
Abstract
Reconstructing environments around archaeological sites is complicated by past land management practices and regional-scale climate proxies that can be contradictory and are often located at a distance from the sites themselves. Here we explore environmental information from fossil snail shells which, even when few in number on an archaeological site, may prove invaluable in constructing site-specific data. The palaeoecology of fossil snails and the stable isotopic composition of their shell carbonate can provide context-specific information on vegetation, water availability, and relative humidity during the occupation of a site. We studied terrestrial and aquatic snails from two later Neolithic archaeological sites in the Jordanian badia, Wadi al-Qattafi and Wisad Pools. At specific archaeological site-scale our study highlights the importance of aquatic snails in the reconstruction of semi-arid environments. At Wisad pools rare aquatic snails in contexts dating between ~8.0 and ~7.6 ka demonstrate episodes of wetness; moreover, their shell isotopic compositions indicate that local watercourses were well established, corroborating previous findings that during this period the immediate environs of Wisad Pools were host to C3 plant species more typical of the Mediterranean zone. Moreover, the δ18O signal in these snail shells allow tentative reconstruction of rainwater isotopic compositions and identify the effects of evaporation. Such fine-grained environmental information is much less evident from the terrestrial snail-shell data alone, showing that an ensemble of snail-shell data can be highly sensitive to environmental differentials across an archaeological site. Finally, at a regional palaeoclimate-scale our Wisad Pools snail-shell stable isotope data are consistent with a sustained, Rapid Climate Change (RCC)-driven wetness between 8.6 and 7.6 ka concurrent with cold and wet conditions in the wider Levant.
Keywords
Introduction
The Early to Middle Holocene (~11.7 to ~5 ka) encompasses two major cultural transitions in the Near East; the first from hunting and foraging to farming and herding and the second from small-scale agricultural communities to fully urban centres. Explanations for both transitions have considered the role played by climate change and climate variability at macro and micro scales (Blockley and Pinhasi, 2011; Brooks, 2012; Clarke et al., 2016; Roberts et al., 2018; Rosen and Rivera-Collazo, 2012). Despite the increasing resolution of palaeoclimatic data for the Eastern Mediterranean and surrounding regions, coupled with recent high-resolution regional archaeological and environmental landscape survey data (Palmisano et al., 2021), local short and medium-term environmental conditions around archaeological sites are still poorly understood. To move beyond ‘big-picture’ studies, where the linkage between environmental change and human action is often weakly articulated, local environmental proxies, specific to archaeological sites and independent of land management strategies, are required. Stable isotopes of terrestrial and freshwater snails found on archaeological sites have the potential to provide the kind of fine-grained, localised environmental reconstruction independent of land use practices needed for more nuanced environmental reconstructions. Here we sample and analyse snails found at two archaeological sites in the Jordanian badia, Wisad Pools and Wadi al-Qattafi.
Recent palaeoenvironmental evaluations of the climate and environment of the Near East and Eastern Mediterranean for the last 20,000 years (Jones et al., 2019; Rohling et al., 2019) suggest that the Early Holocene was wetter than today (Clarke et al., 2016; Rohling et al., 2019). During the Early Holocene, centennial scale, cold (possibly wetter) winter-focussed, Rapid Climate Change (RCC) events punctuated warmer, dryer interstadials (Rohling et al., 2019).
The impacts, nature and scale of these RCCs on society are still debated (Mayewski et al., 2004; Rohling et al., 2019). The so-called 8.2 ka event, a 150-year cold interlude, is thought to have reduced sea-surface temperatures in the Eastern Mediterranean by up to 3°C (Rohling et al., 2019). At Jeita Cave (Lebanon) stalagmites record this event as an arid interlude superimposed upon a cold, wet 8.6–8.0 ka RCC (Cheng et al., 2015; Rohling et al., 2019). It is this event that has tended to dominate archaeological discussion around the drivers of contemporaneous cultural change, societal adaptation, and even societal collapse (Biehl and Nieuwenhuyse, 2016; Flohr et al., 2016; Nieuwenhuyse et al., 2016; van der Horn et al., 2015). As yet no clear correlation has been identified between the 8.2 ka event and stratigraphic discontinuities or site abandonments, instead, where relationships can be drawn, the evidence points to accommodation. The lack of evidence for societal dislocation across the 8.2 ka event has led to speculation that communities were resilient to RCCs (e.g. Flohr et al., 2016) but it is much more likely that two processes were taking place. First, that there was considerable environmental and hydrological variation across the region, with well-watered regions (where settlements tended to aggregate) being more resilient to climatic variability than regions where water stress existed. Second, that within the longer 8.6–8.0 ka RCC, the 8.2 ka event was correspondingly muted; societies had already adapted to climatic variability during the preceding centuries and were likely already resilient.
In this article we present pilot isotopic analyses of Jordanian snail-shell carbonate to investigate environmental conditions associated with human occupation between ~8.4 and ~7.6/7.5 ka 1 . Non-marine snail shells can be common fossils in archaeological settings and while some have a burrowing life mode (notably the genus Cecilioides) and are therefore likely to be an intrusive element of fossil assemblages, most species are surface dwellers. Thus, in certain archaeological contexts snail palaeoecology can provide important environmental information. Moreover, there is a strong relationship between snail-shell chemistry and local environmental conditions (Leng and Lewis, 2016; Prendergast et al., 2015). In particular, stable isotopic data from surface-dwelling snail-shell carbonate can provide key information on relative temperature, humidity, and vegetation type. The principal control on shell aragonite δ18O (δ18Oshell), albeit modified by vital effects (discussed later), is the isotopic composition of local rainfall (δ18Orain; e.g. Goodfriend and Ellis, 2002; Prendergast et al., 2015; Rech et al., 2021; Yanes et al., 2019). Higher δ18Oshell ratios broadly correspond to higher δ18Orain ratios and/or reduced rainfall amount. As such, δ18Oshell values can be a crude indicator of wetter (lower values) or drier (higher values) climatic conditions. There is also a relationship between the carbon isotopic composition of local vegetation and snail δ13Cshell (e.g. Baldini et al., 2007; Colonese et al., 2014; Goodfriend and Ellis, 2002). Plants in semi-arid settings, such as the Mediterranean climatic zone, use either a C3 or C4 photosynthetic pathway, responding largely to water stress. C4 grasses, being more water efficient, become more common as aridity intensifies and this metabolism synthesises organic matter with less negative δ13C values. Thus, snails from regions of predominately C3 vegetation and low water stress typically have more negative δ13Cshell values than snails from regions of predominately C4 vegetation and higher water stress, with intermediate values from those in mixed areas.
Local environments
Wisad Pools and Wadi al-Qattafi are respectively located on the eastern and western margins of the harra, an arid, stony desert plateau underlain mainly by Quaternary basalts (Figure 1). The archaeological structures are found on the edge of the plateau where the basalts overlie impure limestones of Palaeocene to Eocene age (Bender, 1975). Topography on these Northern Plateau basalts is gently undulating with elevations of ~800 m just north of Wisad, increasing northward to a maximum of ~1200 m in the foothills of the Syrian Jebel Druze (Allison et al., 2000). The modern climate is seasonal with hot, dry summers and cool winters (Wigley and Farmer, 1982). Mean annual maximum summer temperatures are ~35°C–38°C, while mean annual winter temperatures are between 2°C and 9°C (Al-Homoud et al., 1995; Allison et al., 2000). Mean annual rainfall ranges from <50 mm a−1 in the southern harra to ~250 mm a−1 in the north, the latter reflecting the orographic influence of the Jebel Druze. Eighty percent of this rainfall occurs between December and March, often as discrete storm events (Allison et al., 2000). Of this rainfall between 85% and 92% is lost to evaporation, 5%–11% infiltrates and just 2%–4% becomes surface flow (Allison et al., 2000). The resulting intermittent surface flow is focussed into wadis, the largest radiating out from the Jebel Druze (Al-Homoud et al., 1995). Some of these wadis are deeply incised, suggesting wetter climates in the past. Wadis may feed closed depressions or evaporative pans, the largest known as qi’an (sing. = qa’). Soils in the eastern badia are either absent or very poorly developed (Allison et al., 2000) except in the topographic lows of wadis or in qi’an. At Qa’ al-Qattafi and ‘small’ Qa’ al-Wisad (immediately below the pools), the sediments are between 1.0 and 3.0 m thick (Jones et al., 2021) with underlying bedrock variously basalt (‘small’ Qa’ al-Wisad) or limestone (Qa’ al-Qattafi). At 60 cm depth OSL ages show the qa’ sediments as being <7.9 ka and at these depths CaCO3 content is between 10% and 15% (Jones et al., 2021). This carbonate is probably derived from bedrock (Qa’ al-Qattafi) or windblown dust, although Allison et al. (2000) describe gypsitic and calcitic deposits in some areas which they regard as ‘secondary’, that is, calcrete. There is currently little or no permanent vegetation in the south of the harra, but some in the north (Al-Homoud et al., 1995) and in wadis to the west such as the Azraq oasis and Shomari Wildlife Reserve.
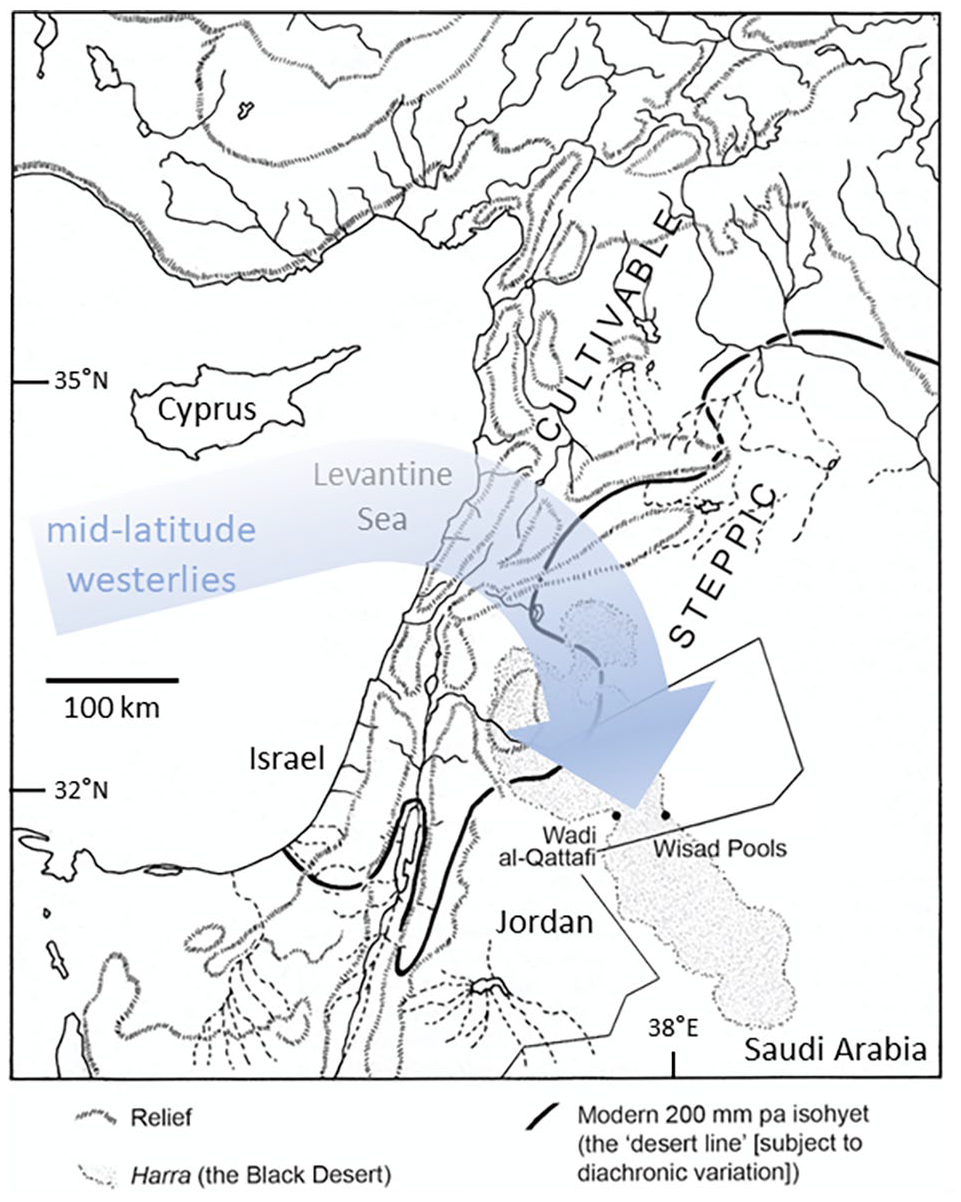
Map showing sites mentioned in the text and inferred likely source and transport pathway of rainfall to the study area.
Winter precipitation in the Middle East is mostly generated in the Mediterranean/Cyprus low system, where cool, dry air from continental Euro-Asia interfaces with the warmer Mediterranean Sea (Burstyn et al., 2019). Much of this precipitation, however, is unlikely to reach the eastern Jordanian desert directly from the west, being in the rain shadow of the Jordanian highlands. Instead, recharge is mostly sourced from the north (Figure 1) as recorded by the drainages radiating from the Jebel Druze (Al-Homoud et al., 1995) and by flow directions in the subsurface aquifers (Al-Homoud et al., 1995; Allison et al., 2000). Wet-season flows in the wadis of the Azraq Basin, west of the basalt plateau (e.g. Wadi Rajil) can be considerable (Al-Homoud et al., 1995). Until the 1980s four springs discharged a large quantity of water from the upper aquifer into the Azraq Wetland Reserve, creating shallow pools up to 2 m deep, including marshland extending to 8 km2; these subsequently dried out due to large-scale abstraction in the 1980s (Bajjali and Al-Hadidi, 2006). Today the shallowest aquifer is at ~50 m below surface in the Azraq Basin because of abstraction.
Modern interpolated wintertime (DJFM) meteoric water in the area has δ18O of −7.3‰ VSMOW (Figure 2). These modern values can be cross-compared with data from Bajjali and Abu-Jaber (2001) who studied Holocene-aged palaeogroundwaters from the Azraq Basin and the Ashquaf Heights (just west and north of the harra study area respectively). The Azraq groundwaters gave DIC 14 C ages between 2.5 and 8.0 ka and δ18O values in the range −6.8 to −5.2‰VSMOW (Figure 2), the Ashquaf groundwaters having a similar δ18O range between −6.3 and −5.5‰ VSMOW. The most negative groundwater compositions are likely to be indicative of unevaporated winter recharge and compare well with the mean modern OIPC interpolated value of −7.3‰ VSMOW (Figure 2).
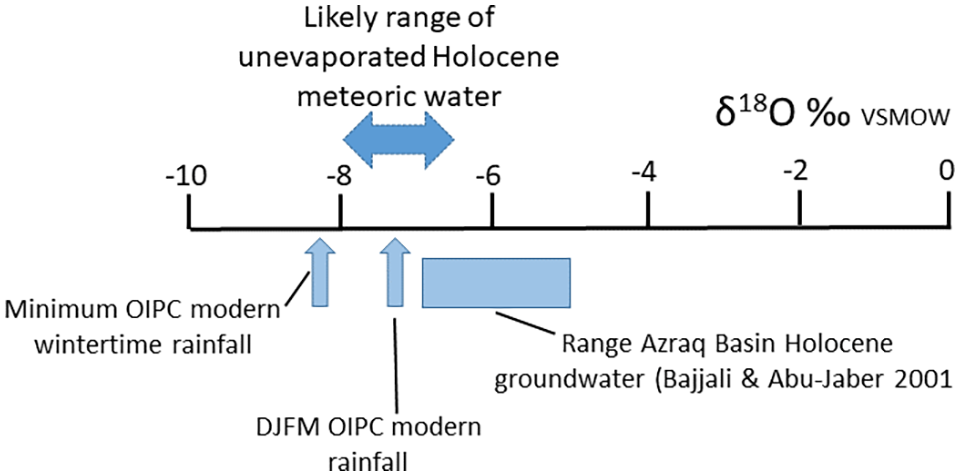
Schematic to show likely range in rainwater δ18O corrections for the Wisad area. OIPC is the Online Isotopes in Precipitation Calculator (OIPC v3.1; Bowen, 2020; Bowen and Revenaugh, 2003; Bowen and Wilkinson, 2002) see Supplemental Table S4 and www.waterisotopes.org. The isotopic composition of modern meteoric recharge in the region is interpolated from the Global Network of Isotopes in Precipitation (GNIP) database of long-term monthly values. We assumed that most Holocene recharge to the eastern badia was sourced from local wintertime runoff (DJFM; Allison et al., 2000) as it is today.
Wisad Pools
Wisad Pools is a series of nine pools along a short (c. 1.5 km) wadi leading from the surrounding plateau down to Qa’ al-Wisad (Figure 3). Over 500 structures and more than 400 petroglyphs concentrate around the pools (Hill et al., 2020) in an area of approximately 1.5 km2 with additional structures, animal pens and enclosures diminishing in density away from the pools. Radiocarbon dates combined with artefactual evidence from the ground surface date the structures to ~9.0 to ~7.5 ka (Figure 4; Supplemental Table S1), however, one surface concentration of chipped stone artefacts dates to ~9.5 to ~8.9 ka indicating localised earlier use of the region.

Aerial photograph of Wisad Pools showing the locations of W-400 closer to the qi’an (far left side of image) and W-80 a short distance from Pool 8. (Aerial Photographic Archive of Archaeology in the Middle East (APAAME), archive accessible from: www.humanities.uwa.edu.au/research/cah/aerial-archaeology)
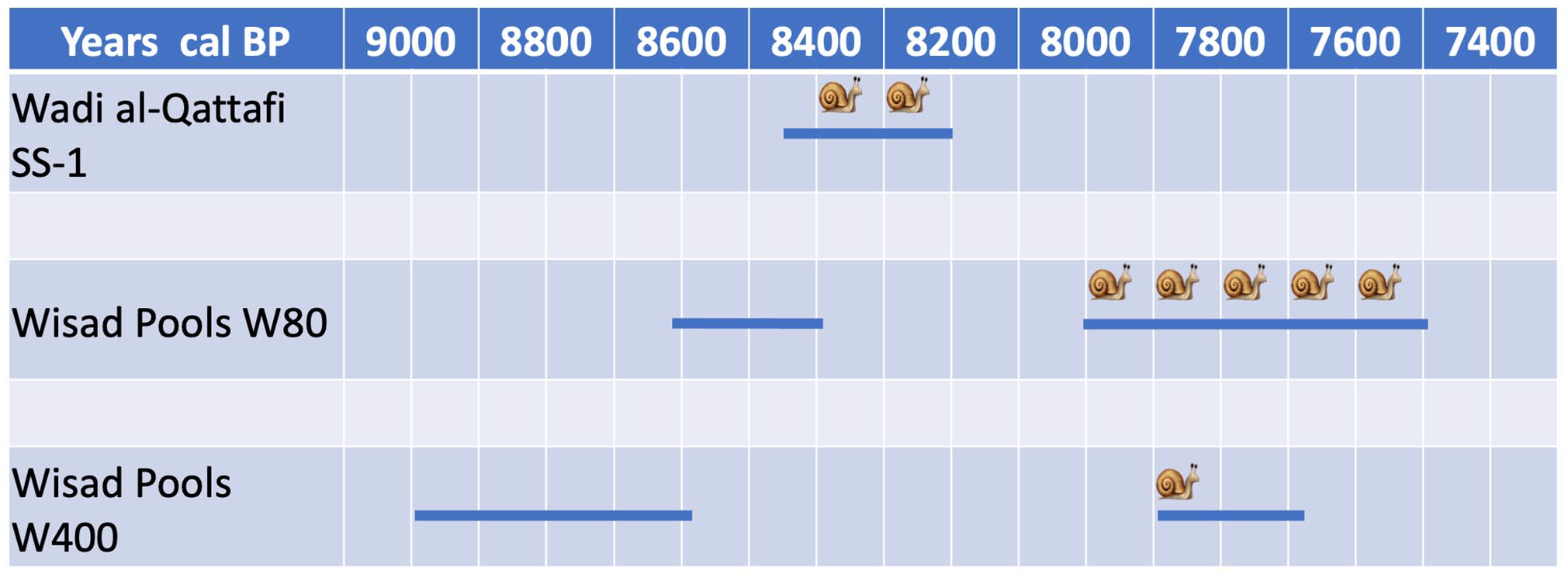
Schematic radiocarbon date ranges for Wisad Pools and Wadi al-Qattafi. Blue horizontal lines represent calibrated radiocarbon years BP, based on existing radiocarbon dates. Snail icons show the approximate chronological windows of the snail samples.
Meticulous excavation at Wisad Pools and a 100% archaeological collection strategy accounts for only three structures having been excavated to date, W-400, W-80 and W-66. Throughout the archaeological deposits snails were extremely rare, deriving from W-400 and W-80 only.
Located near Pool 8 (Figure 3; Supplemental Figure S1) W-80 is a large curvilinear structure (~12 m), modified, abandoned and reused (Rollefson et al., 2018). Gazelle hunting, supplemented by onager, hare and other wild-animal taxa, provide evidence for a subsistence economy that was largely based around wild fauna but supplemented by sheep and goat herding and wild-plant-food processing using – in later phases – massive grinding slabs with inset mortars.
Approximately 650 m to the north of W-80 on the edge of the qa (Figure 3; Supplemental Figure S2) is located structure W-400. The interior of W-400 is much smaller than W-80 (~2.60 m in diameter) and the building style – if not the general layout of interior features – is strikingly different, with smaller basalt blocks and less care in construction. Small finds are far fewer in number, most notably fewer arrowheads, contrasting with the 789 projectile points so far found at W-80 (Rowan, 2020). The different construction techniques combined with fewer artefacts suggests that W-400 was a type of structure associated with a herding element within the Neolithic cultural complex that characterised this region (Wasse et al., 2020).
Wadi al-Qattafi
Wadi al-Qattafi is a major north-south drainage located approximately 60 km west of Wisad Pools, characterised by basalt-capped ‘mesas’ that rise 40–60 m above the wadi bed (Hill et al., 2020). On and around the mesas are hundreds of prehistoric structures of great diversity, including burial mounds, tower tombs, enclosures, ‘wheels’, hunting traps (‘kites’) and dwellings (Hill and Rowan, 2017). Two structures have been excavated to date, SS-1 on Mesa 7 and SS-11 on Mesa 4. SS-1 is a well-built, curvilinear stone structure with an interior plastered basin and a fire pit (Rollefson et al., 2016). SS-11 also had a corbelled wall construction, two low entranceways and a carefully paved floor (Wasse et al., 2012). Adjacent storerooms, and internal installations attest to a degree of domestic use for both.
Chronological summary
There are six radiocarbon dates for W-80 (Supplemental Table S1). These, combined with archaeological evidence, suggest that the building had two main foci of utilisation, one lasting about 200 years between ~8.6 and ~8.4 ka, and the second beginning at ~8.0 ka and ending at ~7.5 ka. Within this sequence are three dated archaeological phases, the first beginning at ~8.6 ka, the second beginning at ~8.0 ka, and the third phase dating around ~7.5 ka.
W-400 has not yet been assigned archaeological phases but two radiocarbon dates place the utilisation of the structure between 9.0 ka and 7.7 ka. RC sample 53 comes from an early room fill, while the second (sample 70) comes from a fill representing final utilisation before abandonment. The two dates top and tail the utilisation of the structure and indicate long-term intermittent use.
Structure SS-1 at Wadi al-Qattafi has six archaeological phases; the earliest is Phase 1 and the latest is Phase 6. Phase 6 and to a certain extent, Phase 5, postdate the primary use of the structure. Four radiocarbon dates (Supplemental Table S1) place Phases 2 and 3 between ~8.4 and ~8.2 ka, while lithics from SS-1 corroborate a reasonably compressed period of use compared to W-80 (Rollefson et al., 2016).
A single radiocarbon date from an expedient hearth sealed by flagstones places SS-11 at ~7.4 to ~7.3 ka (Rowan et al., 2015).
Materials and methods
Two hundred and one terrestrial and six freshwater molluscan remains, found within interior and exterior fills of W-80 and W-400 at Wisad Pools, SS-1 and SS-11 at Wadi al-Qattafi, were collected through flotation of archaeologically derived sediment. The surface dwelling terrestrial snails lived, died and were subsequently sealed in the archaeological contexts they were discovered in. As no mud brick or mud mortar was used in construction material at either site, aquatic snails could not have been introduced as part of such material. Instead, we suggest that these snails were introduced to the archaeological deposits as living individuals on reeds or rushes, that had been growing around the qa’in and the pools, brought into the structures as roofing or flooring material (discussed in detail later).
Snail identification
The assemblages of molluscan remains were sorted under a low-powered Wild binocular microscope and all identifiable specimens were quantified (see also supplemental information). To arrive at a minimum number of individuals (MNI) for each taxon, only apical fragments (representing the tip of the spire of the shell) were counted. For some taxa (e.g. clausiliids) apices were poorly preserved and other diagnostic elements were counted instead, usually the aperture (mouth). Species identified from shell fragments possessing distinctive microsculpture are listed as present (+) in Supplemental Table S2a, S2b, and S3. These were not quantified since it is impossible to know whether the fragments were derived from a single shell or from several specimens.
The taxonomic status of many ‘Middle East’ species requires revision, as minor conchological differences have given rise to a plethora of names across a wide region and are likely to be synonyms. This applies to land snail species within Hygromiidae (see, e.g. Neubert and Bariche, 2013), although since many of the differences in these taxa are based on shell characteristics it is still possible to identify them from well-preserved fossil specimens. The species listed here are therefore in some cases tentative, since the preservation of the material was not good enough for definitive species-level identifications.
Snail-shell sampling: Stable isotopes
For stable isotopes we sampled >10% of the terrestrial snails at each site and all the available freshwater snails from Wisad Pools as follows.
W-80, Wisad Pools
Six snail samples for isotopic analysis came from structure W-80 at Wisad Pools (Supplemental Table S2a). The samples derive from post-8.0 ka deposits except one terrestrial sample deriving from a pre-8.0 ka deposit. The three terrestrial samples were recovered from contexts within the building, one from a pre-8.0 ka hearth and the other samples from post-8.0 ka hearths or firepits (Supplemental Table S2a). The three aquatic samples (all different species) were all from post-8.0 ka occupation deposits.
W-400, Wisad Pools
Two aquatic snails were recovered from Structure W-400 from post-8.0 ka deposits (Supplemental Table S2a), which likely represent the final occupation of the structure sometime after ~7.7 ka.
M4 SS-11, Wadi al-Qattafi
A single terrestrial snail sample came from structure M4 SS-11. This single example was collected from an upper fill of the structure. As the sample is identifiable as Pupoides coenopictus it has been included in the isotopic analyses, but its location suggests its chronological and archaeological integrity may be in doubt. Accordingly, it has not been included in the subsequent discussion and interpretation.
M7 SS-1, Wadi al-Qattafi
Twenty terrestrial snail samples came from structure M7 SS-1 at Wadi al-Qattafi (Supplemental Table S2b). Snail samples were derived from Phases 1 to 5, extending the chronology of the snail data back beyond ~8.4 ka. The contexts from which the snail samples derive are mostly from hearths or firepits, although some samples derive from a plaster basin and another from a paved surface.
Mineralogy and stable isotopes methods
Thirty-five fossil snail shells were selected for stable-isotope analysis after identification, including all six aquatic snails from Wisad Pools; all but two specimens were adults that varied in size from 3 to 10 mm. We avoided known burrowing species to eliminate the risk of analysing material younger than the context sediment. Four bulk sediment samples were also analysed for stable isotopes.
Snail shells were crushed gently to expose internal surfaces and adherent sediment was removed with a steel seeker. Shell fragments were then immersed in 30% H2O2 at room temperature for 24 h, and then placed in an ultrasonic bath for 30 min. The fragments were air dried, then ground gently in an agate pestle and mortar to a fine powder. Isotope data from these homogenised powders should thus represent mean values representative of the snails’ lifetime. Shell mineralogy from a subsample of individuals was checked by powder X-ray diffraction (XRD).
Isotopic analyses (University of East Anglia Stable Isotope Laboratory) were made on 75 ± 5 µg samples, run alongside 75 ± 5 μg internal standards of UEACMST (University of East Anglia Carrara Marble Standard; δ18O −2.05‰VPDB; δ13C 1.99‰VPDB), reacted with 105% (ρ = 1.92 gml−3) phosphoric acid (H3PO4) at 90°C in an on-line common acid bath. The evolved CO2 was purified and analysed for δ18O and δ13C using a Europa SIRA II dual inlet isotope ratio mass spectrometer. The data are calibrated to international reference scales (VPDB and VSMOW) using IAEA Certified Reference Material NBS-19 (δ18O −2.20‰VPDB; δ13C 1.95‰VPDB). Repeat analysis of both international and internal reference materials gave 1σ errors of less than ±0.1‰ for both δ18O and δ13C. Isotope data discussed in the text are relative to VPDB unless indicated otherwise.
Results
Molluscan assemblages
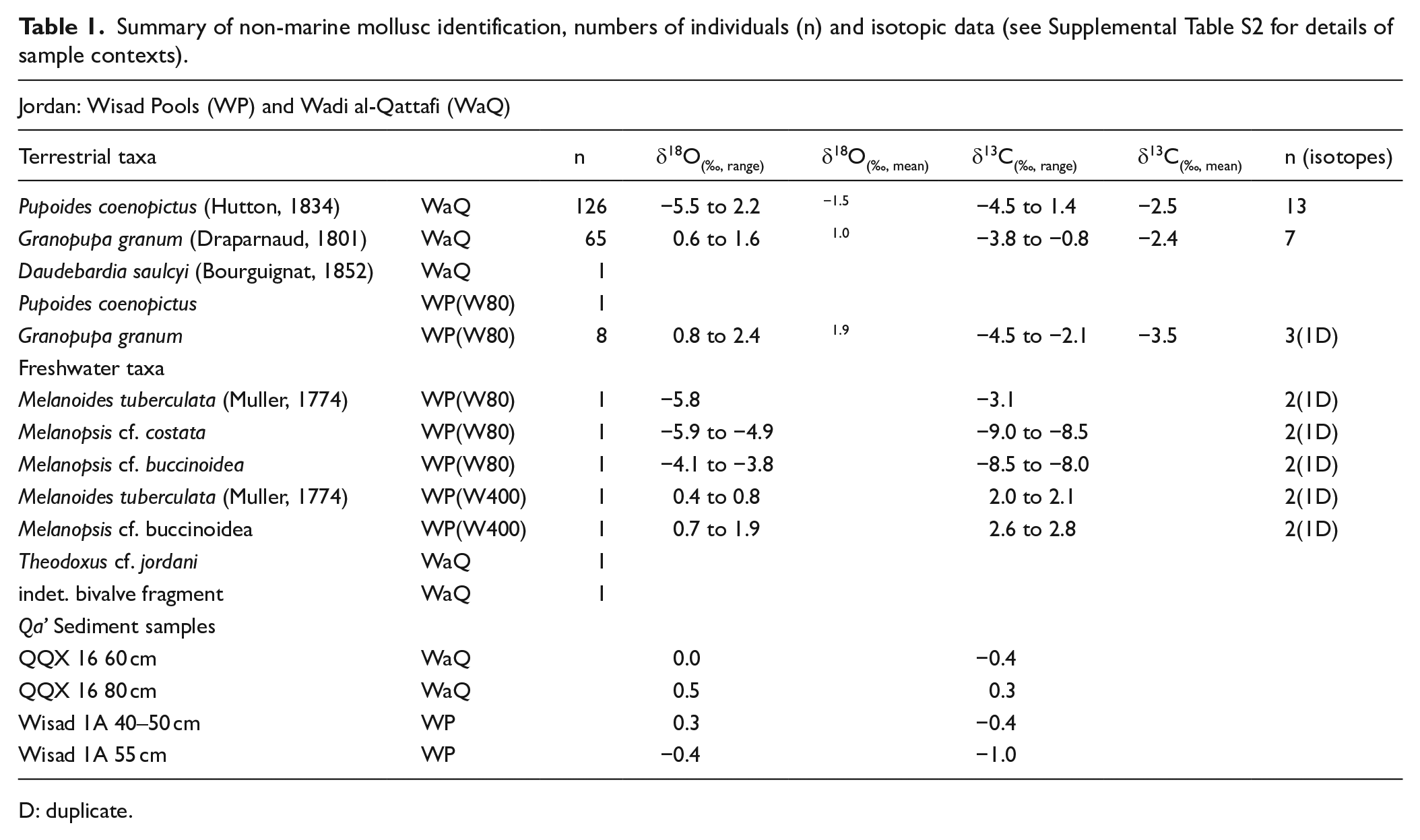
The most numerous molluscan remains recovered from Wadi al-Qattafi, Mesa 7 (structure M7 SS-1) and Wisad Pools (structure W-80) comprise two tiny terrestrial species: Pupoides coenopictus (Hutton, 1834) and Granopupa granum (Draparnaud, 1801) (Table 1). Adult snails are typically 1–3 mm long. A few fragments of cm-sized freshwater taxa were also present in samples from structures W-80 and W-400 at Wisad Pools (Table 1), although it was notable that terrestrial and freshwater species were mutually exclusive and never occurred within the same sample, suggesting incorporation into archaeological contexts followed different pathways. The only other terrestrial species represented in the Wadi al-Qattafi samples was a single shell of the semi-slug Daudebardia saulcyi (Bourguignat, 1852).
Summary of non-marine mollusc identification, numbers of individuals (n) and isotopic data (see Supplemental Table S2 for details of sample contexts).
D: duplicate.
Snail-shell stable isotopes
All snail shells were composed of pure aragonite. Terrestrial snails G. granum and P. coenopictus from Wadi al-Qattafi (M7 structure SS-1) and Wisad Pools (structure W-80) have similar shell δ13C values ranging from +1.4 to −4.5‰ (Table 1 and Figure 5). P. coenopictus from M7 SS-1 has δ18O values ranging from +2.2 to −5.5‰ while G. granum has more restricted δ18O from +1.6 to +0.6‰. Three of the 4 W-80 G. granum individuals had the most positive δ18O compositions (+2.1 to +2.4‰; Figure 5). Three species of freshwater aquatic snails (Melanoides tuberculata, Melanopsis cf. buccinoidea and Melanopsis cf. costata) from structures W-80 and W-400 (Wisad Pools) have quite different isotopic values; individuals from W-400 have low positive isotopic compositions (δ18O between +0.4 and +1.9‰; δ13C between +1.9and +2.8‰) while individuals from W-80 have the most negative Wisad Pools isotopic compositions (δ18O between −3.8 and −5.9‰; δ13C between −3.1 and −9.0‰; Figure 5).
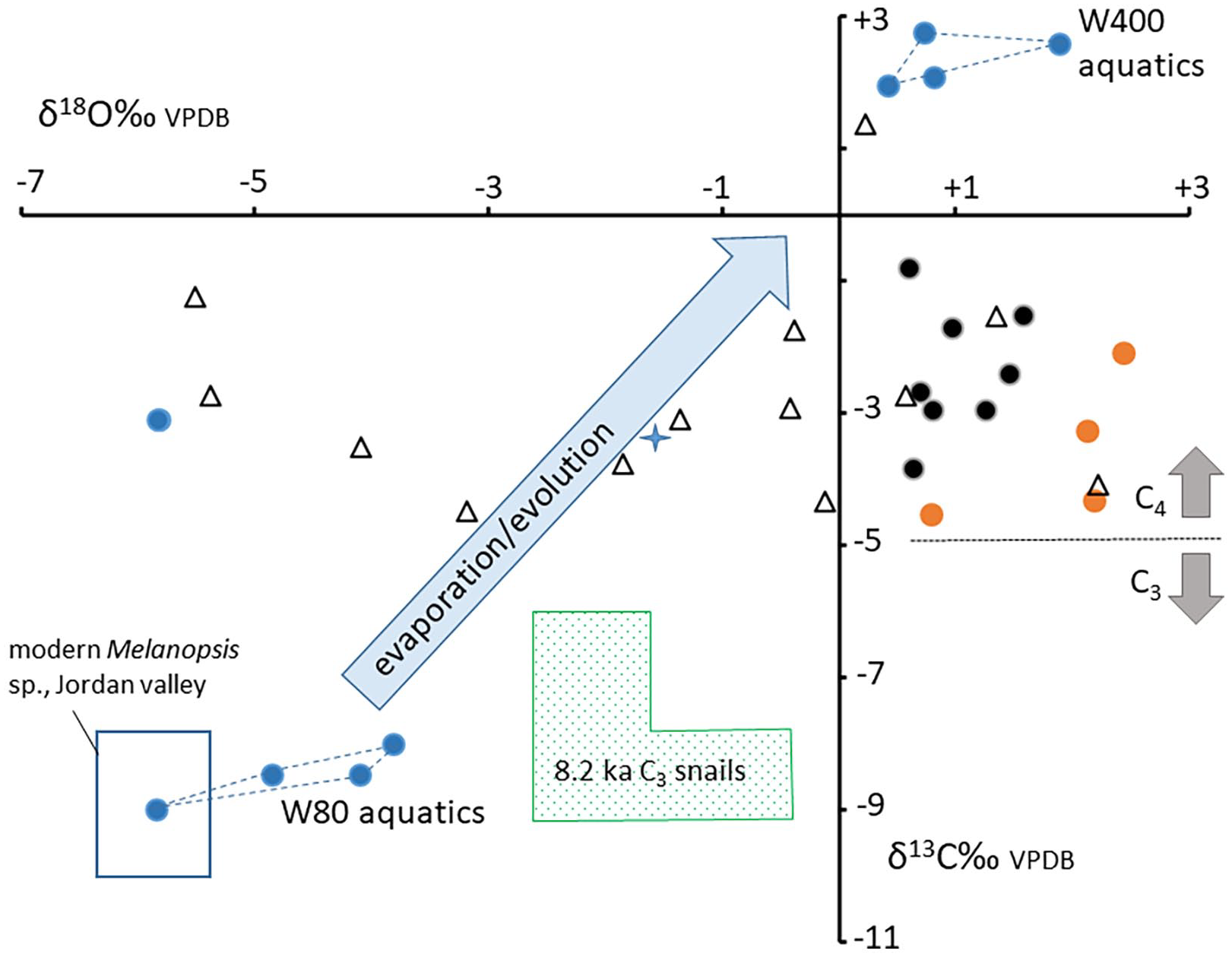
Crossplot of the snail-shell aragonite stable isotope data. Blue dots and fields show the aquatic snail data from structures W-80 and W-400. These are interpreted as near equilibrium values and the blue arrow shows likely evaporation trend between end members. The blue outlined box shows where modern Melanopsis sp., data from the Jordan Valley plot (Zaarur et al., 2016) while the blue star is a modern aquatic snail from evaporated Sea of Galilee water (Zaarur et al., 2016). All other symbols are terrestrial snail shell values; orange dots are G. granum (W-80), black dots are G. granum (SS-1) and open triangles are P. coenopictus (M7 SS-1). Green stippled box shows field for comparative ~8.2 ka BP C3 terrestrial snail data from Italy (Colonese et al., 2010, 8.8 to 6.7 ka BP), Algeria (Padgett et al., 2019, ~8.0 ka BP), and Spain (Yanes et al., 2012 8.4 ka BP).
We do not have any local modern snail data with which to compare the fossil data. Today the region is hyper-arid; it is possible that these tiny terrestrial snails are present but inactive during our field seasons. However, comparison with modern snails from the southern Levant and Negev desert show that Sphincterochila sp., collected from the same sites as Trochoidea seetzeni, are enriched in both 13 C and 18O by an average of 3 and 2‰ respectively (Goodfriend and Magaritz, 1987). These differences are tentatively attributed to the relative metabolic activity of the taxa, perhaps the snails’ overall activity, or shell deposition at different times in the activity cycle of the snail species (Goodfriend and Magaritz, 1987). Whatever the precise cause, the data show that isotopic variation of 2-3‰ in the fossil Jordanian snails should not be attributed to environmental drivers without supporting evidence. There are no aquatic taxa living at Wisad Pools today; however, there is comparative stable isotope data for modern Melanopsis sp. from the Jordan Valley (Zaarur et al., 2016; Figure 5).
Interpretation
Terrestrial snails
Pupoides coenopictus and G. granum are both characteristic of rocky habitats in relatively arid regions. The distribution of P. coenopictus is most extensive in tropical regions, centred on Asia and tropical sub-Saharan Africa, but at the edges of its range it is known from northwest Africa, around the Arabian Peninsula, and across southwest and central Asia (Neubert, 1998; Seddon, 1992). Granopupa granum, which is often found in association with P. coenopictus, is also found in rocky habitats and in screes and leaf litter amongst rock ledges and crevices (Gittenberger, 1973; Kerney and Cameron, 1979), with a circum-Mediterranean distribution. On a more local scale G. granum is widely distributed in Israel and Palestine, whereas P. coenopictus seems to be rarer in this region (Amr et al., 2018; Heller, 2009). The carnivorous semi-slug D. saulcyi feeds on earthworms and lives within the soil (Mienis, 1976), and the presence of both the slug and its inferred prey implies some development of humic soils derived from decaying plant matter, in contrast to the arid environments suggested by the aforementioned species.
Terrestrial snails: stable isotopes
It is well known that terrestrial snail shells are composed of aragonite such that presence of calcite is usually thought indicative of diagenetic alteration (e.g. Colonese et al., 2014). The Jordanian snail-shell mineralogy is 100% aragonite indicative of little or no alteration.
Snail-shell aragonite forms from body-fluid bicarbonate which has three potential carbon sources with different isotopic signatures: (1) metabolic CO2 from respiration, (2) atmospheric CO2 that exchanges (and fractionates) through the snails’ body tissues and (3) CO2 generated in the stomach from ingested CaCO3 (Goodfriend and Ellis, 2002). In our Jordanian samples, calcrete or limestone bedrock could be a component of ingested CaCO3. The significance of atmospheric CO2 exchange through snail tissues is not well understood. However, as terrestrial snails are exposed mostly to plant root respired CO2, shell isotopic compositions are strongly related to the feeding behaviour of the snail which in turn reflects the vegetation type the animal consumed during its life (Balakrishnan et al., 2005; Colonese et al., 2014).
The δ13C composition of plants is primarily controlled by their photosynthetic pathway, C3, C4 or CAM which are also linked to water stress (Cerling et al., 1993; Colonese et al., 2014; Deines, 1980; Liu et al., 1996; Smith and Epstein, 1971). In brief, C3 plants are not well adapted to water stress and typically thrive in moist or wet climates, whereas C4 (including most grasses) and CAM plants like cacti are better adapted to aridity and can be common in semi-arid steppe conditions and arid deserts.
Laboratory studies with Cornus (Helix) aspersa showed that shell carbonate has δ13C ~12‰ less negative than shell-bound organic carbon, this organic carbon being related directly to the vegetation type ingested (Stott, 2002). The difference is caused by fractionations during production of metabolic CO2 and between this CO2 and the bicarbonate from which shell aragonite precipitates. As C3 plants typically produce organic matter with a δ13C around −26‰, these fractionations mean that shell carbonate should have a value of ~−14‰. In practise values between −8 and −12‰ are commonly reported for C3 ingesting snails (e.g. see Balakrishnan et al., 2005), because in nature they can ingest other vegetation types and soil carbonate, have differing water use efficiencies (Prendergast et al., 2017), and because the exact metabolic fractionations probably differ between species. In contrast C4 and CAM plants have less negative δ13C which typically translate to shell aragonite δ13C less negative than ~−5‰ (e.g. see Balakrishnan et al., 2005).
The δ13C compositions of G. granum and P. coenopictus shell aragonites, ranging from +1.5 to −4.5‰ (Table 1), largely reflect ingestion of C4 arid scrub vegetation (Goodfriend and Magaritz, 1987), much more enriched in 13C relative to C3 ingesting fossil snails of similar age (Figure 5) from southern Europe and North Africa. This result is consistent with the discovery of dry-adapted Anabasis sp., other Chenopdioideae, tamarisk and salt tolerant Zygophyllum sp. charcoal in both Structure W-80 (Wisad) and M7 SS-1 at Wadi al-Qattafi (Rowan et al., 2017; Wasse et al., 2022), plants that are broadly indicative of arid steppe environments. The C4 signal can be modulated by ingestion of soil carbonate. To explore this we analysed qa’ sediments ~8.0 ka in age (from Jones et al., 2021) from both Wisad Pools and Wadi al-Qattafi. These sediments have δ13C between -1.0 and 0.3‰ (Table 1) suggesting a bedrock or blown dust carbonate source which would have trivial impact on the plant source signal. There is no obvious species-dependent or geographic difference in δ13C, despite Structure M7 SS-1 being ~50 km from W-80.
Interpretation of terrestrial snail-shell δ18O is complicated by significant offsets, of between 2‰ and 8‰, between shell aragonite and local precipitation (Goodfriend and Ellis, 2002; Goodfriend and Magaritz, 1987; Yanes et al., 2008; Zanchetta et al., 2005), an effect especially strong in semi-arid settings (Yanes et al., 2019). More recent clumped isotope data suggest that while terrestrial snail-shell temperatures may be close to growth season temperatures (Dong et al., 2020) most record higher than expected snail activity temperatures (Eagle et al., 2013; Wang et al., 2016; Zaarur et al., 2011) which is probably caused by a kinetic (vital effect) fractionation (Dong et al., 2021). Put simply, modern terrestrial snail-shell δ18O values do not usually record ambient environmental air temperature even when rainfall δ18O is known.
Overall, the Wisad terrestrial snail-shell δ18O values are similar to those reported for Levantine arid and semi-arid settings (Goodfriend and Magaritz, 1987). At the local scale there is no difference in the W-80 G. granum δ18O value from the earlier contexts when compared to the later contexts. However, the W-80 values are about 1.0‰ more positive than data from the same species collected from Structure SS-1 in Wadi al-Qattafi (situated approximately 50 km to the west of Wisad Pools), which correspond temporally with the earlier contexts at W-80 (Supplemental Table S2a). This 1.0‰ difference could indicate a slightly more arid signal at W-80 reflecting an isohyet gradient from wetter to dryer in a south-easterly direction as exists today. We accept, however, that the difference could also be attributed to slightly different shell precipitation temperatures, metabolic factors (Goodfriend and Magaritz, 1987; Zaarur et al., 2011), or differing ambient relative humidities (cf. Balakrishnan et al., 2005).
The P. coenopictus δ18O data from M7 SS-1 at Wadi al-Qattafi show a much wider spread than those from co-occurring G. granum in contexts that span just 250 years (Figure 5; Supplemental Table S2b). The more positive P. coenopictus values are similar to the M7 SS-1 G. granum values (Figure 5) suggesting arid, water-stressed conditions, while the more negative values, below −1‰, and down to −5.5‰, are lower than values expected from species specific variation (cf. Goodfriend and Magaritz, 1987). This suggests these P. coenopictus were accessing relatively unevaporated meteoric water ‒ with a much more negative isotopic composition ‒ perhaps during more sustained rainy conditions (cf. Goodfriend et al., 1989). Modern interpolated DJFM meteoric water in the area has δ18O of −7.3‰ VSMOW (Figure 2) and the most negative interpolated modern wintertime δ18O for the eastern badia are ~−8.2‰ (Figure 2; Supplemental Table S4), indicating that individual rainfall events are likely to attain the required negative isotopic compositions today. These negative P. coenopictus δ18O values suggest less arid conditions; decreased water stress could have been accompanied by increased relative humidity (RH), a parameter which affects diffusive evaporation of the snail body fluid, increasing RH related to decreasing shell carbonate δ18O (Balakrishnan and Yapp, 2004; Balakrishnan et al., 2005). However, there is no evidence that such conditions were similarly available to G. granum at either Wadi al-Qattafi or Wisad Pools. δ18O in G. granum shells always appears to indicate arid conditions accompanied by low RH.
Aquatic snails
While aquatic taxa were very rare, the few recovered from structures W-80 and W-400 at Wisad Pools included M. tuberculata, a common species ranging across North Africa and southern Asia known for its tolerance of fluctuating salinities and water temperatures, and also rapid powers of dispersal and colonisation due to its parthenogenetic mode of reproduction. At least two species of the variable genus Melanopsis (see Grossowicz et al., 2003) were also noted rarely in several samples, although due to the fragmentary nature of the specimens the precise identity of these taxa remains tentative (Table 1). A single fragment of another variable species, Theodoxus cf. jordani, was recovered from one of the Wadi al-Qattafi samples (Table 1). Both Melanopsis and Theodoxus are widespread in the Near East found in desert springs (Tchernov, 1975) and clear, relatively fast-flowing waterbodies across the region, the latter preferring stony substrates. Melanopsis favours the presence of succulent aquatic vegetation (Moshkovitz and Magaritz, 1987), feeds on algae and can live for several years.
The clear distinction between samples containing freshwater taxa and those containing terrestrial species can be explained if the freshwater snails entered W-80 on plant material – perhaps reeds or rushes – found growing in or around nearby pools and qi’an, while the terrestrial snails lived and died in the archaeological contexts in which they were found. Two shells of M. costata were found in the interior of W-80 in an occupation layer with a high density of chipped stone and bone associated with large grinding slabs and fire pits (Table S2a). Two shells of M. buccinoidea came from an area of paving outside W-80 adjacent to the main doorway and two shells of M. tuberculata came from an area of basalt corbelled roof collapse and may be associated with squatter occupation within the upper part of the collapsed structure, likely dating to the end of the structure’s sequence (Supplemental Table S2a).
Aquatic snails: stable isotopes
Freshwater aquatic snail-shell δ18O is more typically an equilibrium value and may thus record ambient water temperatures (Grossman and Ku, 1986; White et al., 1999). They are less prone to species-specific effects (White et al., 1999) and clumped isotope data from modern Melanopsis sp. in the Jordan Valley confirm that this genus forms its shell carbonate under isotopic equilibrium (Zaarur et al., 2016).
The Wisad pools freshwater aquatic snails (M. tuberculata, M. cf. buccinoidea) plot in quite distinct isotopic space when compared to the terrestrial snail data (Figure 5). Individuals collected from structure W-400 have low positive isotopic compositions, while individuals from Structure W-80, near a semi-permanent seasonal runoff-fed pool (pool 8, Rollefson, 2013; Rowan et al., 2017), have the most negative isotopic compositions. The difference in δ18O values reflect local pool-water isotopic compositions which, although fed by the same seasonal runoff (as today), appear to have suffered strong evaporation and desiccation in the upper qa near W-400, compared to pool 8 near W-80 where evaporative effects are less marked (see also Rollefson, 2013). If the snails were feeding on cyanobacteria and algae, the shell δ13C values reflect the isotopic composition of this food source inherited from the dissolved inorganic carbon (DIC) in the pool water (cf. Deines, 1980). By contrast at W-400 the δ13C values are consistent with very shallow pool environments where DIC had fully or partially equilibrated with atmospheric CO2, resulting in 13C enrichment. The freshwater aquatic snail data thus define end-members of an ‘evaporative/evolved trend’ (Figure 5) with W-80 values representing pool environments with lower water stress/evaporation and W-400 values representing evaporative environments (cf. McLaren et al., 2012) with higher water stress and a snail food source dominated by cyanobacteria that had inherited isotopically enriched 13C from the DIC. This interpretation is corroborated by comparison with modern Melanopis data from the Jordan Valley (Zaarur et al., 2016). Unevaporated springs and streams in the Jordan valley have Melanopis shell stable isotope values that partially overlap the W-80 fossil data suggesting that rainwater isotopic compositions were broadly similar. A more evaporated Sea of Galilee datum from Zaarur et al. (2016); blue star on Figure 5) plots on the ‘evaporative/evolved trend’ between the Wisad fossil end members.
Discussion
Regional palaeoclimate
During the Early Holocene of the Near East and Eastern Mediterranean, centennial scale, cold (possibly wetter) winter-focussed, Rapid Climate Change (RCC) events punctuated warmer, dryer interstadials (Rohling et al., 2019). The presence of aquatic snails and the aquatic snail isotopic data from Wisad Pools between ~8.4 to ~7.6/7.5 ka define a ~800-year time window with clear evidence of wetter conditions than today. This is in broad agreement with wetter climatic conditions in northern Arabia between 8.7 and 8.0 ka (Dinies et al., 2015; Petraglia et al., 2020) and recharge ages in the Azraq Basin (Bajjali and Abu-Jaber, 2001). This time window is coincident with the 8.6–8.0 ka RCC event which, downstream of the Aegean and Levantine seas, corresponds to modelled wintertime cold and wet conditions in the coastal Levant (Rohling et al., 2019). While it is therefore tempting to ascribe wintertime wetness between 8.6 and 7.6 ka in the eastern badia to an RCC driven mechanism there is currently no evidence of Holocene speleothem deposition in the rain shadow region east of the central mountains in Israel (Bar-Matthews et al., 2019) and none until at least ~6 ka and mostly after 3.3 ka in northwestern Jordan (S. Robinson pers. comm. 2021). This suggests that even when the coastal Levant was wet, for example during sapropel one (S1) deposition in the early Holocene, Mediterranean moisture did not penetrate east into Jordan. This raises the possibility that early Holocene moisture in the eastern badia was sourced from either more northerly or southerly sources.
It is well established that southern Arabia can receive southerly monsoon-associated precipitation resulting from a northward displacement of the summer Inter Tropical Convergence Zone (ITCZ) (Fleitmann et al., 2007). Such a scenario, based on climate models, has been proposed to explain a significantly wetter than present central Arabian peninsula during the last interglacial (Jennings et al., 2015), that is, increased rainfall sourced from the North African summer monsoon. This mechanism also explains well-documented lake development south of 25°N during the early Holocene (10.5–7.0 ka; Berger et al., 2012). However, the eastern badia is beyond the northernmost penetration of this modelled ITCZ displacement. While it is likely that most palaeoclimate models underestimate northward movement of the ITCZ and coincident increase in rainfall compared to palaeoclimatic reconstructions (Braconnot et al., 2007; Perez-Sanz et al., 2014) it remains to be proven that this source penetrated beyond 30°N.
On balance it seems plausible that Holocene rainfall in the eastern badia was mostly sourced from the north or northwest, that is, by mid-latitude westerlies (Petraglia et al., 2020) that picked up moisture in the eastern Mediterranean (Figure 1). The W-80 aquatic snail δ18O values suggest the isotopic composition of this rainfall was similar to present day (Figure 2) and most likely fell locally because the drainages feeding Wadi al-Qattafi and Wisad Pools both originate on the nearby basalt plateau. This scenario is consistent with an RCC-driven timing of sustained wetness at Wisad Pools between 8.6 and 7.6 ka concurrent with cold and wet conditions in the Levant from 8.6 to 7.8 ka (Rohling et al., 2019). At the regional-scale, wetter conditions are recorded in northwestern Arabia, where ‘grasslands . . . reached their maximal expansion ca 8.600–8.000 cal BP’ before retreating abruptly at ~8.0 ka (Dinies et al., 2015). Pollen records from Tayma indicate the period of ~8.7–8.0 ka to be the wettest conditions of the Holocene (Petraglia et al., 2020) although other proxy records indicate that a freshwater palaeolake at Tayma, present between ~9.3 and 8.5 ka shrank to a wetland environment between 8.5 and 4.8 ka (Petraglia et al., 2020). The available palaeoenvironmental and archaeological data therefore indicate that the whole region experienced climatic variability – with changing availability of surface water particularly in more arid regions – but broadly characterised by wet (and possibly cold) winter conditions between 8.6 ka and 8.0 ka.
Local archaeological and environmental reconstruction
Wisad Pools
There were no terrestrial snails found at W-400 and the isotopic signature of the aquatic snails indicate that they lived in a locale characterised by a highly evaporative hydrological environment. This aligns well with what is understood of the qa immediately north of W-400. Although the sample is small the aquatic snails indicate water must have been periodically available. However, the absence of terrestrial snails from W-400 indicates that the immediate environment was generally very arid.
In contrast, at W-80 both terrestrial and aquatic snails were present although the terrestrial snails all had isotopic signatures characteristic of feeding on C4 vegetation. The presence of terrestrial snails at W-80 may thus indicate an environment better vegetated than W-400, but one that was still relatively arid. The aquatic snails found in W-80 were presumably transported in on plants growing in nearby pools and their δ18O values indicate relatively unevaporated freshwater. Thus, between 7.6 and 8.6 ka, Wisad Pools may have looked rather similar to Azraq today with the general landscape being arid, while the pools themselves supported lush, marshy, Mediterranean type vegetation (pollen dated by OSL to 7.9 ± 0.7 ka; Jones et al., 2021; Rowan et al., 2017) that included C3 deciduous species such as Quercus, Prunus, Tamarix sp. and Salix sp. (Rowan et al., 2017; Wasse et al., 2022).
The reason why pool-marginal plants, perhaps rushes and reeds, were brought into the structures can only be surmised. However, prior to modern building materials becoming available, Levantine bedouin are known to have used reed matting to roof their dwellings (Supplemental Figure S4) (Hula reed hut 1920, https://en.wikipedia.org/wiki/Hula_Valley)
Wadi al-Qattafi
Wadi al-Qattafi today is hyper-arid like Wisad Pools but both the molluscan and isotopic data suggest that during the 9th millennium BP it may have been marginally more humid than Wisad. The fossil snail P. coenopictus is much more abundant at M7 SS-1 than at Wisad (Table 1); moreover, some of their δ18O values were markedly negative which imply less arid conditions with decreased water stress and increased relative humidity. This is also supported by the much larger number of terrestrial snails found at M7 SS-1 and the presence of the semi-slug D. saulcyi; this feeds on earthworms, which require humic conditions. It should be noted that the snail samples from M7 SS-1 are all from deposits dating to ~8.4 to 8.2 ka.
One important observation arising from the discussion above is that at Wisad Pools snails were present in contexts postdating the regional 8.6–8.0 ka RCC. This is not the case at Wadi al-Qattafi where snails were found only within the chronological window of the 8.6–8.0 ka RCC. This, combined with an isotopic signature suggesting decreased water stress and increased relative humidity at Wadi al-Qattafi in comparison to Wisad Pools, supports wetter conditions in the region prior to 8.0 ka. Notwithstanding the inexplicable absence of snails in pre-8.0 ka contexts at Wisad Pools (Figure 4), it is not surprising that snails continued to survive later (albeit in more water-stressed conditions) in an environment where the pools may have continued to contain water well after the wider region became more arid. This supports the theory mentioned in the introduction that considerable environmental and hydrological variation has had the tendency to mask regional patterns of human and environmental responses to periods of RCC.
Conclusions
The presence of aquatic snails at two sites in the Jordanian badia (dating between ~ 8.4 and 7.6 ka) define a ~800-year time window when wetter conditions than present day prevailed. Water courses were established and C3 vegetation grew on the marshy margins, while the wider environment was largely arid stony desert dominated by C4 vegetation. These inferences are consistent with a sustained, RCC-driven wetness between 8.6 and 8.0 ka concurrent with cold and wet conditions in the wider Levant. While the isotope data are not diagnostic regarding moisture provenance, the timing is consistent with a north or north westerly source from mid-latitude westerlies that picked up moisture in the Eastern Mediterranean.
In an archaeological context our pilot study shows the importance of aquatic snails in the reconstruction of semi-arid environments. The combination of terrestrial and aquatic species, combined with plant fossil data, allows a detailed reconstruction of land cover in the immediate vicinity of the structures, as well as land cover around nearby water sources. The difference in the stable isotope signatures of the qi’an and the pools allow for a nuanced reconstruction of a varied but mostly arid environment. The most negative δ18O signal in these aquatic snails, are closest to an equilibrium value, and are consistent with modern regional rainwater isotopic compositions, while the more positive values record the effects of evaporation. Such fine-grained environmental information is not available from the terrestrial snail-shell data when taken alone.
Methodologically, we have identified a bridge between differences in scale of palaeoenvironmental proxies for use in archaeology. Clarke et al. (2016) highlighted the lack of environmental proxies local to archaeological sites that can be used to reconstruct environments relative to human generations. This study has demonstrated that when snails with appropriate ecologies are collected from well-dated contexts, supporting isotopic data can inform on environmental change at the kind of localised spatial, and short-term temporal scales that should be broadly compatible with the lived experience of past human communities. As human populations would have chosen to modify (or not) their cultural and economic practices in response to locally observable environmental circumstances, the approach presented here offers a means to address the gap between large-scale environmental proxies, and human action as evidenced through changes in the archaeological record at individual sites. In that sense it may start to close one of the gaps in the chain of evidence that was noted by Clarke et al. (2016).
Supplemental Material
sj-docx-1-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-docx-1-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Supplemental Material
sj-jpg-5-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-jpg-5-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Supplemental Material
sj-jpg-6-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-jpg-6-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Supplemental Material
sj-jpg-7-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-jpg-7-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Supplemental Material
sj-jpg-8-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-jpg-8-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Supplemental Material
sj-xlsx-2-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-xlsx-2-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Supplemental Material
sj-xlsx-3-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-xlsx-3-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Supplemental Material
sj-xlsx-4-hol-10.1177_09596836221138345 – Supplemental material for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites
Supplemental material, sj-xlsx-4-hol-10.1177_09596836221138345 for Local-scale environmental gradients in ‘snail-shell’ stable isotopes from Holocene Jordanian archaeological sites by Holly Jenkins, Julian Andrews, Yorke Rowan, Alexander Wasse, Tom White, Graham Philip, Alina Marca and Joanne Clarke in The Holocene
Footnotes
Acknowledgements
The analyses and conclusions reached in this paper would not have been possible without the valued input from colleagues, in particular Professor Gary Rollefson, Dr Brita Lorentzen and Dr Jennifer Ramsay of the Eastern Badia Archaeological Project (EBAP). Financial help was provided by the Palestine Exploration Fund, The Oriental Institute of the University of Chicago and the University of East Anglia. We thank in particular Sam Royle, who helped developed the isotope work as part of his post-doctoral research and Matthew Jones (Nottingham) for sediment samples. Emma Hooper, Linda Turner and Bertrand Leze (UEA) provided laboratory technical support in the School of Environmental Sciences (UEA). We also thank the editor and journal reviewers whose comments helped clarify our interpretations and presentation.
Author contributions
Conceptualisation (JA, JC); Methodology (HJ, JA, JC); Validation (HJ, JA, AM); Formal analysis (HJ, JA, TW, AM); Investigation (HJ, JA, GP, YR, AW, TW); Data curation (HJ, JA, TW, AM); Writing original draft (HJ, JA, JC); Writing, review and editing (JA, GP, YR, AW, TW, JC); Visualisation (JA, JC); Resources (YR, AW); Supervision (JA); Project administration (JC); Funding acquisition (JC).
Declaration of conflicting interests
The authors report no competing interests as defined by The Holocene, or other interests that might be perceived to influence the interpretation of the article. The authors have no non-financial competing interests, or other interests that might be perceived to influence the interpretation of the article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.