
Abstract
Changes in marine productivity of the last five centuries in the Gulf of Tehuantepec were investigated using a high-resolution record of diatoms, organic carbon (Corg), total nitrogen (TN), Ni/Al, and Cu/Al. The laminated sediments were dated by using 210Pb and 14C, with a bayesian age model providing a new ΔR = 247 ± 30 years for the bulk sediment. The Little Ice Age (LIA) (~1500 to ~1858 CE) was characterized by the predominance of cold-water and high productivity diatoms (Chaetoceros spores, Thalassionema nitzschioides, Lioloma pacificum, Thalassiosira nanolineata, and Rhizossolenia setigera) and high values of geochemical productivity proxies. A transition period (~1860 to ~1919 CE) toward warmer conditions related to the end of the LIA and the beginning of the Current Warm Period (CWP), was indicated by the appearance of warm-water diatoms (Neodelphineis pelagica, Thalassiosira tenera, and Rhizossolenia bergonii), as well as lower values of Corg, TN, Ni/Al, and Cu/Al. The most recent period of the CWP (~1920 CE to today) was characterized by the increased abundance warm-water taxa (N. pelagica, Cymatodiscus planetophorus, T. tenera, Plagiogramma minus, Nitzschia interruptestriata, and R. bergonii), and by the prevalence of low values of Corg, TN, Ni/Al, and Cu/Al. These changes in productivity during the LIA and CWP were likely driven by changes in solar irradiance and the migration of the Intertropical Convergence Zone. This study highlights the spatial extent of the LIA in the Eastern Tropical North Pacific and contributes to the knowledge of the productivity response to climate in tropical regions.
Introduction
The climate of the Eastern Tropical North Pacific (ETNP) is primarily modulated by changes in the strength of the trade winds and the latitudinal shifts of the Intertropical Convergence Zone (ITCZ) (Lavín et al., 2006). Also, variations at diverse scales (centennial, decadal and interannual) have impacted the ETNP climate in the last half millenium . A prolonged cold period called the Little Ice Age (LIA ~1250 to ~1850 CE, e.g. Crowley et al. (2008), Miller et al. (2012), ~1400 to ~1700 CE, e.g. Mann et al., 2009) has been related to the lowest solar irradiance in the past millenium (Spörer, Maunder and Dalton solar minima, Bard et al., 2000; Lean, 2018), and increased volcanic activity (Crowley et al., 2008). Although the LIA has been widely studied, its timing and impact on a global scale are still being discussed, mainly in tropical regions (Juárez et al., 2014) and in the ETNP (Barron and Bukry, 2007; Choumiline et al., 2019; Goni et al., 2006; Juárez et al., 2014; Staines-Urías et al., 2009). In paleolimnological studies in central and southern Mexico, the LIA has been associated with temperature decrease of ~2.0°C and mountain glacier advance (Lozano-García Mdel et al., 2007; Vázquez-Selem, 2011). A change to warmer condition is documented worldwide since ~1850 (IPCC, 2014) identified as the Current Warm Period (CWP) (e.g. Griffiths et al., 2016; Salvatteci et al., 2014).
The Gulf of Tehuantepec (GoT) is in the ETNP (Figure 1). Water masses in the GoT are: Equatorial Surface Water (ESW, up to 29°C and salinity >34), Subtropical Subsurface Water (SSW, <25°C and salinity >35), North Pacific Intermediate Water (NPIW, 4°C–9°C and salinity <34.5), and North Deep Pacific Water (NDPW, 1.2°C–2°C, <34.5) (Fiedler and Talley, 2006; Machain–Castillo et al., 2008); and their vertical distribution is influenced by the atmospheric circulation regime.
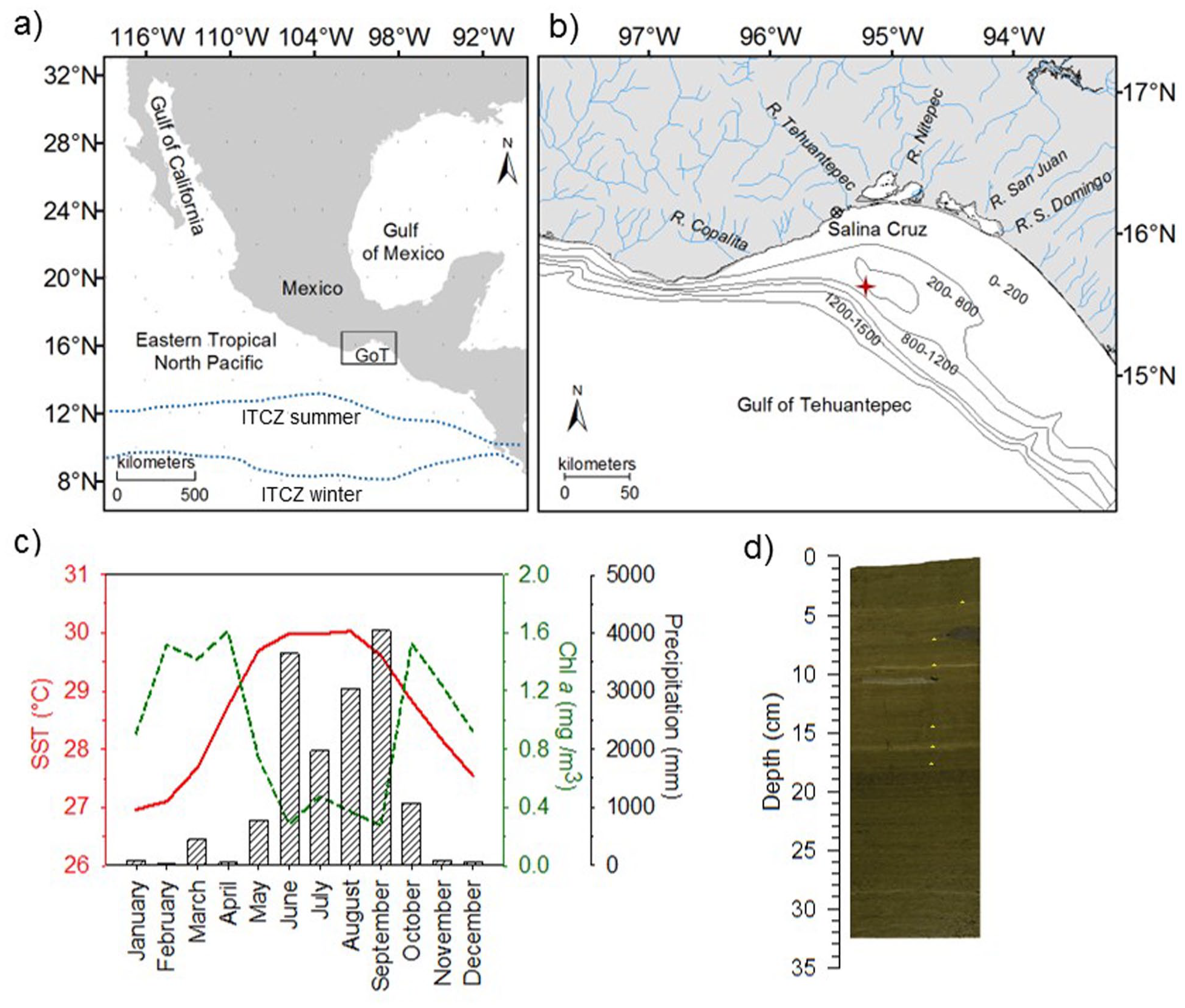
(a) Gulf of Tehuantepec (GoT) location in the Eastern Tropical North Pacific and mean position of the ITCZ in summer and winter (Amador et al., 2006). (b) sampling site of core Tehua XII E03 (red star) and main rivers in the Tehuantepec coast. (c) Monthly average of Sea Surface Temperature (SST), Chlorophyll a (Chl a), and Precipitation in the study area (data taken from IRI, 2015). (d) Photography of core Tehua XII E03b, the yellow asterisks indicate the key laminae used to correlate subcores.
During winter and early spring seasons, the ITCZ is located at its southernmost position, and easterly trade winds across southern Mexico and Central America are stronger than the rest of the year (Amador et al., 2006). These conditions allow the high-pressure systems south-eastward migration associated with cold-air outbreaks coming from the northwestern United States. The outbreaks generate airflows channeled through the low elevation gap of the Sierra Madre del Sur and reach the GoT as intense northerly winds (Amador et al., 2006), locally named Tehuanos (Trasviña et al., 1995). Tehuanos are perpendicular to the coast and cause mixing and stress curling of the surface water column on the wind axis (Trasviña et al., 1995). These winds can break up the stratification or raise the thermocline near the surface (Kessler, 2006) to even less than ~30 m depth (Lluch-Cota et al., 1997), and cold (~20°C), nutrient-rich subsurface waters fertilize the euphotic zone resulting in high primary productivity, including diatom blooms, as revealed by high chlorophyll-a concentrations (Figure 1). At this time, the ESW occupied the upper 35–40 to 70 m, the SSW is found until 430–470 m, the NPIW to 900–1200 m, and the NDPW below this depth (Machain–Castillo et al., 2008).
In contrast, during the summer and autumn seasons, the ITCZ is at its northernmost position, rainfall dominates in the ETNP due to the associated convective belt of the ITCZ, and trade winds are weaker than during the winter-spring period (Amador et al., 2006). Surface water circulation is dominated by tropical waters from the equatorial region flowing north through the Costa Rica Coastal Current (Kessler, 2006). The ESW is found between 55 and 70 m, SSW until 450 m, and the NPIW and NDPW depth limits are similar to winter (Fiedler and Talley, 2006; Machain–Castillo et al., 2008). Tehuanos are relatively sporadic and weak (Romero-Centeno et al., 2007), and the thermocline is deeper at ~70 m (Lluch-Cota et al., 1997). Hence upwelling and productivity decrease (Meave del Castillo and Hernández-Becerril, 1998), and the highest sea surface temperatures (SST) ~30.0°C are found (Figure 1).
Diatoms are unicellular algae and one of the main components of phytoplankton in the GoT, together with cyanophytes, dinoflagellates, and coccolithophorids (Meave del Castillo and Hernández-Becerril, 1998). One must consider that preserved diatoms silicate frustules on the seafloor may be a mixed assemblage (freshwater, marine, benthic, and planktonic). Furthermore, most of the information is lost during settling to the seafloor by dissolution, grazing, transport, reworking, and bioturbation (Crosta and Koc, 2007). However, upwelling regions with high productivity usually reflect the overlaying hydrographic conditions of the surface waters (Treppke et al., 1996), and have been used to reconstruct ocean environmental conditions during climate episodes such as the LIA and the Medieval Warm Period (MWP ~900 to ~1200 CE, e.g. Barron and Bukry, 2007; Barron et al., 2003). In addition, geochemical proxies such as organic carbon (Corg) and total nitrogen (TN) in sediments have been widely used to infer exported production to the seabed (Sifeddine et al., 2008; Choumiline et al., 2019; Salvatteci et al., 2014). The C:N ratio and δ13 C have been used to track the origin of sedimentary Corg, since marine phytoplankton and land vegetation carry a distinct C:N and δ13C signal (Lamb et al., 2006). Ni and Cu concentrations are also used as productivity tracers since they are used as a micronutrient for the phytoplankton (Smrzka et al., 2019; Tribovillard et al., 2006). Although, these elements can be affected by terrestrial input (Calvert and Pedersen, 2007; Smrzka et al., 2019), the normalization with a terrigenous element (such as Al or Ti) removes the terrestrial signal. Contamination by human impact is another factor that can alter trace element concentrations in sedimentary records (e.g. Ruiz-Fernández et al., 2004). In the GoT coastal zone, previous studies have reported increments in trace metal associated with the development of anthropogenic land-based activities, mainly oil production, urban wastes, and agricultural residues (Pica-Granados et al., 1994; Ruiz-Fernández et al., 2004). Ruiz-Fernández et al. (2004) reported that Cu background level in sediments (before 1860) was 138.9 mg/kg, and a moderate anthropic influence on Cu (from 150 to 480 mg/kg) since the 1980s off Salina Cruz at ~240 m water depth. Concerning Ni, Pica-Granados et al. (1994) reported current high Ni concentrations in water (from 49.8 to 81.7 mg/l) collected near the “Antonio Dovalí Jaime” oil refinery (~5 km from Salina Cruz Port, Figure 1) at <5 m water depth.
Redox processes can also affect the Ni and Cu signals deposited in hypoxic environments (Calvert and Pedersen, 2007; Smrzka et al., 2019). However, when Ni and Cu profiles have similar trends to other productivity proxies, such as Corg and biogenic silica, they can be considered as reliable productivity tracers.
In the GoT, most of the studies on phytoplankton have been focused on taxa distribution, species richness (Torres-Ariño et al., 2019 and references therein), and seasonal variability (e.g. Meave del Castillo and Hernández-Becerril, 1998; Moreno-Ruiz et al., 2011). This study provides the first high-resolution reconstruction of paleoproductivity based on diatoms, Corg, TN, Ni/Al, and Cu/Al of the last five centuries in a sequence of laminated sediments from the GoT and its relationship to ocean-climate variability. The study site is in a more tropical latitude. Nevertheless, we expect that the climate variability of the last centuries seen in more northern regions (e.g. Barron and Bukry, 2007; Choumiline et al., 2019; Goni et al., 2006; Juárez et al., 2014; Staines-Urías et al., 2009) can also be registered in the GoT sediments. . Tropical climates are naturally sensitive to climate variations (e.g. Li et al., 2020; Yamaguchi and Suga, 2019), as small SST increases further strengthen upper water column stratification (Amador et al., 2006; Fiedler and Talley, 2006), affecting productivity due to lower nutrient availability (Li et al., 2020; Yamaguchi and Suga, 2019). Therefore, this study helps to understand the effects of warming on tropical regions and their implications for future climate and ocean dynamics.
Material and methods
Sediment core
The present study is based on a Reineck box core, retrieved in March 2014 from the GoT (Tehua XII E03, 15.6442° N and 95.3071° W) at 743 m water depth, aboard the R/V “El Puma” from the National Autonomous University of Mexico (UNAM) (Figure 1). Two subcores were obtained; subcore Tehua XII E03a (34.5 cm long, 6.5 cm diameter) was used for 210Pb dating, Ni, and Cu analysis, and subcore Tehua XII E03b (32.5 cm long and 14 × 14 cm wide and high; Figure 1) was used for diatom, and the rest of geochemical analyses (Corg, TN, and δ13C), and radiocarbon (14C) dating.
Age dating
The radiochronology of the upper core segment (0–20 cm) of the subcore Tehua XII E03a was estimated through the 210Pb age dating method at 1 cm resolution (Table S1). 210Pb activities were determined at the laboratory of Isotopic Geochemistry and Geochronology at UNAM (Mexico) through high-resolution gamma ray spectrometry (HPGe well detector, Ortec-Ametek) as described by Díaz-Asencio et al. (2020). For validation, 239,240Pu was determined in selected sediment samples by alpha spectrometry (Ortec-Ametek Alpha Spectrometry system) (Ruiz-Fernández et al., 2014). Data quality was assessed through the analysis of the reference material IAEA-300 (Radionuclides in Baltic Sea sediment) and the results were within the reported range of recommended values.
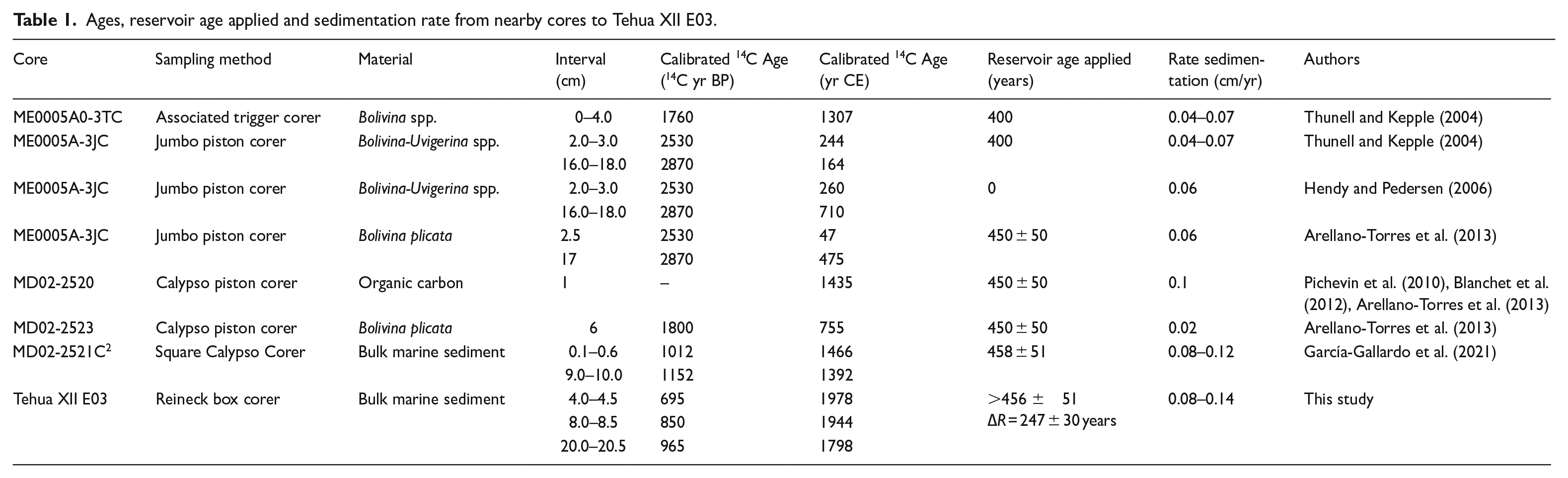
Radiocarbon ages were determined by accelerator mass spectrometry (AMS), at the National Platform LMC14, France, on five bulk sediment samples (4.0–4.5, 8.0–8.5, 20.0–20.5, 25.0–25.5, and 29.0–29.5 cm depth) of the subcore Tehua XII E03b (Table S2). We used the bulk sediment because of the poor preservation (Arellano-Torres et al., 2013) and recalcification of planktonic foraminiferal shells (Gibson et al., 2016). Previous studies in the GoT using AMS14C age dating on bulk sediment (Table 1) have shown that this method is reliable in the area (e.g. Arellano-Torres et al., 2013; Blanchet et al., 2012; García-Gallardo et al., 2021; Pichevin et al., 2010). To produce an integrated 210Pb-14C age model, both subcores were stratigraphically correlated by identifying common key laminae (light laminae visually identified in both subcores; Figure 1, Table S1) and 210Pb-derived dates for subcore Tehua XII E03a were transferred to subcore Tehua XII E03b (Table S1).
Ages, reservoir age applied and sedimentation rate from nearby cores to Tehua XII E03.
Geochemical procedures
Twenty-four dry sediment samples (~0.5 cm each) ground in a porcelain mortar, were used for geochemical analyses at the ALYSES platform (IRD/SU, Bondy France). The content of Corg and TN and the stable carbon isotopic ratio 13C/12C of the organic matter were determined on a Flash HT 2000 elemental analyzer, coupled to an isotopic ratio mass spectrometer Delta Vplus via a combustion-ConFlow IV interface from Thermo Fischer Scientific. The samples for Corg and δ13C analysis were treated with HCl 10% for removal of the carbonate fraction. Bulk sediment was used for TN analysis. The stable carbon isotopic ratio 13C/12C of OM is reported in the conventional δ-notation with respect to the PDB (Pee Dee Belemnite) carbonate standard defined with the equation
To evaluate the productivity changes along the core, Ni and Cu concentrations were used as productivity proxies (Smrzka et al., 2019; Tribovillard et al., 2006). As the Ni and Cu concentration in the sediments can be affected by terrestrial input, we normalized them with Al concentrations (Calvert and Pedersen, 2007). Ni, Cu, and Al concentrations were determined on 35 samples (~1 cm each) by X-ray fluorescence spectrometry (XRF, Spectrolab Xepos-3). Analytical precision was assessed through the replicate analysis (n = 3) of a single sediment sample, and the coefficients of variation were <0.6% for Ni and Cu, and <0.01% for Al). The accuracy of the measurements was evaluated through the analysis of the reference material IAEA-158 with results obtained within the reported certified values.
Diatom processing
Ninety-two sediment samples (~0.4 cm average thickness and ~6 years temporal resolution average; Table S3) were obtained for diatom analysis. Samples were processed at the Micropaleontology Laboratory of the Marine Sciences and Limnology Institute, UNAM. Laminae were visually recognized and separated using an x-ray digitalized acetate template. The dry weight of each sediment lamina was registered for abundance calculations. Sediment samples (~0.2 to ~0.5 g dry weight) were added with HCl 10% to eliminate carbonates and digested at ~80°C with H2O2 30% and HNO3 70% to remove organic matter. Acids and salts were removed through several washes with distilled water, settled for at least 24 h, and the liquid was discarded. The samples were diluted to a standard volume (30 mL).
For diatom slides, samples were homogenized and 200 µL aliquots were taken. In most samples, dilution was needed owing to the large number of particles, these volumes were considered for the calculation of total abundance. Aliquots were placed on coverslips (18 mm diameter) and dried at room temperature. Slides were mounted in Naphrax resin (refraction index = 1.74). All diatoms were identified at the genera or species taxonomic level using Cupp (1943), Round et al. (1990), Moreno et al. (1996), Hasle and Syvertsen (1997), Hernández-Becerril et al. (2021), and specialized literatures. The samples were counted by transects (Schrader and Gersonde, 1978), at least 500 valves per sample were identified under a light microscope (Nikon eclipse Ni) at 1000x magnification with Nomarski interference contrast. The precision of the diatom analysis was evaluated through the replicate analysis (n = 8) of diatom slides; the relative standard deviation was <8%. The species composition is reported as relative abundance (%).
Data analysis
In order to quantitatively define assemblages’ zone (periods) in the studied core, a CONISS (CONstrained Incremental Sums of Squares; Grimm, 1987) analysis was applied to a simplified diatom species matrix (taxa with total abundances >0.8%; Table S3) together with geochemical data (Corg, TN, δ13C, C:N, Ni/Al, and Cu/Al). A squared Euclidian matrix approach was used to quantify the dissimilarity between samples, and the number of statistically significant zones was established with a broken stick model (Bennett, 1996). These procedures were performed with the R packages “rioja” (Juggins, 2020) and “vegan” (Oksanen et al., 2020).
Results
Age-depth model
The 210Pb chronology was established a) with the Constant Flux model (Sanchez-Cabeza and Ruiz-Fernández, 2012), with uncertainties estimated through a Monte Carlo method with 106 simulations (Sanchez-Cabeza et al., 2014), and b) with a bayesian method (Aquino-López et al., 2018) by using the R package rplum (Blaauw et al., 2021), which has been successfully used in contrasting aquatic environments, including marine sediment cores (Aquino-López et al., 2020). Both chronologies were almost identical and were successfully validated with the 239,240Pu onset, which is a robust age-dating marker in regions far from nuclear testing grounds (Sanchez-Cabeza et al., 2021) (Figure 2).
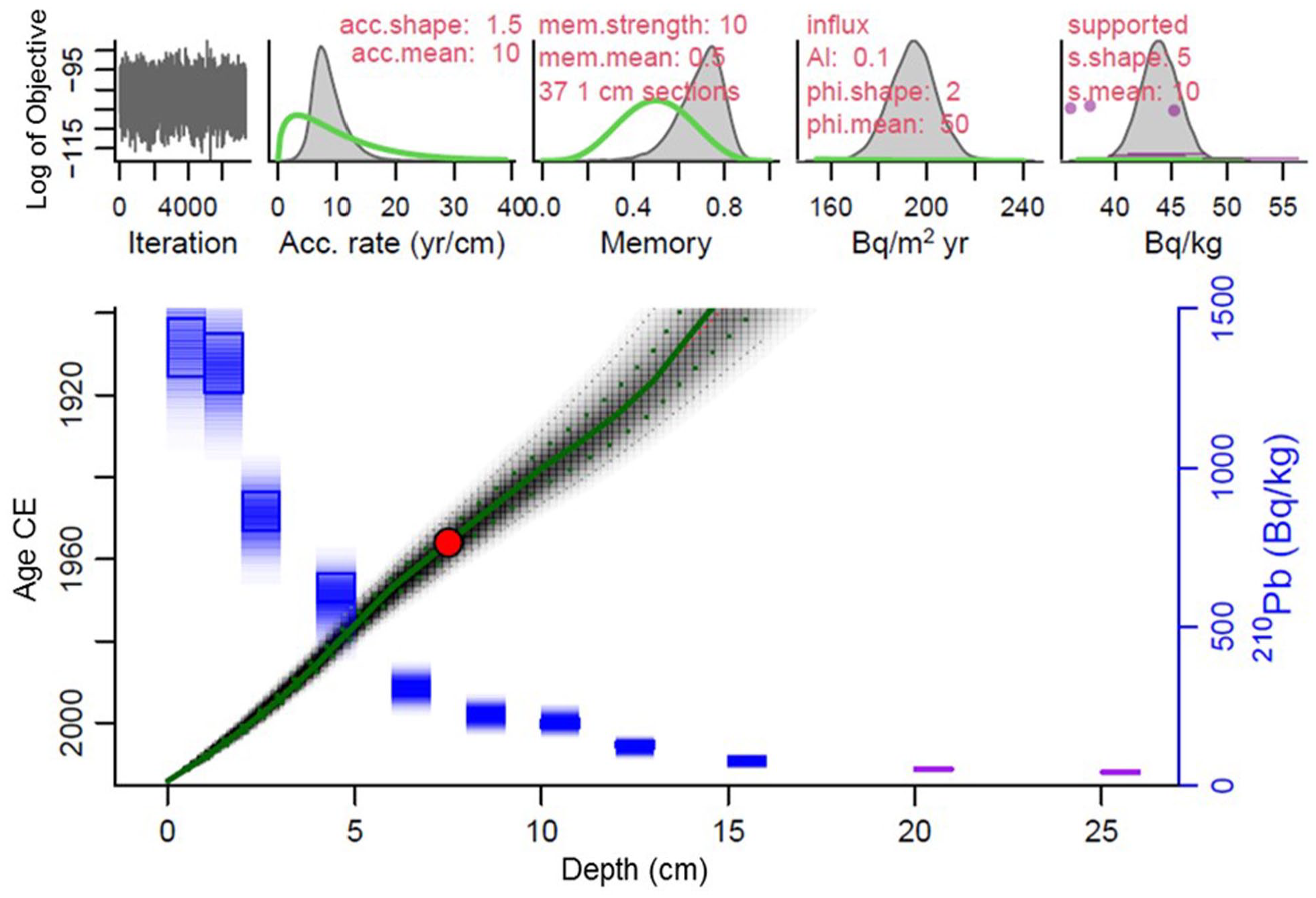
210Pb-derived age model for the core Tehua XII E03 using the Constant flux (green line) and Plum models (red line). The blue squares represent the 210Pb activities (± 2σ; right axis). The red point represents the 239,240Pu onset (red point) in 1954 CE (Common Era), used to validate the age model.
Following common practice, the radiocarbon ages were calibrated with the Marine20 curve (Heaton et al., 2020) and corrected for a regional marine reservoir age (ΔR) of 456 ± 51 years (Berger et al., 1966), but ages of sections 4.0–4.5 and 8.0–8.5 were significantly older than the 210Pb-derived ones (Table S1-S2). These discrepancies have been attributed to regional differences in the marine reservoir age (e.g. Hendy and Pedersen, 2006; Thunell and Kepple, 2004), which can be large in upwelling areas (Goodfriend and Flessa, 1997; Gutiérrez et al., 2009).
To estimate the local ΔR, we followed the method described by Reimer and Reimer (2017), which has been successfully used with 210Pb in the Baja California continental margin, also affected by upwelling (Treinen-Crespo et al., 2021). To calculate uncertainties, we used a Monte Carlo approach with the R language (R Core Team, 2021). The 210Pb-derived ages were expressed as BP (before present; age BP = 1950 – calendar age) and produced 106 simulations of the 210Pb age BP (pb210.bp) and the sample radiocarbon age (c14) following normal distributions. Simulated 210Pb BP ages younger than 1950 were discarded as they could not be calibrated with the MARINE20 curve. Each pb210.bp simulation was reverse-calibrated with the MARINE20 curve (pb210.c14) by using the calBP.14C function of the IntCal package (Blaauw, 2022). Then, the ΔR simulations were determined as the difference between the sample radiocarbon age simulations (c14) and the 210Pb reverse-calibrated age simulations (pb210.c14) as
The integrated 210Pb-14C age-model was produced by a bayesian approach (Blaauw and Christen, 2011) with the rbacon R package (Blaauw et al., 2022). We used all 210Pb-derived ages and the three radiocarbon 14C ages beyond the validated 210Pb chronology (sections below 20.0 cm), with the MARINE20 curve and the calculated ΔR. The integrated age-model showed a satisfactory agreement between 210Pb and 14C ages, and a common and smooth trend (Figure 3).
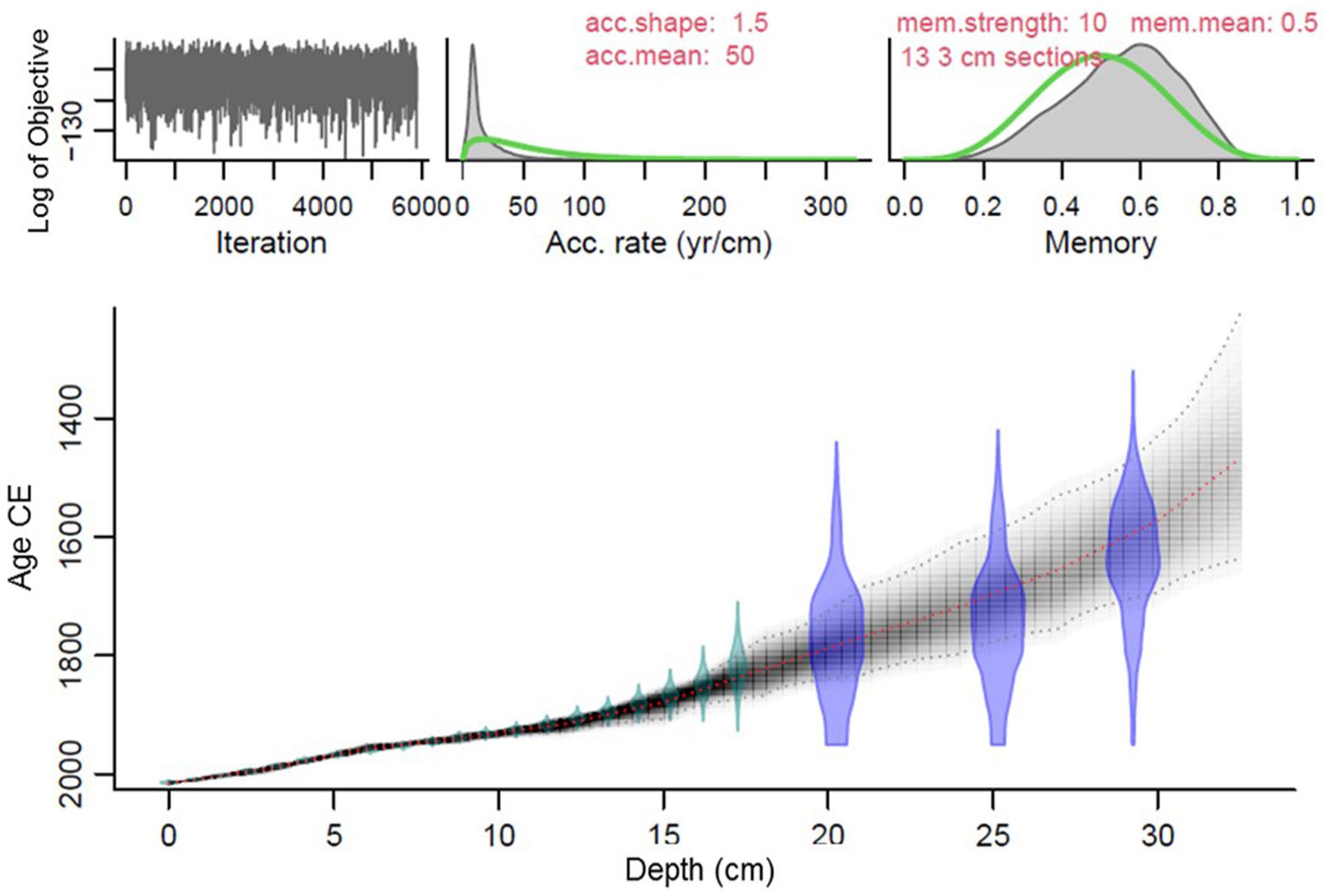
Integrated bayesian age model of the core Tehua XII E03 using MARINE20 curve and the calculated ΔR = 247 ± 30 years. 210Pb dates in cyan color and 14C dates in blue. CE = Common Era.
The integrated 210Pb and 14C age model indicated that the base of the core reached ~1500 CE (Table S4; Figure 3). The average sediment accumulation rate from ~1907 to ~2014 CE was 0.14 ± 0.07 cm/year; whereas for the older sediment, the sedimentation rate was of 0.08 ± 0.06 cm/year (Tables S1 to S2).
Geochemistry
The Corg (%) and TN (%) trends were similar along the core, with values ranging from 4.94 to 7.33% (average Corg = 6.35%) and 0.52 to 0.87% (average TN = 0.68%), respectively. Both elements displayed the highest values between ~1717 and ~1820 CE. The δ13C varied from -21.24 to -18.22‰ (average δ13C = −19.40‰) (Figure 4; Table S3). The Ni/Al ranged from 0.01 to 0.11 (average = 0.50), Cu/Al from 3.55 to 8.25 (average = 5.58) (Table S3). From ~1500 to ~1847 CE, Ni/Al and Cu/Al showed the highest concentrations (Figure 4).
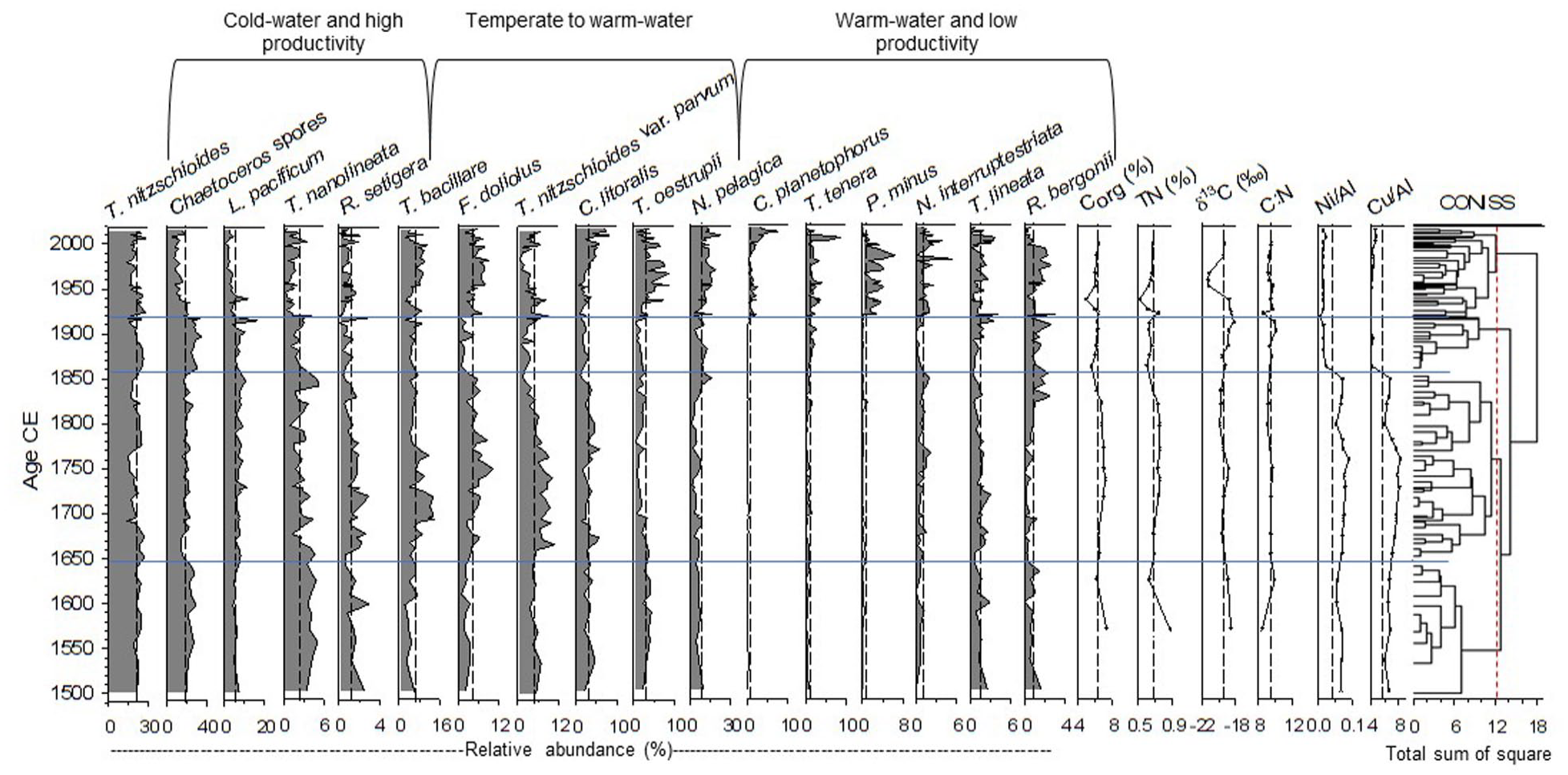
Relative abundance (%) and ecological group for the most common diatoms (>0.8%), Corg (%), TN (%), δ13C (‰), C:N, Ni/Al, Cu/Al, and CONISS dendrogram of the Tehua XII E03. Vertical dashed lines indicate the average values of each set. Horizontal (blue) lines indicate the four zones defined by CONISS analysis (at the 12.1 of total sum of squares indicated in red dotted line).
Diatoms
A total of 103 taxa belonging to 54 genera were determined; marine taxa were predominant and benthic and freshwater taxa represented only < ~9% of all studied sample. Specimens in the sediment core Tehua XII E03 were common to adjacent tropical Pacific regions and other upwelling areas. Seventeen taxa with a total relative abundance >0.8% made up 85.8% (Figure 4; Plate 1; Table S3). Thalassionema nitzschioides (20.9%) and Chaetoceros spores (mainly C. affinis, C. costatus, C. curvisetus, and C. radicans) (18.5%) were the dominant taxa, reaching up to 57% of total assemblages in the samples; followed by Neodelphineis pelagica (8.5%), Thalassionema bacillare (6.6%), Lioloma pacificum (5.8%), Thalassionema nitzschioides var. parvum (5.0%), Fragillariopsis doliolus (4.4%), Cyclotella litoralis (3.1%), Thalassiosira oestrupii (3.0%), Thalassiosira nanolineata (2.4%), Rhizosolenia setigera (1.5%), Thalassiosira lineata (1.4%), Thalassiosira tenera (1.4%), Nitzschia interruptestriata (1.1%), Rhizosolenia bergonii (0.9%), Plagiogramma minus (0.8%), and Cymatodiscus planetophorus (0.8%).
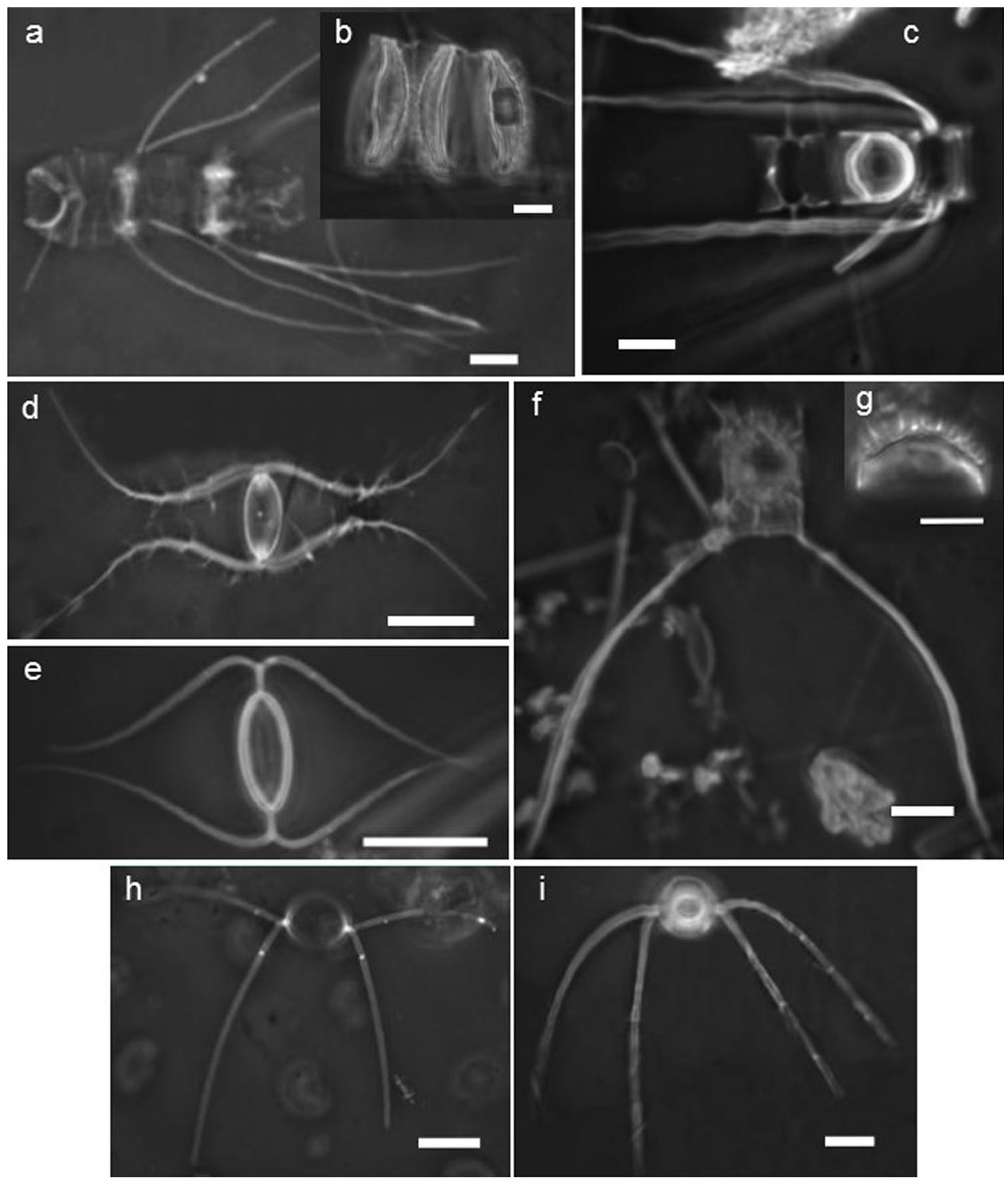
Light microscope photographs of vegetative cells with their resting spores of some of the predominant Chaetoceros species in the sediment traps from the GoT. (a) Chain valves and (b) chain resting spores of Chaetoceros costatus. (c) Chain valves and resting spore of Chaetoceros compressus. (d) Valve of Chaetoceros radicans and (e) its resting spore. (f–g) Terminal valve and resting spore of Chaetoceros affinis. (h) Valve of Chaetoceros curvisetus and (i) its resting spore. The scale bar is 10 μm in all images.
Ecological affinity of the main diatom taxa in the Gulf of Tehuantepec
Thalassionema nitzschioides is widely distributed from tropical to temperate waters (Hasle and Syvertsen, 1997) and usually appears throughout the year (Meave del Castillo, 2002; Meave del Castillo and Hernández-Becerril, 1998; Romero et al., 2009a; Sancetta, 1995); however, its largest abundance in subtropical to tropical regions occurs during the most productive season (e.g. Almaraz-Ruiz, 2017; Barron et al., 2010; Romero et al., 2011; Sancetta, 1995; Schrader et al., 1993; Treppke et al., 1996). Similarly, the Chaetoceros genus is usually predominant in regions with high nutrient availability (Hasle and Syvertsen, 1997; Lange et al., 1997; Meave del Castillo, 2002; Meave del Castillo and Hernández-Becerril, 1998; Rines and Hargraves, 1988; Romero et al., 2001, 2009a; Treppke et al., 1996), where it is commonly represented by resting spores in the sediment (e.g. Ren et al., 2014; Romero et al., 2009a; Sancetta, 1995). Unpublished data from sediment traps (~500 m depth) in the sediment core location during an ENSO-neutral year (February to July 2006) indicate that T. nitzschioides was more abundant during the most productive season, together with the Chaetoceros genus and L. pacificum, where Chaetoceros was represented mainly by coastal species in their vegetative forms (C. affinis, C. compressus, C. costatus, C. curvisetus, C. decipiens, C. diversus, C. laciniosus, C. lorenzianus, and C. radicans). Resting spores were only found for C. affinis, C. compressus, C. costatus, C. curvisetus, and C. radicans (Plate 1). In the sediments of core Tehua XII E03, no vegetative forms were found, only resting spores. We identified mainly four resting spore forms (C. affinis, C. costatus, C. curvisetus, and C. radicans; Plate 2, 7–10) that correspond to the most abundant Chaetoceros taxa found in the sediment traps during the upwelling season. Also, in the core sediments, the highest abundances of Chaetoceros spores occurred with T. nitzschioides and L. pacificum, as in the sediment traps during upwelling events. Therefore, we considered that the association of Chaetoceros spores, T. nitzschioides and L. pacificum recovered from the GoT sediments reflects the upwelling season conditions.

Scanning Electron Microscope (SEM) and Light Microscope (LM) images of the most abundant diatom taxa in the sediment core Tehua XII E03. (1) Cymatodiscus planetophorus (LM), (2) Thalassiosira tenera (LM), (3) Cyclotella litoralis (SEM), (4) Thalassiosira lineata (LM), (5) Thalassiosira nanolineata (SEM), (6) Thalassiosira oestrupii (SEM), (7) Chaetoceros affinis resting spore (SEM), (8) Chaetoceros curvisetus resting spore (SEM), (9) Chaetoceros radicans resting spore (internal view of the flat secondary valve, SEM), (10) Chaetoceros costatus resting spore (SEM), (11) Fragillariopsis doliolus (LM), (12) Neodelphineis pelagica (SEM), (13) Plagiogramma minus (LM), (14) Thalassionema nitzschioides var. parvum (SEM), (15) Thalassionema nitzschioides (SEM), (16) Rhizosolenia bergonii (LM), (17) Rhizosolenia setigera (LM), (18) Nitzschia interruptestriata (LM), (19) Lioloma pacificum (a-b in SEM, c in LM), and (20) Thalassionema bacillare (LM). Scale bar: 5 µm in 7–10 ; 10 µm in 1–6 and 11–15 ; 30 µm in 16–18, and 20 µm in 19a-c and 20.
Rhizossolenia setigera is sometimes found together with Chaetoceros in the California Current System, reflecting high productivity (Lange et al., 1997; Sancetta, 1995; Sautter and Sancetta, 1992). Thalassiosira nanolineata is recorded in Panama Basin as a coastal taxon; however, the species also showed a high factor score in the high productivity group (Romero et al., 2011, Table 1). Since in this study T. nanolineata exhibited its highest abundance together with T. nitzschioides, Chaetoceros spores, L. pacificum, and R. setigera, it was considered part of the assemblage of cold-waters and high-productivity taxa.
In the California Current System and the Eastern Equatorial Pacific, during warm and low productivity conditions, diatom assemblages are composed of one or more of the following species: F. doliolus, T. oestrupii, N. interruptestriata (Barron et al., 2010, 2013; Lange et al., 1987, 1990; Romero et al., 2011; Sancetta, 1992, 1995; Sautter and Sancetta, 1992; Schrader et al., 1993), T. bacillare (Baumgartner et al., 1985), T. nitzschioides var. parvum (Romero et al., 2011), T. lineata, R. bergonii (Baumgartner et al., 1985; Kemp et al., 2000; Lange et al., 1987), C. litoralis (Barron and Bukry, 2007; Barron et al., 2004, 2005; Sancetta, 1995), C. planetophorus (Estrada Gutiérrez et al., 2022), N. pelagica (Almaraz-Ruiz, 2017), and other taxa in minor amounts. In particular, R. bergoni is a deep-dwelling taxon (up to ∼ 130 m, Kemp et al., 2000) that indicates strong stratification in the water column in tropical to subtropical regions (Baumgartner et al., 1985; Kemp et al., 2000; Lange et al., 1987, 1994; Romero et al., 2011). For example, in the Santa Barbara Basin, R. bergonii (and other warm-water diatoms) showed high abundances during the 1983 El Niño (Lange et al., 1987). This trend also was observed in the Guaymas Basin during the 1957-1959, 1965, 1968–1969, and 1972 El Niño events (Baumgartner et al., 1985). El Niño on the Eastern North Pacific coast are characterized by a strong upper water column stratification and a deeper thermocline, which limit nutrients availability, and consequently low productivity (Pennington et al., 2006), where R. bergonii is one of the common species. However, some of these species have also been observed in temperate water, such as F. doliolus (Almaraz-Ruiz, 2017; Lange et al., 1994; Sancetta, 1995; Treppke et al., 1996), T. oestrupii (Almaraz-Ruiz, 2017; Hasle and Syvertsen, 1997; Romero et al., 2009b; Sautter and Sancetta, 1992), T. bacillare (Sancetta, 1995), T. nitzschioides var. parvum (Almaraz-Ruiz, 2017; Schrader et al., 1993; Treppke et al., 1996), and C. litoralis (Lange and Syvertsen, 1989; Romero et al., 2009a, 2011). Therefore, in this study T. bacillare, F. doliolus, T. nitzschioides var. parvum, C. litoralis, and T. oestrupii were considered temperate to warm-water taxa. Thalassiosira tenera is reported as cosmopolitan, except for polar regions (Hasle and Syvertsen, 1997; Li et al., 2013; Naya, 2012) and P. minus, as a widely distributed species (Guiry and Guiry, 2020); since these taxa exhibited in our record a similar distribution to warm-water N. pelagica, C. planetophorus, N. interruptestriata, T. lineata, and R. bergonii, they were interpreted as such (Figure 4).
CONISS analysis
According to the abundance of the main diatom species and their ecological affinities based on the literature, three broad ecological assemblages were adopted: cold-water and high productivity taxa, temperate to warm-water taxa, and warm-water and low productivity taxa (Figure 4). These assemblages and the CONISS zonation are described below.
Visual observation of the CONISS and the broken stick model (Fig. S1) suggest four significant zones (at the 12.1 of total sum of squares) in the studied core (Figure 4). The classification on the first hierarchical level presents two significant diatom assemblage zones, the lower part of the core between ~1500 and ~1919 CE (32.5 to 11.5 cm), and the upper part between ~1920 and ~2014 CE (11.4 to 0 cm). The classification on the second level subdivides the lower part into two zones from ~1500 to ~1858 CE (32.5 cm to 16.6 cm) and ~1860 to ~1919 CE (16.5–11.5 cm). Finally, a third level separates another zone between ~1500 and ~1648 CE (32.5–28.0 cm).
The lowest zone (~1500 to ~1648 CE) was characterized by the predominance of cold-water and high productivity taxa (T. nitzschioides, Chaetoceros spores, L. pacificum, T. nanolineata, and Rhizosolenia setigera) and most temperate to warm-taxa (T. nitzschioides var. parvum, C. litoralis, and T. oestrupii). The zone from ~1650 to ~1858 CE showed a relative increase of the temperate to warm-taxa (T. bacillare, F. doliolus, T. nitzschioides var. parvum, and C. litoralis). From ~1860 to ~1919 CE some cold-water and high productivity taxa (T. nitzschioides, Chaetoceros spores, and L. pacificum) and most warm-water and low productivity diatoms (N. pelagica, T. tenera, and R. bergonii) increased. The upper part (~1920 to ~2014 CE) was characterized by high abundances of all warm-water and low productivity taxa and almost all temperate to warm-water taxa (except T. nitzschioides var. parvum) (Figures 4 and 5).
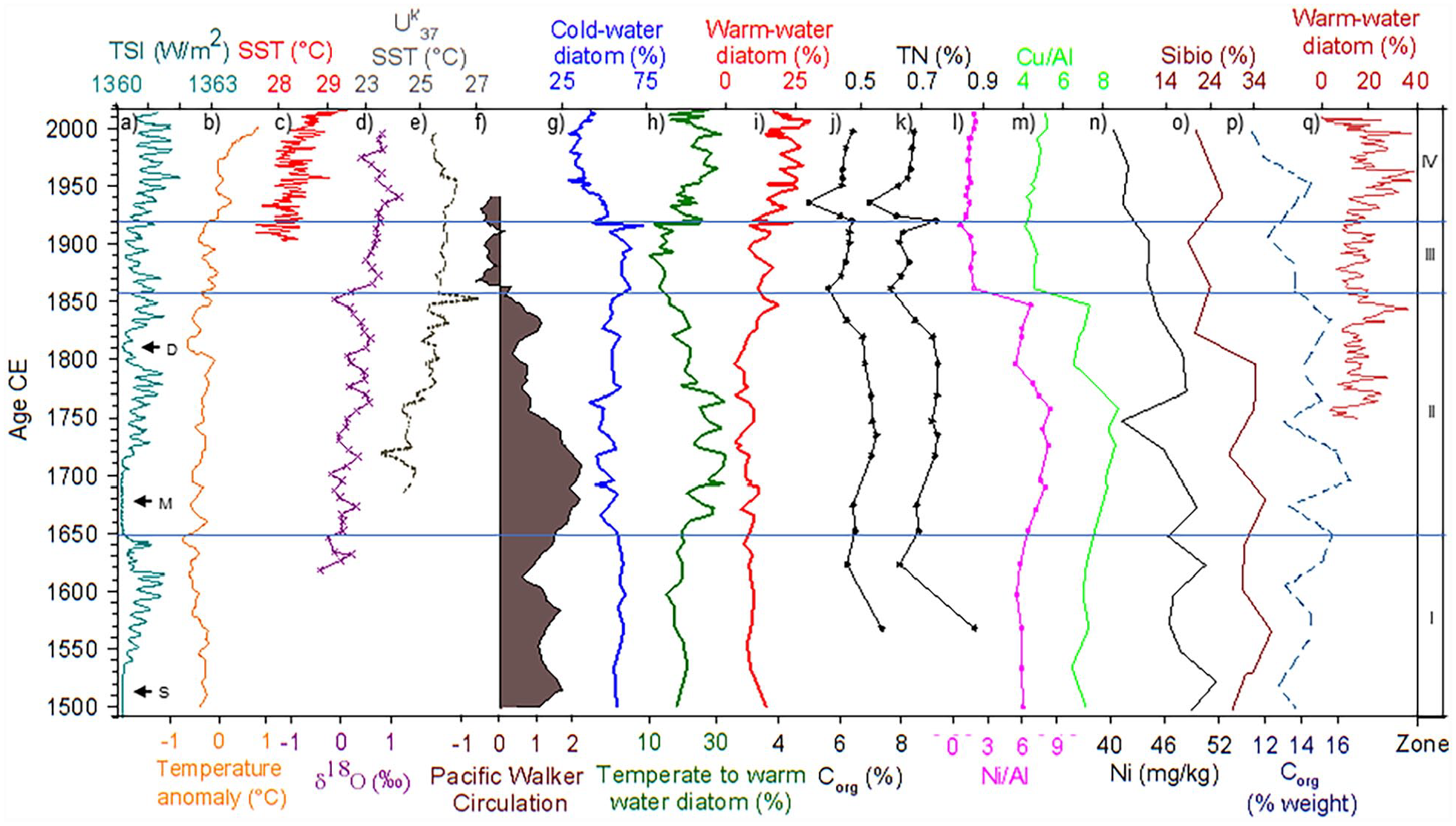
Paleoclimate records and the Tehua XII E03 record during the studied period. (a) Total Solar Irradiance (TSI, W/m2), Spörer (S), Maunder (M) and Dalton (D) minima (Lean, 2018). (b) Surface temperature anomaly (°C) from the Northern Hemisphere (Mann et al., 2009). (c) SST (°C) from GoT (IRI, 2015). (d) stratification of the water column (Staines-Urías et al., 2009) from Gulf of California. (e) Alkenone-derived SST (Uk′37 °C) from Guaymas Basin (Goni et al., 2006). (f) Pacific Walker Circulation strength (Griffiths et al., 2016). (g) Cold-water diatom (%), (h) temperate-water diatom (%), (i) warm-water diatom (%), (j) Corg (%), K) TN (%), (l) Ni/Al, and (m) Cu/Al from GoT (this study). (n) Ni concentration (mg/kg) (Choumiline et al., 2019) and (o) biogenic silica (Sibio %) (Barron and Bukry, 2007) from Gulf of California. (p) Corg (weight %) from off the Baja California margin (Juárez et al., 2014). (q) Warm-water diatom from Santa Barbara Basin (Barron et al., 2013). Horizontal lines indicate the periods (I to IV) identified with CONISS analysis.
Discussion
Chronology and sedimentation rate
The 210Pb dating is the most suitable method to obtain the chronology of recent sediments (~100 years) (Sanchez-Cabeza and Ruiz-Fernández, 2012), whereas the 14C method is widely used to date sediments beyond the 1700–1800s (e.g. Choumiline et al., 2019; Gutiérrez et al., 2009; Staines-Urías et al., 2009). Although there are several palaeoceanographic studies in the GoT, these studies were focused on climate changes at the millennial scale and according to their reported ages they do not cover the last hundred years (e.g. Arellano-Torres et al., 2013; García-Gallardo et al., 2021; Hendy and Pedersen, 2006; Pichevin et al., 2010; Thunell and Kepple, 2004; Table 1). This is the first high-resolution study in the region of the last ~500 years, where the environmental changes, linked to climatic variability, are based on an integrated 210Pb-14C age model.
A new 14C reservoir age (∆R = 247 ± 30 years; Figure 3) was calculated for bulk sediments of the core Tehua XII E03 (section 2.2). This ∆R value is higher than the reference reservoir age of the GoT proposed by Berger et al. (1966) (456 ± 51 years, ∆R = 18 + 50 years), the marine reservoir effect applied by Thunell and Kepple (2004) (400 years), and Hendy and Pedersen (2006) (Table 1). Our reservoir age is consistent with the independently obtained 210Pb dates (Figure 2; Tables S1-S2), which are reliably validated by the Pu onset (Figure 2).
The method to estimate ΔR has been used in sediment cores at Soledad Basin, Baja California, Mexico (Treinen-Crespo et al., 2021), where different ΔRs are found for organic matter and planktonic foraminifera. This suggests that the reservoir age in Turritella leverostoma shell (Berger et al., 1966) cannot be used for bulk sediment. In recent and high-resolution sedimentary records (such as Tehua XII E03), it is essential to obtain reliable local reservoir corrections, as ages can be shifted by decades, or even centuries (Treinen-Crespo et al., 2021), thus invalidating the paleoreconstructions.
The sedimentation rate for the last century was 0.14 ± 0.07 cm/year, within the range reported by Ruiz-Fernández et al. (2009) of 0.03–0.21 cm/year for the GoT. The sedimentation rate of 0.08 ± 0.04 cm/year for the older sediments was similar to values obtained in previous studies in GoT for the late-Holocene of ~0.1 cm/yr (Table 1) (Arellano-Torres et al., 2013; Blanchet et al., 2012; García-Gallardo et al., 2021; Hendy and Pedersen, 2006; Thunell and Kepple, 2004).
Origin of organic matter in the Tehua XII E03
The origin of sedimentary organic matter was inferred through the C:N ratio and δ13C that indicate the relative contributions of organic matter from different sources (Lamb et al., 2006). Typically, C:N ratio values above 12 and δ13C values that range from −33 to −25 ‰ suggest the predominance of terrestrial organic matter. C:N ratio values between 4 and 10, and δ13C values ranging from −21 to −18 ‰ indicate a marine origin (Lamb et al., 2006). Thus, our values of C:N ratio (~9) and δ13C (~-22 to ~−18 ‰) suggest that the organic matter preserved in the GoT sediments is predominantly of marine origin (Figure 4; Table S3).
Productivity variation in the Gulf of Tehuantepec over the last five centuries
The diatom abundance and geochemical proxies used in this study (Corg, TN, δ13C, Ni/Al, and Cu/Al) suggest relative productivity changes related to LIA and CWP conditions during the last five centuries. According to CONISS analysis, four significant zones were recognized within these two conditions (Figure 4). These periods are compared below (Figure 5) with solar irradiance (Lean, 2018), temperature records (Goni et al., 2006; Mann et al., 2009; Staines-Urías et al., 2009), and the Pacific Walker Circulation strength through the Southern Oscillation Index (SOI) (Griffiths et al., 2016). Also, we compared our record with other ETNP exported productivity records (Barron and Bukry, 2007; Barron et al., 2013; Choumiline et al., 2019; Juárez et al., 2014).
Little Ice Age (zone I ~1500 to ~1648 CE and zone II ~1650 to ~1858 CE)
During the LIA period, the predominance of cold-water and high productivity assemblage and the low abundance of temperate to warm-water and warm-water taxa, as well as the highest values of Corg, TN, Ni/Al, and Cu/Al (Figures 4 and 5), overall suggest the presence of upwelling and high productivity, presumably linked to LIA. Similar findings are observed in the Gulf of California and Baja California margin records during the LIA (Barron and Bukry, 2007; Barron et al., 2003; Choumiline et al., 2019; Juárez et al., 2014; Figure 5).
The LIA in the Northern Hemisphere has often been observed as negative surface temperature anomalies (~−0.7 to ~−0.3°C, Mann et al., 2009), associated with lower solar irradiance (Bard et al., 2000; Lean, 2018). Under these conditions, the mean position of the ITCZ is displaced southward (Griffiths et al., 2016; Haug et al., 2001; Sachs et al., 2009) related with a strengthened Pacific Walker Circulation (Griffiths et al., 2016). Although during the LIA, presumably, La Niña-like conditions were predominant (Beaufort and Grelaud, 2017; Griffiths et al., 2016; Yan et al., 2011), as it is suggested by the SOI (Figure 5, Griffiths et al., 2016), the temporal resolution of our samples does not allow direct comparison with ENSO variability.
The southern position of the ITCZ reinforced northeasterly trade winds into the Gulf of Mexico and the Caribbean Sea (Black et al., 1999; Nyberg et al., 2002), which reached the Pacific coast of Central America (Glynn et al., 1983) and the GoT. Also, a southward migration of the high-pressure systems may have resulted in more frequent outbreaks, particularly during winters (Lozano-García Mdel et al., 2007; Nyberg et al., 2002). As a result, strong Tehuanos winds blowing during the LIA, likely enhanced mixing and upwelling and therefore caused high productivity due to the nutrient input from subsurface waters into the photic zone (Lluch-Cota et al., 1997; Pennington et al., 2006). In the Gulf of Papagayo, close to the GoT, Glynn et al. (1983) reported the demise of coral reefs likely caused by low SST, triggered by an increase in duration or intensity of seasonal upwelling during the LIA (~1600 to ~1900 CE).
Our diatom record suggests that the LIA was not a homogeneous cold period. The increased abundance of the temperate to warm-water assemblage (Figure 4) suggests less cold and less productive conditions from ~1650 to ~1858 CE (zone II), although our geochemical productivity proxies in this interval show a slight decrease after ~1850, similar to those found in the Gulf of California (biogenic silica %, Barron and Bukry, 2007) and the Baja California margin (Corg, Juárez et al., 2014) (Figure 5).
The relative decrease in productivity suggested by our diatom record between ~1650 and ~1858 CE is associated with the increase in solar irradiance after the Maunder minimum and the decrease in Pacific Walker Circulation (Figure 5). Other studies in the ETNP have also indicated the heterogeneity of the LIA, also attributed to solar activity variations (Barron and Bukry, 2007; Barron et al., 2003; Choumiline et al., 2019; Cuna et al., 2014; Lozano-García Mdel et al., 2007; Rodríguez-Ramírez et al., 2015; Staines-Urías et al., 2009).
Transition period (zone III ~1860 to ~1919 CE)
Although there is still no consensus in the ETNP records about the end of the LIA, we identified a transition period from ~1860 to ~1919 CE that could be interpreted as the end of LIA and the beginning of the CWP. The records closest to the GoT report the end of LIA at about ~1750 CE (Ricaurte-Villota et al., 2013), ~1820 CE (Juárez et al., 2014), ~1850 CE (Barron and Bukry, 2007; Barron et al., 2003; Choumiline et al., 2019; Cuna et al., 2014; Del Castillo-Batista et al., 2018), and even ~1900 (Hodell et al., 2005).
In this interval, though some cold-water and high productivity taxa (Chaetoceros spores, T. nitzschioides, and L. pacificum) remained abundant, the increased abundance of warm-water and low productivity taxa (N. pelagica, T. tenera, and R. bergonii) characterized the period, indicating the transition toward warmer and less productive conditions (Figure 4). Likewise, the sediments from this period were characterized by lower values of Corg, TN, Ni/Al, and Cu/Al ratio, suggesting a reduction in the productivity of the GoT, similarly to some productivity proxies from Gulf of California and off the Baja California margin (Barron and Bukry, 2007; Barron et al., 2003; Choumiline et al., 2019; Juárez et al., 2014; Figure 5).
As mentioned in section 3.4, R. bergonii is a deep-dwelling taxon that indicates strong stratification in the water column (Baumgartner et al., 1985; Kemp et al., 2000; Lange et al., 1987, 1994; Romero et al., 2011). In the GoT, greater stratification occurs during the summer and autumn when the thermocline and nutricline are found beyond ~50 m (~30 m during winter), and the upwelling and mixing events are restricted (Lluch-Cota et al., 1997). Also, this taxon is common during El Niño events on the Eastern North Pacific coast. Thus, the increased abundance of R. bergonii suggests that the GoT water column became more stratified around the mid-1800s, similar to the findings of Staines-Urías et al. (2009) (Figure 4).
The general trend of this period shows a progressive warming since ~1850 CE (IPCC, 2014) largely associated with solar irradiance (Bard et al., 2000; Hoyt and Schatten, 1993; Lean et al., 1995) and global warming (IPCC, 2014). This warming trend is also observed in the Gulf of California by Staines-Urías et al. (2009), who pointed out rapid warming since the mid-1800s and enhanced water column stratification. Also, Goni et al. (2006) found a general trend of increased SST of 1–2°C in Guaymas and Cariaco Basins since ~1800 CE (Figure 5) and related them to the end of the LIA. Likewise, this period coincides with the weakening of the Pacific Walker Circulation since ~1860 CE (Griffiths et al., 2016; Yan et al., 2011) and the northward migration of the ITCZ (Sachs et al., 2009). Lacustrine sediments from central Mexico recorded wetter conditions after the LIA, coinciding with the ITCZ’s northward displacement (Cuna et al., 2014). These conditions probably led to a weakening of the northeasterly trade winds over the Caribbean Sea (Nyberg et al., 2002), as well as to lesser frequent polar outbreaks over the Gulf of Mexico (Lozano-García Mdel et al., 2007), which would result in weaker Tehuanos winds in the GoT and consequently less upwelling and lower productivity. These findings together with those reported in the northwestern region of Mexico suggest warmer climate conditions and a likely decline in productivity during this transitional period in the ETNP.
Current warm period (zone IV ~1920 CE to ~today)
The most recent period is characterized by the highest abundance of the warm-water and oligotrophic assemblage together with a high abundance of moderate to warm conditions taxa (Figure 4). High abundances of N. interruptestriata, T. nitzschioides var. parvum, and F. doliolus have been interpreted in La Paz, to reflect an incursion of tropical/subtropical water during anomalously warm periods (Acevedo-Acosta et al., 2021). Also, the rapid increase up to ~7.4% of the C. planetophorus (Figure 4), a tropical taxon commonly found at temperatures above 23.3°C in the northwestern coasts of Mexico (Estrada Gutiérrez et al., 2022), supports the idea that warmer conditions prevailed during this period. Diatom sedimentary records (Barron and Bukry, 2007; Barron et al., 2013; Esparza-Alvarez et al., 2007; Martinez -Lopez et al., 2007) and planktonic foraminifera (Field et al., 2006) from the Eastern North Pacific also showed evidence of the global warming trend. For example, the percentage of warm-water diatoms (and silicoflagellates, not graphed in Figure 5) from Santa Barbara Basin exhibited a clear increase since ~1920 CE (Barron et al., 2013), in agreement with the peak of Alkenone-derived SST (Uk′37) from the Guaymas Basin during the same time (Goni et al., 2006) (Figure 5). Goni et al. (2006) attributed this increase in Uk′37 SST to the northernmost Subtropical High and ITCZ migration in response to the Northern Hemisphere insolation. Under boreal summer conditions, the atmospheric regime results in weak winds over the central and southern regions of the Gulf of California; consequently, upwelling shuts down, leading to progressive warming and thermal stratification in the Guaymas Basin. In addition, the Uk′37 SST peak after ~1920 CE could also reflect the northernmost incursion of warmer tropical water in the Eastern Tropical Pacific, similar to El Niño conditions in this region (e.g. Lavín et al., 1997).
On the other hand, our Corg, TN, Ni/Al, and Cu/Al records also remain with low values (Figures 4 and 5; Table S6). Similarly, geochemical productivity proxies from the Gulf of California indicated reduced productivity in this period (Barron and Bukry, 2007; Choumiline et al., 2019; Juárez et al., 2014; Figure 5). In the GoT, human impact contamination has been identified by the increase in trace metals in the coastal and shelf zones (Pica-Granados et al., 1994; Ruiz-Fernández et al., 2004); however, since our sedimentary record is further seaward and deeper, our Ni and Cu concentrations were lower (16. 8 mg/kg for Ni and 47.0 mg/kg for Cu) than in the coastal and shelf zones. Therefore, we considered that our Ni and Cu records are not masked by human impact. Also, these elements’ trend has been decreasing since ~1860 CE (together with those of the other productivity proxies) before the anthropic influence was evident (~1980) in the region (Ruiz-Fernández et al., 2004).
During this period, a generalized warming trend is observed in diverse records (IPCC, 2014; Mann et al., 2009), including the Eastern North Pacific (Barron et al., 2013; Goni et al., 2006; Staines-Urías et al., 2009). In the SST record of the GoT, the evident warming trend of surface water in the 20th century (IRI, 2015) (Figure 5) leads us to infer that the reduced productivity during the CWP is associated with the warming trend, which caused a sharp upper ocean stratification, as is observed in the δ18O of the planktonic foraminiferal record from the Gulf of California (Staines-Urías et al., 2009; Figure 5).
Our observations document the dominance of warm conditions and low productivity since ~1920 CE in a more tropical area (~15°N latitude). These results agree with previous studies further north; therefore, this study documented the regional scale of the warming trend and its consequence on productivity in the tropical oceans.
Conclusions
This study provides a new ΔR = 247 ± 30 years for the last 500 years in the Gulf of Tehuantepec, estimated through the combinations of 210Pb and 14C methods in a bayesian model.
The diatom and geochemical proxies analyzed in the Gulf of Tehuantepec sediments reflected two predominant productivity conditions during the last five centuries, higher productivity conditions during the Little Ice Age and lesser productivity conditions during the Current Warm Period.
During the part of the LIA found in our record, low solar irradiance (Spörer, Maunder and Dalton minima) promoted the southward migration of the Intertropical Convergence Zone, which likely promoted more Tehuanos winds in the Gulf of Tehuantepec. This scenario resulted in enhanced upwelling and higher productivity as evidenced by the prevalence of cold and high productivity taxa (Chaetoceros spores, T. nitzschioides, L. pacificum T. nanolineata, and R. setigera) and high values of Corg, TN, Ni/Al, and Cu/Al. Temperate conditions and lesser productivity from ~1650 to ~1858 CE, were suggested by the increase of some temperate to warm-water taxa (T. bacillare, T. nitzschioides var. parvum, F. doliolus, and T. lineata).
A transitional period (~1860 to ~1919 CE) toward the Current Warm Period was indicated by the appearance of most warm-water taxa (N. pelagica, T. tenera, and R. bergonii) as well as lower values of Corg, TN, Ni/Al, and Cu/Al. From ~1920 to ~2014 CE, the dominance of temperate to warm-water and mainly warm-water taxa (T. bacillare, F. doliolus, C. litoralis, T. oestrupii, N. pelagica, C. planetophorus, T. tenera, P. minus, N. interruptestriata, T. lineata, and R. bergonii) and low values of geochemical productivity proxies indicated even warmer conditions and lower productivity. These conditions were associated with diminished upwelling, and probably increased upper water column stratification and deeper thermocline derived from prevalence of warming trend, the northward migration of the ITCZ and weakening of the Pacific Walker Circulation.
Our findings highlight the importance of high-resolution productivity reconstructions and contribute to the knowledge of the response of the tropical regions to the Little Ice Age and Current Warm Period climates.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836231183057 – Supplemental material for Diatom-based paleoproductivity and climate change record of the Gulf of Tehuantepec (Eastern Tropical Pacific) during the last ~500 years
Supplemental material, sj-xlsx-1-hol-10.1177_09596836231183057 for Diatom-based paleoproductivity and climate change record of the Gulf of Tehuantepec (Eastern Tropical Pacific) during the last ~500 years by Laura Almaraz-Ruiz, María Luisa Machain-Castillo, Abdelfettah Sifeddine, Ana Carolina Ruiz-Fernández, Joan-Albert Sanchez-Cabeza, Alejandro Rodríguez-Ramírez, Perla Guadalupe López-Mendoza, Mercedes Mendez-Millan and Sandrine Caquineau in The Holocene
Footnotes
Acknowledgements
We thank the crew of the R/V “El Puma” for their assistance during the TEHUA XII expedition and core collection. Libia Hascibe Pérez Bernal performed the radiometric analysis for 210Pb dating. Laura Elena Gómez-Lizárraga provided technical aid with the SEM images at the ICML, UNAM. We are also indebted to anonymous reviewers for their valuable suggestions to improve this paper.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Financial support for the development of this project and publication costs was provided by the Instituto de Ciencias del Mar y Limnología, UNAM. Support for field sampling was provided by the UNAM through oceanographic mission TEHUA XII. The first author (LAR) thanks the Graduate Program in Marine Science and Limnology at UNAM, Mexico, and the financial support provided by the National Council of Science and Technology (CONACYT) for the doctoral scholarship (Grant number: 556646), as well as the Institut de Recherche pour le Développement (IRD) for the fellowship grant for an academic stay at the Laboratory of Oceanography and Climate (LOCEAN), IRD France-Nord. Also, the authors are grateful to the ALYSES platform for their support for carbon and nitrogen analyses.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.