
Abstract
At the end of the last glacial period in the northern hemisphere, meltwater from receding ice sheets accumulated into large proglacial lakes, potentially limiting postglacial afforestation. We explored whether former islands of proglacial Lake Ojibway (Canada) (hilltops in the current landscape) could have acted as migration outposts and thus accelerated the postglacial migration. We extracted sediments from two small lakes located on “paleo-islands” and used XRF to detect changes in soil erosion and vegetation biomass. We also used plant macro-remains and wood charcoal to determine if (and which) tree species colonized the sites and to detect local fire events. Organic sediment accumulation started around 9657 and 9947 cal. yr BP at Lakes Perché and Despériers, respectively, before the level of Lake Ojibway started to decrease and liberate parts of the studied landscape ca 9400 cal. yr BP. Lithogenic elements (Ti, K, Sr, Fe, Zr, and Rb) decreased between the beginning of organic sediment accumulation and 8800–8700 cal. yr BP, indicating reduced soil erosion, possibly due to soil stabilization by vegetation. Then, the S/Ti ratio, a proxy of organic matter increased around 8800 and 8400 cal. yr BP. The earliest tree macro-remains (Larix laricina and Pinus spp.) were found between 9850 and 9500 cal. yr BP. Local fires were detected around 9820 and 8362 cal. yr BP. Early afforestation occurred on the islands of Lake Ojibway, 200 and 450 years before its level started to decrease, confirming that some islands acted as migratory outposts accelerating postglacial migration.
Keywords
Introduction
At the end of the Pleistocene, large portions of the northern hemisphere were covered by ice sheets which were progressively retreating to the north. From the beginning of the Holocene (11,700 years ago) to the Mid-Holocene (ca 6000 years ago), the Laurentide Ice Sheet slowly melted over north-eastern Canada (Dalton et al., 2020). As landscapes became free of ice, they were available for colonization by vegetation from forests located at the southern margins of ice sheets.
Meltwater from receding ice sheets sometimes accumulated at their front and created so-called proglacial lakes (Carrivick and Tweed, 2013) that prevented vegetation colonization of deglaciated areas for centuries. Several Holocene proglacial lakes have been identified in Canada, including Lake Ojibway, one of the largest proglacial lakes (115,000 km3), covering what is currently western Quebec and eastern Ontario (Godbout et al., 2020; Veillette, 1994; Vincent and Hardy, 1977). Lake Ojibway was formed because the crustal depression beneath the ice caused the adjacent terrain to slope down toward the north, which prevented meltwater from leaving the ice front (Figure 1). The ice sheet produced meltwater, which was prevented from flowing north by the ice itself acting as a wall. Water accumulated at the ice sheet front and created proglacial Lake Ojibway around 10,570 cal. yr BP (calibrated years before present). Then, as the ice sheet continued its northward retreat, the southern end of James Bay was eventually freed from ice and Lake Ojibway gradually drained into it. Different phases of lake level have been recorded before the final drainage of Lake Ojibway. The level of proglacial Lake Ojibway reached its maximum during the Angliers phase ca 9400 cal. yr BP (Brouard et al., 2021; Godbout et al., 2020). Then, lake level gradually decreased and was ca 65 m lower around 9000 cal. yr BP when the Kinojévis phase started. As the level of Lake Ojibway decreased, more and more islands emerged. The level of Lake Ojibway had lowered ca 35 m more, at the beginning of the La Sarre phase ca 8300 cal. yr BP (Brouard et al., 2021). Several dates were proposed for the final drainage of proglacial Lake Ojibway, with most studies placing it at ca 8200 cal. yr BP (Brouard et al., 2021; Daubois et al., 2015; Godbout et al., 2020; Roy et al., 2015).
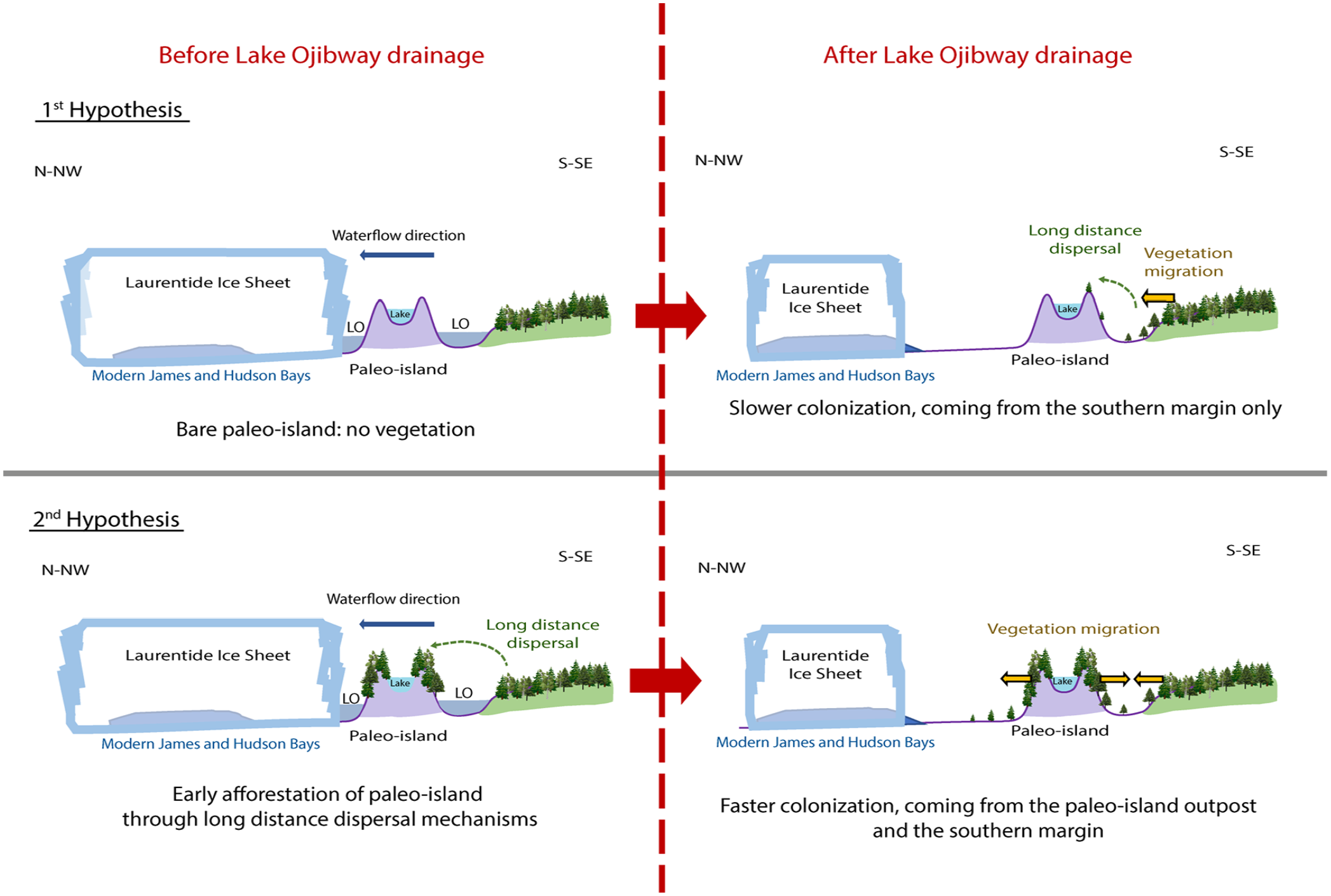
Schematic representation of proglacial Lake Ojibway (LO) creation and hypotheses concerning the colonization of paleo-islands before the drainage of Lake Ojibway and implications on postglacial migration thereafter.
The observed postglacial migration of several tree species in western Quebec was faster than the estimated migration time based on the mean distance dispersal capacities observed nowadays, a phenomenon called Reid’s paradox of rapid plant migration (Clark, 1998). Indeed, several vegetation reconstructions based on palynological and macro-remain studies in Western Quebec indicated that afforestation closely followed the drainage of Lake Ojibway (Ali et al., 2008; Carcaillet et al., 2001; Richard, 1980). Such rapid colonization can either be explained (1) by the presence of glacial refugia (Brochmann et al., 2003; Varga, 2010); or (2) by long distance dispersal events (Bullock et al., 2017; Cain et al., 2000). As there is no evidence that glacial refugia were ever present in western Quebec, the second hypothesis can be retained but remains to be supported by empirical data.
During its >2300 years presence over western Quebec and north-eastern Ontario, Lake Ojibway should have acted as a barrier slowing down postglacial forest migration, unless its islands were colonized early and acted as outposts facilitating colonization of lowlands following the drainage of Lake Ojibway into James Bay. For the islands of Lake Ojibway to be colonized, long-distance dispersal events from the vegetation front south of the lake were necessary. According to Schurr et al. (2009), long-distance seed dispersal via abiotic or biotic action happens when ⩽1% of seeds travel >1 km. Although such events are rare, they can strongly affect plant biogeography and population dynamics (Cain et al., 2000).
If the “paleo-islands” on Lake Ojibway had remained uncolonized, while Lake Ojibway was present, that is, if no long-distance dispersal events succeeded, colonization following the drainage of the proglacial lake would have followed a south-north trajectory limited by average seed dispersal distance, as most tree species in north-eastern Canada are wind-dispersed (Burns and Honkala, 1990) (Figure 1 – first hypothesis). This scenario does not explain how sites located several hundreds of kilometers north of the migration front could have been afforested almost immediately after the drainage of Lake Ojibway (Ali et al., 2009). An alternative hypothesis would be that some islands on Lake Ojibway were colonized through long-distance seed dispersal before the final drainage of the proglacial lake into James Bay, which would explain how sites far from the postglacial migration front could have been afforested quickly, then our study would enlighten an early colonization of paleo-islands, that is, while the Lake Ojibway still covered Quebec (Figure 1 – second hypothesis, Figure 2).
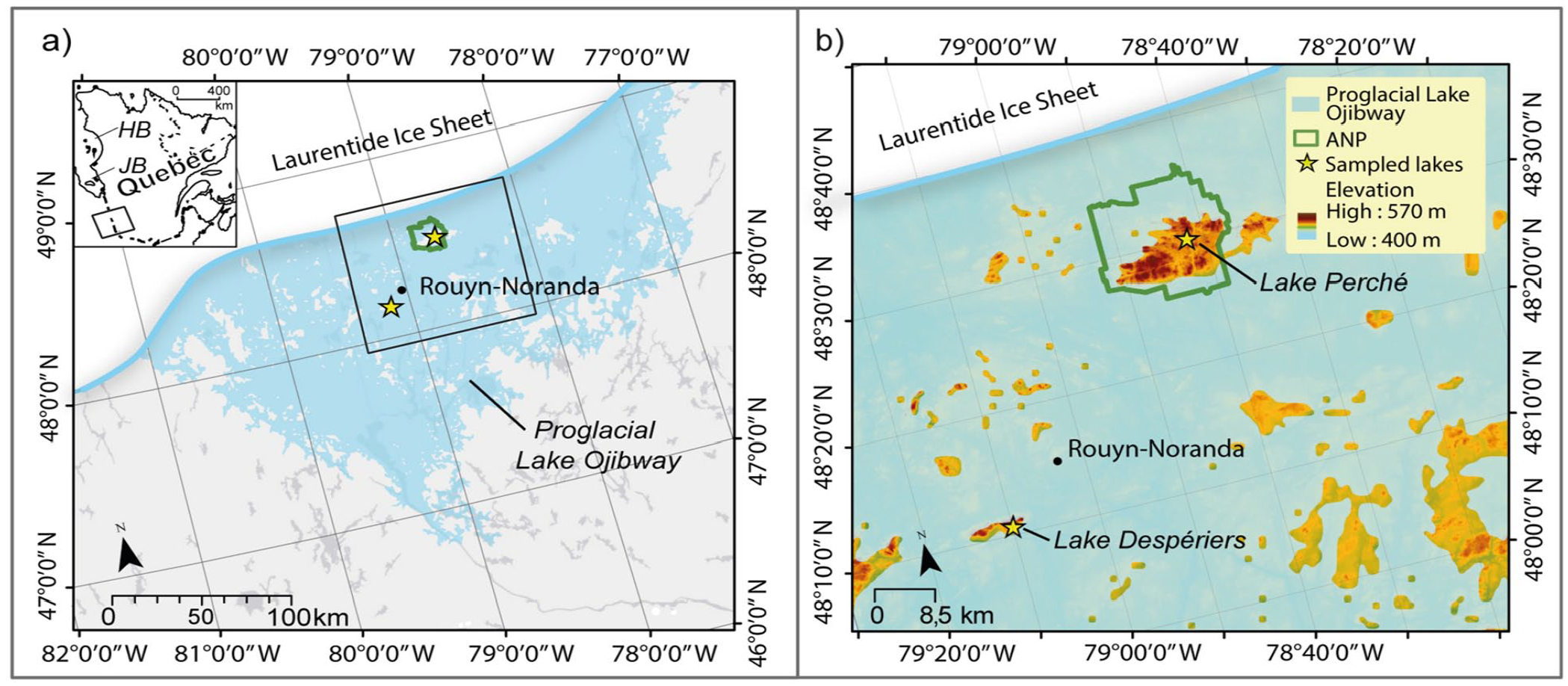
(a) Extent of proglacial Lake Ojibway in western Quebec around 9600 cal. yr BP (Brooks, 2020), showing the locations of the two studied paleo-islands (yellow stars). The location of Aiguebelle National Park (ANP) is also shown. The inset shows the location of ANP relative to Hudson Bay (HB) and James Bay (JB), where proglacial Lake Ojibway drained around 8200 cal. yr BP. (b) Close-up of the study area showing paleoislands (in light to dark orange), including the Abijévis and Kekeko Hills, where Lakes Perché and Despériers were sampled, respectively.
To test the hypothesis of early afforestation of paleo-islands through long-distance dispersal events (second hypothesis), we sampled sediments from two small lakes located on paleo-islands (current hilltops above the maximum shoreline level of Lake Ojibway) (Godbout et al., 2017), and we radiocarbon-dated bulk sediment and macro-remains to build age-depth models. We used multiple proxies to infer afforestation: X-ray fluorescence (XRF) geochemical analysis to detect variations in soil erosion as a proxy for plant cover; plant macro-remain analysis to attest local vegetation presence; and charcoal analysis to reconstruct local wildfire activity as an indicator of biomass burning. Only a few studies have been conducted so far on the afforestation of sites located near the maximum shoreline levels of proglacial lakes in eastern Canada (Richard, 1980; Richard and Larouche, 1989) and they did not allow to clearly demonstrate the potential role of paleo-islands on postglacial forest migration. Our study documents environmental conditions above the maximum limit of Lake Ojibway, therefore allowing us to test the hypothesis of early colonization of paleo-islands on a proglacial lake.
Material and methods
Study area
The study area is located in the Abitibi-Témiscamingue region of western Quebec and is part of the clay belt of Quebec and Ontario created by deposits of proglacial Lake Ojibway (Figure 2a). The Abijévis hills are located within the Aiguebelle National Park, whereas the Kekeko hills are 50 km southward of the park. Both hill formations originated from marine volcanism and are composed of mafic rocks, that is, mostly non-carbonate rocks such as gabbro and andesite. Surface deposits are glacial till (MERN - Système d’information géominière, 2022; Nutz et al., 2020). The hills were aerially exposed following ice sheet retreat ca 10,650 cal. yr BP (Dyke, 2004; Veillette, 1994). Although they were rounded by ice sheet action, the hills remain among the highest elevations in Abitibi-Témiscamingue, with maximum elevations of 570 and 488 m above sea level (a.s.l.) for Abijévis and Kekeko, respectively. They are surrounded by a generally flat topography (between 180 and 350 m a.s.l.).
Ancient shorelines of proglacial Lake Ojibway are still visible on hillsides in the current landscape and allowed reconstructions of lake level history (Godbout et al., 2017, 2020) and the identification of several paleo-islands that became hills following the final drainage of Lake Ojibway ca 8200 cal. yr BP (Richard, 1980; Richard et al., 2020; Veillette, 1994). Lake Ojibway’s highest shorelines (ca 9400 cal. yr BP) are estimated to be slightly below 400 m a.s.l. and 380 m a.s.l. for the Abijévis and Kekeko hills, respectively (Godbout et al., 2017). Areas above these elevations are therefore considered to have always been paleo-islands (Figure 2b).
Site selection and lake sediment sampling
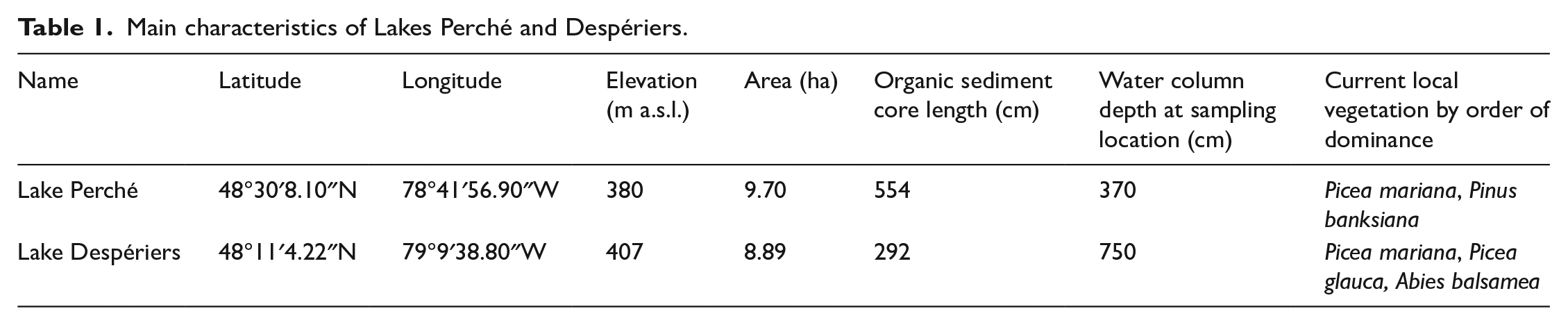
We selected two small headwater lakes surrounded by steep shores to maximize sediment collection: Lake Perché (formerly known as Lake Edrien) in the Aiguebelle National Park (Abijévis hills) and Lake Despériers on the Kekeko hills (Table 1; Figure 2). Lake Perché is located at 380 m a.s.l. While this is slightly below the highest shoreline of Lake Ojibway in the area (400 m a.s.l.), Lake Perché was formed before the complete drainage of proglacial Lake Ojibway and could have recorded part of the paleo-island’s history. Lake Despériers is located at 407 m a.s.l., that is, almost 30 m above the highest shoreline of Lake Ojibway in the area (Table 1). Current forest vegetation around the sampled lakes is dominated by black spruce (Picea mariana [Mill.] BSP), white spruce (Picea glauca [Moench] Voss), jack pine (Pinus banksiana Lamb.) and balsam fir (Abies balsamea [L.] Mill.). Both lakes are surrounded by rolling hills less than 100 m in height.
Main characteristics of Lakes Perché and Despériers.
We extracted lake sediment cores from the center of each lake with a 60 mm diameter Livingstone corer. We selected headwater lakes to avoid sediment disturbance by incoming water flows. We collected complete gyttja sequences (organic sediments), from the water-sediment interface to the contact with the underlying mineral glacial sediments. We stored sediments in plastic tubes and kept them in cold conditions (4°C) until further analyses.
Age-depth models
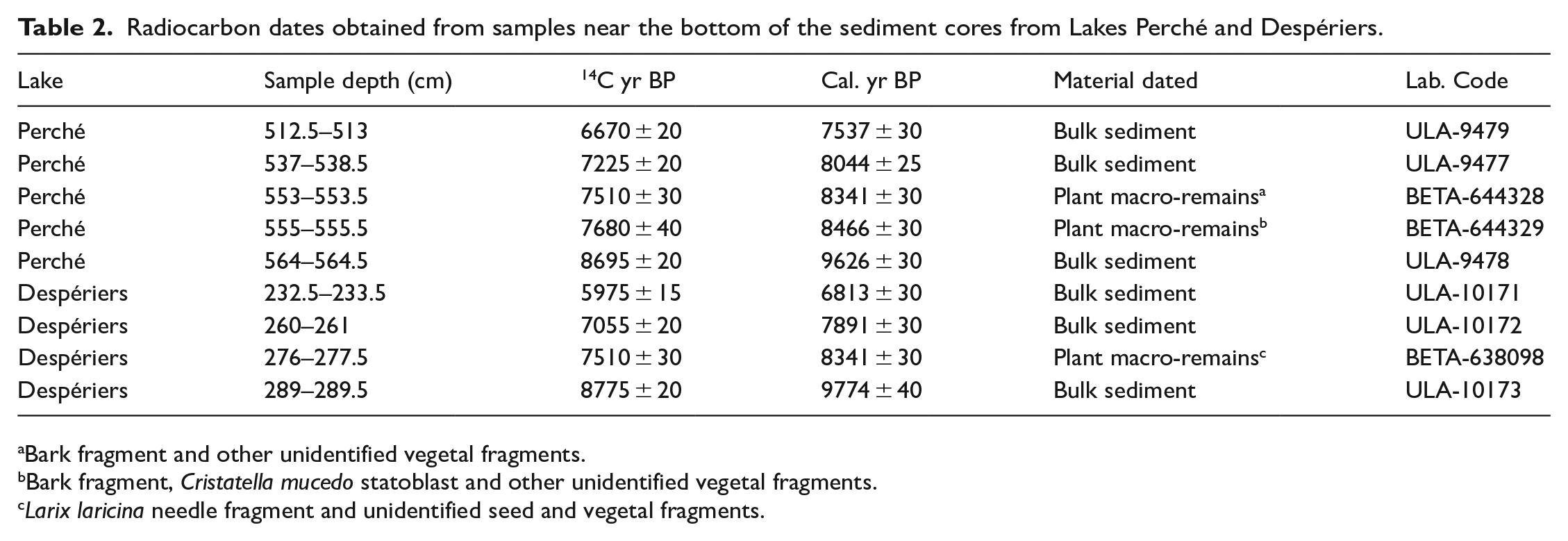
We built age-depth models with the Bayesian statistics-based “Bacon” v2.2 package in R (Blaauw and Christen, 2011) for the bottom part of the sediment sequences at each lake, as we wanted to focus on the period corresponding to the beginning of organic sediment accumulation on the paleo-islands. Despite our efforts, we found few macro-remains and we only managed to obtain 14C dates for two levels at lake Perché and one level at lake Despériers (Table 2). Hence, we also dated bulk sediment from three levels from each core to complete the age-depth models (Table 2). We calibrated the 14C dates with the IntCal20 calibration curves (Reimer et al., 2020).
Radiocarbon dates obtained from samples near the bottom of the sediment cores from Lakes Perché and Despériers.
Bark fragment and other unidentified vegetal fragments.
Bark fragment, Cristatella mucedo statoblast and other unidentified vegetal fragments.
Larix laricina needle fragment and unidentified seed and vegetal fragments.
Geochemical analyses
We performed XRF analyses on the basal sediments corresponding to the first millenium of the paleo-islands’ existence. To detect semi-quantitative variations of geochemical composition, we used the TestAllGeo analysis mode on the Niton XL3t 900SHE portable scanner and continuously scanned the sediments every 0.5 cm along the cores. Changes in the concentration of several major and minor elements can be used as proxies of ecological phenomena (Croudace and Rothwell, 2015). We used K, Ti, Rb, Sr, Zr, and Fe (lithogenic elements) as indicators of erosion from the watershed to the lakes, which we expected to decrease following afforestation of the paleo-islands. In addition to being an erosion indicator, low levels of K indicate extensive vegetation presence (Foerster et al., 2012). While S and Ca have various environmental interpretations (Croudace and Rothwell, 2015), we considered S to be indicative of organic matter presence and Ca to be indicative of biogenic carbonate (algae production). As plants fix sulfur (S), we used the S/Ti ratio as an indicator of organic matter presence (Moreno et al., 2007). The fact that S, Ca, and the S/Ti ratio are indicators of organic matter presence compensates for our inability to perform loss-on-ignition (LOI) analysis due to lack of material. For each lake, we applied principal component analysis (PCA) with the “factoextra” package in R (Kassambara, 2017) to differentiate portions of the sediment sequences based on their chemical signature. We used the Kaiser criterion to select significant principal components (PCs), that is, those with eigenvalues >1 (Kaiser, 1960). As variations along the PCs were semi-quantitative because of the XRF scanning method and scanner calibration, we could only compare trends in element dynamics (increase or decrease), and not their absolute values.
Identification of plant macro-remains
We bathed half of each 0.5 cm slice of basal sediments in a hexametaphosphate (Na6P6O18) solution for at least 12 h under agitation, for deflocculation purposes. We then filtered each sample through a 250 µm sieve to keep plant macro-remains. We identified plant macro-remains under a dissecting microscope using a published key (Lévesque et al., 1988) and recorded the presence and absence of taxa for each level studied.
Charcoal extraction and analysis
To perform charcoal extraction, we cut the basal sediments continuously into 0.5 -cm-thick slices. We extracted a 1-cm3 subsample from the middle of each slice to avoid contamination risk (Mustaphi and Pisaric, 2014). We bathed the subsample in a solution of potassium hydroxide (KOH), sodium hexametaphosphate (Na6P6O18), and sodium hypochlorite (NaClO) for at least 24 h under agitation to perform deflocculation and to help differentiate charcoal particles from organic matter (Whitlock and Larsen, 2001). We sieved the solution through a 160 µm mesh to extract macro-charcoal particles and measured their surface area under a dissecting microscope using the WinSEEDLE ™ image analysis software (Regents Instruments Inc., https://regent.qc.ca/, August 2021).
To reconstruct local fire history, that is, fire events that occurred within the lakes’ watersheds and thus on the paleo-islands, we applied the Charcoal Size Distribution (CSD) method (Asselin and Payette, 2005; Oris et al., 2014a; Remy et al., 2018) to every fire event detected using the CharAnalysis software v1.1 (Higuera et al., 2008; http://CharAnalysis.googlepages.com). We calculated geometric mean diameters of charcoal particles from the square root of the surface area (Clark and Hussey, 1996) and converted them to log mm. We sorted charcoal particles of each sample according to their size into six classes ranging from −0.9 log mm to 0.3 log mm (every 0.2 log mm). We applied a linear regression to the distribution of the proportion of charcoal particles in each size class (−0.8, −0.6, etc.) for each sample. As large charcoal particles travel less far, local fires are characterized by a greater proportion of large charcoal particles than regional fires, which is reflected by a gentler slope of the size distribution regression (Asselin and Payette, 2005). According to this principle, we classified as local fires (on the paleo-island) the samples with a slope more gentle than a threshold fixed at −1.77 based on a previous study (Oris et al., 2014a). In addition, we considered as local fires only the samples having at least one charcoal particle >−0.3 log mm (Supplemental Figure 1). We considered that other fires (extra-local or regional) did not burn on the watersheds of the paleo-islands.
Results
Organic sediment accumulation during Lake Ojibway presence
Basal 14C dates obtained from organic sediments at or close to the contact with the glacial mineral sediments were 9657 cal. yr BP and 9947 cal. yr BP for Lakes Perché and Despériers, respectively (Figure 3). These basal ages are older than the estimated date of Lake Ojibway’s final drainage (8200 cal. yr BP). Radiocarbon dates based on macro-remains that are coeval with the gradual lowering of the level of Lake Ojibway were recorded 10 cm above the basal dates obtained from bulk sediments. Despite being a little younger than expected, these radiocarbon dates based on macro-remains are the oldest dates obtained for the region. Moreover, they are from the Kinojévis phase (9000 to 8300 cal. yr BP), before the final drainage of Lake Ojibway. Radiocarbon dates based on bulk sediment and on plant macro-remains are consistent, suggesting that no or little reservoir effect altered the age-depth models and basal age estimations (Grimm et al., 2009).
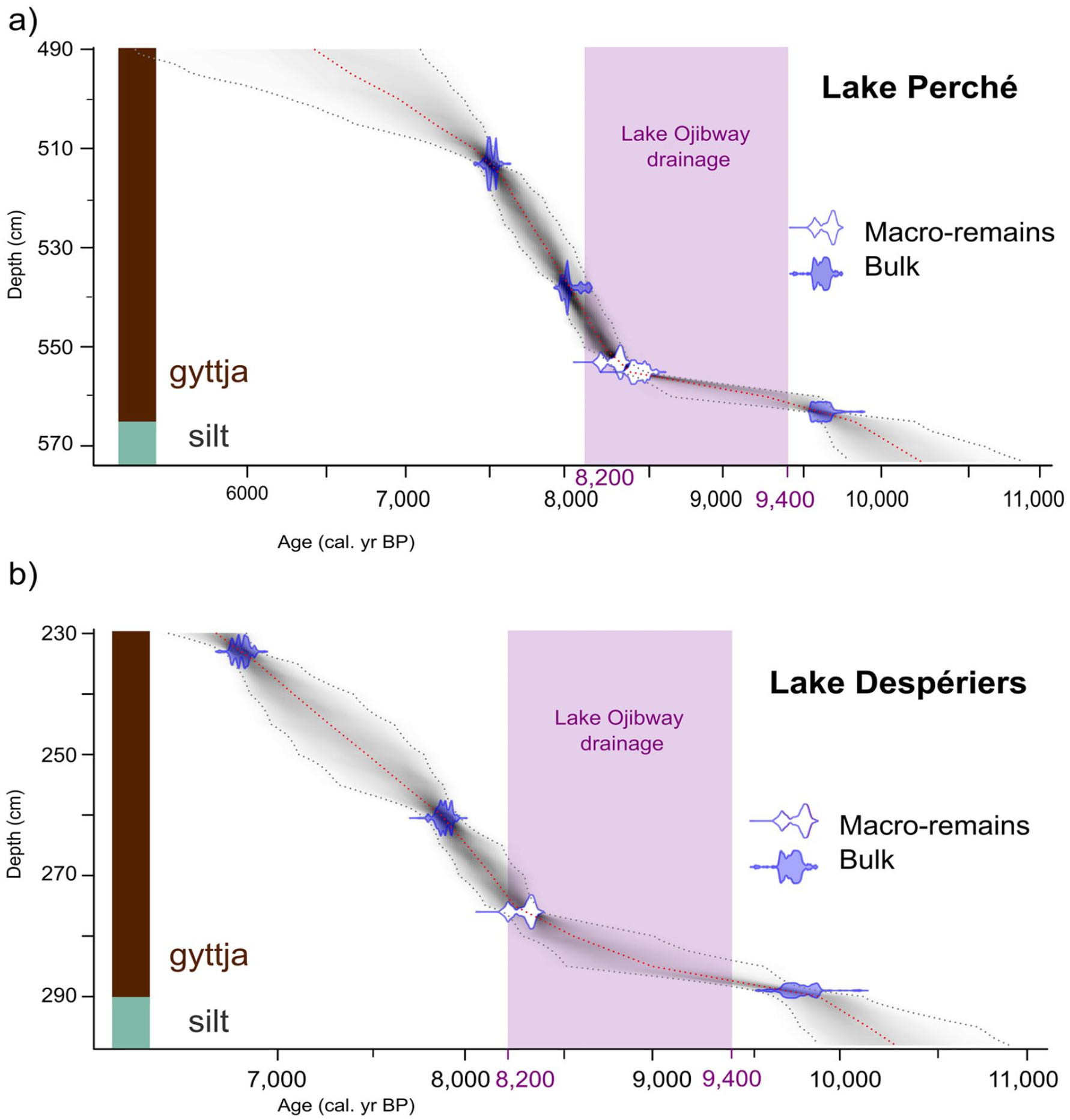
Calibrated age-depth models based on the radiocarbon dates (Table 2) obtained from basal sediments (brown = organic sediments and olive-gray = mineralogic sediments) at (a) Lake Perché and (b) Lake Despériers. The purple rectangle illustrates the period when the level of proglacial Lake Ojibway gradually decreased until its final drainage ca 8200 cal. yr BP.
At Lake Perché, the sediment sequence we analyzed spanned from 9657 to 6503 cal. yr BP, and at Lake Despériers from 9947 to 6850 cal. yr BP (Figure 3). The mean sedimentation rates were relatively similar at both lakes at the beginning of the Holocene (44 yr.cm−1 for Lake Perché and 53 yr.cm−1 for Lake Despériers). The mineral sediments found below the gyttja were olive-gray colored silt for both lakes, which was markedly different from the clay characterizing sites located below the shorelines of proglacial Lake Ojibway. Silt is indicative of glacial till associated to the direct action of the ice sheet on the bedrock without a proglacial lake intermediary (Boulton, 1978; Normandeau et al., 2013).
Geochemical signature of the organic sediments
Two groups were detected by the PCA analysis in the gyttja of Lake Perché: (1) from the silt-gyttja transition at 9600 cal. yr BP to 9000 cal. yr BP and (2) between 9000 and 6500 cal. yr BP (Figure 4a). The first two principal components explained 88.66% of the variability in the dataset (PC1 = 73.67% and PC2 = 14.99%). PC1 was positively associated to lithogenic elements (K, Ti, Fe, Rb, Zr, Sr) coming from the steep shores surrounding the lake, while PC2 was positively correlated to S. One element (Ca) was positively related to both PCs (Figure 4a).
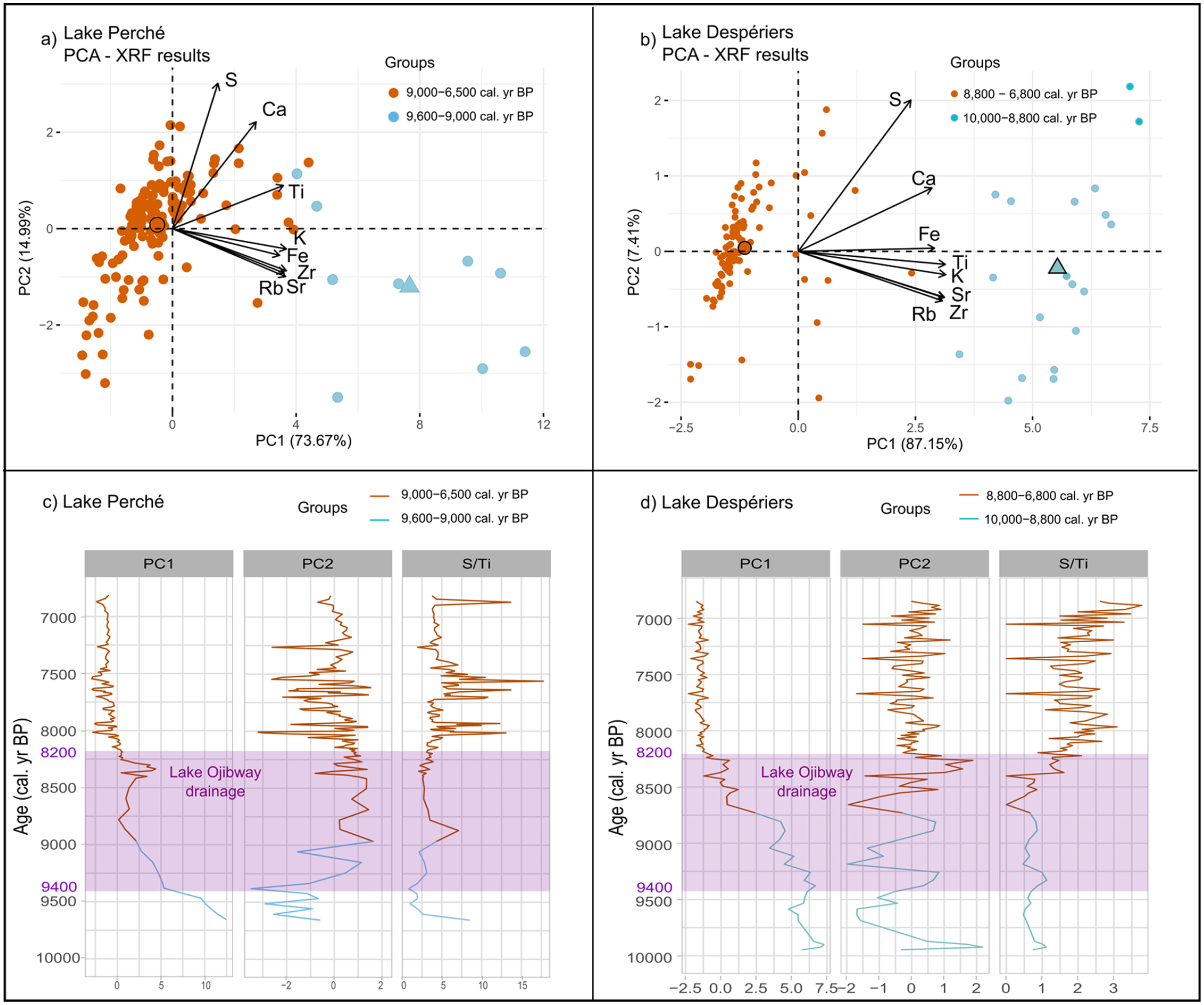
Principal component analysis (PCA) of sediment samples (dots), associated centroids for each cluster (larger dots and triangles), and XRF results (arrows) for (a) Lake Perché and (b) Lake Despériers. At both sites the sediment samples pre- and post-dating afforestation are clearly separated along PC1. Variations of PC1 and PC2 loadings, and of the S/Ti ratio across time at (c) Lake Perché and (d) Lake Despériers. The purple rectangle illustrates the period when the level of proglacial Lake Ojibway gradually decreased until its final drainage ca 8200 cal. yr BP. Note that the data for PC2 at Lake Despériers does not respect the Kaiser criterion of eigenvalue >1 and should be interpreted with caution.
The PCA also differentiated two groups in the gyttja of Lake Despériers: (1) from the silt-gyttja transition at 10,000 cal. yr BP to 8800 cal. yr BP and (2) between 8800 and 6800 cal. yr BP (Figure 4b). The first two principal components explained 94.56% of the variability (PC1 = 87.15% and PC2 = 7.41%). As was the case for Lake Perché, the lithogenic sediments were positively correlated to PC1 at Lake Despériers, S was positively associated to PC2, and Ca was positively related to both PCs.
For Lake Perché, PC1 values (associated with lithogenic elements) quickly decreased until 8800 cal. yr BP, and then increased until 8300 cal. yr BP. After 8250 cal. yr BP, values slowly decreased and reached their minimum until the end of the record, that is, 6500 cal. yr BP. PC2 values (associated with S) increased between 9350 and 9000 cal. yr BP (Figure 4a and c) and then remained relatively stable (despite short-term variations) between 9000 and 7900 cal. yr BP (Figure 4c). The S/Ti ratio slowly increased with peaks around 8800 cal. yr BP, between 8000 and 7800 cal. yr BP, between 7700 and 7500 cal. yr BP, and ca 6900 cal. yr BP (Figure 4c). The absence of a distinguishable pattern at 8200 cal. yr BP indicates that the environment remained stable during the drainage of proglacial Lake Ojibway.
For Lake Despériers, PC1 values (associated with lithogenic elements) gradually decreased from the beginning of gyttja accumulation around 10,000 cal. yr BP until 8700 cal. yr BP, then they strongly decreased around 8700 cal. yr BP to reach minimum values after 8200 cal. yr BP (Figure 4b and d). PC2 did not respect the Kaiser criterion (eigenvalue = 0.59) and therefore could not be interpreted (Supplemental Table 1). The S/Ti ratio remained stable during the early Holocene at Lake Despériers, with peaks at ca 9900, 9300 and 8800 cal. yr BP (Figure 4d). Then, ca 8400 cal. yr BP, the S/Ti ratio started to increase until 7900 cal. yr BP. Around 7700 cal. yr BP, the S/Ti ratio decreased and remained stable until 7000 cal. yr BP, then it increased until 6500 cal. yr BP (Figure 4d).
Plant macro-remains: Early presence of local trees
Despite the paucity of macro-remains in the sediments, we were able to extract and identify some of them. The earliest tree macro-remains that we found were larch (Larix laricina [Du Roi] K. Koch) needles dating from around 9513 cal. yr BP at Lake Perché (Figure 5a; Supplemental Figure 2a, b). Spruce (Picea spp.) needle fragments were also found ca 8385 cal. yr BP (Figure 5a; Supplemental Figure 2c). At Lake Despériers, the earliest tree macro-remains that we found were pine (Pinus spp.) needles, dating from around 9845 cal. yr BP (Figure 5b; Supplemental Figure 2d). Needles of spruce (Picea spp.) and larch (Larix laricina), as well as birch (Betula spp.) seeds were also found ca 8585–8300 cal. yr BP (Figure 5b; Supplemental Figure 2e–i). Trees were therefore present on the studied paleo-islands at least 1400 years before Lake Ojibway drainage.
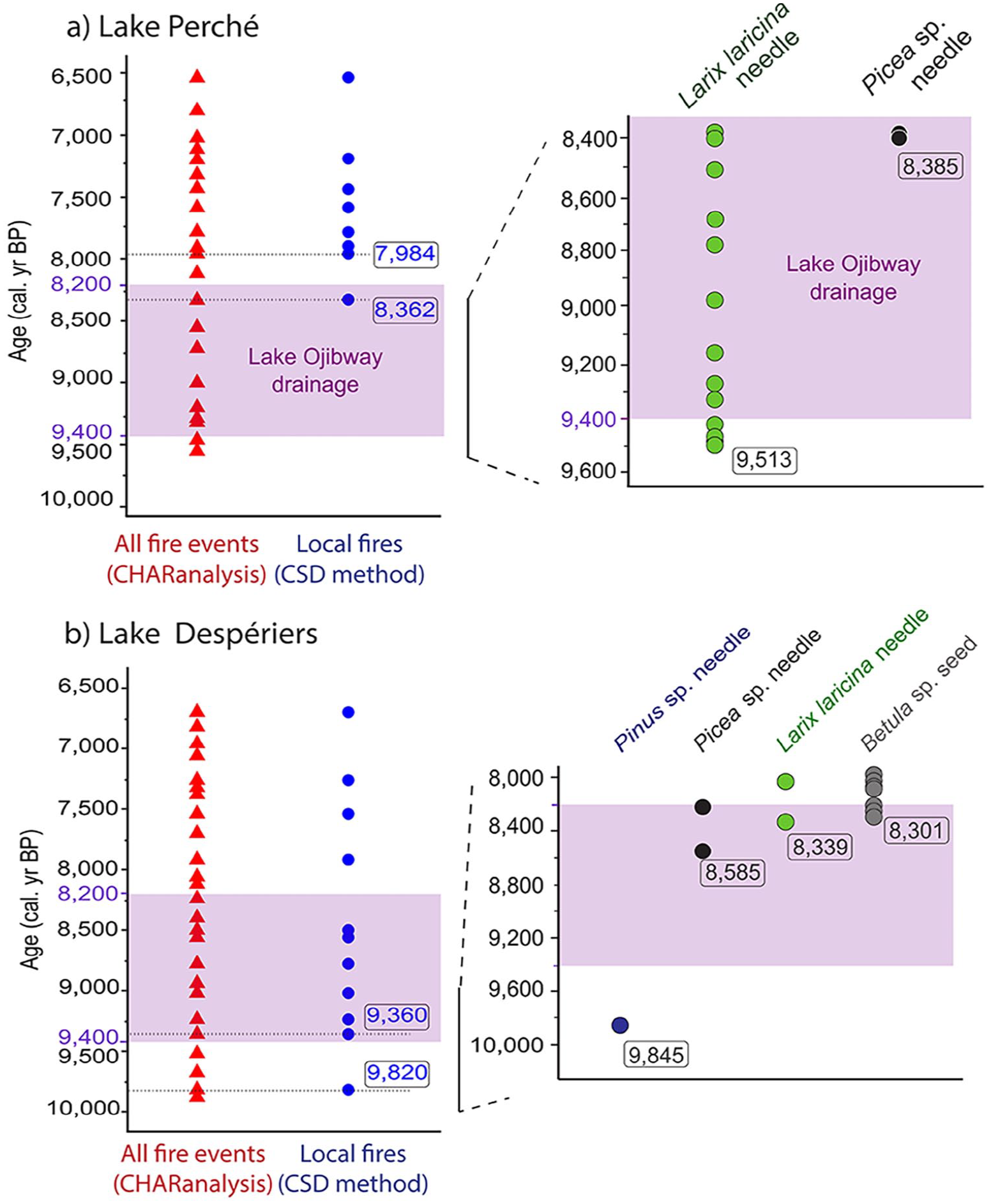
Fire events detected with CHARanalysis (red triangles), and local fire events detected with the Charcoal Size Distribution (CSD) method (blue circles) and oldest tree macro-remains retrieved from the sediments at (a) Lake Perché and (b) Lake Despériers according to the age-depth models (Figure 3). The purple rectangle illustrates the period when the level of proglacial Lake Ojibway gradually decreased until its final drainage ca 8200 cal. yr BP.
Evidence of local forest fires on a paleo-island
CHARanalysis detected 21 and 25 fire events in the Lake Perché and Lake Despériers sediments, respectively (Figure 5). At Lake Perché, eight events met the conditions to be considered local fires having burned within the watershed, the earliest local fires were dated at 8362 (Figure 5a) when charcoal influx was maximal (Supplemental Figure 3), 7984 and 7942 cal. yr BP (Figure 5a) when charcoal influx was lower (Supplemental Figure 3). The earliest local fire, evidenced by burned needle fragments (possibly Picea spp.) (Supplemental Figure 2c), occurred around 160 years before the drainage of proglacial Lake Ojibway. At Lake Despériers, 11 events met the conditions to be considered local fires having burned within the watershed. The earliest local fires at Lake Despériers were dated at 9820 and 9360 cal. yr BP, while Lake Ojibway was still present (Figure 5b).
Discussion
Our analyses of two sites located 50 km apart revealed that both paleo-islands were colonized by tree species several centuries before the level of Lake Ojibway lowered enough for paleo-islands to become connected with the mainland to the south ca 9000 cal. yr BP (Godbout et al., 2020; Roy et al., 2015). We used proxies suited to reveal local vegetation establishment, and we avoided proxies such as pollen, which can be influenced by extra-local vegetation inputs.
Early afforestation of islands on proglacial Lake Ojibway
Gyttja started to accumulate at the bottom of small lakes on paleo-islands while Lake Ojibway was still at its maximum level, that is, about 250–550 years before it started to gradually decrease. The presence of silt rather than clay beneath the gyttja at both sites is evidence that they were never flooded by proglacial Lake Ojibway. Radiocarbon dates obtained from plant macro-remains testify that shrubs and trees were present on the paleo-islands at least 140–260 years before the final drainage of proglacial Lake Ojibway (Table 2). According to the age-depth models, the oldest tree macro-remains we found were (indirectly) dated to ca 9600 cal. yr BP and 9850 cal. yr BP at Lakes Perché and Despériers, respectively, 200 and 450 years before the level of Lake Ojibway first started to decrease (Brouard et al., 2021; Godbout et al., 2020). Hence, while deglaciation time has been estimated at 10,570 cal. yr BP in the study area (Daubois et al., 2015; Godbout et al., 2020; Roy et al., 2015), our results indicate that the afforestation process needed about 1000 years to unfold after the glacier retreat. This is much longer than what has been documented at recently deglaciated sites in Alaska, where primary succession took less than 200 years to complete (Fastie, 1995). The difference would likely be due to the distance to seed sources having been much greater at our sites (80 km over mostly water) compared to those in Alaska (<30 km over ground). Furthermore, cold temperatures during the early Holocene (Viau et al., 2006) likely implied lower vegetation productivity, as evidenced in our age-depth models by lower sediment accumulation rates compared to the middle Holocene.
Geochemical analyses showed that lithogenic elements - therefore erosion (Joannin et al., 2022; Lefebvre et al., 2021) - decreased rapidly at both sites from the beginning of gyttja accumulation until 8800 cal. yr BP (Lake Perché) and 8700 cal. yr BP (Lake Despériers). As soil cohesion and stabilization are supported by plant root systems (Easterbrook, 1999), it is reasonable to assume that decreasing erosion is an indirect indicator of vegetation colonization on paleo-islands. Indeed, more individual plants produce more roots which stabilize the ground, meaning that less soil erosion is recorded in the lake sediments. Therefore, PC1 variations suggest the presence of vegetation rapidly after the beginning of gyttja accumulation at both lakes. The plateau PC1 values reached for both lakes around the final drainage of proglacial Lake Ojibway ca 8200 cal. yr BP likely indicates densification of the vegetation. Increased PC2 values (mostly linked to S fixation by plants) and a peak of the S/Ti ratio around 8800 cal. yr BP indicate a strong organic matter input and suggest the arrival (or densification) of local vegetation, coherent with the decrease of the level of Lake Ojibway. Therefore, the variations in the concentrations of lithogenic elements and elements indicating organic matter input into the lake sediments provide indirect support of early vegetation establishment before the level of Lake Ojibway started to decrease and possible densification on the paleo-islands before Lake Ojibway’s final drainage.
Macro-remains analysis confirmed local presence of trees at both sites, while Lake Ojibway was still present and before it gradually drained. According to the age-depth model, the earliest tree species identified at Lake Perché is Larix laricina, possibly indicative of wet conditions around the lake ca 9500 cal. yr BP (Johnston, 1990). It was followed by Picea spp. around 8400 cal. yr BP, at a time when geochemical analyses indicate that vegetation became denser. At Lake Despériers, Pinus spp. was the earliest species found at the paleo-island around 9850 cal. yr BP and could be indicative of environmental conditions drier than at Lake Perché (Rudolph and Laidly, 1990). Then, between ca 8585 cal. yr BP and 8300 cal. yr BP, Picea spp., Larix laricina and Betula spp. were also present, possibly indicating that environmental conditions had become more mesic, as at Lake Perché.
The fact that a local fire was dated at 8362 cal. yr BP at lake Perché (burned unidentified needle) is yet another confirmation of early afforestation of the paleo-island, as fire ignition and propagation are fuel-dependent (i.e. biomass-dependent) (Finney et al., 2021). At Lake Despériers, the earliest local fire around 9820 cal. yr BP (burned unidentified needle fragment, Fig. S.3.3) confirmed biomass presence and afforestation on the paleo-island (Finney et al., 2021) before the drainage of proglacial Lake Ojibway started.
The paleo-island colonization process: an answer to Reid’s Paradox
For primary succession to be successful on a bare island on a proglacial lake, several biotic and abiotic barriers must be overcome (Crofts and Brown, 2020). Climatic and meteorological factors are at play, substrate characteristics are also important, but distance to seed sources is the main hurdle.
At the time of full extension of proglacial lake Ojibway, the distance between the studied paleo-islands and the continental forest margin was about 80 km, with other islands located in between (Figure 2b). Colonization of the studied paleo-islands could have resulted from seeds coming directly from the colonization front and traveling almost 100 km above water. Such long distance dispersal events have been documented before (Davis et al., 1986). Seeds could also have come from other paleo-islands that could have acted as “stepping stones” (Harris et al., 2018). In either case, our results support the hypothesis of tree population outposts on paleo-islands, far from the main source population of propagules at the southern margin of the proglacial lake, as previously theorized (Davis et al., 1991). Even with the “stepping stones” scenario that would have reduced seed traveling distance to steps of ca 15–20 km, such dispersal would be considered very long, as the vast majority of boreal tree seeds are dispersed less than 100 m from the parent tree (Asselin et al., 2001; Burns and Honkala, 1990; Greene and Johnson, 1996). Estimating the seed rain at distances longer than a few 100 m is very difficult technically (Nathan, 2006). A modeling study estimated the proportion of seeds traveling 1–10 km (long distance dispersal events) at 0.1% (Higgins and Richardson, 1999). Knowing that yearly seed production of most boreal tree species reaches 1–5 million seeds/ha (Burns and Honkala, 1990), between 12,500 and 62,500 seeds could have reached the studied paleo-islands each year. Multiplied by 1000 years between deglaciation and vegetation establishment on paleo-islands and adding the probability of germination and survival, the possibility of colonization through long distance dispersal is, after all, not so unlikely. Moreover, as the level of Lake Ojibway gradually decreased from ca 9400 cal. yr BP, more and more land became available for colonization, which would have decreased the travel distance between mainland and islands, thus gradually increasing the probability of successful dispersal events and colonization of paleo-islands.
Catabatic winds blowing from the ice sheet to the south could have slowed down the migration process (Richard and Larouche, 1989). Reconstructions of paleo-wind directions indicate a dominant wind blowing toward the east or south-east (Veillette and Paradis, 1996), but do not indicate a predominance of catabatic winds (Nutz et al., 2020). Active (biotic) dispersal (e.g. by birds) could have contributed to long-distance dispersal, even if most boreal tree species are generally dispersed by wind (Higgins et al., 2003; Nathan et al., 2008; Wilkinson, 1997). Indeed, nonstandard dispersal of wind-dispersed seeds by omnivorous birds has been hypothesized and later confirmed by empirical data (Nogales et al., 2012; Rumeu et al., 2014). We do not have paleoecological proxies confirming or denying bird presence or passage on the paleo-island, but they could have facilitated tree seed dispersal. In the winter, secondary seed dispersal over snow or ice, or on ice rafts in the spring, could have also favored long-distance dispersal (Alsos et al., 2016; Greene and Johnson, 1997). Paleo-island plant colonization could also have occurred with seeds from the Lake Ojibway free water at Lake Perché, as it was located close to the proglacial lake’s shoreline. In the spring, melting snow would have flushed plant material (including seeds) into Lake Ojibway, and windrows would have facilitated their buoyancy and transport by wind up to the paleo-island shores (Glaser, 1981).
Simulation studies have estimated the migration rate of Picea spp. at about 50 m.yr−1 (Snell and Cowling, 2015; Sulphur et al., 2016), which would mean that trees would have needed ca 4000 years to migrate the 200 km from south to north of the area covered by proglacial Lake Ojibway following its final drainage around 8200 cal. yr BP, if it were not for paleo-islands. However, basal dates of bulk sediments sampled in lakes located at the northern end of the area yielded ages very close to drainage time (Ali et al., 2009; El-Guellab et al., 2015; Oris et al., 2014b). Our demonstration of early afforestation of two paleo-islands implies that they could have acted as migration outpost following the drainage of proglacial Lake Ojibway, hence accelerating colonization of the lowlands. The total area covered by paleo-islands represented ca 3600 km2, that is, about 10% of the area covered by proglacial Lake Ojibway at its maximal extent (Figure 2a). This means that several possible outposts were available for early afforestation, from near the colonization front in the south, up to the ice sheet margin in the north (Figure 2b). The contribution of paleo-islands as migration outposts is the most likely mechanism that could explain Reid’s paradox of rapid plant migration in western Quebec (Clark et al., 1998).
Supplemental Material
sj-docx-1-hol-10.1177_09596836231169988 – Supplemental material for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts
Supplemental material, sj-docx-1-hol-10.1177_09596836231169988 for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts by Marianne FS Vogel, Hugo Asselin, Sébastien Joannin, Yves Bergeron, Sabrina Leclercq, Cécile Latapy and Adam A Ali in The Holocene
Supplemental Material
sj-docx-2-hol-10.1177_09596836231169988 – Supplemental material for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts
Supplemental material, sj-docx-2-hol-10.1177_09596836231169988 for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts by Marianne FS Vogel, Hugo Asselin, Sébastien Joannin, Yves Bergeron, Sabrina Leclercq, Cécile Latapy and Adam A Ali in The Holocene
Supplemental Material
sj-docx-3-hol-10.1177_09596836231169988 – Supplemental material for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts
Supplemental material, sj-docx-3-hol-10.1177_09596836231169988 for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts by Marianne FS Vogel, Hugo Asselin, Sébastien Joannin, Yves Bergeron, Sabrina Leclercq, Cécile Latapy and Adam A Ali in The Holocene
Supplemental Material
sj-docx-4-hol-10.1177_09596836231169988 – Supplemental material for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts
Supplemental material, sj-docx-4-hol-10.1177_09596836231169988 for Early afforestation on islands of proglacial Lake Ojibway as evidence of post-glacial migration outposts by Marianne FS Vogel, Hugo Asselin, Sébastien Joannin, Yves Bergeron, Sabrina Leclercq, Cécile Latapy and Adam A Ali in The Holocene
Footnotes
Acknowledgements
We thank Thibaut Petry and his team at Aiguebelle National Park for their help and support. We also thank Mary Robles, Raynald Julien and Danielle Charron for their help with fieldwork and data processing. We are grateful to the Institut de recherche en mines et en environnement (UQAT) for letting us use their XRF scanner, and to Yvan Poirier and Mathieu Villeneuve for their help with the scanner. We thank Nils Ambec and Alayn Larouche for their help to identify plant macro-remains. We also thank Jean J. Veillette for constructive discussions, and Raphaël Chavardès, Dorian Gaboriau and the proofreading committee “Corrige-moi” who provided helpful comments on a first draft. We thank the anonymous reviewers, whose comments improve our manuscript. This is a contribution of the Laboratoire International de Recherche sur les Forêts Froides and ISEM contribution No 2023-049.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Mitacs Acceleration in partnership with Aiguebelle National Park [grant number IT25924].
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.