
Abstract
The site of Körtiktepe in southeastern Turkey is one of few sites in the Upper Mesopotamia basin that attests continuous, permanent occupation across the boundary from end of the colder, drier Younger Dryas (YD) into the comparatively wetter and warmer Early Holocene (EH). This allows for the study of the degree of environmental change experienced on a local level over this boundary as well as for the study of the adaptations that the occupants of the site undertook in response to these changes. The mammal assemblage of Körtiktepe remains relatively stable across the YD – EH transition with the main contributors to diet being mouflon (Ovis orientalis) and red deer (Cervus elaphus) in approximately the same quantities, although the contribution of aurochs (Bos primigenius) increases in the EH. The most significant changes can be seen in the shift in avifauna remains, with a sharp increase of waterbirds during the EH. It is proposed that these shifts reflect changes in the local environment with an increase in woodland cover as well as expansion of local waterways, which is generally consistent with previously published archaeobotanical studies. In terms of species exploited, mortality profiles as well as size distribution of mammals, a great deal of continuity is observed. This suggests that over this particular period the local impact of the beginning of the Early Holocene was not overly dramatic, allowing for cultural continuity of previously established subsistence strategies.
Keywords
Introduction
Excavations in the last decades have revealed the importance of the Upper Mesopotamian basin in understanding the Epipalaeolithic-Neolithic transformation that took place in southwest Asia during the first half of the 10th millennium BC. The transition from the Late Pleistocene to Early Holocene saw the beginning of numerous lasting changes in socio-cultural behaviour including an explosion in visible symbolic iconography and the movement to a plant cultivation and livestock husbandry-based subsistence system. Whilst research into early farming communities in the Near East was for many decades concentrated in the Southern Levant, the finding of sites such as Çayönü Tepesi in the 1960s (Braidwood et al., 1969, 1971), followed by a wealth of recent work as part of rescue excavations preceding the construction of numerous hydroelectric dams in southeastern Turkey has revealed the extent and nature of early settled communities in this region (Figure 1). The excavation of Pre-Pottery Neolithic (PPN) sites such as Nevali Çori (Hauptmann, 1988, 1999, 2011) and Hallan Çemi in the 1990s (Rosenberg and Davis, 1992; Rosenberg et al., 1995), as well as more recently: Hasankeyf Höyük (Hongo et al., 2019; Miyake et al., 2012), Gusir Höyük (Karul, 2011, 2020), as well as Körtiktepe (Benz et al., 2015; Özkaya and Coşkun, 2011; Rössner et al., 2018), has demonstrated, contrary to the previous models, that year-round settlements indeed preceded the cultivation of plants and the management of animals (Braidwood and Braidwood, 1953; Childe, 1951: 282; Kenyon, 1965: 44). Archaeozoological work has recently turned its attention to understanding numerous subsistence strategies employed by hunter-gatherers at the end of the Terminal Pleistocene and beginning of the Early Holocene in order to better understand the varied circumstances prior to the transition to an agriculturally based economy as well as to better understand local histories and trajectories (Arbuckle, 2014; Arbuckle and Erek, 2012; Atici, 2009). One of the many fervent debates that remains in the discussion of the transition to control over animal and plant reproduction is the role of climatic change, namely the onset of the Early Holocene (EH) climatic amelioration (from approximately 11,590 BP/9,640 BC) following the Younger Dryas (YD), a period of worldwide abrupt climate change characterised by cooler and dryer weather conditions (Friedrich et al., 2004: 1120). The end of the Younger Dryas brought with it an increase in average temperature and rainfall which resulted in profound changes in vegetation and waterways that led to local changes in plant and animal ecology (Roberts et al., 2001: 732; Willcox et al., 2009: 152).
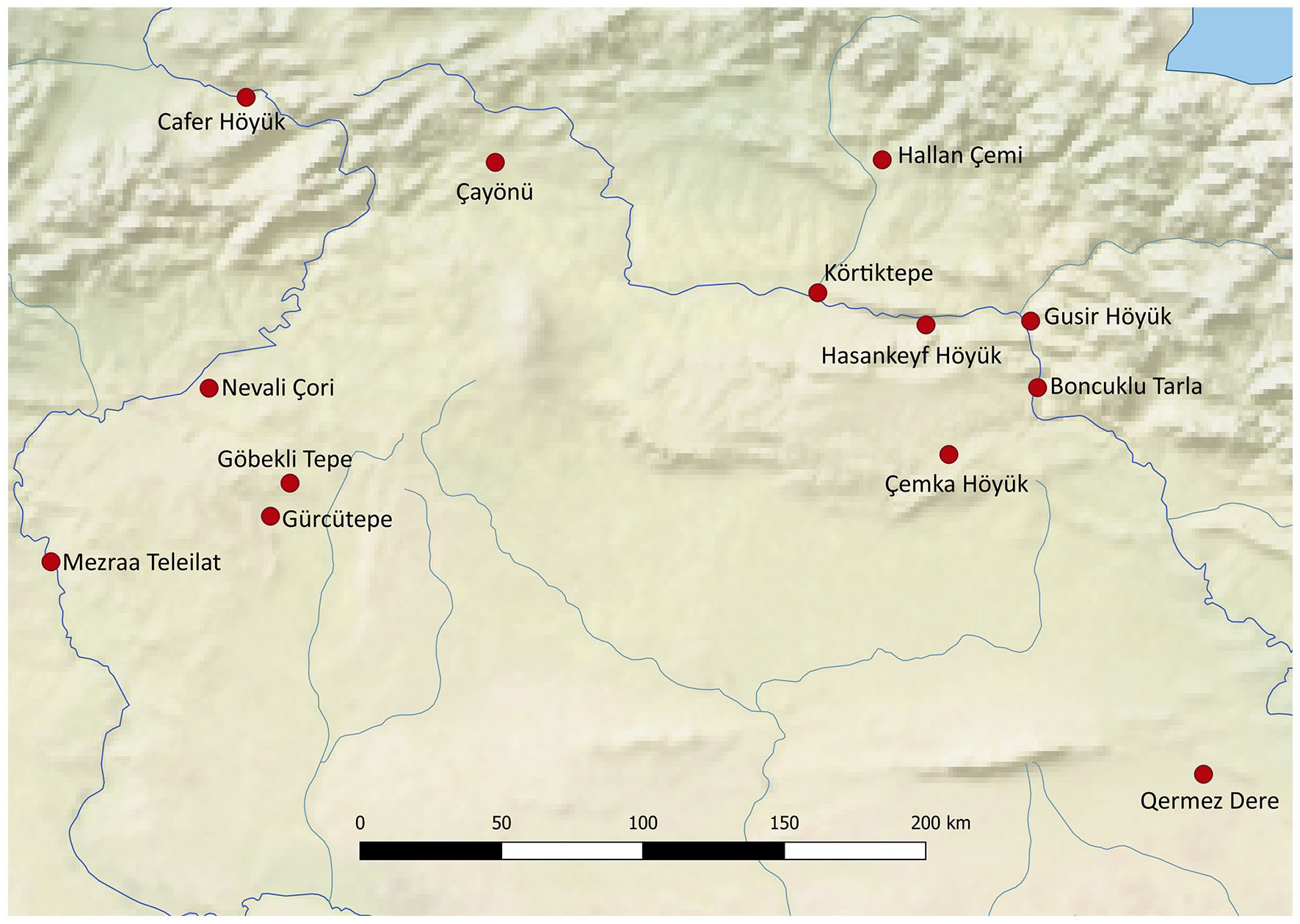
Map of Pre-Pottery Neolithic sites mentioned in text.
Körtiktepe (alongside Hallan Çemi) is the earliest site in Anatolia that appears to be occupied year-round and is one of few sites excavated in this region that attests what appears to be continuous occupation across the Pleistocene-Early Holocene boundary, allowing for the study of the effects of this period of climate change (Benz et al., 2012a: 302, 2016: 231). Presently, only Qermez Dere in northern Iraq has published faunal results spanning this time period, and whilst showing shifts of emphasis across its phases, remains relatively stable in taxonomic composition across its occupation (Watkins et al., 1989, Dobney et al., 1999: 49, Fig 6.2, 55). Additionally, there are the newly found sites of Boncuklu Tarla, which is currently awaiting further excavations and results from zooarchaeological analyses, (Kodaş, 2019a: XII; Kodaş, 2019b: 7), as well as Çemka Höyük which is currently unexcavated and without precise published dating information (Kodaş et al., 2020), both sites being located in the Upper Tigris basin of southeastern Turkey. The site of Hallan Çemi in the foothills of the Taurus mountains, also within the Tigris River basin, has also yielded radiocarbon dates from the end of the 11th millennium BP (uncalibrated) that would correspond to the last part of the Younger Dryas (Rosenberg, 1994; Rosenberg et al., 1998: 27). However, the earliest radiocarbon dates display high standard deviations, and the bedrock of the site was never reached in excavation, so the dates of the earliest occupation layers remain uncertain, although it is considered that settlement would have already started in the terminal Pleistocene (Rosenberg et al., 1998: 28) (For discussion see: exoriente.org/associated_projects/ppnd_site.php?s=26). Zooarchaeological results from Hallan Çemi have in any case largely been published altogether, without chronological differentiation (Starkovich and Stiner, 2009: 49; Rosenberg et al., 1998: 32–33). The exception is Zeder and Spitzer (2016) who differentiated between different building levels, with building level (BL) 3 being the earliest, radiocarbon dated to the very end of the 11th millennium BC/ beginning of the 10th millennium BC (Higham et al., 2007: S22–S23; Zeder and Spitzer, 2016: 142, tab.1). As such, Körtiktepe offers a rare opportunity to investigate the transformation in cultural and subsistence practices that took place at the Pleistocene-Holocene boundary within a single location. Zooarchaeological study across this boundary aims to understand the degree of continuity over the occupation of the site, the manner this climate amelioration impacted on the local fauna, and the response of the inhabitants of Körtiktepe to these potential changes in their local environment and associated animal resources.
Körtiktepe
Körtiktepe (previously published as Körtik Tepe) (37°48′51.90″N, 40°59′02.02″E) is a tell settlement located in southeastern Anatolia within the province of Diyarbakır, Turkey. The mound rises approximately 5 m above the plain with an area of around 100 × 150 m (Özkaya and Coşkun, 2011: 90). Radiocarbon dating suggests that the site has been continuously occupied since at least 10,400/10,200 cal. BC, the Final Pleistocene, into the Early Holocene, dated to 9,600–9,250 cal. BC (Benz et al., 2012a: 293, tab.1, 2015; Coşkun et al., 2012: 28).
The site is situated by the confluence of the Batman Çayı and the River Tigris, within a plain on the cusp of the foothills of the Taurus Mountains. The settlement was likely situated much closer to the Batman Çayı than it is today as well as another palaeochannel previously located west of the site, placing the settlement on a kind of peninsula (Benz et al., 2015: 14). Körtiktepe was identified as an aceramic prehistoric site and excavated as part of rescue excavations carried out ahead of the building of the Ilısu Dam and hydroelectric power plant (Özkaya and Coşkun, 2011: 90). From 2000 onwards, excavations were directed by Vecihi Özkaya of Dicle University (Özkaya and Coşkun, 2011: 90) and between 2009 and 2012, as well as in 2014 and 2015 were joined by German co-operation partners from the Universities of Freiburg, Tübingen and Mainz (Benz et al., 2015: 11; Rössner et al., 2018: 17). In 2010 to 2012, epipalaeolithic occupation layers were reached in some deep soundings of the site (Benz et al., 2015: 11). Detailed reports of the stratigraphy and architecture of the Younger Dryas levels of Körtiktepe have been previously published (Benz et al., 2012b, 2015, 2017; Coşkun et al., 2012).
The buildings excavated at Körtiktepe fall broadly into three categories, round-planned buildings of approximately 2.3–3 m in diameter, smaller structures of approximately 1.1–2.1 m in diameter and a small number of larger buildings with a diameter of 3.4–3.8 m (Özkaya and Coşkun, 2011: 91–92). These have been interpreted as likely representing respectively: dwellings, storage units, and potentially a form of public or ‘special’ building (Özkaya and Coşkun, 2011: 91–92). Already in the Younger Dryas levels occupation is considered to be year-round with a great deal of building tradition showing continuity into the Early Holocene, where it becomes more sophisticated, and the site more densely occupied (Coşkun et al., 2012: 30; Schreiber et al., 2014: 14). Especially of note amongst the artefactual finds are over 500 examples of carved stone vessels, many of which display incised geometric patterns and depictions of plants and animals (Özkaya and Coşkun, 2011: 96; Özkaya and Siddiq, 2020: 46).
One of the most remarkable aspects of the site is the impressive quantity of interments that has been uncovered, around 2000 single and double burials (Özkaya and Siddiq, 2020: 45). Körtiktepe contains one of the largest sets of burial data available in the Pre-Pottery Neolithic Near East, allowing for detailed analysis regarding burial practices, bioarchaeological examination, and grave good analysis (Benz et al., 2016, 2017; Erdal, 2015; Özkaya and Coşkun, 2011: 93). A range of burial customs have been observed including skeletons having been plastered or coloured, with a small percentage showing cutmarks (Erdal, 2015: 7–8).
As Younger Dryas levels were not reached at Körtiktepe until 2011, previous initial reports and short summaries of the faunal assemblage have related exclusively to the Early Holocene (Arbuckle and Özkaya, 2006; Özkaya and Coşkun, 2011: 100–102; Özkaya et al., 2011). Archaeobotanical analysis has revealed the changing character of the local environment, as well as subsistence strategies, between the Younger Dryas and Early Holocene occupation levels (Rössner et al., 2018). The charcoal remains from the Younger Dryas levels largely reflect the local riverine environment (Rössner et al., 2018: Tab.5, 23) whereas the other plant materials show the wider landscape as being dominated by small-seeded grasses, suggesting a relatively open landscape (Rössner et al., 2018: 22). The Early Holocene assemblage, however, shows a shift away from small-seeded cereals towards a more open oak-woodland environment, although the riverine taxa would have still contributed significantly to the plant spectrum (Rössner et al., 2018: 25–26). Possible small-scale cultivation of legumes and cereals is suggested for the Early Holocene due to their slight increased importance, the presence of increased quantities of chaff, as well as cultivation-associated weed flora in the case of the cereals (Rössner et al., 2018: 25–26). As such, the zooarchaeological findings of the study, in combination with the archaeobotanical results from the site, will be of great value in considering the impact on the local ecology of these Early Neolithic settlements at the end of the Pleistocene and in considering the responding adaptations in subsistence strategy and social organisation.
Methodology
The vast majority of the faunal remains presented in this study were excavated in the seasons of 2014 and 2015. Only well-stratified contexts were selected for zooarchaeological identification. Flotation took place on a limited number of contexts that would have been excavated after 2009 (mesh size: 0.2 mm), and additionally numerous microfauna including quantities of small fish remains were recovered from the bone-washing process. Sieving was conducted on contexts associated with graves with sieve sizes of 7–8 mm in addition to sieves 3.5 mm in size. This will naturally bias the results against smaller elements and taxa. However, the collection methods for both Younger Dryas and Early Holocene contexts were the same, so any observed species change is not due to change in methodology.
Identification was carried out with aid of a modern reference collection housed at the Institute for Paläoanatomie, Domestikationsforschung und Geschichte der Tiermedizin at the Ludwig Maximilian University of Munich. Identification of avian remains was conducted using the reference collection of the Staatssammlung für Anthropologie und Paläoanatomie München (SAPM). Multiple bones judged to be coming from the same individual were collectively given a number of identified specimens (NISP) of one. Antler fragments were not included within the statistics as the specimens may have been collected shed. Material was recorded using Ossobook, a specialist database for zooarchaeological findings (Kaltenthaler et al., 2021). Where possible all information relating to taphonomy, ageing or pathology were recorded. Measurements followed von Den Driesch (1976). Differentiation of mouflon and wild goat was based on published literature (Boessneck et al., 1964; Halstead et al., 2002; Zeder and Lapham, 2010; Zeder and Pilaar, 2010).
Tooth eruption and mandibular wear stages (Grant, 1982; Munro et al., 2009; Payne, 1973) were recorded, but due to the small sample sizes, are not presented in this study. It was not possible to construct mortality profiles based on teeth wear and eruption stages for any taxa due to low sample sizes of mandibular teeth, likely resulting from differential transportation of carcass portions. As such, mortality profiles have been reconstructed using post-cranial epiphyseal fusion data, which is only intended to suggest general mortality trends. For caprines, the epiphyseal sequence and approximate age of fusion follows Zeder (2006) based on a modern collection. The percentage of epiphyses fused is expressed for each age category, independent of one other. For the construction of the Körtiktepe mortality profiles for mouflon, both elements positively identified as mouflon, as well as those identified as being mouflon/wild goat (Ovis orientalis/Capra aegagrus) were used. This approach is aimed to increase the sample size of elements used, as well as to mitigate the bias towards elements where distinction between Ovis and Capra are more easily made (e.g. humerus, coxa, metapodials). Additionally, some morphological features used for distinguishing between Ovis and Capra only develop with age, this then allows for the inclusion of these younger specimens where this has not yet developed. It is considered that despite the potential occasional inclusion of wild goat into the mortality profile of mouflon through this method, the overall trends should still be valid as mouflon appears to outnumber wild goat considerably in the assemblage (sheep-to-goat ratio: YD: 28: 1, EH 5: 1). This was also done in the case of the mortality profile constructed for Göbekli Tepe, as no wild goats were found within the assemblage, so all undetermined caprines are most likely mouflon, but in the cases of Gusir Höyük and Gürcütepe II and III, only elements identified as being mouflon were used due to a larger percentage of the assemblage being wild goat.
The use of epiphyseal fusion for red deer (Cervus elaphus) is, however, more problematic than in the case of caprines, with few studies looking into the timing of fusion of the skeleton (Calderón et al., 2019; Habermehl, 1985; Mariezcurrena and Altuna, 1983). The problem is compounded by the relatively wide range of ages when particular epiphyses fuse, which is for instance affected by sex, with females generally fusing at an earlier age than males (Calderón et al., 2019: 205). As such, the fusion age categories of red deer have been collapsed into three phases, being: epiphyses fused by 20 months, those at 36 months and those above 48 months. The elements assigned to each of these categories are summarised in Appendix 1. Individuals under 36 months can be considered juvenile (Calderón et al., 2019: 207).
Due to limited sample sizes, biometric data have been presented using the logarithmic size index (LSI) method, comparing archaeological data against standard modern individuals (Meadow, 1999). The standard animal used for comparison with mouflon is a female published in Uerpmann and Uerpmann (1994). Red deer measurements were compared to a male standard from Upper Bavaria kept in the Staatssammlung für Anthropologie und Paläoanatomie München (SAPM) (SAPM-MA-01295) (Appendix 2). The standard used to compare aurochs is based on a Pinzgau cow from Austria from the same collection (SAPM-MA-01259), published in Manhart (1998): table 103). Measurements for wild goat (Capra aegagrus) and wild boar (Sus scrofa) were too few to present LSI information, but the raw measurements are presented in Appendixes 3–11. In the case of the red deer remains from the Körtiktepe Early Holocene contexts, a number of outlying, smaller values were generated from measurements from the scapula. It is considered that this is due to the scapula being an early fusing bone, which then continues some appositional growth, so despite being fused, may be still subadult (Habermehl, 1985: 36–37; Mariezcurrena and Altuna, 1983: 184, Tab. 54). Due to this possibility, values generated from scapula measurements have not been used for either the Younger Dryas or Early Holocene phases. Measurements from the scapula were included in the Gusir Höyük data, the LSI values being 0.14 and 0.02 (still fusing), well within the general expected range.
In order to assess the impact of the Early Holocene climatic transformation on the taxonomic diversity or heterogeneity of the mammalian assemblage the Shannon entropy, or ‘archaeobiodiversity’ (aD), was calculated (for aD methodology see Pöllath et al., 2008: 65). Heterogeneity in this context is meaning the measure of both richness (the number of taxa present or NTAXA) as well as the ‘evenness’, how evenly the quantity of different taxa are distributed (Lyman, 2008: 175,192). High heterogeneity is correlated with taxonomic richness and evenness, making the identity of any one random individual difficult to predict (Lyman, 2008: 176–177). Archaeobiodiversity was calculated considering mammalian and avian faunal material separately. Within the mammalia, microfauna (smaller than hedgehog) was excluded and caprines were considered together.
Where possible, the results from Körtiktepe were compared with other zooarchaeological datasets from PPN sites in the region, those being: Gusir Höyük, Göbekli Tepe, and Gürcütepe II and III. These sites were selected for comparison due to the availability of data (including measurements, sexing, and epiphyseal data) and comparable recording methodologies.
Gusir Höyük is located beside Gusir lake near the intersection with the Tigris and Botan rivers, in the foothills of the Taurus mountains and, with the currently excavated remains dating to the Early Holocene, although as yet the earliest remains have yet to be excavated (Karul, 2011: 1–2, 2020: 78–79). Preliminary zooarchaeological results from Gusir Höyük provide an example for comparison of subsistence strategies from a nearby, near-contemporary community (Kabukcu et al., 2021; Neuberger et al., 2019). Göbekli Tepe is located approximately 15km NE of Şanlıurfa on top of a limestone ridge rising from the Harran Plain (Schmidt, 1995: 9, 2000: 45). The site consists of a number of buildings, the oldest and largest having rings of T-shaped pillars gathered around two central pillars, many of which are decorated with carvings of animals (Peters and Schmidt, 2004; Schmidt, 2006, 2011, 2012). The site dates to the PPNA through to the middle of the PPNB, but as the majority of the faunal remains come from the infilling of the large buildings, the dating of these remains cannot be certain (Dietrich, 2011). Gürcütepe consists of a series of hills in the Harran plain, 4km SE of Şanlıurfa with settlements II and III dating to the late PPNB (Beile-Bohn et al., 1998: 5, 13–14; Schmidt, 1995: 9). This site provides a comparison with an assemblage considered to consist of largely domestic animals (von Den Driesch and Peters, 1999: 32). The first zooarchaeological data published from Göbekli Tepe and Gürcütepe can be found in von Den Driesch and Peters (1999). Subsequent publications on the archaeozoology of Göbekli Tepe can be found in Peters and Schmidt (2004), Peters et al. (2005) and Pöllath et al. (2018).
The faunal assemblage
Taphonomy
A total NISP of 21,783 was recorded from the Younger Dryas contexts with a weight of 27.8 kg, of which 2950 (27.2 kg) could be identified to a size-category or better taxonomic level. From the Early Holocene contexts, a total NISP of 21,936 (49.1 kg) was recorded, with 4018 (47.7 kg) being identified to a size-category or better level. Overall, the remains were well preserved, with frequent finds of more fragile bones such as bird elements. However, despite the excellent bone surface preservation, the remains were brittle and frequently showed modern breaks, which inhibited, in particular, the taking of measurements. This brittle nature is seen similarly in both the Younger Dryas and Early Holocene contexts with 86.3% from Younger Dryas contexts being unidentified mammal remains and 81.5% in the Early Holocene. Bone surface preservation was also similarly good in both phases (Table 1). Percentage of elements with calcareous deposits on the surface of the bone is, however, higher in the Early Holocene assemblage, which is disproportionately higher on remains of aurochs likely effecting the ability to identify surface modifications (Table 2). The higher percentage of calcareous deposits found on EH remains may be due to the level of the water table at the site. Both rodent and carnivore gnawing is low in both phases and elements with some degree of burning being a little higher in the Younger Dryas phases (Table 2). The good condition of the bones, as well as frequent finds of bones from the same context that could be re-articulated, suggests that the material was rapidly buried after deposition resulting in minimal attritional effects. A small proportion of remains, that are considered to be intrusive, are from domestic species, as determined by their extremely small size (Appendix 12). These intrusive specimens were found in various contexts across the site, at various depths, and were introduced likely as the result of animal or root action. A potential source of these specimens is the medieval remains that are also present on site, and sometimes disturbed the Neolithic layers (Özkaya and Coşkun, 2011: 90).
Percentage of level of bone surface preservation of mammal bones (without unidentified mammal remains) by phase (YD: n = 2509, EH: n = 3123).
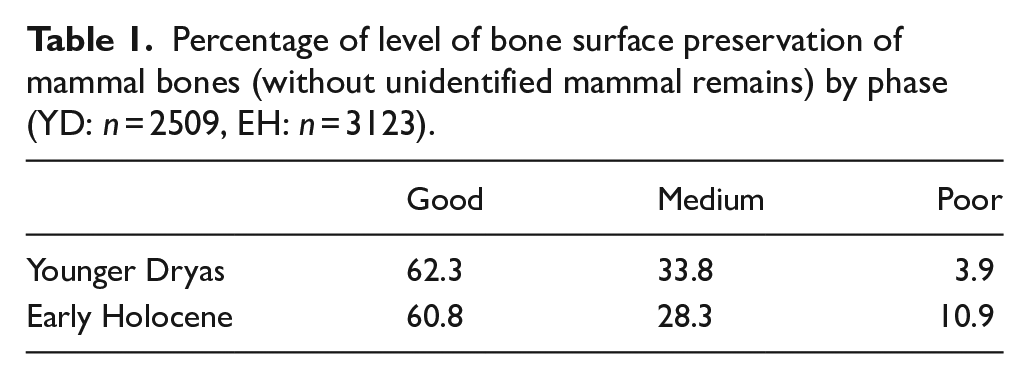
Percentage of mammal bones with taphonomic modifications (without unidentified mammal remains) by phase and species (YD: 2509, EH: n = 3123).
Mammals
The mammal remains from both the Younger Dryas and Early Holocene contexts are summarised in Table 3, the full species list can be found in Appendix 12. In Appendix 12, where a species was recorded as, for example, ‘probably Ovis aries’, this has been combined with the category ‘Ovis aries’.
Identified mammal taxa from Körtiktepe.
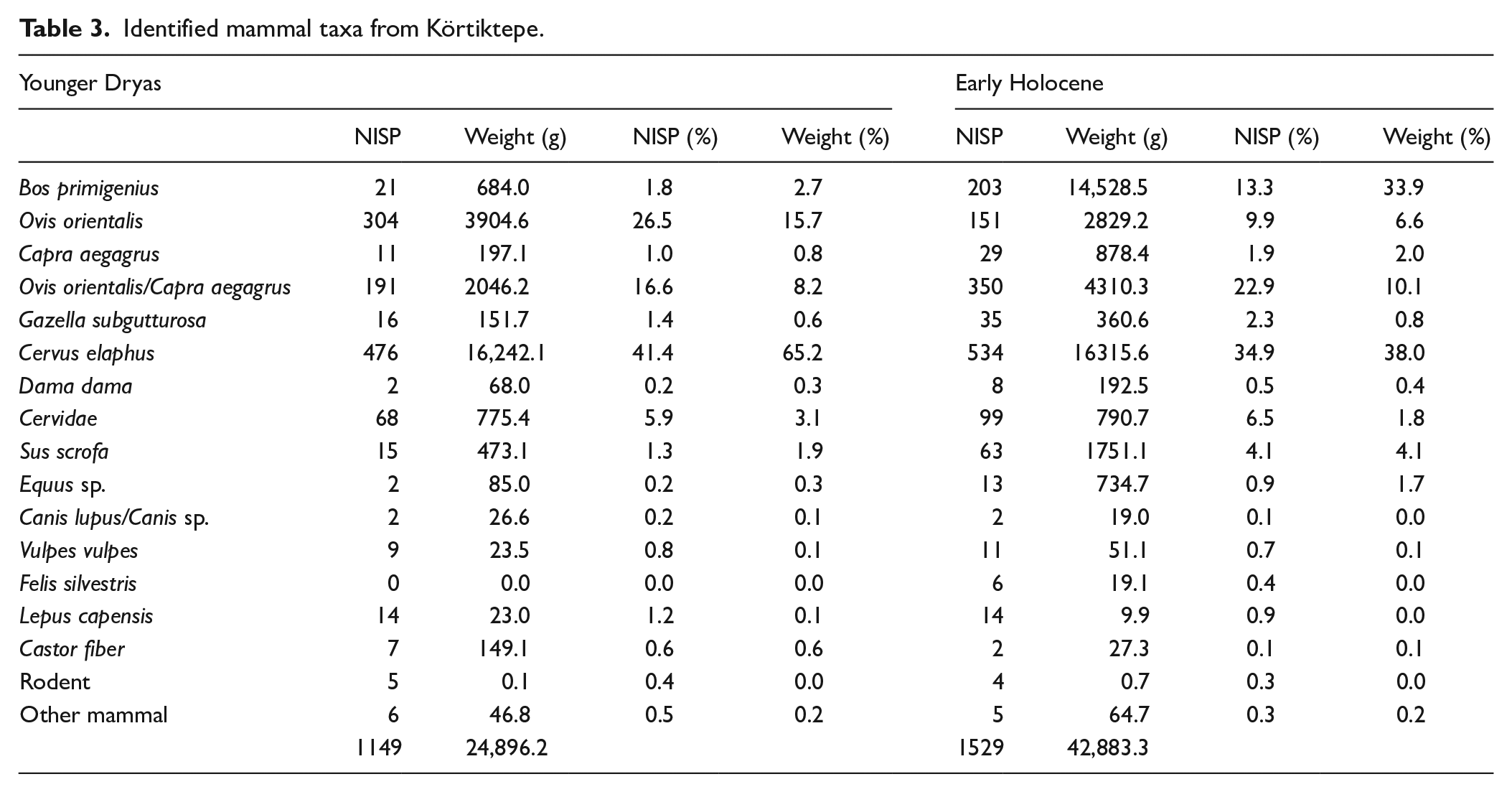
In both the YD and EH phases the most abundant taxon by NISP is red deer, although if it is assumed that the vast majority of the undetermined caprine remains are belonging to mouflon, then in both phases the number of red deer and mouflon remains are found in similar proportions (Table 3). The main changes between the two phases are a marked increase in the importance of aurochs as well as a minor increase in the wild boar (Sus scrofa) in the EH. The increased importance of aurochs is particularly seen when considering the relative weight of the remains which is of a near-negligible amount in the Younger Dryas (2.7%), rising to over one third of the identified mammal remains in the Early Holocene contexts. In both phases mouflon significantly outnumber the finds of wild goat (Capra aegagrus) with the YD having a mouflon to wild goat ratio of approximately 28: 1 and the EH 5 : 1, although the number of wild goat may be too low to consider the change in ratio between phases to be particularly meaningful.
Size reconstruction
Red deer
The size of red deer remains relatively stable across the late Pleistocene/Early Holocene boundary with the YD having a median LSI value of 0.08 and the EH 0.07 and a mode of 0.09, 0.08 and 0.08 respectively. However, whilst the upper end of the LSI range is the same for both the YD and EH data (0.15), a number of smaller individuals are present in the EH (Figure 2). The Younger Dryas data appears to show a dimorphic distribution with a peak that would correspond to the males at 0.08 and 0.09 (Figure 2). A second, smaller peak is also perhaps seen at 0.05 which may represent a lesser number of females being hunted. In the Younger Dryas there is a clear pattern of more males being hunted, whereas in the Early Holocene there is a slight shift towards an increasing proportion of females being represented in the assemblage. In the Early Holocene data, the same larger peak at 0.09 is also seen, perhaps representing the males, and a second peak being at around 0.06, although it is not as clear as in the Younger Dryas data. Too few remains were able to be sexed in order to be used to help reconstruct sex-specific patterns, with the coxae from the YD levels numbering 1 male, 6 female and 2 probable females. In the Early Holocene 4 male were identified, 6 female and 1 probable female. The LSI data from Körtiktepe was compared with the nearby late PPNA site Gusir Höyük. The overall LSI size range exhibited is similar in both sites, having a median LSI value of 0.07 and the mode for Gusir Höyük being 0.07, against 0.08 in Early Holocene Körtiktepe. Gusir Höyük has a more unimodal distribution than Körtiktepe, which may be the result of different hunting strategies, perhaps with Gusir Höyük having less of an emphasis on large males. Too few measurements for red deer were available from Göbekli Tepe and Gürcütepe II and III for comparison.
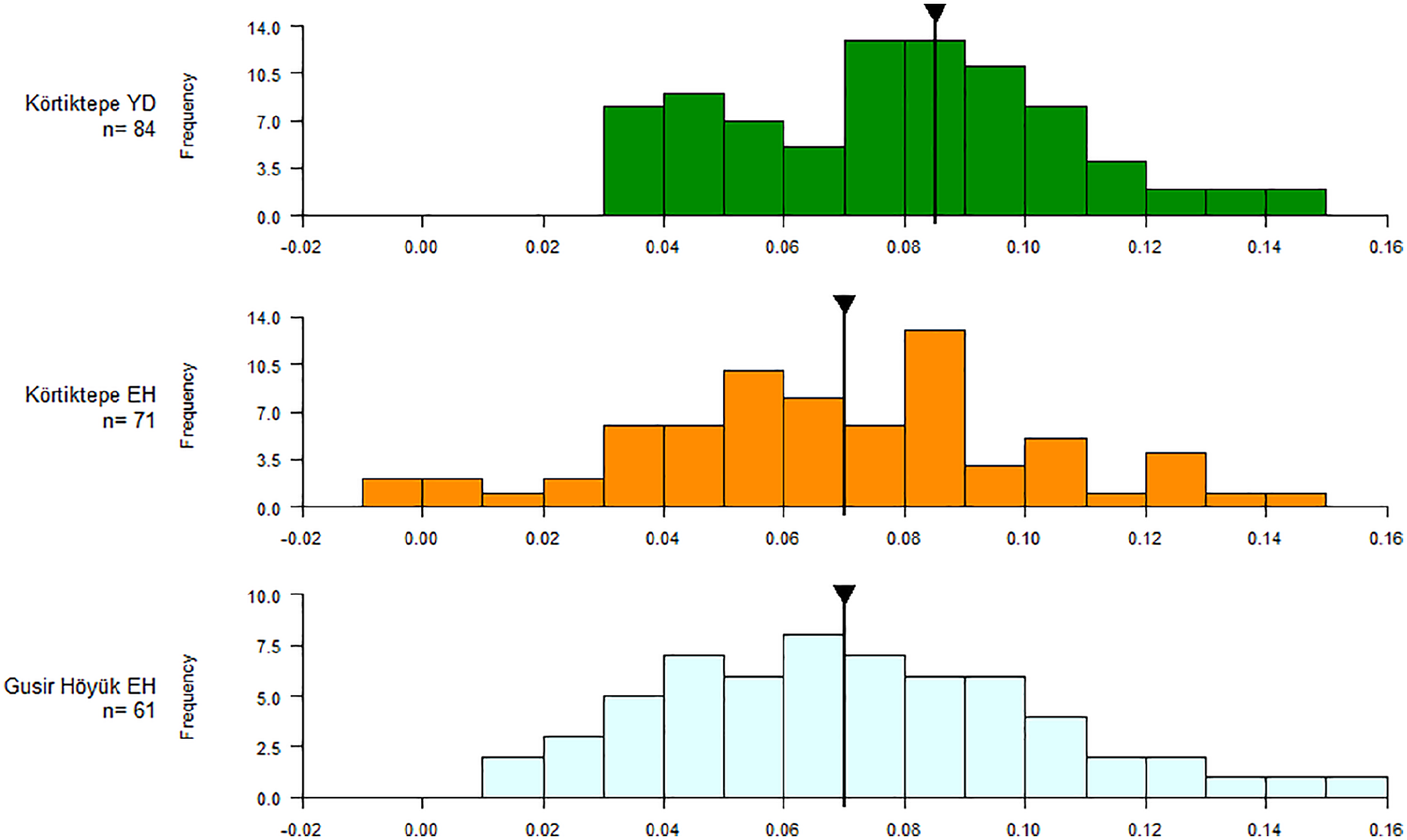
LSI values for red deer (Cervus elaphus) from YD and EH Körtiktepe and Gusir Höyük. The black line and arrow represent the median value.
Mouflon
In both the Younger Dryas and Early Holocene contexts mouflon at Körtiktepe have a median LSI value of 0.07 and mode of 0.08. Both data sets show an upper peak at 0.08, likely representing the males (Figure 3). In the Younger Dryas a peak perhaps representing the females appears to be around 0.04 and 0.05, and whilst less convincingly present in the Early Holocene, there also appears to be this second peak at around 0.05. Whilst generally there is a lot of continuity between the two periods, there is perhaps a perceptible shift towards more females represented in the Early Holocene. Sexed skeletal elements are only few in number but show an even split between the sexed with the YD having five males, five females and one that was probably female. The EH levels likewise show a near even split with five females and four males. The size ranges from Körtiktepe fits well with the size ranges observed at the late PPNA site of Gusir Höyük, the site having a median of 0.06 and mode of 0.08, with 0.08 also likely being the upper peak, representing rams (Figure 3). A similar range is also expressed at Göbekli Tepe, although there appears to be more females than males, a tendency also expressed in the sexed coxae with 10 elements identified as being female to six being male. As such, the median and mode are much lower both being 0.06. Whether the emphasis on female mouflon from Göbekli Tepe is the result of targetted hunting, or from the remains representing early animal management is a topic for future study. In comparison, at LPPNB Gürcütepe II and III, where the majority of Ovis sp. are from domesticated animals, the median and mode are again considerably lower at 0.01 and 0.02. The size of mouflon at Körtiktepe fits well with the broadly contemporary Round-Grill building subphases at Çayönü and at PPNA Hasankeyf (von Den Driesch and Peters, 1999: 28, tab.4; Hongo et al., 2005: 117, fig.3; Hongo et al., 2019: 165, Fig.7a). Gradual size diminution at Çayönü in sheep is seen from the Channelled subphase onwards with a definitive shift in the Large Room subphase (LPPNB) representing the beginnings of caprine husbandry at the site (Hongo et al., 2005: 116–117, 120). As such, the sizes of mouflon of Körtiktepe are clearly in the size ranges of the wild animals of the region, with no indication yet of the size diminution evidenced at other sites indicative of animal management taking place.
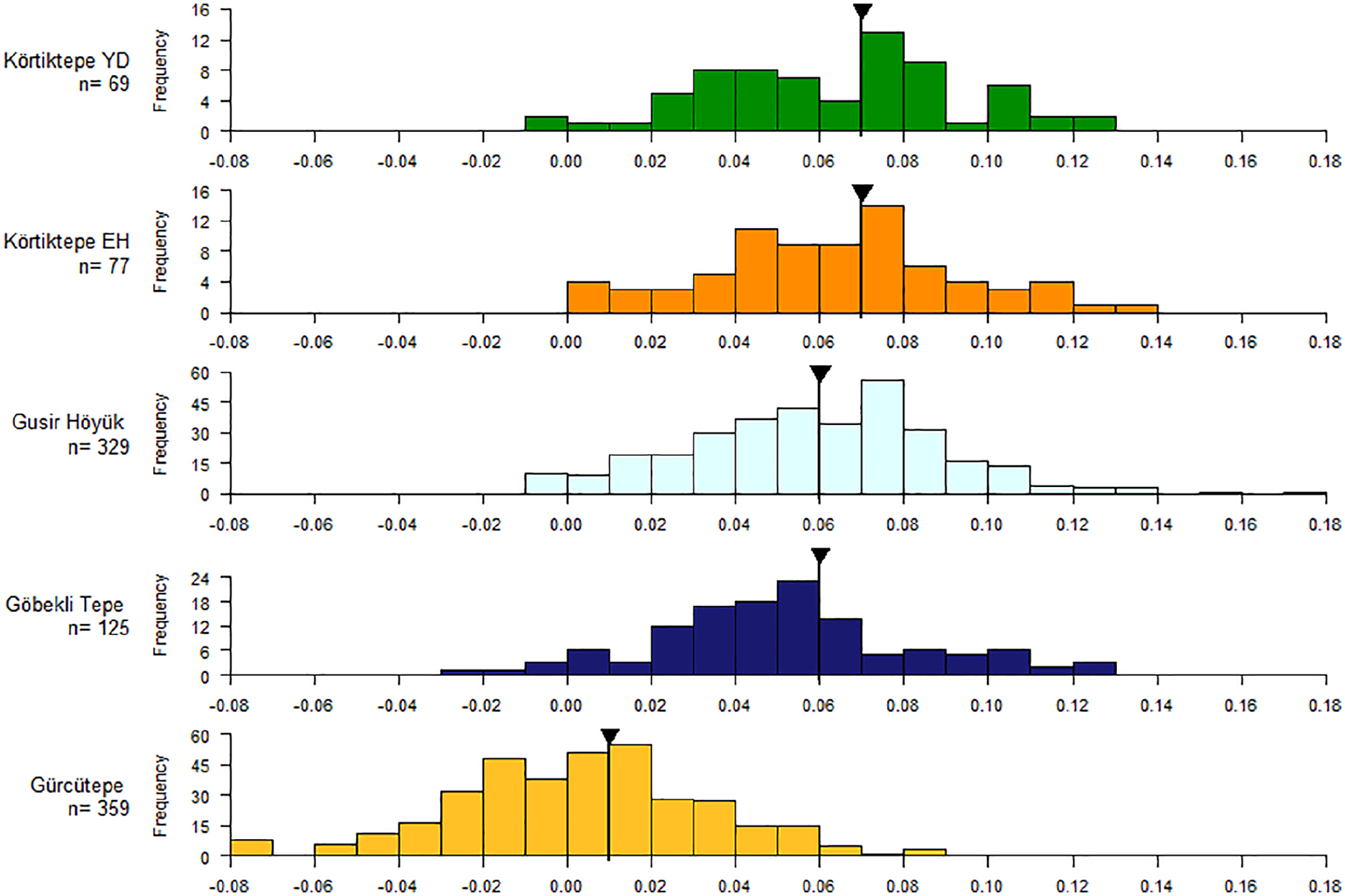
LSI values for mouflon (Ovis orientalis) from YD and EH Körtiktepe, Gusir Höyük, Göbekli Tepe and Gürcütepe II and III. The black line and arrow represent the median value.
Aurochs
Due to the small number of aurochs (Bos primigenius) remains identified from the Younger Dryas levels only one measurement was available for LSI which gave the value (0.00), putting it on the lower end of the expected size range. From the Early Holocene contexts, one value was considerably smaller than would be expected (−0.04, from a fused calcaneus) and so is presumed to be intrusive cattle and removed from considerations. Other than this specimen, the aurochs at Körtiktepe display a wide variation in size from an LSI value of 0.01–00.22. Overall, Early Holocene Körtiktepe, Gusir Höyük and Göbekli Tepe show relatively similar sizes with the medians being 0.11, 0.13 and 0.10 respectively. Körtiktepe, Gusir Höyük and Göbekli Tepe all have a mode of 0.10 (Figure 4). Although only a small number of measurements are available from the previously published PPNA round building phase of Çayönü (n = 9), the general size also appears to be similar, having a median of 0.12 (Öksüz, 2000: 162, tab.1). This is compared against LPPNB Gürcütepe II and III, by which time it is considered that cattle have come under human control (Peters et al., 2017: 9). The size range is, however, roughly similar to that of the wild animals, also with a median of 0.10, which has previously been attributed to a lack of genetic isolation of the managed animals (von Den Driesch and Peters, 1999: 30). Sexed elements from Gürcütepe suggest a predominance of females with two male and nine female coxae being found, with a general shift chronologically towards more females being interpreted as a potential result of a degree of human control (Peters et al., 2017: 9). No sexed elements were available from Körtiktepe, so whilst the domestication status cannot be independently determined from either size or sex ratio, from what is known in the region, it is considered that these specimens represent wild individuals.
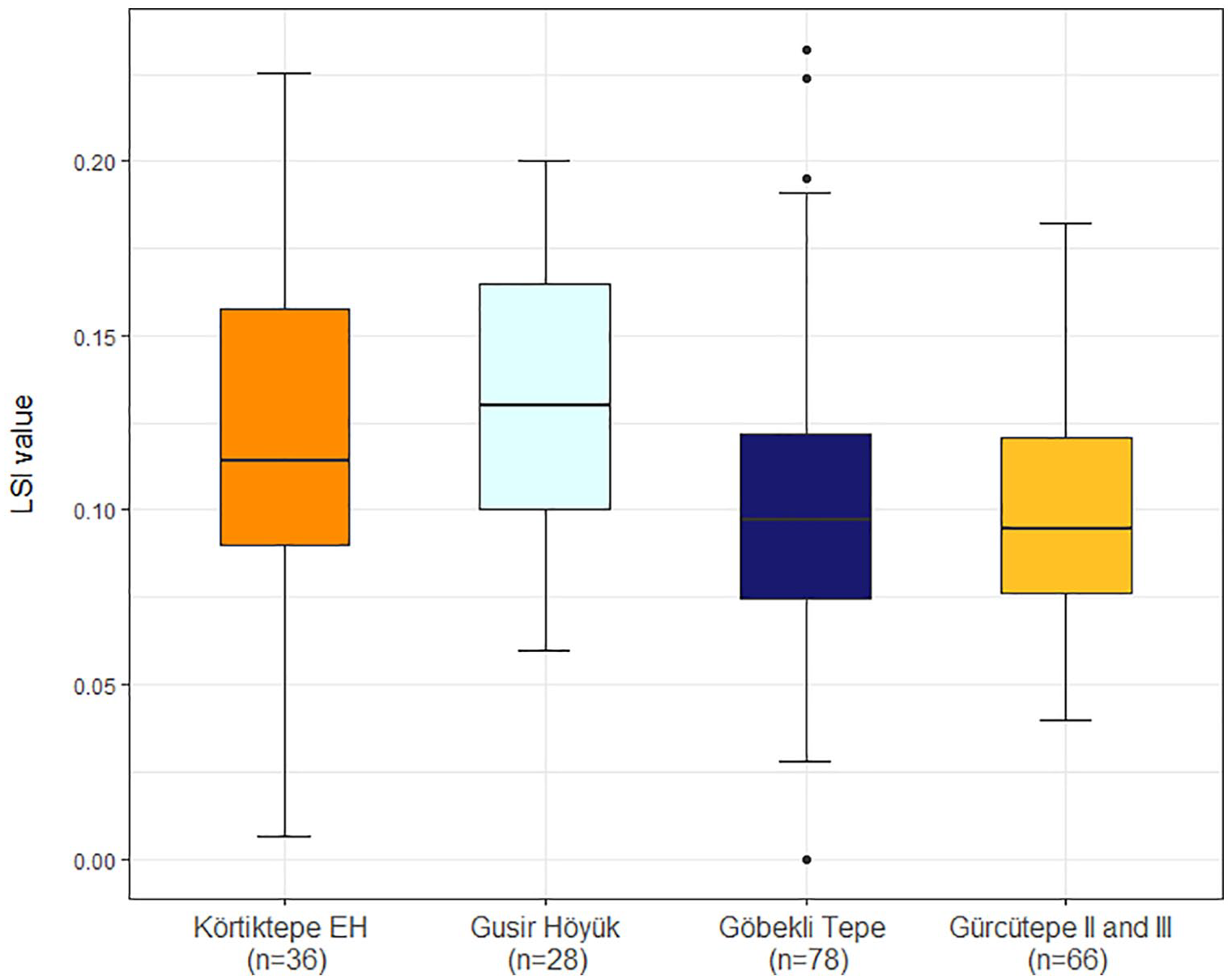
Box plot of LSI values for Aurochs (Bos primigenius) from the Early Holocene contexts of Körtiktepe and Gusir Höyük, Göbekli Tepe as well as the LSI values of Bos taurus and Bos taurus/Bos primigenius from Gürcütepe II and III.
Mortality profiles
Red deer
Whilst only a rough picture can be gained from the mortality profiles construction from post-cranial information, generally in both the Younger Dryas and Early Holocene phases, relatively few juvenile individuals are represented, with the majority making it past 4 years, suggesting the targetting of prime-aged individuals (Figure 5). An attritional or ambush-hunting model would more closely resemble the living structure, with a greater emphasis on younger individuals than that has been exhibited here (Stiner, 1990: 323). These general trends are found in both the Younger Dryas and Early Holocene levels, indicating that hunting strategies did not greatly change over this transition. At Gusir Höyük few young individuals are represented, and few also made it past 4 years of age, suggesting the targetting of young adult individuals. The LSI measurements from Gusir Höyük, as well as the sexed elements (excluding antler), suggests a slight emphasis on the hunting of females (male: 6, female: 10). Too few red deer remains from Göbekli Tepe and Gürcütepe were recovered in order to provide a comparison.
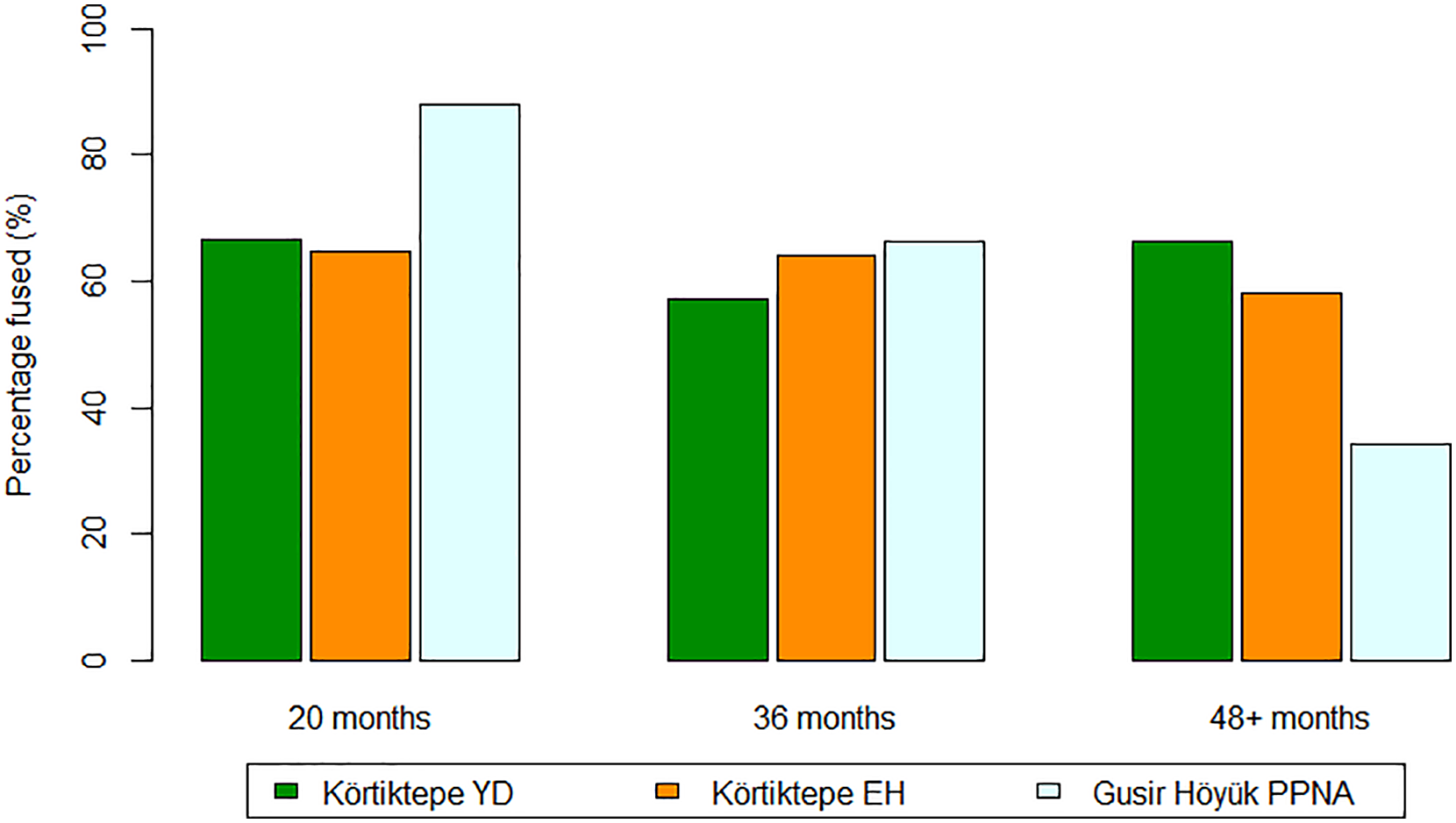
Mortality profile for red deer based on post-cranial fusion. Körtiktepe Younger Dryas (n = 192), Körtiktepe Early Holocene (n = 162) and Gusir Höyük Early Holocene (n = 234) ‘Percentage fused (%)’ refers to the proportion of elements that would fuse at that age group, it is independent of other age groups. The elements included in each epiphyseal group are given in Appendix 1.
Mouflon
In both phases of Körtiktepe, relatively few infantile individuals were identified with instead there being steady attrition of the age classes, until in both cases only just over half are making it into maturity (Figure 6). Generally, with caprines, after a high mortality of lambs, survivorship of yearlings and older age-classes is high (Hesse, 1982: 407). The significant proportion of sub-adults here suggests that nursery herds may have been targetted by hunters. LSI data suggests that amongst adult animals, males were predominantly targetted. As such these mortality profiles might be the result of the hunting of a mixture of both nursery herds that would contain females and juveniles as well as either bachelor herds or lone males. As in the case of red deer, the overall profile remains relatively similar in both the YD and EH levels. At Gusir Höyük, survivorship of sub-adults is even higher, but there a significant drop-off by 3–4 years, suggesting the targetting of adult individuals. As suggested by the LSI results, sexed elements (mostly coxae, but also based on sexed atlas and axis vertebrae and horncores) also show that males were particularly targetted, with over double the number of males identified than females (male: 67, female: 28, probably male: 1, probably female: 1). This suggests that the hunters of Gusir Höyük particularly targetted bachelor herds and/or lone males. Göbekli Tepe also exhibits a similar pattern with the main targets being individuals around 2 years and as well as adults above 3–4 years. Unlike in the other two sites however, the sexed elements suggest a similar ratio of males and females (male: 15, female: 19). In contrast, the post-cranial remains from LPPNB Gürcütepe II and III show a high survivorship into maturity and sexed elements shows a slight preference of females (male: 91, female: 112). Previously published mortality profiles constructed using tooth wear data from Gürcütepe II, however, indicates that the majority did not reach 2 years of age (Peters et al., 2017, 8: 16.2). This discrepancy in results between the tooth wear and epiphyseal fusion data maybe be due to taphonomic considerations such as the role of dogs on the site. Dogs are likely to preferentially select the softer elements of juveniles, particularly the meatier post-cranial elements, and carry them off-site for consumption. The consumption of domestic animals by dogs at Gürcütepe has already been strongly indicated by isotopic data that suggests that dogs consumed remains from animals that were pastured with C4-plants (Grupe and Peters, 2011: 74). This in opposition to the signature demonstrated by the wild animals of the site that were feeding exclusively on C3 plants, such as cereals (Grupe and Peters, 2011: 74). The preferential destruction of juvenile post-cranial elements by dogs may lead to this result where the mortality profiles constructed by tooth-wear or post-cranial fusion may widely differ. With this in mind, post-cranial fusion-based mortality profiles for all sites should be considered cautiously. Nonetheless, from what can be considered, both phases at Körtiktepe suggest an emphasis on male adults, indicating that the assemblages are the result of hunting rather than the early management of mouflon.
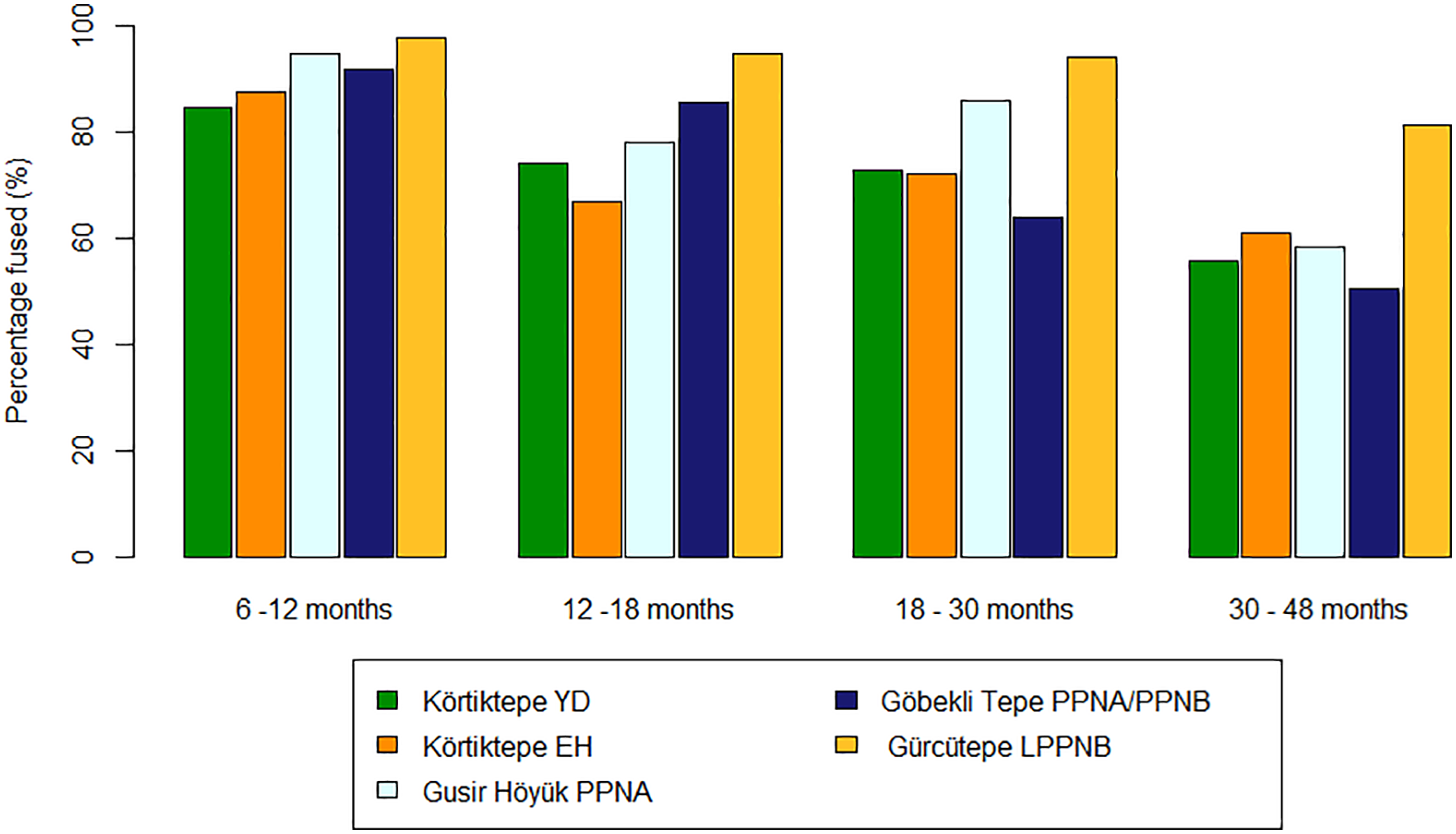
Mortality profiles for combined mouflon (Ovis orientalis) based on post-cranial fusion (Zeder, 2006: 107, fig.15) Körtiktepe Younger Dryas (n = 118), Körtiktepe Early Holocene (n = 149) Gusir Höyuk Early Holocene (n = 359), Göbekli Tepe (n = 363) and Gürcütepe (n = 347). ‘Percentage fused (%)’ refers to the proportion of elements that would fuse at that age group, it is independent of other age groups.
Wild goat
In both the Younger Dryas and Early Holocene, the sample size for wild goat was too small to make anything but very general observations about the slaughter profile. In both cases very few juveniles were found (Appendix 13). Little sexing information is available; one wild goat was sexed as female from the Younger Dryas, and one female, one probable female and four males were sexed from the Early Holocene contexts. The small numbers of goat and slight tendencies towards older and male individuals perhaps suggests that these findings are the result of hunting of lone males or small ram bands.
Aurochs
No fusion information was available for the Younger Dryas due to limited sample size. The remains from the Early Holocene only gave a small dataset to consider mortality profiles for aurochs, which is presented in Appendix 14.
Wild boar
In the case of wild boar, there were again too few specimens from the Younger Dryas to consider age of death, and only a small number from the Early Holocene, these are summarised in Appendix 15. A number of quite young individuals are presented including one ulna from a neonate. Little sexing information is available, with just a find of a mandible and a lower canine, both from males. Dominance of an assemblage of juveniles does not necessarily reflect any sort of human management and instead may reflect a natural kill-off pattern from hunting (see discussion on the Hallan Çemi boar remains (Lemoine, 2012; Peters et al., 1999: 40–41; Starkovich and Stiner, 2009: 56).
Avifaunal remains
Bird remains from the Younger Dryas and Early Holocene levels are summarised in Figure 7, with the full species list found in Appendix 12. In both phases the Great Bustard (Otis tarda) is the most commonly found species, comprising around one third of the assemblages. Between the Younger Dryas and Early Holocene there is, however, a major shift towards the exploitation of waterbirds, which increases from 20.3% in the YD to 59.6% of the EH bird assemblage. This is largely driven by the Greylag goose (Anser anser), which increases from 3.8% to 27.1%. In contrast, in the Younger Dryas layers there is more of an emphasis on raptors which makes up 26.6% of the YD remains, as opposed to 6.0% in the EH. Despite the smaller assemblage size of bird remains coming from the YD, the range of bird species represented is wider with 24 different species identified compared to 19 in the EH.
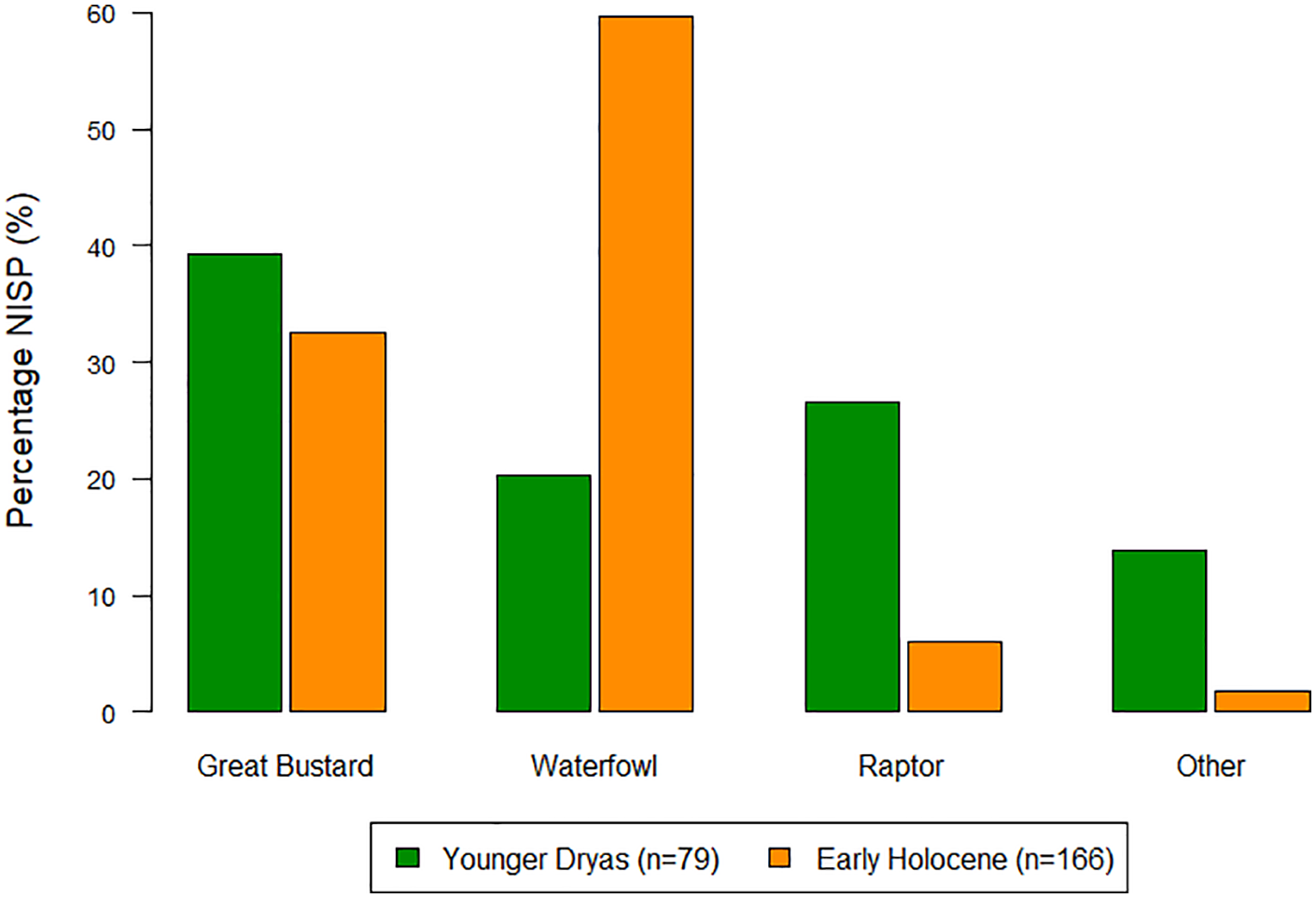
Overview of avifaunal remains from Körtiktepe.
Seasonality
Seasonality has been exclusively addressed through the presence of avian species, due to a lack of suitable data from the mammalian assemblage. Due to a general lack of cranial elements being found at Körtiktepe, addressing seasonality through mammalian tooth wear (as per. Gourichon, 2004) is not possible. There are also very few neonate specimens found on site (one neonate bone from the YD level and one from the EH, both being caprine humeri) and so shows some evidence for winter/spring hunting activities. However, the lack of neonate remains might be easily explained by a lack of targetting of very young individuals, or a lack of survival of young bones due to a variety of taphonomic reasons. Additionally, at this time isotopic data for the animals of Körtiktepe has not been published, so seasonality cannot be addressed through oxygen and strontium isotopic signatures (e.g. Lang et al., 2013).
Avifaunal assemblages can help establish the seasonality of archaeological sites by considering the migratory habits of the various species present. This is a particularly important issue to consider for this period of time as Körtiktepe is considered to be one of the earliest sedentary sites in the region (Özkaya, 2009: 3–4; Schreiber et al., 2014). The geographical limits of ancient populations of birds as well as migratory patterns are in most cases based on what is known about the species’ modern habits. In a number of cases however, modern migratory patterns and population extent will be different from those seen at the end of the Pleistocene and in the Early Holocene due to changes in climate, as well as through human action such as over-hunting or the destruction of habitats. This problem is compounded by gaps in knowledge on population extent and migration patterns of modern birds in some parts of the world, with many areas of southeastern Turkey having only been systematically surveyed relatively recently. As such, modern seasonality data must be considered with a great deal of caution.
Seasonality at Körtiktepe has been addressed in previous publications, with year-round occupation suggested on the basis of substantial architectural investment at the site, presuming that the remains of a seasonal camp would have been more ephemeral, as well as evidence from strontium isotopic data (Benz et al., 2016) (although see Asouti and Fuller, 2013). Using avifaunal data, Arbuckle and Özkaya (2006) concluded that Körtiktepe shows evidence of multiple-season occupation, on the basis of the presence of numerous winter visitors to the site, as well as the presence of the mallard (Anas platyrhynchos), considered to be a summer visitor (based on Harrison, 1982). Taking into account more regionally specific and modern survey data however, we would consider the mallard to be a year-round resident, as well as a winter visitor (Biricik and Karakas, 2012: 99, tab.2; Karakaş, 2010: 474, tab.2; Karakaş and Kiliç, 2004: 304, tab.2). Additionally, Özkaya and Coşkun (2011) argue that seasonality is implied by the presence of the greylag goose and the great bustard, which are breeding in the region in spring and summer (Özkaya and Coşkun, 2011: 101). Nowadays there are only small groups of the greylag goose that ‘probably represent no more than relicts of a once much more widespread breeding population’ in the region (Scott and Rose, 1996: 10), but generally these populations are sedentary or ‘locally dispersive’ (Scott and Rose, 1996: 73). Whilst it has previously been stated that the graylag goose vacates their breeding grounds in eastern Turkey during the winter (Scott and Rose, 1996: 73), this species has more recently been found at the reservoir at the Göksu dam, south of Diyarbakır, there apparently being an important winter-feeding ground for this species (Karakas and Kilic, 2002: 49). With the population extent of the graylag goose seemingly having greatly changed in the recent past, the presence of this species is not considered to be a reliable indicator for seasonality.
The migratory habits of the great bustard, being the most common bird species found in both the YD and EH occupations, is poorly understood (Kirwan and Martins, 1994: 91). It is however considered to be a resident species, although until recently (1980s) it is reported that their numbers used to hugely swell in the winter in this region (Kirwan et al., 2008b: 177; Kirwan and Martins, 2000: 25). As such, its presence cannot be used as a seasonal indicator, but if in the ancient past this species also frequented southeast Anatolia as a wintering ground, this bird could have been considered an important winter resource.
The majority of bird species from the Younger Dryas levels, including the most-commonly found (great bustard and the Eurasian eagle-owl) are both known to be locally resident species, although, as mentioned, the numbers of the great bustard may be larger in the winter (Kirwan et al., 2008a: 171, 251). There are a small number of species that are known to be winter visitors including the hen harrier (Circus cyaneus) and the greater white-fronted goose (Anser albifrons) (Kirwan et al., 2008a: 132, 60). No species that are found in southeastern Anatolia exclusively in the summer were identified in the Younger Dryas levels. A number of taxa are locally resident, but their numbers will be larger in the winter when migrating populations will join them, such as the northern pintail (Anas acuta), the greylag goose, the common crane (Grus grus) and the common buzzard (Buteo buteo) (Kirwan et al., 2008a: 71, 61, 166, 139). This profile is much the same within the Early Holocene period, with a large overlap in terms of species found, with the addition of the presence of other winter visitors such as the red-breasted goose (Branta ruficollis) and the grey heron (Ardea cinerea) (Kirwan et al., 2008a: 63, 110).
The lack of presence of summer visitors is however not that surprising for southeastern Anatolia. With southeastern Anatolia being at a relatively low latitude, comparatively few bird species would be exclusively summer visitors (Newton, 2008: 370–371). Southeastern Anatolia is however an important stopover point for migrating birds, which is reflected by the higher numbers of birds and higher range of species observed in the spring today (Biricik and Karakas, 2012:104, fig.2; Karakaş, 2010, 479, fig.2, fig.3; Karakaş, 2015, 109, fig.2; Karakaş and Kiliç, 2004: 307, fig.2). Overall, in both the YD and EH levels, bird exploitation, and by extension site occupation can be inferred for the winter months. Summer occupation of the site cannot be inferred from the observed bird species. However, the lack of strong seasonality in terms of which birds would be present at different points in the annual cycle means that it is difficult to use birds as a seasonality indicator in this region. The lack of species indicating a summer occupation is however not necessarily evidence for the site being a seasonal encampment, but is more a reflection of the habits of birds in this region. Presuming year-round occupation, local bird populations may have provided a valuable food resource through winter and spring, as well as during the spring/autumn seasonal migrations, when bird populations in the region swell, and likely played a less important subsistence role in the summer, when bird numbers are lower.
Reptilia
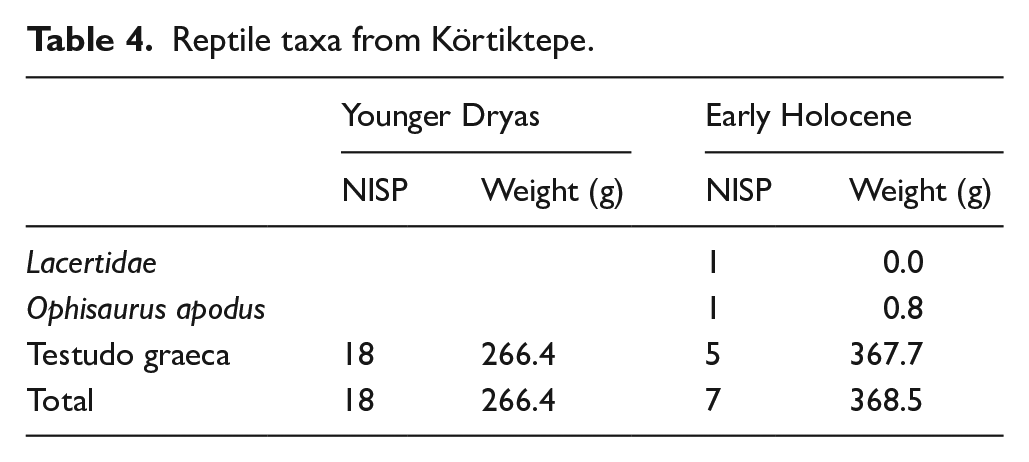
The reptile taxa from Körtiktepe are summarised in Table 4. The most common species found in both phases is the Greek Tortoise (Testudo graeca). It has been previously published that a number of tortoise carapaces have also been found within burials (Özkaya and Coşkun, 2011: 94). Additionally, from the Early Holocene contexts several skull elements from a European glass lizard (Ophisaurus apodus) and a fragment of a vertebra from the family of wall lizards (Lacertidae) were identified, both of which are potentially intrusive.
Reptile taxa from Körtiktepe.
Fish
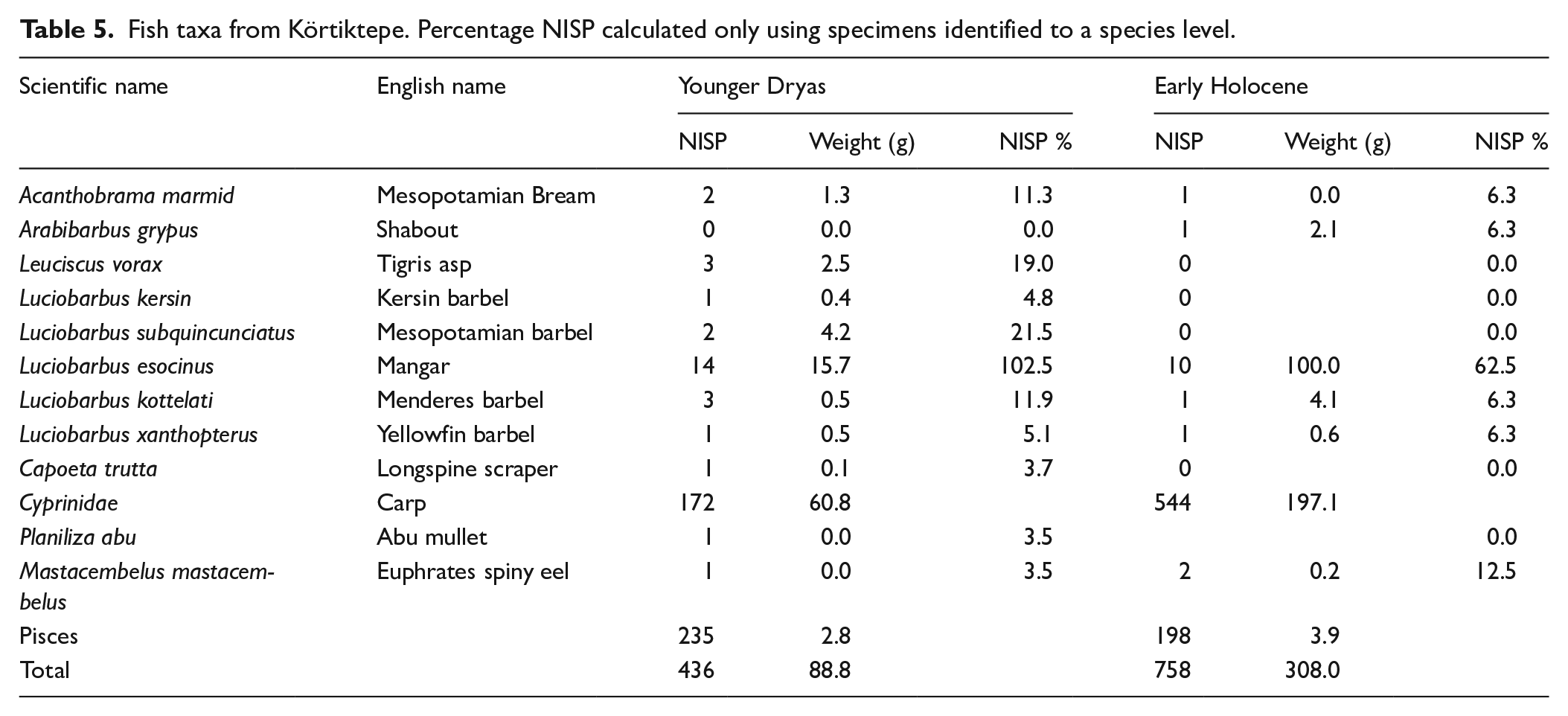
A total of 436 fish specimens from the Younger Dryas were recorded and 758 from the Early Holocene contexts. The vast majority of fish specimens could not be identified to a species level, with 53.9% and 26.1% of fish remains being recorded simply as Pisces in the Younger Dryas and Early Holocene respectively. Where species could be determined, all were freshwater taxa, unsurprising due to the location of Körtiktepe, on the bank of the modern-day Batman Çayı and the Tigris River. In both phases the vast majority of taxa are from the carp family (Cyprinidae) with the most commonly identified species being the mangar (Luciobarbus esocinus) (Table 5). Mangar can reach lengths of over 2 m and weights of in excess of 150 kg (Coad, 2021:1163-1164). A small number of particularly large vertebrae from EH Körtiktepe suggests that mangar of this size were being caught. The Mangar’s environmental requirements are not well understood but they are generally found in modern times in rivers, streams, lakes, marshes and reservoirs (Coad, 2021:1164). Apart from members of the carp family, a few examples of the spiny eel (Mastacembelus mastacembelus) were also identified. The spiny eel is a freshwater fish living in both still and running water, usually found near the bank amongst rocky crevices or plant stumps (Coad, 2015: 6).
Fish taxa from Körtiktepe. Percentage NISP calculated only using specimens identified to a species level.
Whilst too few fish specimens were identified to a species level in order to convincingly speak on differences in taxa between the Younger Dryas and Early Holocene at Körtiktepe, potentially there is some shift in the size of fish being caught between the two phases. Figure 8 shows that the height of pre-caudal vertebrae of cyprinids, as a rough indicator for the size of fish, is potentially increasing between the Younger Dryas to the Early Holocene, with certainly all of the larger specimens being recovered in the later stage of the settlement.
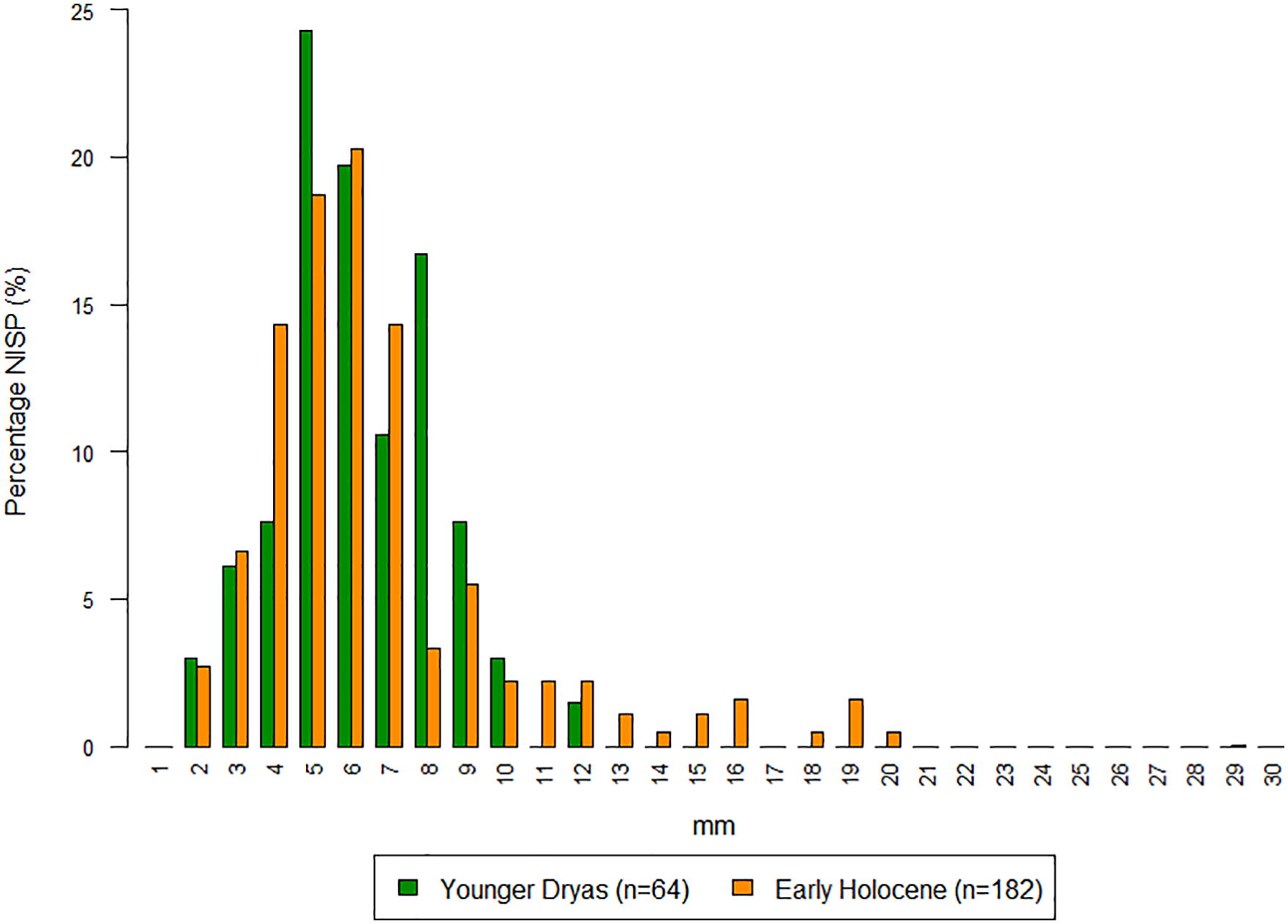
Percentage NISP (%) of the height of pre-caudal vertebrae of Cyprinids from Younger Dryas (YD) and Early Holocene (EH) of Körtiktepe.
Downstream from Körtiktepe, Hasankeyf Höyük also reports an ichthyological assemblage dominated by Cyprinidae, most commonly Barbus sp. (although these may have been reclassified into genera such as Arabibarbus sp. or Luciobarbus sp.), as well as Capoeta sp. (Itahashi et al., 2017: 42; Hongo et al., 2019: 160). Similarly, Cyprinidae was the most commonly found family at Gusir Höyük with the mangar also being the most commonly identified species.
Molluscs
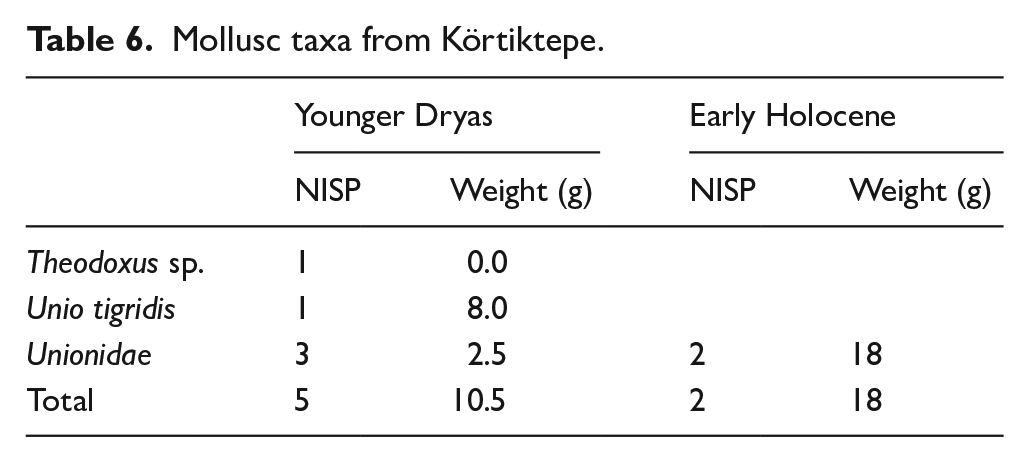
Finds of molluscs were exceptionally rare in both phases of the site. These were limited to one find of Theodoxus sp. a small freshwater snail, from the Younger Dryas, a small number of fragments belonging to freshwater mussels (Unionidae) in both phases, and single case of (Unio tigridis) being identified in a context from the Younger Dryas levels (Table 6). It should be noted, however, that finds of molluscs as grave goods in form of beads were extremely common, the full analysis of beads found is forthcoming (Özkaya and Coşkun, 2011: 101).
Mollusc taxa from Körtiktepe.
Taxonomic diversity
It has been proposed that in the Late Epipalaeolithic/Early Neolithic communities of the Near East, domestication was preceded by an increase in diet diversity, termed the Broad Spectrum Revolution (BSR) (Flannery, 1969). The BSR model has been adopted into studies of early domestication worldwide but has been intensely studied and debated in the context of the wider Mesopotamian region, particularly in regards to whether this broadening should be attributed to resource stress, or resource richness (Dobney et al., 1999; Starkovich and Stiner, 2009; Stiner, 2001; Stiner et al., 2000; Stutz et al., 2009; Wallace et al., 2019; Zeder, 2012, 2016). If the BSR were to be seen taking place at precisely this transition to the Early Holocene, heterogeneity is expected to increase.
As has been previously discussed in Peters et al. (2013), southeastern Turkey (largely sites from the Euphrates basin) sees a decrease in mammalian archaeobiodiversity (aD) over time. This increasing specialisation in terms of mammal species exploited is in particular related to the development of animal husbandry (Peters et al., 2013: 89: Fig.5.5). This pattern is, however, not observed within their ‘eastern Turkey’ assemblages (largely sites from the Upper Tigris Basin) (Peters et al., 2013: 89: Fig.5.5). The analysis for eastern Turkey however heavily relies on samples with relatively small sample sizes (<800 NISP), and a low sample size will mean that low heterogeneity is likely to be observed. It also heavily relies on the data from just a small number of sites to see what happens in the later stages of the PPN (particularly Çayönü and Cafer Höyük). Çayönü is also notably further west than the other sites compared in the Tigris valley assemblage, and Cafer Höyük is in the Euphrates basin, so may not fit into any regional pattern, if there is one to be observed. It was noted by the authors that Çayönü in particularly retains a high level of diversity throughout its occupation, which may be due to a location in a particularly lush environment, with many species readily available to exploit (Peters et al., 2013: 89). This would be in congruence with the knowledge that Çayönü is the only known site in the Tigris Valley basin to be continuously inhabited throughout the PPNB (Clare and Kinzel, 2020; Erim-Özdoğan, 2011: 62). The particular local environments and site-specific strategies tailored to those environments may then be very individualistic, rather than conforming to a wider regional trend. Körtiktepe, like Çayönü, offers an opportunity to consider these strategies over a longer period of time.
Körtiktepe sees only a small increase in NTAXA from the YD to EH from 14 to 15 mammal species, although the sample size from the EH is almost 50% higher. Nonetheless, the Shannon Entropy or aD increases from 3.0 to 4.3, showing that whilst the number of taxa utilised has remained more-or-less the same, there is a broadening on the focus of diet, with less dependence on just one or two taxa (Figure 9). The opposite can, however, be said in the case of the avifaunal remains with a decrease in NTAXA from 18 to 15 and corresponding decrease in the aD from 8.4 to 3.9, reflecting the increasing emphasis in particular on geese in the Early Holocene. The observed decrease in variance of δ13C values as well as δ15N in males found buried at the site, suggests a restricting of diet spectrum from the YD to EH, is then likely not driven by a change in the mammal spectrum, but instead perhaps from the relatively limited range of birds exploited in the EH, or by a narrowing of the plant diet (Benz et al., 2016: 241). Whilst a broadening of the mammal spectrum can be observed from the YD to EH, with the YD levels showing similarly lower levels of heterogeneity as later sites practicing animal husbandry, a uniform pattern is not observed, with the site of Gusir Höyük also displaying lower levels of aD. The restriction in bird species exploited, and indications from isotopic data also do not show conclusive evidence for a diet broadening at Körtiktepe in the PPNA. However, the slight widening of the mammalian species spectrum, with a restriction of the bird species exploited might be seen in terms of more opportunities, with more choice over which species hunted leading to a broadening of diet in some realms, but specialisation in others. This would then suggest that in our study area any observed broadening of diet was a result of climate amelioration allowing for experimentation in strategy, rather than necessity forcing the utilisation of low-ranking foodstuffs. This limited broadening of diet at Körtiktepe, largely being driven by an increase in aurochs rather than the ‘low-ranked’ species that would be expected in a BSR scenario, as well as there being little evidence of a broadening of diet in the PPNA more regionally, argues against evidence for a BSR in the Upper Mesopotamian basin, in line with previous studies’ results (Arbuckle, 2015: 227; Peters et al., 2013: 89: Fig.5.5; Starkovich and Stiner, 2009: 57–58). It should however be kept in mind that sieving of sediment was not widely undertaken at Körtiktepe, except for burials, and so smaller species are likely to have been generally underrepresented in both the YD and EH contexts.
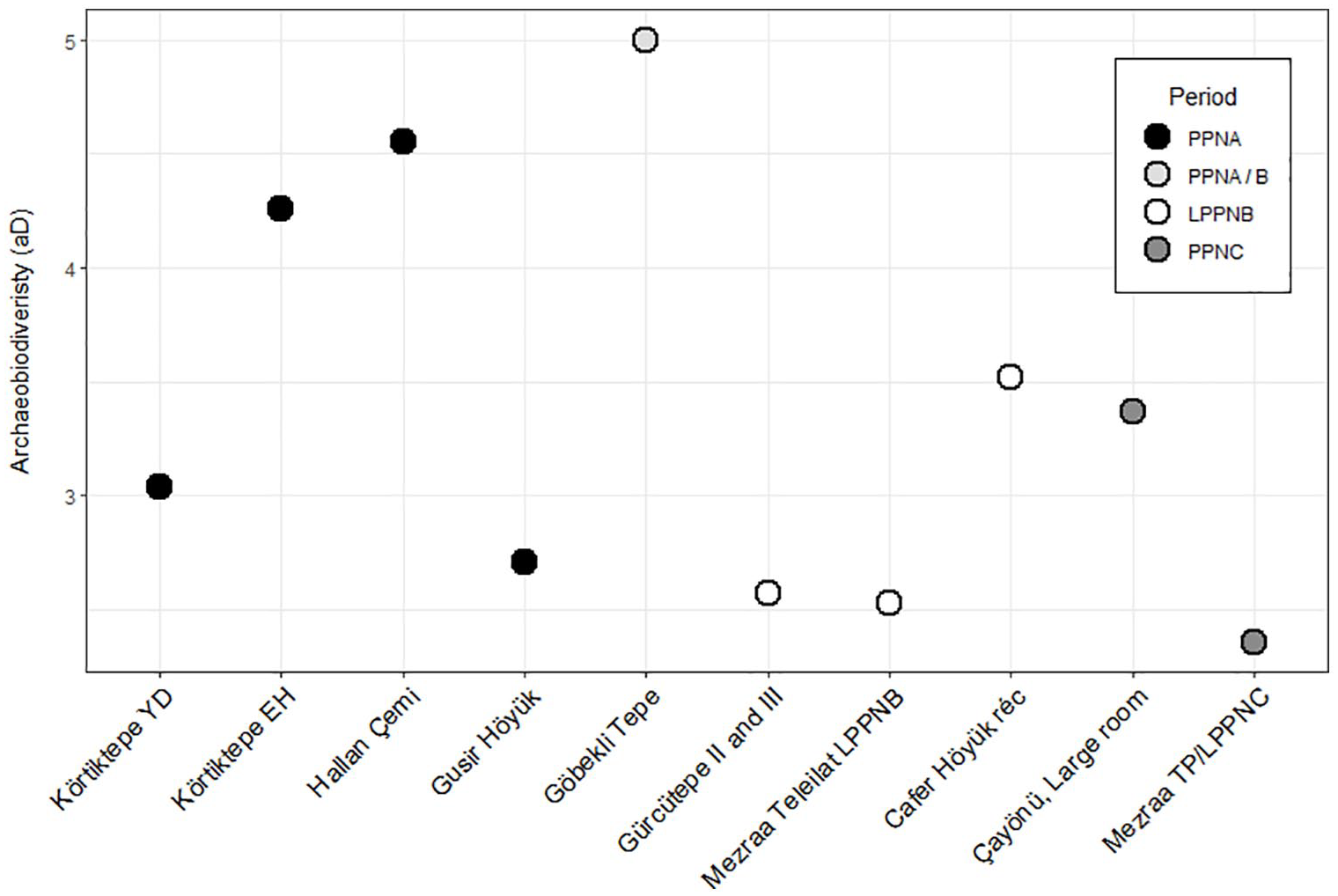
Shannon Entropy/ archaeobiodiveristy (aD) of Körtiktepe mammalian fauna and comparison with PPN sites in the Upper Tigris and Euphrates basins. Hallan Çemi data from (Starkovich and Stiner, 2009: 50, tab.3), Mezraa Teleilat data from (İlgezdi, 2008: 85, tab.11), Cafer Höyük and Çayönü after Peters et al. (2013).
Discussion
The increase in humidity, temperature, rainfall, and forest expansion that characterises the beginning of the Early Holocene in this region is clearly reflected in the relative proportion of species exploited at Körtiktepe. A major shift, particularly within the avifaunal assemblage to an emphasis in the exploitation of waterfowl in the Early Holocene, suggests changes in local waterways took place that became particularly favourable for these species. This may have included an increase in marshland with large reed beds and wet meadows or the formation of lakelets (Del Hoyo et al., 1992: 582). In the mammalian assemblage, these changes are also indicated by an increase in aurochs remains, a species that likely inhabited largely riverine, marshy, and forested environs (van Vuure, 2005: 236, 239–240, 245). This is accompanied by a moderate increase in wild boar, which whilst is an adaptable species, particularly thrives in gully side vegetation, oak forest, and mixed wood, but tends to avoid open spaces, making the local Early Holocene landscape more suitable than that of the Younger Dryas (Abaigar et al., 1994: 208; Wilson and Mittermeier, 2011: 290). The appearance, although in small numbers, of the European wildcat (Felis silvestris) is also indicative of expanding woodland in the vicinity of the settlement. Whilst riparian gallery forest may have increased generally in the EH, regionally the, if anything, slight decrease in bird species associated with woodlands, suggests that any such new woodland areas in the vicinity of Körtiktepe in the EH, still likely remained relatively sparse.
Changes in local waterways were also likely responsible for the increase in the size of fish found in the Early Holocene. Whilst it is not possible in this assemblage to know whether this increase is due to a change in the species available and exploited, or in the size of the same species, it nevertheless indicates a change in either the location or character of the local rivers or the number and size of the bodies of water. This is however caveated that this shift might be a reflection of changes in human choice or fishing technique. For instance, in one of the later layers of Körtiktepe a fishing hook with two holes in it was discovered, and it has been proposed that if the two holes were connected with string the hook would be better protected from breaking when catching large fish (Coşkun et al., 2010: 62). The importance of the role of human choice, cultural preference or changes in hunting strategy is exemplified by the avifaunal assemblage at Gusir Höyük, which despite the settlement’s location on the edge of a lake and being not far from two large rivers, is almost exclusively dominated by partridges (Alectoris chukar, Perdix perdix) whose preferred habitats are rocky hills and grasslands respectively (Kabukcu et al., 2021: 4; Kirwan et al., 2008a: 50, 53).
Small increases in frequency of a number of thermophile mammals such as the goitered gazelle, equids, and potentially also in fallow deer hints at the beginnings of a changing faunal spectrum, the result of warming at the beginning of the Early Holocene (Table 3). The increases in these species are only very slight, representing only the very beginnings of the climatic amelioration, with the faunal spectrum changing only after a delay as a period of adjustment. Overall, however, there is a great deal of continuity with species such as mouflon and great bustard remaining well represented in the assemblages. The continued presence of these species suggests continued access and exploitation of steppe-like, grassland habitats in the vicinity of the site, which is also supported by the archaeobotanical results (Rössner et al., 2018: 25).
Whilst being of relatively lesser importance, wetland environment was however exploited in the Younger Dryas, with various species of ducks, waders, and raptors such as harriers (Circus sp.) often associated with wetland habitats (Harrison, 1982: 101). Generally lacking from the Early Holocene layers, but making up a sizeable percentage (12.7%) in the Younger Dryas, however, is the Eurasian eagle-owl (Bubo bubo), a species that would primarily roost in rocky environments such as the nearby Raman mountains approximately 6 km east of the site (Harrison, 1982: 171; Kartal et al., 2018: 92). Exploitation of mountainous terrain is also perhaps indicated in both periods by the hunting of wild goat, as well as the presence of a find of Chamois (Rupicapra rupicapra) in the YD. These taxa were perhaps exploited especially in the winter when they descend from the mountains, particularly to avoid heavy snowfall (Ballo, 2010: 36; Nowak and Paradiso, 1983: 1299). The lack of Eurasian eagle-owl in the EH period may suggest either a movement away from the utilisation of this environment, or lack of interest in hunting this particular species.
Overall, whilst in both the Younger Dryas and Early Holocene multiple ecozones (wetland, grassland, mountainous habitats) were exploited, a feature that has been observed to characterise early Holocene sites in this region (Zeder and Spitzer, 2016: 144), a shift towards the exploitation of species associated with wetland environments suggests a change in the nature of the waterways in the surroundings of the site. The higher representation of wetland associated species (aurochs, boar and waterbirds) indicates that the increased rainfall of the Early Holocene has translated into an increase in marshland and/or fluvial lakes locally. This new ecological habitat, in addition to the presence of the steppes and mountainous regions that the inhabitants of the site had previously exploited, allowed for the broadening, and change in emphasis of, the mammalian taxa exploited, leading to higher heterogeneity, as indicated by the increase in archaeobiodiversity, seen in the Early Holocene. Conversely, the introduction of more geese in the local environment led to an emphasis on the hunting on these species, particularly the greylag goose, meaning a decrease in the archaeobiodiversity amongst the bird remains. From a purely optimal foraging theory perspective, geese make an attractive prey due to their relatively large size and their habit of flocking in relatively large numbers, allowing for the prospective of multiple catches in one hunt. Their size also means large feathers, which would be another attractive product to the hunters of Körtiktepe.
The increasing archaeobiodiversity amongst mammals, and restricting aD amongst the avifaunal at Körtiktepe, as well as a lack of noticeable trend over time in the Upper Tigris Basin suggests a lack of overarching trends in this region regarding climate change and heterogeneity. Instead, amongst the PPNA sites, a wide range of strategies can be seen from the relatively specialised, such as at Gusir Höyük (aD = 2.71), compared to the more diverse, such as at Hallan Çemi (aD = 4.55) or EH Körtiktepe (aD = 4.26). Instead, the particular circumstances of the site seems to be the most important factor, with the EH amelioration, in the case of Körtiktepe, allowing for the introduction of a more diverse range of mammalian taxa, and in the case of birds allowing for the specialisation on taxa that suits the inhabitants needs, either in terms of calories, convenience, use of fat or feathers, or ritual uses.
This change in local environment does not, however, translate into substantial change in mammal size ranges with both red deer and mouflon showing remarkable continuity in this respect across the Late Pleistocene/Early Holocene boundary. The few examples of smaller red deer individuals that appear in the Early Holocene is unlikely to be explained by the environment changing to one that is unsuitable for the species, as red deer are thermophiles that thrive particularly in the interface between woodland and grassland, which more closely characterised the local Early Holocene conditions rather than that of the Younger Dryas (Wilson and Mittermeier, 2011: 422). Over-hunting, particularly of large males leading to selection of smaller individuals might be considered, although the largest sizes of red deer in the Early Holocene are still of the same size as those in the Younger Dryas layers, making this scenario unlikely. It may also be explained by a change in hunting strategy or herd composition, leading to more females being hunted and a more complete representation of their body size range in the LSI data. Alternatively, this might be the beginning of some size diminution due to Bergmann’s rule, which observes that in populations members of the same species will exhibit smaller body sizes in warmer climates and conversely, larger body sizes in colder conditions (Bergmann, 1847; Meiri and Dayan, 2003; Post et al., 1997). It has also been found that Bergmann’s rule leads to size diminution of female red deer in warmer climates, but size increase in males, which might explain why the upper size range has not also reduced (Post et al., 1999). Bergmann’s rule is however controversial and the mechanisms that might lead to changes in body sizes are numerous and particular to different species (for controversy over Bergmann’s rule see: Teplitsky and Millien, 2014; Watt et al., 2010). This size diminution is additionally not replicated in mouflon, whose size range remains stable across the transition. The smaller individuals in red deer may then simply be a result of relatively low sample sizes, and further recording may show that these smaller individuals are present in the landscape in both periods. It is not possible to consider size change of aurochs across the YD-EH boundary due to low sample size. However, the wide size variation of EH aurochs could also be explained by similar reasons as the variation seen in red deer. Size diminution due to rising temperatures may be responsible for the smaller individuals, but not being far into this process, the larger individuals are also present. Alternatively, the comparatively wide size range may be due to the sex ratio. If the sex ratio is more balanced compared to the other sites, a greater size range may be observed due to the less targetted hunting strategy. However, with no sexed elements being available, it is not possible to confirm this possibility.
Continuity is observed in the mortality profiles of both red deer and mouflon, which in both species remains stable across time. This may indicate that the movements of particular sex and age cohorts of the population (bachelor herds, nursery herds as well as lone males) continued without substantial interruption across this time span, meaning hunting strategies persevered uninterrupted. This may be due to the resilience of the taxa, but also the relatively slow change in vegetation in this region, despite the rapid change in humidity and temperature at the onset of the Early Holocene (Wick et al., 2003: 670–671). The radiocarbon dates available for Körtiktepe’s early Holocene contexts begin only just after the end of the Younger Dryas and should still be considered to be within this transition phase. As such the lack of indication of change in animal movements or prey selection may then not be considered so surprising, but more dramatic changes to animal behaviour, as well as species composition, may have been observed in the assemblage had the occupation at Körtiktepe not been abandoned.
Conclusion
Körtiktepe is one of few sites within the Upper Mesopotamian basin that exhibits occupation across the Late Pleistocene/Early Holocene boundary, and thus gives a rare opportunity to consider climatic impact on the fauna at the onset of the Early Holocene amelioration and the human response to it. In both the Younger Dryas and the Early Holocene occupation levels the people of Körtiktepe primarily exploited red deer and mouflon, as is typical of this region (Peters et al., 2014: 140). The size of these species, as reconstructed through LSI data, remains relatively stable across the transition, with the exception of a small number of smaller red deer individuals being present in the Early Holocene levels. The culling profiles of both red deer and mouflon also remain similar in both periods, suggesting that the migration patterns and hunting strategies of these species remained stable, and that the environment remained hospitable for these taxa. Whilst early small-scale cultivation is suggested for Körtiktepe in the early Holocene, at present there is no indication of early animal management, although it is not possible to assess the status of wild boar and aurochs due to small sample sizes.
In the Early Holocene there was however a shift to greater exploitation of aurochs as well as a major shift towards geese within the avifauna. This indicates changes in the local environment, such as increased woodland in the case of aurochs, or changes in the waterways, perhaps the emergence of more marshland, fluvial lakes or larger or more permanent rivers. This is consistent with both the archaeobotanical evidence from the site as well as more general environmental proxies that the region received greater precipitation with the onset of the Early Holocene (Rössner et al., 2018: 24–25; Wick et al., 2003: 667–670). The results then show both a reflection of the wider environmental changes happening in the region, as well as a very local story and response.
Whilst a great deal of continuity is evidenced in both the immediate surroundings of the site, as well as in hunting strategy, the hunting of aurochs would have required a larger group of hunters in order to kill and process the carcasses of these colossal beasts (see Pöllath et al., 2018 for discussion on aurochs hunting strategies). The hunting of aurochs likely had a deeper socio-ritual meaning, with the necessity for large group co-operation combined with the danger that aurochs impose. The ritual significance of these species is well documented in the neighbouring Euphrates basin during the later PPNB period (Helmer et al., 2004: 157; Stordeur, 2010: 124), but is, for instance, also indicated for the Tigris basin by the find of an aurochs skull at a communal building at nearby PPNA Hallan Çemi, as well as finds of an aurochs horncores at Körtiktepe alongside an intramural burial ( Özkaya and Coşkun, 2011: 112, fig.6; Rosenberg, 2011: 63).
The setting of Körtiktepe allowed for the exploitation of grassland, aquatic, riverine, and conceivably mountain-dwelling species, which facilitated year-round occupation supported by a hunter-gatherer-fisher strategy. This strategy was largely maintained across a period of relatively rapid climatic change, largely without dramatic upheaval, but perhaps with the addition of small-scale cultivation, and a shifting emphasis to species that would likely have been found more frequently under the new climatic conditions. Under these conditions, the site flourished, to be then suddenly abandoned sometime in the Late PPNA for unknown reasons, although the threat of flooding will be explored as a possible reason in future studies.
Footnotes
Appendix
Sus Scrofa post-cranial fusion from Early Holocene Körtiktepe (n = 15). Age fusion stages and approximation of age of fusion from Zeder et al. (2015), p. 139, table 3.
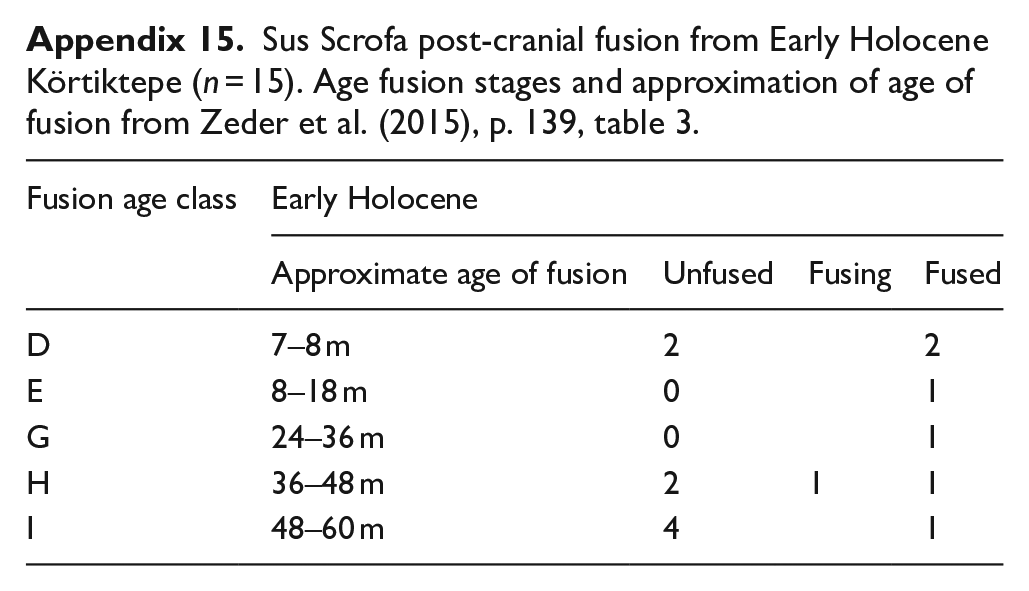
| Fusion age class | Early Holocene | |||
|---|---|---|---|---|
| Approximate age of fusion | Unfused | Fusing | Fused | |
| D | 7–8 m | 2 | 2 | |
| E | 8–18 m | 0 | 1 | |
| G | 24–36 m | 0 | 1 | |
| H | 36–48 m | 2 | 1 | 1 |
| I | 48–60 m | 4 | 1 | |
Acknowledgements
We owe thanks to the General Directorate of Cultural Heritage and Museums, Ministry of Culture and Tourism, Republic of Turkey, and our colleagues and the students of the Körtiktepe team for their hard work and valuable support. Many thanks to Jörg Schibler and Francesca Ginelli (IPNA Basel) for their support in re-sorting and handling of the assemblage. We are also thankful to Joris Peters and Nadja Pöllath for supplying unpublished data for the sites Gusir Höyük, Göbekli Tepe and Gürcütepe, as well as their valuable feedback. We are also grateful to the valuable feedback and kind comments of two anonymous reviewers.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We also gratefully acknowledge the financial support of the Deutsche Forschungsgemeinschaft (DFG) awarded to Joris Peters - Project number 165831460.