
Abstract
In early 2021 several tree stumps embedded in a remnant peat deposit were found in the intertidal zone of Wharton Beach, southwestern Australia by Esperance Tjaltjraak Rangers. Attempts were made to identify the tree using both dendrochronological and anthracological methods but were limited by the lack of reference information from tree species in the southwestern Australia. Radiocarbon dates from one stump indicates the tree lived for approximately 200 years, growing into the peaty sediments sometime before ca. 7340 cal yrs BP and died directly or indirectly as a result of marine transgression. The peat deposit is dated to 7608–7429 cal yrs BP but its exposed seaward edge was unconformably underlain by younger sand, dated by OSL to around 3550 years, which may have intruded as a result of a storm event. An OSL age of 12,600 years was obtained from the base of a laminated dune sequence behind the exposed peat deposit, and is significant given a similar age for a previously documented infant burial site in the area. Together these records provide a rare insight into the former wetland landscape and a cultural and scientific link to the drowned coastal plain. A need for more integrative research along this cultural corridor is clear but this preliminary study has demonstrated the value of combining Cultural Knowledge Systems and Western Science, for Tjaltjraak Healthy Country Program and analogous Aboriginal ‘caring for country’ programmes throughout Australia.
Introduction
Submerged cultural landscapes of Kepa Kurl
This project is situated within lands of the Wudjari Traditional Owners, centred on Kepa Kurl, Esperance, southwestern Australia (Figure 1). Kepa Kurl translates to ‘the place where water lies like a boomerang’. The Native Title Holders are represented by their corporation – Esperance Tjaltjraak Native Title Aboriginal Corporation. The Tjaltjraak, or blue-green Mallee (Eucalyptus pleurocarpa) is a significant cultural plant and adopted as the namesake for the Corporation.

(a) Kepa Kurl country, showing location of Wharton Beach just outside of Cape Le Grand National Park (map modified from McDonald, 2005, his Figure 1) and (b). Photo of Wharton Beach facing south, showing exposed peat with embedded tree stumps, unconformably overlying a conglomeratic siltstone.
Understanding the way landforms have been created and altered over time, and how people have shaped and adapted to change, is critical in all aspects of heritage and environmental management. In the Esperance region, the Tjaltjraak 1 Healthy Country Program is developing a multi-year, collaborative study to examine the ancient, inundated cultural landscapes of Kepa Kurl. These cultural landscapes are found in various forms – as submerged limestone reefs and ancient shorelines, islands that were once coastal headlands, cultural pathways that drowned rivers and ridges, and cultural features and landforms exposed in the intertidal zones of today. Many of these are or were extensions of modern-day estuaries, wetlands and lakes that remain an important component of Wudjari 2 cultural values and connections.
This study focuses on a set of tree stumps embedded in peat sediments exposed during the early summer of 2021 in the intertidal zone at Wharton Beach in Tjaltjraak Boodja Park (a Shire-managed Reserve) (Figure 1). The tree stumps and the peat were discovered by the Tjaltjraak Rangers during an On Country Patrol and have since been naturally reburied by sand. A short-term research programme was undertaken while the stumps were exposed, bringing together a team of Elders, rangers, heritage specialists and scientists from diverse fields with the aim of understanding the origin and formation history of this uncommon exposure.
Wharton Beach, Cape Le Grand NP
Geomorphology and offshore bathymetry
The Esperance coastline is characterised by arcuate sandy beaches located between exposed Proterozoic granite headlands, many over 300 m high (Sanderson et al., 2000). The 105 dome-shaped islands of the Recherche Archipelago resemble the granitic headlands of the mainland coast and, along with offshore seagrass meadows, protect the coast of Esperance from the high energy Southern Ocean. Within the Cape le Grand area, most of the islands are within the 50 m bathymetric contour, with some of the outer islands rising from as deep as 80 m or more (Australian Hydrographic charts). Outcrops of Tertiary limestone and Pleistocene calcarenite occur offshore, and some of the Proterozoic islands are capped by remnants of Tertiary limestone (James et al., 1994; Kendrick et al., 2005).
Wharton Beach borders Cape Le Grand National Park, which extends west towards the town of Esperance and contains some of the most spectacular coastal scenery in Australia. Here, as elsewhere in the Esperance region, the marine, beach and foredune deposits are composed of almost pure white silica sand derived from the surrounding granite rocks. The shelf profile in the region shows a general steepening towards the shoreline, aiding movement of sediments offshore and limiting the accumulation of shell and carbonate material on open beaches (Morgan and Peers, 1973). A recent bathymetric survey to 40 m water depth of the Esperance Bay shows an initial steep slope reaching 20 m water depth ~2 km from the shoreline then a shallower slope reaching ~40 m depth approximately 10 km offshore (Li et al., 1999).
Offshore rock platforms and drowned shorelines have been reported off steep islands and headlands at depths of 6–8, 11–13, 16–21, 27–36 and at 55–100 m below sea level, reflecting marine abrasion and lower eustatic sea-levels during the late Pleistocene (Carrigy and Fairbridge, 1954; Morgan and Peers, 1973; see also Ryan et al., 2008). With each of these drops in base level, rivers were rejuvenated resulting in the dissection of alluvial flood plains and forming gorges near the coasts. Based on marine studies of Esperance Bay, the inner shelf region would have been flooded between 12.5 and 7 kyr BP (Ryan et al., 2008). Rejuvenation ceased when sea levels stabilised around their present-day levels, and now active erosion of the coastline is taking place (Morgan and Peers, 1973).
Wharton Beach has a scarped foredune, backed by vegetated transgressive dunes that typically extend up to 10 km inland, driven by strong southwest winds. Small creeks in the coastal plain flow to small lakes and inter-dune swamps or lagoons near the coast. Some of these swamps may also be fed by springs, with coastal seepage evident along parts of this and other beaches in the region. Coastal seepage has also resulted in the development of organic-rich soils along contact zones between impervious granitic gneisses (Whinam and Hope, 2005). Further details on the geology of the region can be found in Fairbridge and Serventy (1954) and Morgan and Peers (1973).
Oceanography
Southwestern Western Australia features a temperate climate, with very dry summers (average annual evaporation is around 1600 mm), and cool wet winters (mean annual rainfall of 623 mm). Temperatures range from 16°C to 26°C in summer and 8°C to 17°C in winter. The relatively low rainfall coupled with the low-relief onshore topography, results in minimal fluvial discharge into coastal waters. Prevailing winds are SE in summer and SW in winter but with regular storm events from the NW (Sanderson et al., 2000).
Tides are semi-diurnal with a maximum spring tidal range of 1.1 m (Van Hazel et al., 2001). The Leeuwin Current maintains water temperatures near the coast above 13°C in winter and around 22°C in summer (Li et al., 1999), and interacts with upwelling cooler waters from the south. Localised tidal currents occur between some of the islands and in some longshore tidal channels but are generally less significant than wind-generated currents.
For most of the year the Recherche Archipelago is affected by strong, relatively consistent ocean swells from the southwest with big deep-water wave heights (>2.5 m), and long swell periods (>12 s), with wavelengths up to 200 m (Kendrick et al., 2005; Van Hazel et al., 2001). Swell can be reinforced by wind-generated waves, and produce a net eastward littoral drift along the south coast (Fisheries Western Australia, 2000). The waves, swell and currents of this region have a significant influence on the coastal geomorphology. Wharton Beach is exposed to persistent long southerly swell from the Southern Ocean, and comprises a transverse bar and rip system.
Cultural and historical context
The study area is part of the Greater Cheetup cultural landscape that is the spiritual landscape of the Wudjari, and active, ongoing component of the community’s cultural heritage and identity. Cheetup (place of birds) is a sacred granite dome and the location of an archaeological study revealing human occupation extending to at least 13,000 years BP (Smith, 1993). Today, the cultural practice that is ‘caring for country’ defines much of the role of the Tjaltjraak Rangers, in the footsteps of the ancestors. The team are actively managing this area, soon to be named ‘Tjaltjraak Boodja Park’ – and Tjaltjraak Rangers have been working on cultural surveys and mapping, archaeological projects, landcare projects, dune stabilisation and erosion control measures, and visitor management tasks in collaboration with local government agencies. A recent Tjaltjraak ranger-led archaeological heritage preservation project in this area described the holistic work of the Tjaltjraak Healthy Country Program, that involved archaeological open-air test excavations revealing human occupation in the immediate area extending to ca. 3800 yrs BP. 3
A variety of different types of cultural/historical sites are known, both ethnographic (not mentioned here) and archaeological, including burial sites, quarries, stone arrangements, rockshelters, stone-lined fish traps and extensive artefact scatters (Dortch and Morse, 1984; Guilfoyle et al., 2019; Smith, 1993). Archaeological surveys have been carried out across the exposed floors, shorelines and underwater areas in many parts of the southwest including Broke Inlet, Wilson Inlet, Oyster Harbour, inlets of the Fitzgerald River National Park, Torradup Estuary and Stokes Inlet (Dortch, 1995). According to Dortch (1995), several of these places have cultural evidence that pre-dates sea-level rise and the formation of these shallow estuarine basins, although absolute dating is still needed to confirm this. The process of marine transgression may negatively impact certain cultural features and places (Rowland and Ulm, 2012), at the same time offering unique preservation conditions in the study of human-environment relations.
The Esperance coast was first explored by the Dutch in 1627 and later the French in 1792 who both named many of the prominent coastal and island features. It was not settled by Europeans until the 1870s and while farming occupies the inland, most of the coast remains in a natural state, much in national parks. The Archipelago was first charted in detail by Matthew Flinders in 1802 (Ryan et al., 2008).
Methods
Field sampling
Field sampling was undertaken on 15th November 2021, during a period of low neap tides (low tide of 0.41 m at 15:07). There were five tree stumps exposed, with four of these clustered together on the eastern edge of the exposed peat. The top of one of the tree stumps was sawn off for wood identification and age dating analysis.
Attempts were made to trace the peat unit landward and alongshore using an auger. Landward, the peat unit appears to thin out towards the dunes, and alongshore there was no evidence of the peat extending beyond the area that was already exposed. The seaward extension of the peat unit has eroded, showing it to unconformably overlie an indurated, green conglomerate clay, in the top of which were embedded angular pebbles of red quartzite (Figure 2).

Cemented conglomeratic unit with embedded pebbles of angular quartzite and sub-rounded siltstone exposed in the surface where there is no overlying peat.
The black, peaty sediments in which the stumps were embedded was sampled for dating. A sharp spade was used to expose a section of the peat, which revealed two clear sediment layers – an upper unit of compacted black, clayey-sand and a lower unit of organic-rich, dark-grey, moderately sorted, medium–fine quartz sand with a fine-medium gravel component (<5%) (Figure 3). A 15 × 5 cm Kubiena tin was pushed into this profile for later micromorphological analysis (Figure 3).

Exposed peat showing an upper unit of fine black mud overlying a dark grey, well-sorted sand. The grey box shows the position of a 15 cm Kubiena sample taken for micromorphological analysis.
A sample of the peat at 5 cm depth was collected for radiocarbon dating. A sample of the dark grey sand was obtained for Optically Stimulated Luminescence (OSL) dating, using a steel tube hammered into the lower unit at 15 cm depth. Additional sediments were taken at the same depth to determine the background radiation, which is needed in OSL age estimates (Bateman, 2019). These deposits have only been exposed recently and have likely been buried by a metre of more of sand over the past few decades, and even more over the past millennia as dunes have migrated across the coastal plain. Thus, its current depth below surface likely does not reflect its average depth below the surface during its burial history.
The base of the foredunes were also exposed to reveal a series of laminated dark chocolate-brown, light brown and fawn-coloured sands, overlying a lighter grey sand (Figure 4). Multiple horizontal laminations in the dune profile indicate that the deposit is largely in situ, but the dune may have been higher in the past. The base of these sands was a cemented unit, which at a similar level, was assumed to be the same cemented conglomerate unit exposed below the peat. An OSL sample was taken at the base of the profile, again using a steel tube hammered into the sands with a depth estimated at 320 cm below the top of the dunes. A cross-section of the profile was collected in a PVC tube, split longitudinally, with additional samples collected through the profile for sediment characterisation. All samples were transport to the University of Western Australia (UWA) for subsequent analysis.

Photo (left) showing one of the tree stumps before it was cut, behind which are the eroded foredunes. Cutting back the base of these foredunes reveals a series of laminated grey, dark brown and fawn sands.
Wood identification and ageing
A ~2 cm thick cross-section (disc) was cut from the stump sample and prepared for wood anatomical and dendrochronological analysis to attempt to identify the tree species and estimate its age. The section was sanded using successively finer grit sandpaper (up to 1500 grit) to form a smooth surface, then soaked in acetone baths (two times) for 6–8 h each time to remove any resin prior to macroscopic and microscopic visual analysis (Figure 5). The wood anatomy was then visually examined under a stereo-microscope.

Samples were taken from the ‘long’ and ‘short’ side. The ‘inner’ sample is closest to the pith, the ‘mid’ sample is taken from about two-thirds of the way from the centre, and the ‘outer’ sample is taken from the outermost rings. Each site sampled is noted with yellow post-it on the above image. For each sample 1–3 rings were needed to collect sufficient material for a radiocarbon date.
A 4 cm sub-sample of the tree stump was prepared for anthracological examination by heating in foil to 400°C for 30 min. The sample was compared against a reference sample of modern Taxandria juniperina (provided by SH from west of Mt Melville, Albany) that was similarly prepared. Sample anatomy was examined from longitudinal and transverse sections of charcoal stubs using the Neoscope SEM housed at the Centre for Microscopy, Characterisation and Analysis (CMCA) at UWA.
Radiocarbon dating of wood
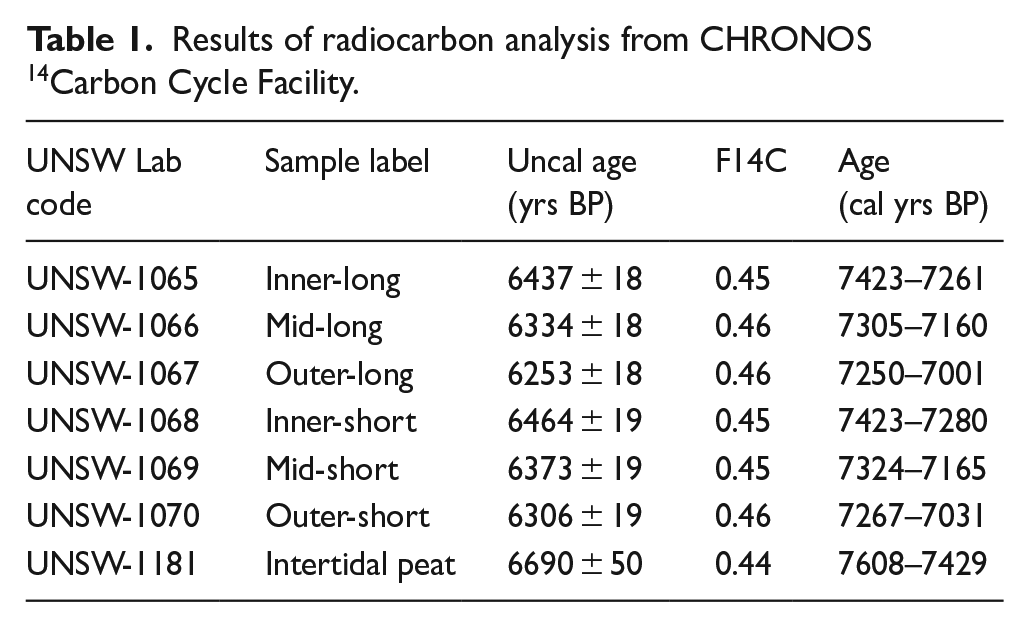
Wood samples were submitted to CHRONOS 14Carbon Cycle Facility at the University of New South Wales. A total of six samples were taken from the inner (closest to the missing pith) to the outer edges of the sectioned wood (Figure 6). A standard Base-Acid-Base-Acid-Bleach (BABAB) chemical treatment was used for all wood samples, whilst an Acid-Base-Acid was used for the peat sample. Full details of the analytical methods and AMS analysis is provided by Turney et al. (2021). To further constrain the age of the tree stump, given the known stratigraphic sequencing of the samples, Bayesian Sequence modelling was undertaken, with a specified interval (with error) between the samples. Final results were calibrated using OxCal using SHCal20 curve (Hogg et al., 2020).
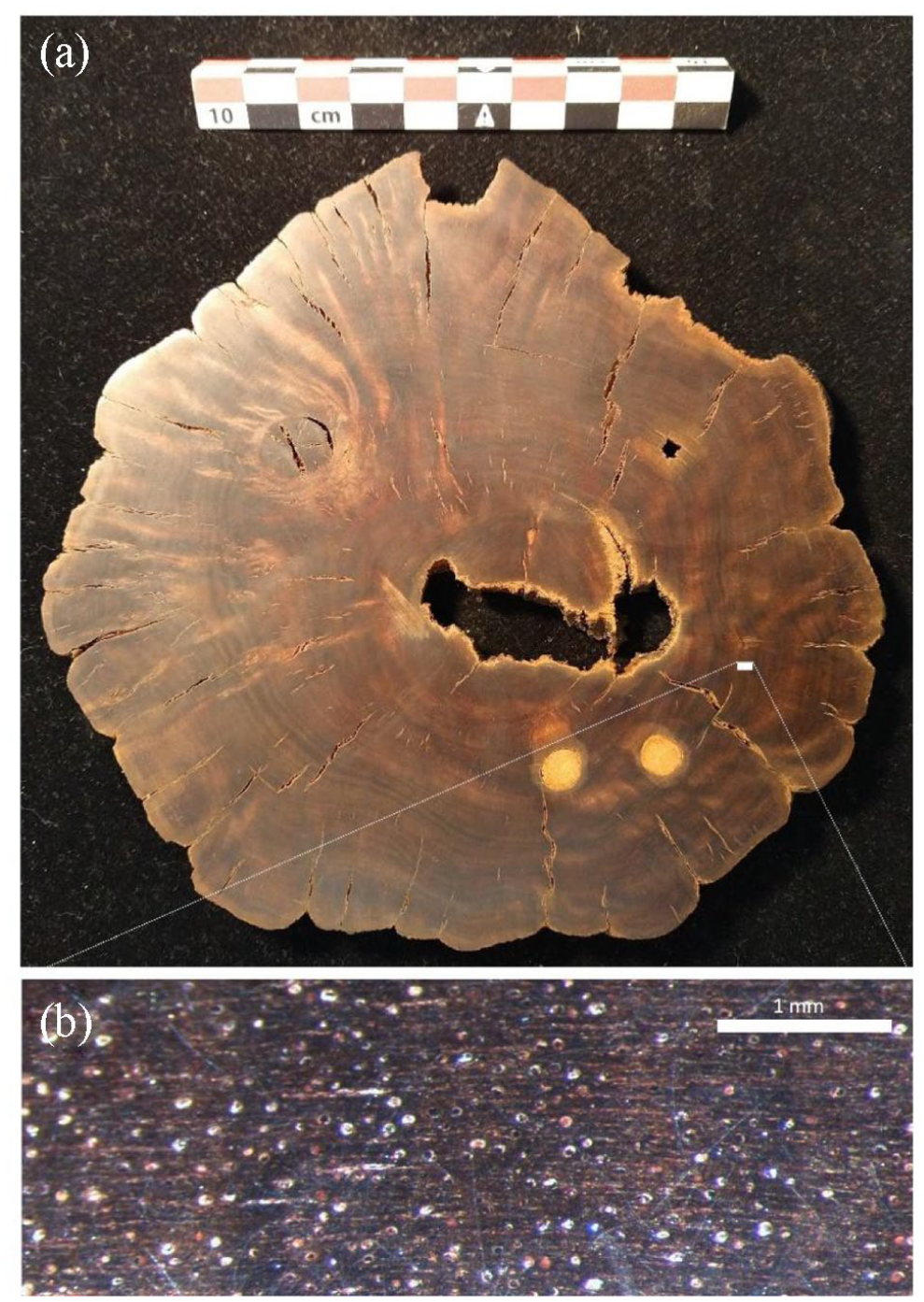
(a) Photograph of the sanded surface of the cross-section used for dendrochronological and anatomical analysis and (b) microscopic image (70×) of vessel (pore) arrangement of the stump sample. Vessels are sometimes partially or completely filled with sanding dust and appear as white circles.
OSL dating of sediments
Single-grain OSL dating was undertaken at Wollongong University, with a report provided by O’Gorman and Jacobs (2022). Both OSL samples – WH01 and WH02 – were prepared using routine optical dating procedures (Aitken, 1998) with grains of 180–212 μm in diameter used for dating. Single-grain measurements to determine the latent luminescence signal or equivalent dose (De) followed the single-aliquot regenerative-dose (SAR) procedure (Galbraith et al., 1999; Murray and Wintle, 2000). OSL measurements were made on an automated Risø TL-DA-20 luminescence reader, equipped with a focused green (532 nm) laser for single-grain stimulation (Bøtter-Jensen et al., 2003). Most quartz grains have inherent luminescence properties that may give rise to inaccurate estimates of De, which are identified through tests built into the measurement sequences. A total of 500 individual quartz grains were measured for each sample, with a total of 34 grains accepted for WH01 and 121 for WH02 (Table 2).
The total environmental dose rate consists of contributions from beta, gamma and cosmic radiation external to the grains, plus a small alpha dose rate due to the radioactive decay of uranium and thorium inclusions inside sand-sized grains of quartz. Beta dose was measured directly by low-level beta counting of dried, homogenised and powdered sediment samples in the laboratory, using a Risø GM-25-5 multi-counter system (Bøtter-Jensen and Mejdahl, 1988). Samples were prepared, measured and analysed following the procedures described and tested in Jacobs and Roberts (2015), with three sub-samples measured for each sample. Gamma dose rate from activities of radionuclides (U, Th and K) were measured by a combination of thick-source alpha counting (TSAC) and GM-25-5 beta counting. Radionuclide activities were converted to gamma dose rates using the conversion factors of Guerin et al. (2011) and corrected for the effect of moisture content (Nathan and Mauz, 2008).
The cosmic-ray dose rate was calculated following Prescott and Hutton (1994), and adjusted for water content (Readhead, 1987). The measured water contents of 17% ± 5% and 8% ± 2% were used to calculate the OSL age for WH01 and WH02, respectively. For WH01, the cosmic-ray dose rate was calculated based on time-averaged depths of 15 cm (i.e. the current depth below surface), 1 and 3 m to observe the impact of depth estimates on ages. The environmental dose rate data together with the total dose rates are provided in Table 2.
Sediment characterisation
The sediment block was air dried before resin impregnation, using a mix of polyester resin diluted with styrene at a ratio of seven parts resin to three parts styrene. The resin was left to cure over several weeks. A slice (<10 mm) was cut from the hardened block using a diamond rock saw, and used to make standard (25 × 76 mm) thin sections (30 µm thick). Thin sections observed using a Nikon polarising petrographic microscope available at the Department of Archaeology at UWA. Identification, description and semi-quantitative evaluation of abundance of various particles were done under plane-polarised light (PPL) and crossed-polarised light (XPL) using different magnifications (2×, 5×, 10×, 25×, 50×) following terminology outlined by Stoops (2003)
Particle size analysis (PSA) was undertaken on selected sediment samples from the peat profile and from the foredune sequence. PSA was undertaken on sieved (<2 mm) fractions of the bulk sediments using a Malvern Mastersizer 2000 (v. 5.6) housed at UWA. pH was also tested.
Pollen analysis
A sample of the peat was submitted to Morgan Goodall Palaeo (MG Palaeo) for pollen analysis, with analyses and full report provided by Macphail (2022). The modern reference pollen used for the analysis include representative specimens for many of the prominent families and genera and some species in the Western Australian flora but is not comprehensive regarding the range of pollen and spore (miospore) morphotypes found in large families such as the Ericaceae, Myrtaceae and Proteaceae.
Results
Wood identification
The stump sample is made up entirely of heartwood; sapwood has likely deteriorated over time (or it could be a very slow growing species). The heartwood is very dense and of a dark reddish-brown colour. Vessel elements are present, indicating that the stump is a hardwood (angiosperm). There are few vessels (i.e. <20/mm2), which are solitary, small (<100 μm) and randomly distributed. Given few southwest Australian species have comprehensive wood anatomical information available, the identity of the sample could not be verified from these features at this time.
The stump was given (by SH) a tentative identification of Taxandria juniperina (commonly known as watti), largely based on the presence of spreading buttress roots. Watti is a native species with prop roots generally confined to freshwater swamps and watercourses. Attempts to confirm the stump’s identification by anthracological (wood charcoal) observation against a modern reference sample of Taxandria juniperina were complicated by poor preservation of the waterlogged sample, which meant that some of the smaller features were not visibly preserved. The reference sample displayed radial clusters (of 3–6) in comparison to the distinctly solitary vessel arrangement in the stump charcoal sample. Whilst multi-seriate rays were present in both samples, they were comparatively longer in the Taxandria reference charcoal (Figure 7). The initial comparison shows these samples are likely different species. Through examination and comparison to UWA anthracology reference collection, the stump is Myrtaceae cf. Melaleuca. However, further comparative reference woods are required to confirm the stump’s identification and to advance to species level identification and rule out the potential of intraspecies variability.
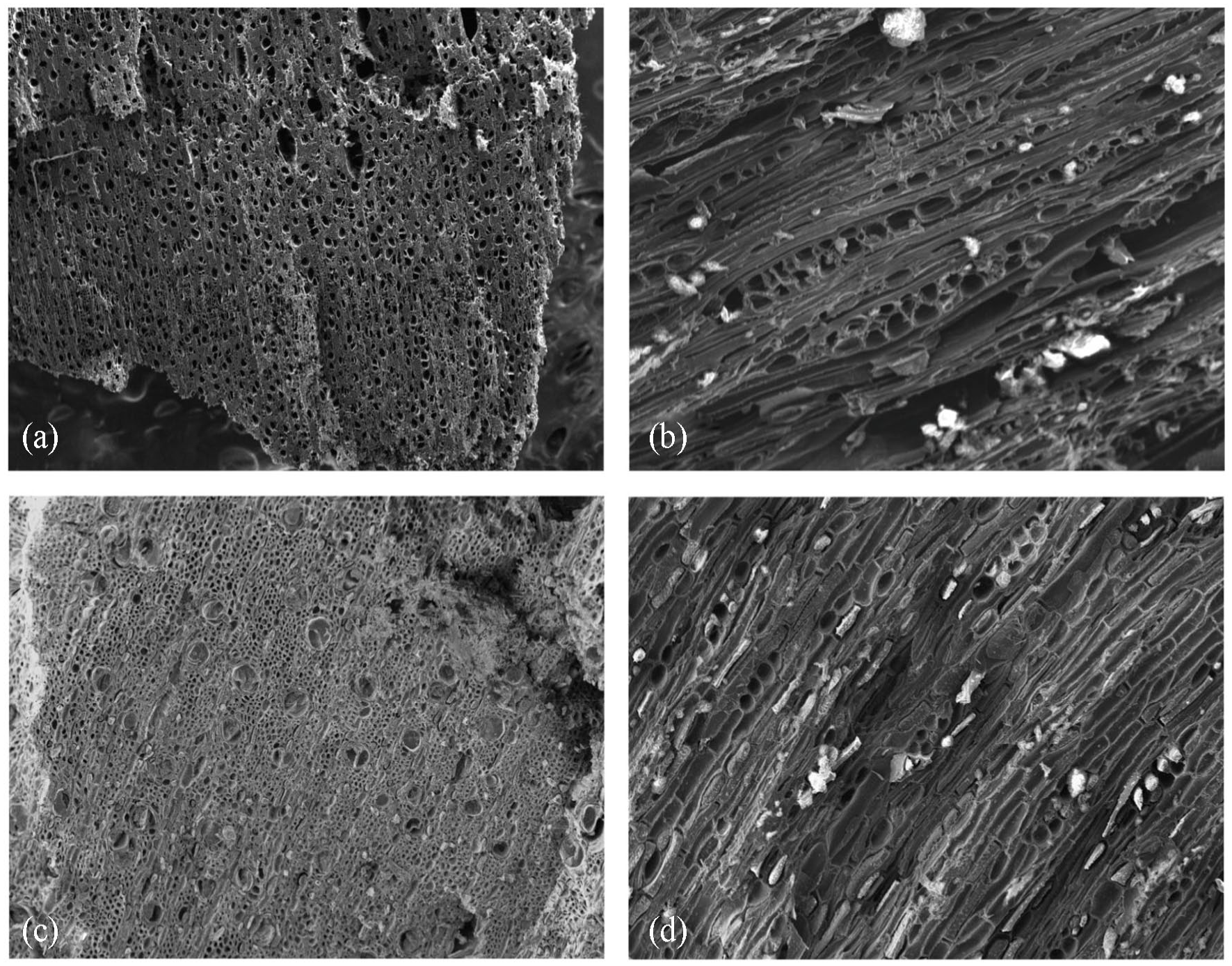
(TOP) Taxandria junipera reference: (a) transverse profile (×30) showing long radial clusters and (b) tangential longitudinal profile (×230) showing rays uni- to bi-seriate 5–15 tall, (BOTTOM) stump sample: (c) transverse profile (×110) showing solitary vessels and (d) tangential longitudinal profile (×170) showing rays uni- to bi-seriate 4–6 tall.
Wood and peat dating
On a macroscopic level, there are subtle variations in colour (wood density), which could be indicative of the earlywood and latewood of growth rings (Figure 6a). However, the wood is diffuse-porous, that is, the distribution and size of vessels are relatively even across earlywood and latewood of ‘growth rings’, hence, on a microscopic level, growth rings are indistinct and not well defined (Figure 6b). Consequently, the sample could not be aged using standard dendrochronological techniques alone.
As both the centre of the tree and the outer portion is missing from the stump sample, the dates provided by radiocarbon dating (Table 1) are estimates of the maximum (i.e. latest) establishment date and the minimum (i.e. earliest) death date. Radiocarbon dates indicate the tree lived for approximately 200 years; establishing before ca. 7340 cal yrs BP and dying after ca. 7160 cal yrs BP (Figure 8). The peaty sediment into which the tree grew was dated between 7608 and 7429 cal yrs BP (Table 1).
Results of radiocarbon analysis from CHRONOS 14Carbon Cycle Facility.
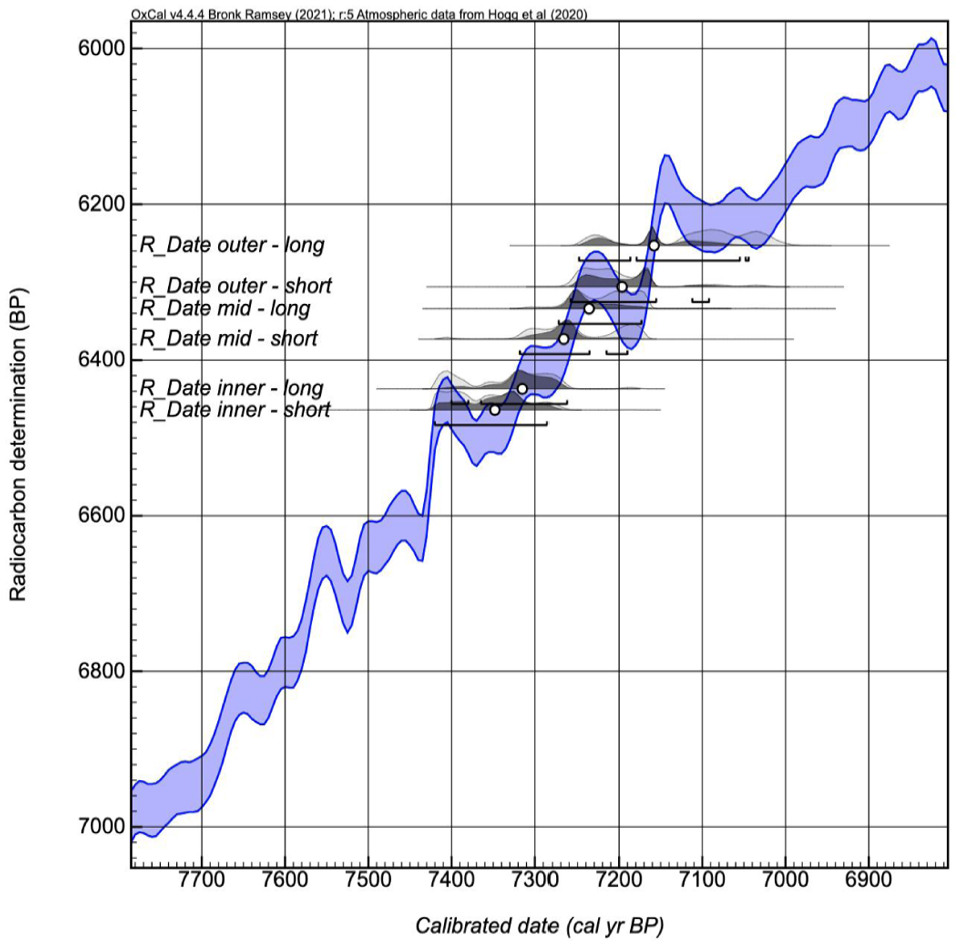
OxCal curve plot showing radiocarbon ages of the seven wood samples plotted on SHCal20 constrained by ‘Sequence’ model with intervals (as provided by CHRONOS 14Carbon Cycle Facility).
OSL dating of sediments
Full details of the OSL dating analysis are provided by O’Gorman and Jacobs (2022), with the main results reproduced here. The radial plots (Figure 9) indicate that the vast majority of grains from both samples have been well-bleached, with only a small number of grains (9%–13%) with either high or low De values that are inconsistent (shown as open circles in the radial plots) with the main component of De values, and resulting in high overdispersion (OD) values of 49% for each of the samples. These grains may have intruded post-depositionally. After the rejection of the outliers, the central age model (CAM) was then used to obtain the final De value.
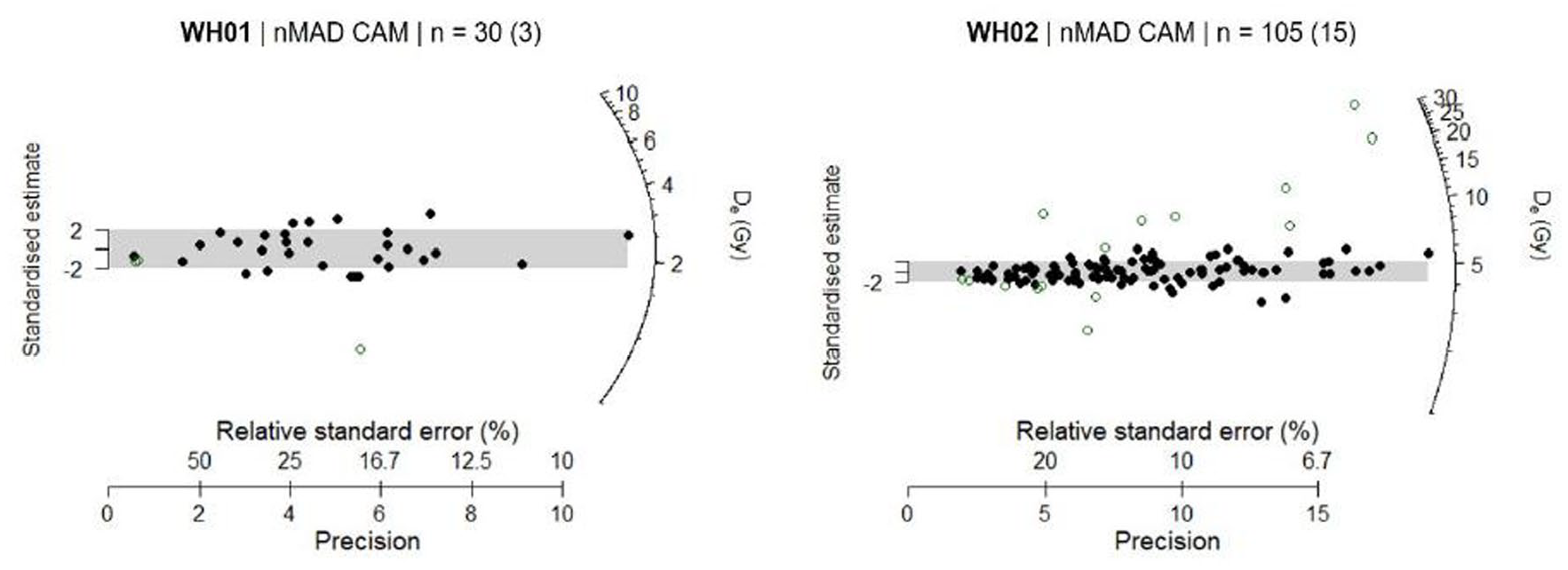
Radial plots for the two samples measured in this study, as provided by Wollongong University. The grey band is centred on the De value determined using a nMAD CAM statistical model. Outliers identified using nMAD are shown as open circles; the numbers of grains identified as outliers are shown in parentheses.
The final OSL age estimates are listed in Table 2, together with the supporting De and dose rate estimates. Uncertainties on the age estimates are given at 1σ (the standard error on the mean) and were estimated by combining, in quadrature, all known and estimated sources of random and systematic error. For the medium–fine quartz sand overlain by the peaty clay (WH01), ages of 3308 ± 335, 3583 ± 353 and 3758 ± 365 years ago were obtained for time-averaged depths of 15 cm, 1 m and 3 m, respectively. These age estimates are all consistent at 2σ, showing that the age estimates (average age = 3550 years ago) for this sample are insensitive to depth. A much older age of 12,597 ± 1194 year ago was obtained for the fine quartz sand overlying the peaty clay (WH02), assuming a time-averaged depth of 320 cm.
Dose rates, equivalent dose (De), overdispersion (OD) values, and ages as calculated using the CAM model.
Sediment characterisation
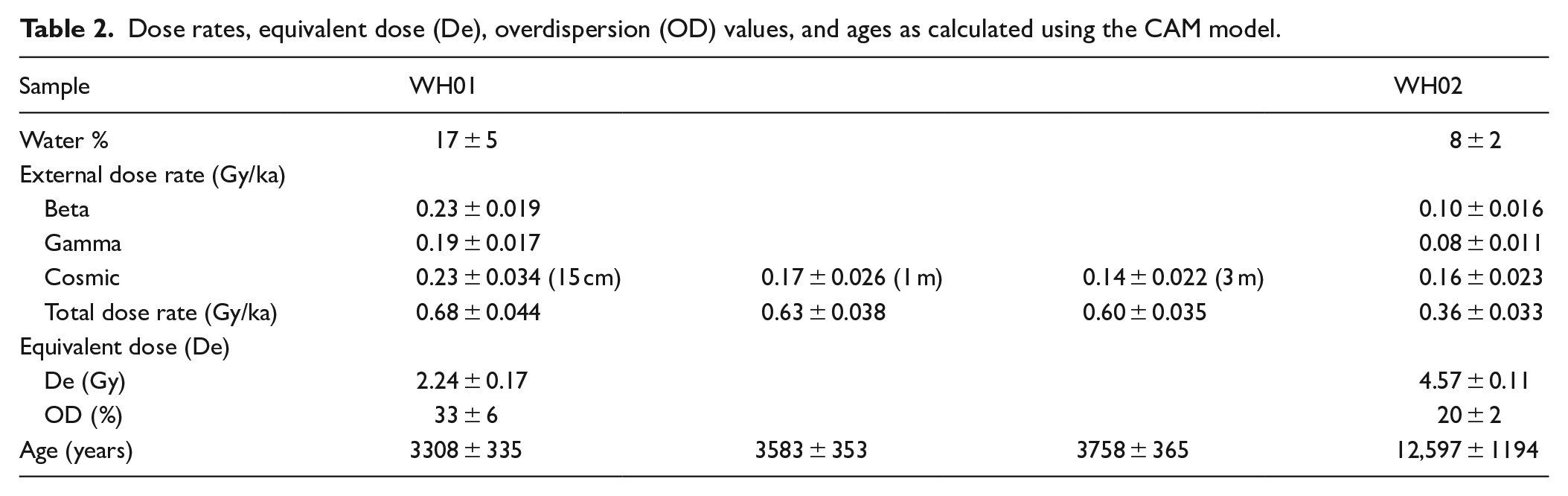
PSA of sediments (<2 mm) from the peat profile indicates sediments are coarsely skewed with four main modes including a (minor) very coarse sand (1200 µm), a slightly more dominant medium sand (240–275 µm), a coarse silt (30–40 µm) and a very fine silt (<10 µm) (Figure 10). The relative volume of coarse sand mode and very fine silts are greater at the base and top of the peat profile, respectively. Under microscope, individual quartz grains from the organic peat are sub-round and comprise a mixture of shiny and matt grains, whereas those from the base of the peat unit are sub-angular and matt (Figure 10). The organic-rich component at the top of the profile is highly acidic at pH 4.5 whilst the base of the profile is more alkaline at pH 6.5.
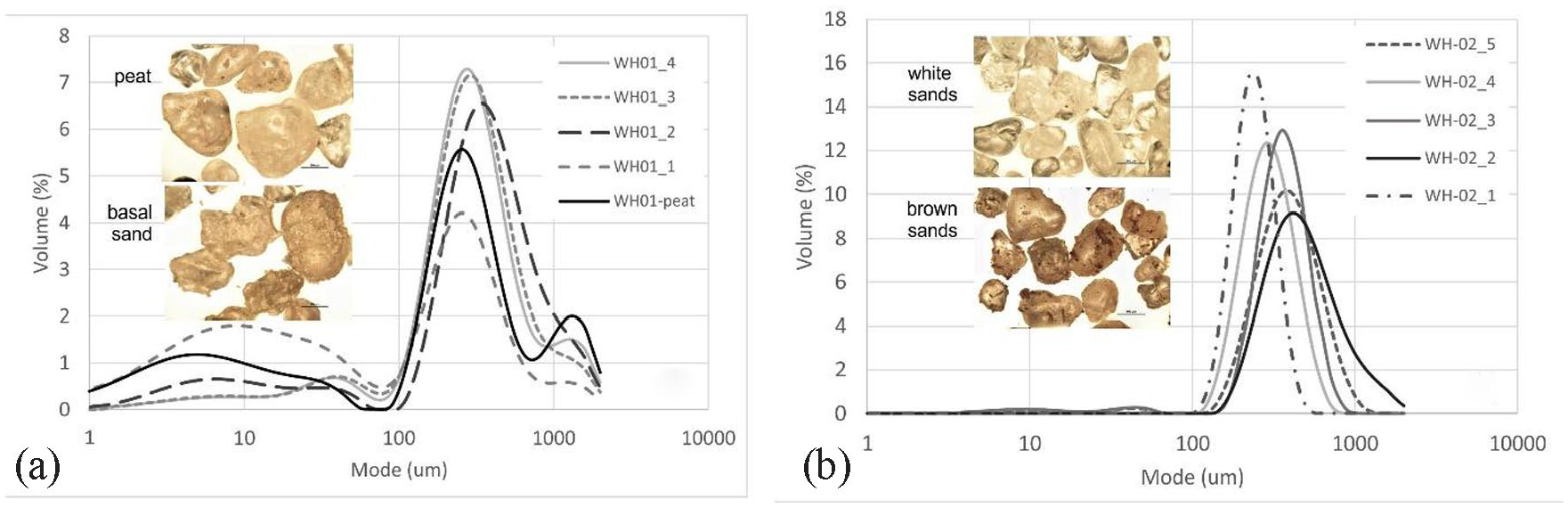
Particle size analysis (<2 mm) of sediments from (a) peat profile and (b) dune. Inset photos show quartz grains under the microscope (scale bar is 100 µm).
PSA of the dunes behind the peat exposure indicates the sediments are moderately well-sorted and bimodal with a dominant medium sand (270–395 µm) mode and a minor coarse silt (~ 41 µm) mode (Figure 10). The top of the dune is made up of slightly finer (225 µm), more well-sorted sands. This sorting is also visible under the microscope with a mixture of matt and shiny sub-round quartz grains, with those from the darker layers showing adhering organic material (Figure 9).
Micromorphological characterisation reveals the peat unit to contain plant material including rare charcoal fragments (Figure 11a), humified plant fragments (Figure 11b), unidentified stem fragments (Figure 11c and d), and abundant phytoliths, pollen (Figure 11e–h) and/or possibly foraminifera (Figure 11e and f, see also 12c). Further analyses are ongoing to identify the phytolith and pollen material. In addition to the plant matter, glauconite (Figure 11i) and pyrite (Figure 11j) are also relatively common. These secondary minerals form under anoxic conditions, with glauconite commonly found in nearshore sands and shallow marine environments.

Micromorphological images from the peat profile, showing: (a) charcoal fragment, (b) humified plant fragment, (c and d) cross-section of a plant stem(?), (e and f) either tetrad pollen grain (possibly Proteaceae) or fungal spore tetrad, (g) unidentified bisaccate structure, (h) single-cell microfaunal grain or foraminifera, (i) glauconite, and (j) pyrite framboids surrounded by rounded quartz grains. Scale bar is in microns.
Pollen analysis
Pollen analysis indicates the organic peat had very high abundance of aquatic algal cysts and zygospores (402%) relative to the terrestrial fossil pollen count. The former comprise aquatic algal cysts including Debarya (12%), Zygnema (42%, Figure 12a) and the green alga Botryococcus (3%, Figure 12b), and zygospores of unidentified organisms including rare trochospiral liners of brackish-saline foraminifera (~16%, Figure 12c). Other well-preserved plant remains including leaf cuticle and carbonised xylem (Macphail, 2022).
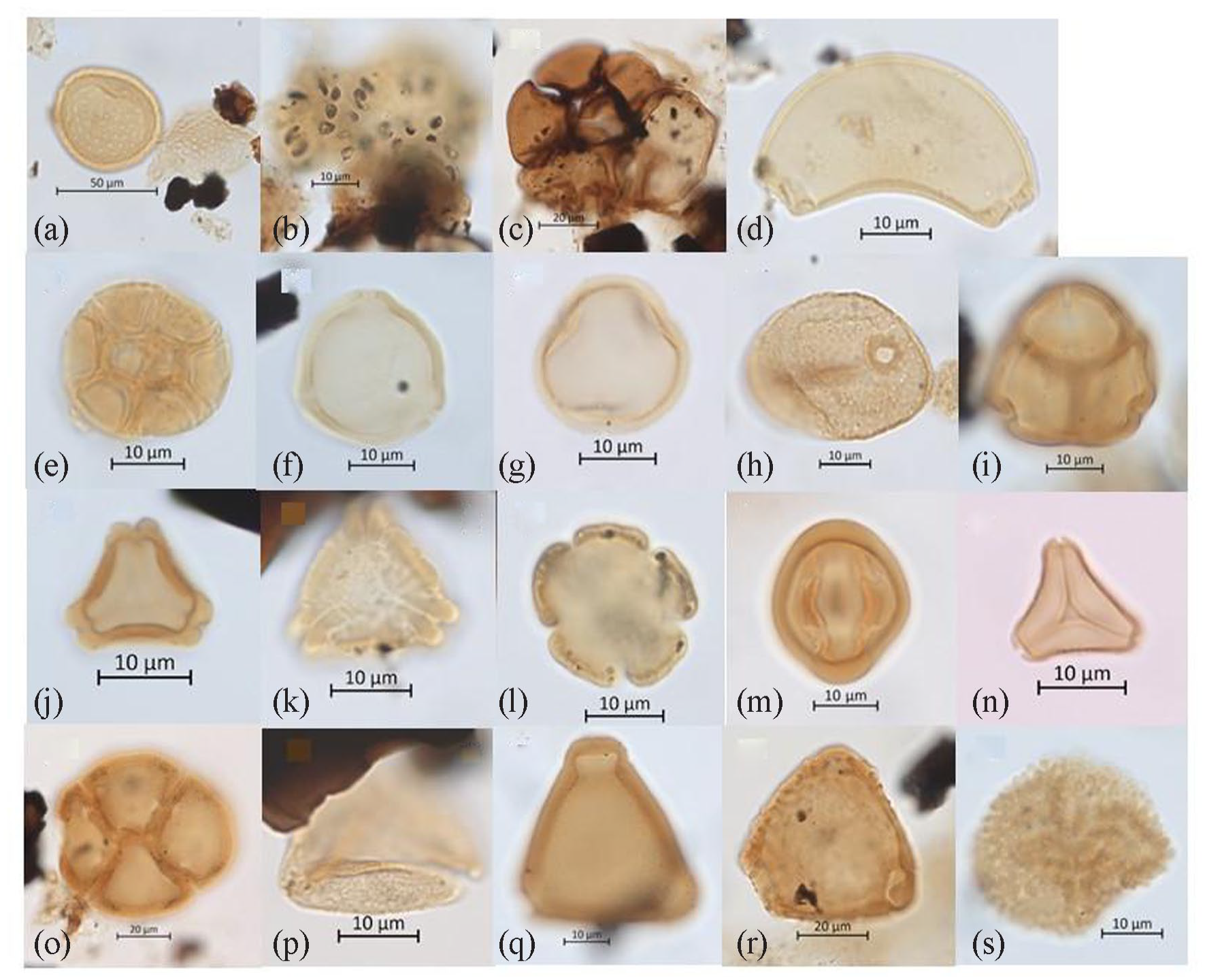
Images of better-preserved environmental indicator zygospores, foraminifera and pollen, including: (a) Zygnemataceae (Zygnema), (b) Botryococcus, (c) trochospiral liners of unidentified foraminifera, (d) Banksia speciosa-type, (e) Acacia, (f) Allocasuarina/Casuarina, (g) Coprosma-type, (h) Anarthriaceae, (i) Ericaceae, (j) Eucalyptus (sensu lato), (k) Eucalyptus spathulata, (l) Galium-type, (m) Gyrostemonaceae, (n) Myrtaceae (Leptospermoidae), (o) Orchidaceae?, (p) cf. Cupaniopsis (Sapindaceae tribe Cupanieae), (q) Proteaceae (Grevillea-type), (r) Proteaceae (Hakea-type), and (s) Ricinocarpos-type.
The diverse fossil pollen and spore assemblage (terrestrial microflora) is dominated by pollen of woody shrubs, in particular unidentified genera of the heath family Ericaceae (42%, Figure 12i) and Allocasuarina/Casuarina (16%, Figure 12f) with frequent Myrtaceae including Eucalyptus (4%, Figure 12j) and diverse Leptospermoidae (7%, Figure 12n), Proteaceae [8%, including Banksia speciosa-type (2%), Grevillea, Hakea and Petrophile types] and Asteraceae (2%). All families and genera are common in the heath flora of the Esperance coast, with Banksia speciosa documented (by SH) on the access track to Wharton Beach, a few hundred metres from the study site.
Several uncommon to rare taxa represent genera and/or species that are restricted to southwest Australia were also identified. Examples include (i) Gyrostemonaceae (Figure 12m) whose possible sources are the tall to under-shrubs Gyrostemon and Cypselocarpus, which occur in the general area and whose pollen are widely transported by wind, and (ii) Eucalyptus spathulata (Figure 12k), which, like T. juniperina, does not appear to be recorded in extant coastal vegetation in the Esperance region although its pollen is routinely transported by water into southwest estuaries (Macphail, 1998, 2007). Unidentified Proteaceae pollen types may represent Stirlingia, with this and other taxa in the Wharton Beach microflora close to the edge of their eastern range, whilst the cord-rush family Anarthriaceae (Figure 12h) is mainly found in swamps and open wetland communities around the coast (Macphail, 2022).
Other rare taxa can be identified with confidence to a plant family but not to the genus level because of the large number of possible affinities for example, Coprosma-type (Rubiaceae), Galium-type (Lamiaceae), Leucopogon-type (Ericaceae) and Ricinocarpos-type (Euphorbiaceae, Figure 12s). Plants known to produce similar pollen morphotypes to Galium and Coprosma, such as Pomax and Lycopus, do not occur in the local flora. A large planar tetrad is provisionally identified as orchid pollen (Figure 12o), which is notable given pollen in the family Orchidaceae are seldom recorded as microfossils. The occurrence of Sapindaceae tribe Cupanieae pollen resembling Cupaniopsis (Figure 12p) is difficult to explain as the only sources extant in the state are restricted to the northwest coast (Macphail, 2022), with possible ‘contamination’ from migrating birds one possibility.
Discussion
Drowned forests
Whilst marine submerged forests are relatively well documented around the coasts of England, Wales, France, Denmark and other parts of Europe (Campbell and Baxter, 1979; Reid, 1913); their occurrence in Australia is more exceptional. There have been reports of intertidal or subtidal tree stumps from localities along the southwest coast, including Broke Inlet (Eucalyptus marginata), Wilson Inlet (Melaleuca sp.), Stokes Inlet (M. cuticularis) and in the surf-zone off Trigelow Beach 300 km to the west of Esperance (Dortch, 1995). The exposure of the peat-embedded tree stumps at Wharton Beach in late 2021 offered a rare opportunity to gain information relating to past vegetation and environments during the early Mid-Holocene (Whinam and Hope, 2005; see also Macphail, 1973). Both the tree stump and the peat were important.
While the wood anatomy of the tree could be clearly observed and described, a lack of reference information of wood anatomy of Australian tree species, particularly from the south coast of Western Australia, limited the ability to identify the species from wood anatomical features. Similarly, the modern pollen herbarium is limited, particularly for large families such as the Ericaceae, Myrtaceae and Proteaceae (Macphail, 2022). Collection and description of the wood anatomy of modern specimens of tree species in the south coast region could help to identify the species of this sample but was beyond the scope of this study. The degraded nature of the wood also limited identification via anthracological analysis. The nature of the remnant prop roots hinted towards a possible identification of T. juniperina, a species generally confined to margins of winter-wet or permanent freshwater swamps and watercourses. The location of the stumps at Wharton Beach is a long way east from extant populations nearer Albany, and a positive identification would have implied a more extensive distribution of these trees in the past. However, comparison with a modern anthracological sample of T. juniperina indicated this identification was unlikely and an exact species identification has yet to be determined.
Radiocarbon dating indicates the tree likely started growing into the peaty sediments sometime before ca. 7340 cal yrs BP and died sometime after 7160 cal yrs BP. These results are complemented by the calibrated age of the sediments of 7608–7429 cal yrs BP, indicating that the sediments had started to accumulate before the tree started growing. There are fewer than 15 sea-level data points for the whole of the southwestern and southern Western Australian region (Baker et al., 2005), with the most recent curves for Western Australia constructed mainly using data from further north (Lewis et al., 2013). Combing the latter with the available dates and inundation record for Esperance Bay (Ryan et al., 2008), indicates that at the time the tree started growing, sea level was probably around 2 m below present (Figure 13). The trees may have initially been protected by a beach barrier that was later breached although this has yet to be resolved.
The age estimates for the stump at Wharton Beach are similar to those recorded for the tree stumps in the floors of Broke, Wilson and Stokes estuaries with five available dates ranging between 7387 ± 74 cal BP (Wk3295) and 8091 ± 195 cal BP (WAIT43) (Dortch, 1995). This indicates a more widespread event of forest inundation, which likely relates to sea-level rise (Figure 13). The closest published analogue to the Wharton Beach site is an organic deposit in the small embayment of Fingal Bay near Newcastle on the Central Coast of NSW (Macphail, 1973). The peat deposit at Fingal Bay includes tree trunks in growth position but differs from Wharton Beach in that the peat outcrops from ~ 0.9 m above the estimated mean high-water mark (HWM) to below LWM, with ages for the organic mud ranging between 3300 and 6500 cal yrs BP. However, a Melaleuca stump at RL -1.8 m also has a similar radiocarbon age of 7353 ± 150 cal yrs BP. The depositional context here is of a coastal swamp impounded behind a probable Pleistocene sand barrier is more or less identical to that inferred for Wharton Beach.
Palaeoenvironmental interpretation
The peat
The Wharton peat deposits unconformably overlie an undated consolidated conglomeratic siltstone (Figure 1). From around 80,000 years to the terminal Pleistocene, sea level was essentially below the shelf edge and so the area of Cape Le Grand was terrestrial. The rejuvenation of rivers in the coastal region during this period of lower sea levels would have resulted in dissection of flood plains across the inner shelf (Myers, 1995). The green conglomeratic siltstone may represent one such alluvial deposit, with the green colour indicative of anoxic conditions and the horizontal surface and fine silt indicative of low energy floodplain, into which later were embedded higher energy colluvial material including locally-derived angular quartzite pebbles. The absence of carbonate indicates the deposit was not formed during the higher sea levels of the last interglacial but could have formed anytime in the last 80,000 years.
The microfloral evidence indicates that the peat had accumulated in a wet environment where fluctuating water table levels allowed the in situ establishment of low to tall shrubs, or possibly tree-sized individuals growing. The paucity of eucalypt pollen is against the microflora representing mallee-heath and the low relative abundances of cord-rushes (Anarthriaceae, Restionaceae) and sedges (Cyperaceae) are against the plant community including extensive areas of low/open vegetation. Allocasuarina and/or Casuarina spp. may have been growing around the peat swamp, although this is complicated by the fact that pollen of the Casuarinaceae are produced and dispersed in astronomical numbers. Similarly, it is uncertain whether the palaeo-community was kwongan (defined by ericoid shrubs <1 m tall, Beard, 1990: 312; see also Hopper, 2014) or a taller form of coastal heath.
The possible presence of orchids pollen polyads is also of interest. Orchids are generally salt-intolerant but around the granite inselbergs adjacent Wharton Beach, the coastal sun orchid (Thelymitra granitora) and common mignonette orchid (Microtis media) have each been located (by SH) within 100 m of the shoreline where they would be exposed to salt spray in storms. In the southwest, orchids are frequently found in near-coastal swamps beneath or adjacent to Taxandria sp. and Melaleuca sp. trees. Combined with the presence of pollen from the Anarthriaceae family, and the confirmed presence of Banksia speciosa, the microfaunal indications are that the surrounding vegetation of the peat and stumps in the Holocene was similar to the near-coastal heath and kwongkan shrublands that are found nearby today.
When considered alongside the radiocarbon age of the peat deposit of ~7500 cal yrs BP, the most plausible explanation is that peat accumulated in a coastal swamp impounded behind a (Pleistocene) sand barrier dune system that wholly or partially isolated the Wharton Beach embayment from the open ocean into the early Holocene. The presence of lithological glauconite, pyrite framboids and foraminifera in the peat (Figure 11) further confirms the marginal marine conditions in the past.
With transgression, the trees will have died either as groundwaters became more saline and/or more directly through drowning assuming breaching of an older dune barrier. The presence of abundant charcoal particles in the peat deposit indicates the palaeo-community had been burnt during the early Holocene. Whether there is any link between burning and dune destabilisation is unknown and probably unknowable. The peat itself would have become increasingly compacted as sediments accumulated over the top of it as beach deposits migrated landward. Between about 7000 and 2000 years ago sea level was somewhere between 1 and 2 m higher than present (certainly no more than 3 m according to Morgan and Peers, 1973: 17) before falling to modern levels (Baker et al., 2005). During that time of slightly higher Holocene sea levels, the Cape area would likely have been shallow marine to intertidal.
The underlying sands and dune deposits
The OSL age of around 3550 years for the dark-grey sands unconformably underlying the peat unit is somewhat anomalous. These sands are more angular and have a matt appearance, and have a fine-medium gravel component implying a different source to those in the overlying organic peat. Different siliciclastic sand facies have been documented for nearshore sediments at different water depths in the Esperance region, with more siliciclastic slightly gravelly sand associated with seagrass meadows to depths of ~30 m (Tecchiato et al., 2019). Storm events may mobilise these sands (Ryan et al., 2008), and erode seagrass meadows (Tecchiato et al., 2019). Hence, it is possible that a storm event in the Late-Holocene may have mobilised sediment including from seagrass meadows off Wharton Beach. The dark-grey sands were exposed at the leading edge of the peat outcrop, and may have intruded and exacerbated erosion along the contact zone of the consolidated mudstone and peat. Over time, it is likely the peat deposit has been periodically protected as sands have accumulated over it or exposed as overlying sands have been eroded, with each exposure allowing more of the deposit to be washed away. Any carbonate component that was present in the source sediments will have been removed through dissolution in the acidic sediments.
The OSL age of 12,600 years for the base of the dunes behind the exposed peat is also somewhat anomalous as it occurs at or just below the level of the peat. The dune stratigraphy shows repeated layers of fine sands with variable levels of organics within the vadose zone colour the sands different shades of brown as the environments shifted between marine, brackish, freshwater and terrestrial conditions. The OSL sample was taken in the grey sands below the darkest chocolate brown layer, and hence within the phreatic zone of the dune. It is possible that this chocolate brown layer is conformable with the similarly-coloured nearshore peat deposit but further dating is needed to confirm this. The sequence is likely indicative of a former inter-dune or back-barrier lagoon or swamp surrounded by once active dunes that have since been stabilised by vegetation.
The early age of the base of the dune is significant. Excavations in the 1970s revealed the remains of a partially cremated infant that had traces of seaweed/seagrass placed in a shallow pit with nodules of red ochre dated to the 12,845 ± 310 years BP (Smith, 1993). At this time, sea level would have been about 40–50 m below present (Figure 13), placing the shoreline in line just beyond the outer islands and some 20 km further offshore. The presence of seaweed/seagrass is therefore indicative of continued human connections with the coast. This area between the remnant peat deposit and the ancient shoreline remains a huge unknown in our understanding of human-environmental dynamics.
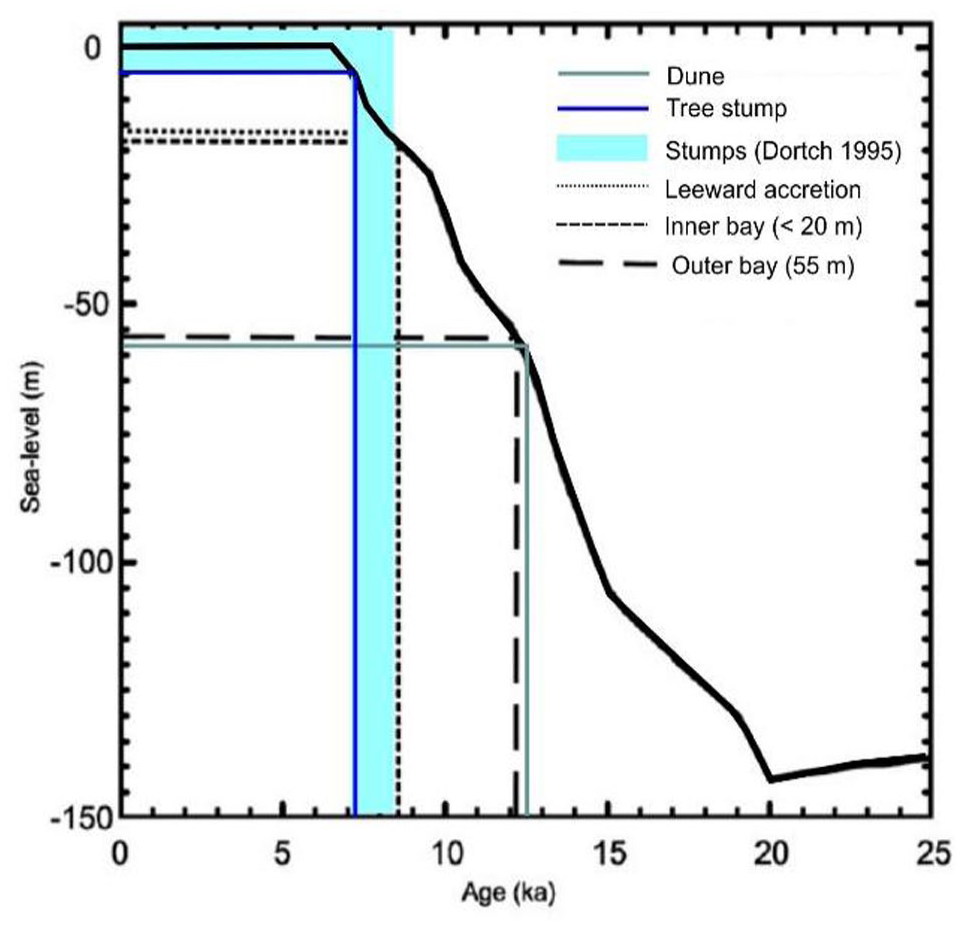
Age of Wharton tree stump (dark blue line), dated stumps elsewhere in the southwest (from Dortch, 1995, light blue line) and Wharton dune base (green line) in relation to eustatic sea level, with approximate ages of inundation for various parts of Esperance Bay (modified from Ryan et al., 2008).
Current situation
With Holocene sea-level rise, shorelines have migrated landwards, resulting in entire ecosystems shifting from being predominantly inland systems to being coastal systems. The changing coastal morphology would have modified once large peatlands to tiny remnants, such as that preserved under the sands at Wharton Beach. Whilst the low tidal range in embayments such as Wharton beach are a key factor in the preservation of the intertidal peat sequence, ongoing erosion including through storm surge is an ongoing threat. Geoscience Australia’s ‘Digital Earth Australia Coastlines’ facility (Geoscience Australia, 2019) indicates the shoreline at Wharton Beach has been eroding at 0.3–0.5 m/year since 1988 (Figure 14). This erosion is further exacerbated by the use of recreational offroad vehicles across the beach, especially during the summer months. The burial of the peat by overlying sands has to date largely protected the peat and the embedded tree stumps but this may not continue.
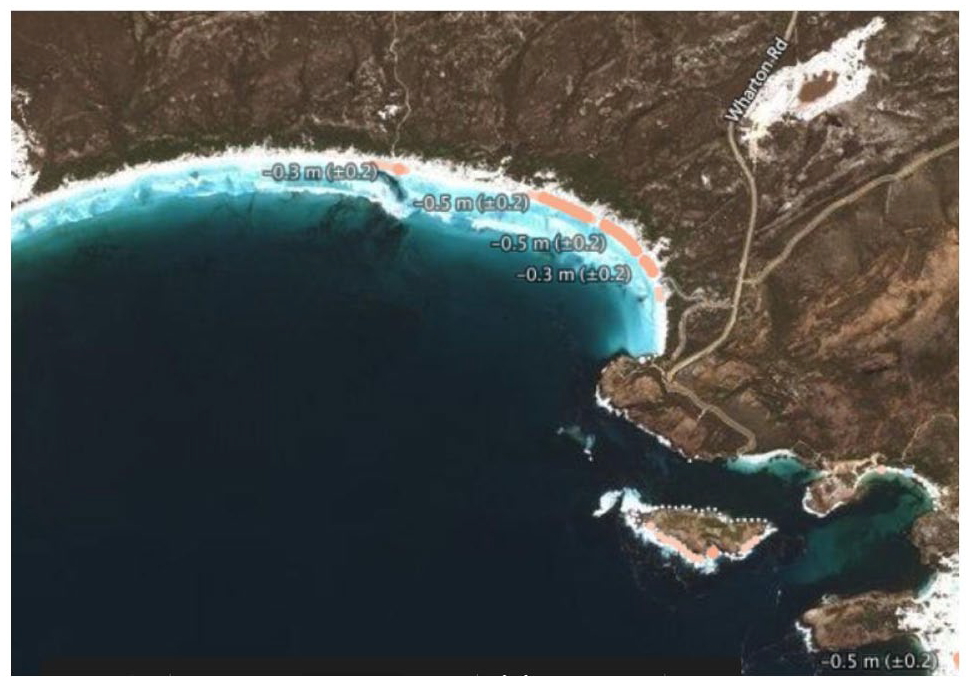
Average erosion rates of the coast at Wharton Beach over the last 33 years as determined from Geoscience Australia (2019) (see https://maps.dea.ga.gov.au/).
Such anoxic, organic-rich, waterlogged deposits are well known to preserve unique ecological records and importantly also undisturbed cultural – including organic – remains (Caple, 2005). While further investigation is needed to map out, characterise and date the peat deposit and the surrounding dune complex at Wharton Beach, such remnants provide a critical link to a former, potentially more extensive coastal wetland landscape (Whinam and Hope, 2005). The very preservation of the peat is also evidence that the late transgression and highstand has not removed all traces of the former land surface. It remains unknown whether other earlier swamp or peatlands remain preserved within the Recherche Archipelago but the preservation of drowned shorelines at depths of around 6–8, 11–13 and 16–21 and 27–36 m (Morgan and Peers, 1973) implies there have been several periods of sea-level stillstand during which swamp development could occur. Preservation of any drowned swamp deposits is partly dependent on how well-consolidated they are and whether or not they have been buried by younger Holocene sands (see also Ward and Larcombe, 2008). A high-resolution geophysical survey of the Recherche Archipelago would aid in this assessment.
Conclusions
The discovery and study of this remnant landform is of cultural value, as it re-affirms stories of the ‘Old People’ who culturally-affirmed what is now a vast submerged coastal plain. The tree stump, although not formally identified, nevertheless provides a sense of connection to place and history, and to the Ancestors, and the environment that they lived in.
“We are strongly connected to this place – the landscapes and seascapes are part of who we are, as a People. When Country reveals itself – like this place, this ancient wetland – we feel a strong sense of pride on how our Old People lived and managed this land and faced all these changes. It is their legacy that we are obligated to protect”. Aunty Donna Beach, Wudjari Elder.
Whilst the exposure and discovery of tree stumps embedded in a remnant peat deposit at Wharton Beach has provided a clue to a former wetland landscape, there remain huge gaps in our knowledge of the now drowned Kepa Kurl coastal plain. Further research on reclaimed coastal wetlands and submerged forests would provide critical information on environmental change linked to both human impacts and natural factors, which in turn will provide important baseline data for conservation and to understanding how these landscapes may respond to future climate and sea level change (Osland et al., 2022). Similarly, development of an anthacological (charcoal) reference akin to that collated for northwestern Australia (Dotte-Sarout et al., 2015) is clearly warranted.
Ongoing research is a critical part of recovering and conserving First Nations knowledge and customary traditions surrounding coastal resource use. In recent cultural values mapping for a proposed new marine park, the Wudjari Elders articulated the importance of collaborative science – Cultural Science and Western Science – with one Elder stating: ‘Healthy Country is when you know a place really well’. In this way, articulating the importance of data collection and research, in every sense, as a cultural value. This study is a model of cultural leadership in adaptive management – that links research data to protection plans, and monitoring programmes, that feed into cultural landscape restoration plans. This also involves explorations into Wudjari Sea Country, investigating the relationship between ancient coastal plain and shorelines, human-environment relations, ecological values, integrative cultural science, and heritage preservation linked to ecosystem management.
Footnotes
Acknowledgements
We wish to thank Chronos radiocarbon lab for undertaking the radiocarbon dating of the wood and peat, and similarly to Wollongong OSL labs for undertaking analyses of the related sand deposits. We thank the two anonymous referees for their helpful corrections on the submitted manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors acknowledge the facilities, and the scientific and technical assistance of Microscopy Australia at the Centre for Microscopy, Characterisation and Analysis, The University of Western Australia, a facility funded by the University, State and Commonwealth Governments. IW is a recipient of an Australian Research Council Discovery Early Career Award (DE180100601). The Tjaltjraak Healthy Country Ranger Program is funded by the National Indigenous Australians Agency, Australian Government.