichuan in China during the Middle Neolithic (5300–4700 cal. BP) by Yiyi Tang, Jiajing Wang, Li Liu and Wei Chen in The Holocene
Footnotes
Acknowledgements
We thank Yinzhi Cui from Northwest University (China) for her assistance in sample collection and processing.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work is supported by Burke Research Initiation Award at Dartmouth College and Min Kwaan Chinese Archaeology Fund at Stanford University.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.