
Abstract
Stable carbon isotope analysis is increasingly used in archaeology as an indicator of crop water status and/or water management regime. While the technique shows promise, robust modern baseline studies are required to inform and validate archaeological interpretations. Here, we test stable carbon isotope values as a crop water status proxy in a monsoonal climatic context for the first time. Specifically, we test the relationship between grain stable carbon isotope values (δ13Cgrain), water availability, irrigation and soil type in barley (Hordeum vulgare L. (Zohary and Hopf.)) in north-west India, with the aim of deriving a locally-appropriate model for isotopic interpretation. We test this relationship across a substantial rainfall gradient (200–1000 mm/year) and find a negative logarithmic relationship between climatic water availability and δ13C. However, there is significant noise in the relationship, and we report δ13Cgrain variation of over 3‰ amongst samples drawn from similar climatic contexts. Soil type, irrigation type and irrigation frequency have no clear modifying effects. We conclude that: (1) barley stable carbon isotope values can act as an archaeological water status proxy in monsoonal areas, but will be most sensitive in areas receiving <450 mm rainfall per year; and (2) it is not possible to precisely infer water management regimes. On the basis of our results, we propose guidelines for archaeological barley stable carbon isotope interpretation in north-west India and analogous monsoonal climates.
Introduction
Stable carbon isotope analysis of crop remains is becoming increasingly popular as a means of reconstructing past plant water availability. In archaeology, it has gained particular interest as a crop water status proxy and/or for elucidating past water management regimes (Aguilera et al., 2011; Araus et al., 2014; Flohr et al., 2019; Riehl et al., 2008, 2014; Styring et al., 2016; Vaiglova et al., 2014; Wallace et al., 2015). Part of its popularity stems from its capacity to act as a direct physiological indicator of plant water status: meaning it does not rely on the availability of other forms of evidence (e.g. detectable irrigation infrastructure), and allows crop-specific inference of past water conditions. To date, the method has been applied in the Mediterranean, Near East, Central Europe and Britain, where it has been used to draw inferences on issues ranging from irrigation practices, crop yields, crop provenance and differential crop treatment, to climate change and climatic conditions at the emergence of agriculture (Aguilera et al., 2009, 2012; Araus et al., 2001, 2014; Bogaard et al., 2016; Ferrio et al., 2005; Fiorentino et al., 2012; Flohr et al., 2019; Lightfoot and Stevens, 2012; Masi et al., 2013; Riehl et al., 2008, 2014; Styring et al., 2016; Vaiglova et al., 2014; Wallace et al., 2015). This paper seeks to broaden the method’s applicability to monsoonal regions, by reporting a modern baseline study on barley (Hordeum vulgare) that has been grown in different soil and watering conditions in northwest India.
Principles of plant stable carbon isotopes as a water proxy
There is well-established theory underlying the use of stable carbon isotopes as a water status proxy (Cernusak et al., 2013; Farquhar et al., 1982, 1989; Tieszen, 1991). The technique is underpinned by the influence of water status on carbon isotope discrimination (Δ13C) during photosynthesis (Farquhar et al., 1982). It is particularly applicable in C3 plants (the sole focus here), which encompass over 95% of the world’s plant species (Kohn, 2010) and include major food staples such as wheat, barley and rice. In C3 plants, Δ13C is strongly influenced by plant water status because of the effect of water status on the degree of stomatal opening (Cernusak et al., 2013). In well-watered conditions, open stomata permit CO2 to diffuse freely into the leaf, allowing the photosynthetic process to discriminate against the heavier 13C isotope (O’Leary, 1988). In drier conditions, closed stomata restrict the diffusion of CO2, which reduces the isotopic discrimination. Plentiful water will thus tend to promote a high degree of carbon isotope discrimination (high Δ13C), while a water shortage will promote low carbon isotope discrimination (low Δ13C, Cernusak et al., 2013). This variation in Δ13C flows on to affect the stable carbon isotope value (δ13C) of the plant tissues that are produced from the photosynthetic sugars (Farquhar et al., 1989, see Supplemental Figure S1.1.1).
In areas where water limits plant growth – for example, arid and semi-arid zones – water status is generally the primary determinant of stomatal activity during photosynthesis, and therefore a dominant determinant of Δ¹³C (Cernusak et al., 2013). This association is reflected in a number of global and regional-scale meta-analyses, which have found clear positive relationships between moisture availability (generally as mean annual precipitation) and Δ¹³C (Diefendorf et al., 2010; Kohn, 2010). However, both Δ13C and the resulting plant tissue δ13C are also influenced by a myriad of other environmental and biological factors, including nutrient availability, light, altitude, temperature, water-logging and salinity, atmospheric CO2 concentration, as well as genotypically- and environmentally-determined physiological characteristics (Cernusak et al., 2013; Farquhar et al., 1989; Hare et al., 2018; Tieszen, 1991; see summary in Supplemental Table S1.1.1).
One factor that is particularly important for archaeological studies is the impact of the δ13Cair value (of the atmospheric CO2 source) on the final δ13C value of the plant tissue. In this paper, we focus specifically on δ13Cgrain, although similar principles apply to charcoal and other types of plant sample. δ13Cair affects δ13Cgrain because it impacts the stable carbon isotope composition of the plant sugars that are produced during photosynthesis, independent of the amount of carbon isotope discrimination. This relationship introduces complexity to archaeological interpretation because δ13Cair has varied substantially through time (Graven et al. 2017). Archaeological studies wishing to compare samples across time periods and/or with modern samples must therefore convert the measured δ13C values of the plant tissue samples to inferred Δ13C using (imperfect) estimates of past δ13Cair.
Overall, these factors reduce the precision with which: (a) variation in δ13Cgrain can be attributed to variability in Δ13C, and (b) variation in (inferred) Δ13C can be attributed to variability in water conditions – which are themselves a complex balance of soil water holding capacity, various forms of water input, and water loss through runoff, evaporation and evapotranspiration. While not insurmountable, each of these components of uncertainty must be accounted for in archaeological stable carbon isotope applications.
Support for archaeological applications
In this context, archaeological applications of carbon isotopic analysis must be underpinned by robust modern baseline studies that define the relationship between climate, water management, plant water status, Δ13C and δ13Cgrain in a way that supports a nuanced archaeological interpretation in given climatic settings. A number of such studies have now been undertaken in order to support stable carbon isotope interpretation in staple cereals (wheat and barley) and pulses (fava bean, lentil) in continental Europe, the European Mediterranean, north Africa and the Levant (Bogaard et al., 2016; Flohr et al., 2019; Styring et al., 2016; Wallace et al., 2013). These studies have resulted in several models (based on converted, i.e. inferred, Δ13C values) that have attempted either to quantify the relationship between Δ13C and precise water inputs (Araus et al., 1999; Ferrio and Voltas, 2005), or, more conservatively, establish broader relationships between Δ13C and ‘bands’ of plant water status and/or water management regime (Flohr et al., 2019; Wallace et al., 2013).
These interpretative frameworks have now been used to support a number of high impact archaeological applications (Araus et al., 2014; Styring et al., 2017), but a key gap in knowledge remains. To date, all interpretative frameworks have been based on results from baseline studies in Mediterranean climatic zones, where the dominant pattern is winter rainfall and hot dry summers. To support the broader application of the technique, research is needed to test the extent to which interpretative frameworks developed in these climatic contexts can be generalised to other ecological zones; providing locally-appropriate interpretative guidance as necessary (Flohr et al., 2019).
Here, we present the first baseline study of crop stable carbon isotope values in a monsoonal climatic zone (characterised by intense summer rainfall and, in general, low to no winter rainfall). As a case study, we examine barley (Hordeum vulgare L. (Zohary and Hopf.)) in monsoonal South Asia: specifically, the region of northwest India once occupied by the Indus Civilisation, which spread across the Indus River Basin and some of the surrounding regions (see Petrie, et al., 2017). Carbon isotope analysis has strong potential in this region, as there are steep rainfall gradients and water is scarce for some or all of the year in many areas. It is also a region where carbon isotope analysis is of strong archaeological interest: for example, there is intense debate about the extent and nature of irrigation utilised by Indus populations, and climate change-induced water scarcity has been invoked as a factor in the Indus Civilisation’s de-urbanisation (Miller, 2015; Petrie et al., 2017; Pokharia et al., 2013). In contrast to the Mediterranean contexts studied to date, much of the Indus River Basin has a monsoonal climate that brings rainfall predominantly in summer, rather than in the winter months, and often in rapid, intense bursts. This study thus seeks to: (a) provide a robust platform for carbon isotope analysis to be deployed specifically in this region’s monsoonal climatic context; and (b) serve as a broader test case to establish the degree to which frameworks developed under Mediterranean climatic regimes can be generalised to other ecological zones. We focus on H. vulgare because it is a common cereal in the archaeobotanical record across a climatically diverse portion of Eurasia (Bates, 2016; Liu et al., 2017), making it a crop for which interpretative frameworks have broad applicability.
This study assesses the relationship between stable carbon isotope values, climatic water availability, water management, soil conditions and sub-species (six-row vs two-row barley) in H. vulgare across a climatic gradient in northwest India. We take a field collection rather than a controlled experimental approach in order to best reflect the uncertainty inherent in archaeological contexts, and explicitly model irrigation, soil texture and salinity effects. This is the first time such a wide range of influences on Δ13C has been included in a modern baseline study. Due to the uncertainty introduced by δ13Cgrain to Δ13C conversions, we use δ13Cgrain in our statistical modelling. We then use our data to derive a set of principles to guide archaeological Δ13C interpretation in NW India and climatically analogous zones.
We specifically ask:
(i) Is δ13Cgrain related to climatic water availability in H. vulgare in northwest India?
(ii) Does this relationship vary with irrigation, soil texture, water logging, salinity and/or sub-species (six-row vs two-row barley)?
Material and methods
Sample collection
Samples of H. vulgare were collected in northwest India over a climatic gradient of 200–1000 mm rainfall per year (Figure 1). Our sampling transect extended over approximately 400 km, running approximately south-west to north-east. Our first sampling campaign was carried out from 14 to 20 March 2014 and over 80% of samples were collected in this season. We undertook a follow-up trip to boost sampling coverage in March 2015. March equates to harvest time for H. vulgare in this region.
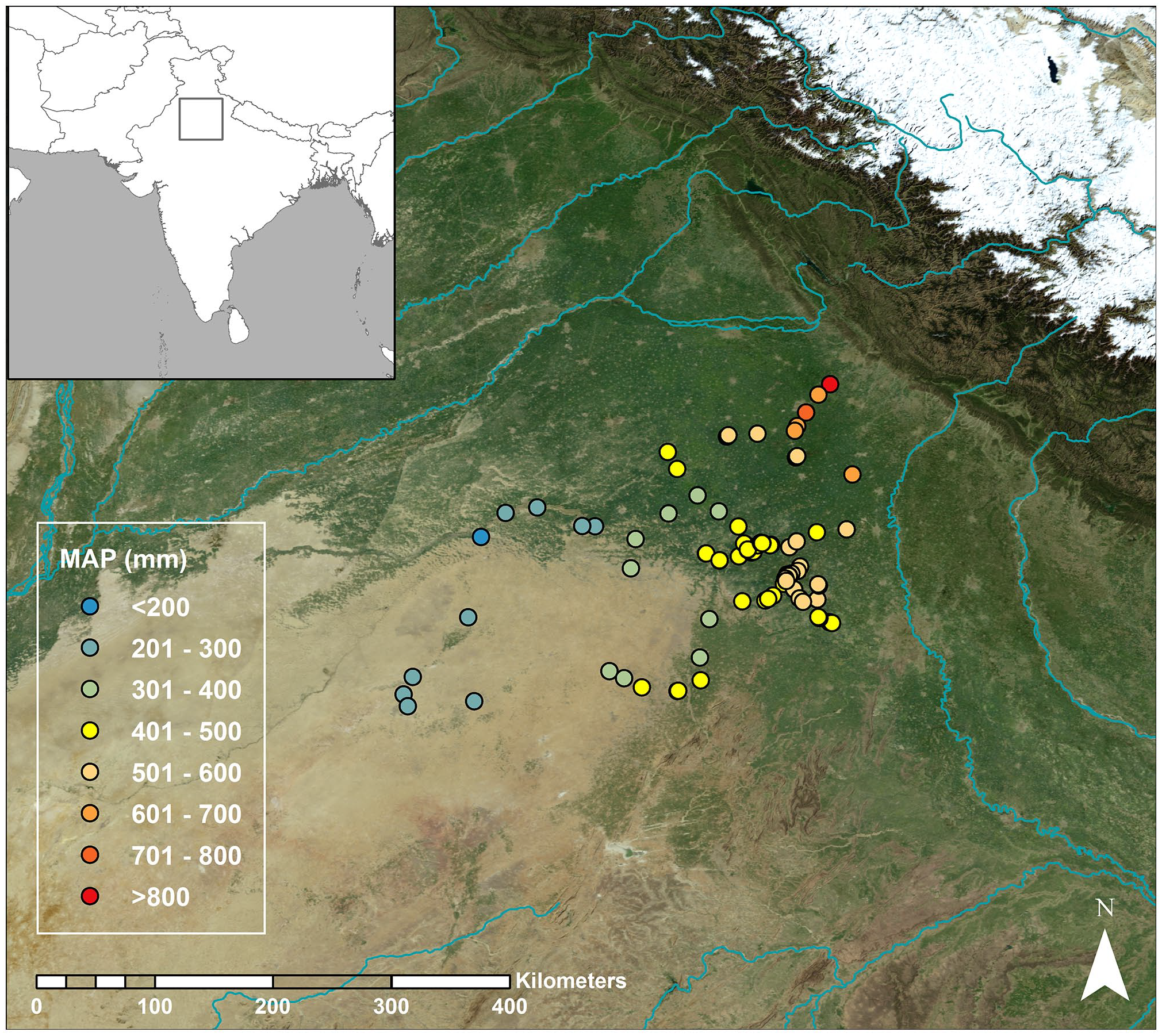
Map of north-west Indian Hordeum vulgare collection sites. Coding shows mean annual precipitation (MAP).
Sampling was based around ‘nodes,’ which were either villages/towns or significant archaeological sites. At each node, samples were collected from multiple sampling sites (that is, fields) in order to: (i) ensure sampling captured a broad representation of the local isotopic range, and (ii) enable sampling of plants grown under the same climatic conditions but under different irrigation regimes, on different soil types, or where the plants were of different phenotypes, in order to test for interactive effects.
At each sampling site, we collected samples of H. vulgare ears from: (a) standing plants within two-three weeks of harvesting, or (b) just-harvested material that was still in the field. Whole ears were taken from 15 to 20 individual plants, distributed across as much of each field as possible, but avoiding the outer ~50 cm to circumvent potential edge effects (Flohr et al., 2016).
At eight sampling sites it was evident that there was considerable intra-site heterogeneity in water availability and/or another variable of interest (soil texture, salinity or waterlogging severity). In these cases, we took separate samples from each clearly distinct ‘patch’ and recorded between-patch differences in plant, soil and water conditions. Similarly, where we could identify multiple phenotypes, we collected and analysed each phenotype separately to evaluate within-field, between-phenotype variation. In total we collected 124 samples from 87 sites across 37 nodes. Full details of all sampling locations are available in Supplemental Appendix S2.
At each sampling site, farmers were interviewed about their water management practices, plant varieties and recent weather conditions. We also photographed each sampling site and recorded details including GPS location, soil type, irrigation regime and topography. Collected samples were stored in sealed plastic bags at −20°C.
Laboratory protocol and mass spectrometry
For each sample, we stripped and pooled the grains from all ears and then randomly selected approximately 30 grains for carbon isotopic analysis. Initially, the grains were de-husked using a Qiagen tissue-lyser to maximise comparability with archaeological samples from the Indus region, the majority of which are unhulled. The grains were then ground to a fine powder using a Fisher Scientific ball-mill in order to homogenise the grains to give an average isotopic value for the growing conditions. The powder was freeze dried.
The ground samples were weighed into tin capsules for carbon isotope analysis in triplicate. Carbon isotope analysis was carried out at the Godwin Laboratory, Cambridge, using an automated Costech elemental analyser coupled in continuous flow mode to a ThermoFinnigan MAT253 mass spectrometer. Isotope values were measured as delta values (e.g. δ13C) relative to the international reference scale VPDB. Measurement uncertainty for δ13C was monitored using the international standard of caffeine (δ13C = −27.50; IAEA-600, IAEA, Vienna, Austria), and the in-house standards of nylon (δ13C: −26.6‰; size standard, nylon 6, Sigma-Aldrich, Gillingham, UK), alanine (δ13C: −26.9‰; L-alanine, Honeywell Fluka, Bucharest, Romania), protein 2 (δ13C: −27.0‰; Protein standard OAS, Elemental Microanalysis, Okehampton, UK), and again caffeine (δ13C: −35.9‰; Elemental Microanalysis). Standard materials comprised >10% of each isotopic run (Szpak et al., 2017). Using the procedures and spreadsheet provided by Szpak et al. (2017), precision (u(Rw)) was determined to be ±0.40‰ for δ13C on the basis of repeated measurements of both the international and in-house calibration standards, and on the measurements of sample replicates.
Environmental variable estimation
In order to model the relationship between δ13Cgrain, water availability and potential confounding factors, we collated the following environmental variables for each sample: climatic water availability (i.e., the climatically driven component of water availability), irrigation inputs (i.e. the anthropogenic component of water availability), soil texture, salinity, water-logging status and two-row versus six-row sub-species. We were not able to obtain quantitative estimates of irrigation inputs from farmers, we thus categorised samples by irrigation type (flood/sprinkler/rainfed) and irrigation frequency (the number of times irrigated over the course of the growing season) on the basis of farmer information. Soil texture, salinity and water-logging were graded on the basis of field observation.
In order to estimate the climatically-driven component of water availability for each sampling site (i.e. water availability un-mediated by irrigation and soil type), we used global climate and hydrology grids. As there is no consensus over which measure of climatic water availability is most appropriate in baseline studies such as this, we compiled precipitation and temperature estimates from WorldClim; the aridity index (AI) from the CGIAR-CSI Global-Aridity and Global-PET Database; and soil water content (SWC) from the CGIAR-CSI Global Soil-Water Balance Database (Trabucco and Zomer, 2010; Trabucco et al., 2008; WorldClim, 2012). All are widely-recognised global models with a spatial resolution of ~1 km2 (see Trabucco and Zomer, 2010; Trabucco et al., 2008; WorldClim, 2012 for details). Monthly estimates for each sampling site were extracted from the source databases using ArcGIS version 10.3; values are based on multi-year trends (from 1950 to 2000) and thus represent average climatic/hydrological conditions. While sampling-year specific data would have been advantageous, no 2014/2015 data were available with sufficient spatial resolution.
We then conducted a preliminary analysis to select the most appropriate proxy for use in our main analysis. On the basis of this preliminary analysis, we modelled soil water content (SWC) calculated over the growth cycle of October to March as our climatic water availability proxy. For full details and justification see Supplemental Section S1.3.
Statistical analysis
We used multiple linear regression models to test the relationship between δ13Cgrain, climatic water availability (as measured by SWC), irrigation and soil type. SWC was centred in all models and log-transformed to achieve linearity. Irrigation and soil texture have the potential to interact but had to be assessed separately because not all soil types were present in all irrigation categories and vice versa. We therefore analysed irrigation and soil texture using separate regression models but performed supplementary analyses on restricted subsets of samples to test for confounding effects. We also performed a sensitivity analysis with data from 2014 only to check whether including data from 2 years introduced additional variability or otherwise impacted our results.
The impacts of salinity, waterlogging and two-row versus six-row sub-species on the δ13Cgrain/water availability relationship were assessed via intra-site or reduced data-set analyses because the number and distribution of samples did not allow for integration into the main regressions. All analyses were performed in R (version 3.4.4) using the base, user-friendly science and car statistics packages (Fox and Weisberg, 2011; Peters, 2015; R Development Core Team, 2018).
Conversion to Δ13C
When we converted δ13Cgrain results into Δ13C to facilitate comparison with archaeological and other modern baseline studies, we used the following equation (Farquhar et al., 1982):

A key consideration in Δ13C conversion is the choice of value for δ13Cair. In addition to long term (centennial to millennial scale) variation, δ13Cair varies both seasonally and geographically. However, no seasonally resolved δ13Cair measurements from our study region or elsewhere in South Asia were obtainable. In our conversion we therefore used the 2014 and 2015 estimates of global mean δ13Cair from a compilation based on flask measurements, which are −8.42‰ and −8.44‰ respectively (Graven et al., 2017). These values differ substantively from the −8.00‰ that has been applied in a number of previous baseline studies (Flohr et al., 2011; Wallace et al., 2013); a difference that partially reflects the rapid change in δ13C values over the past two decades. While the best estimate available, the seasonal and geographical variation in δ13Cair means that the δ13C value of the atmospheric carbon that was assimilated into our sampled grains may differ from the global annual average estimate of Graven et al. (2017). Based on the degree of seasonal and spatial variation in δ13C across sites from the NOAA Earth System Research Laboratories network, we apply an estimated uncertainty of ±0.4‰ on our chosen value for δ13Cair into our Δ13C conversion to account for potential error. For full justification of our choice of δ13Cair and the ±0.4‰ uncertainty, see Supplemental Section S1.4.
Results
The relationship between δ13Cgrain and climatic water availability
Our field collections yielded H. vulgare samples from 124 ‘patches’ at 87 sites across a gradient of 194–1042 mm mean annual precipitation (MAP) (Figure 1). The range of δ13C across all samples was −21.2‰ to −28.6‰ (Table 1, full raw data in Supplemental Appendix S3).
Summary statistics for H. vulgare δ13C and converted Δ13C (‰) by irrigation type.
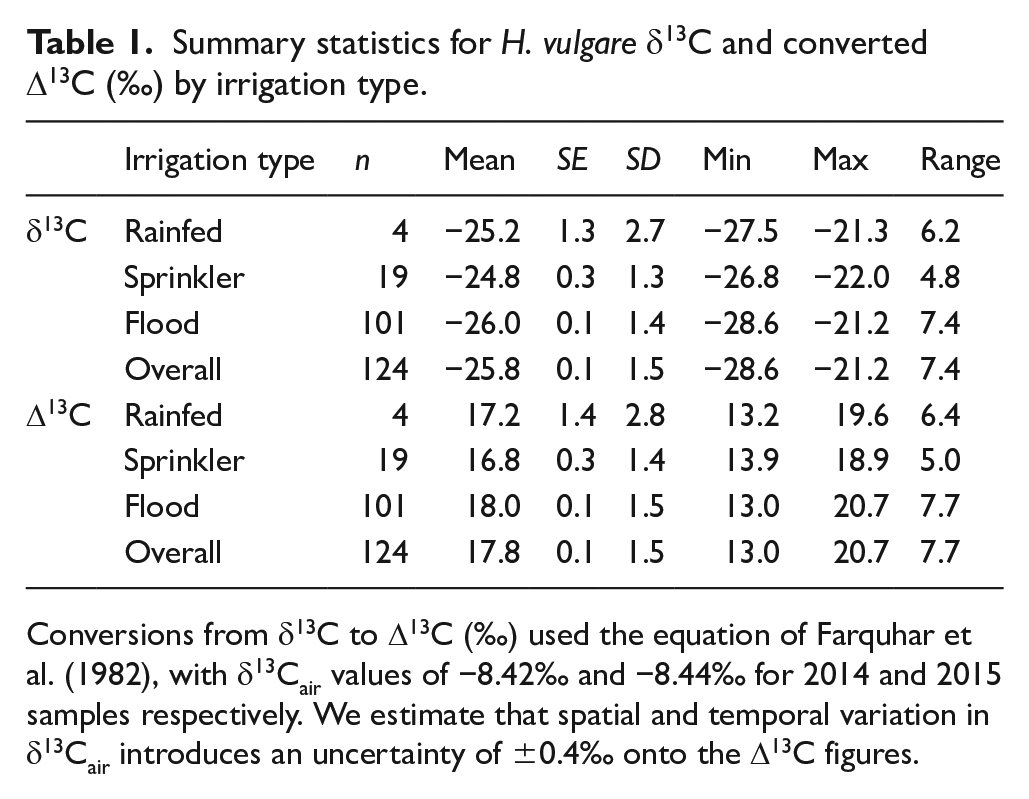
Conversions from δ13C to Δ13C (‰) used the equation of Farquhar et al. (1982), with δ13Cair values of −8.42‰ and −8.44‰ for 2014 and 2015 samples respectively. We estimate that spatial and temporal variation in δ13Cair introduces an uncertainty of ±0.4‰ onto the Δ13C figures.
Our first aim was to test the relationship between δ13Cgrain and climatic water availability (as measured by soil water content, SWC). We found a negative logarithmic association between soil water content and δ13Cgrain, though with wide scatter in the relationship (δ13Cgrain = ln(SWC), adjusted R2 = 0.18, beta = −1.77 ± 0.33SE, Figure 2, Table 2, Model H1A).
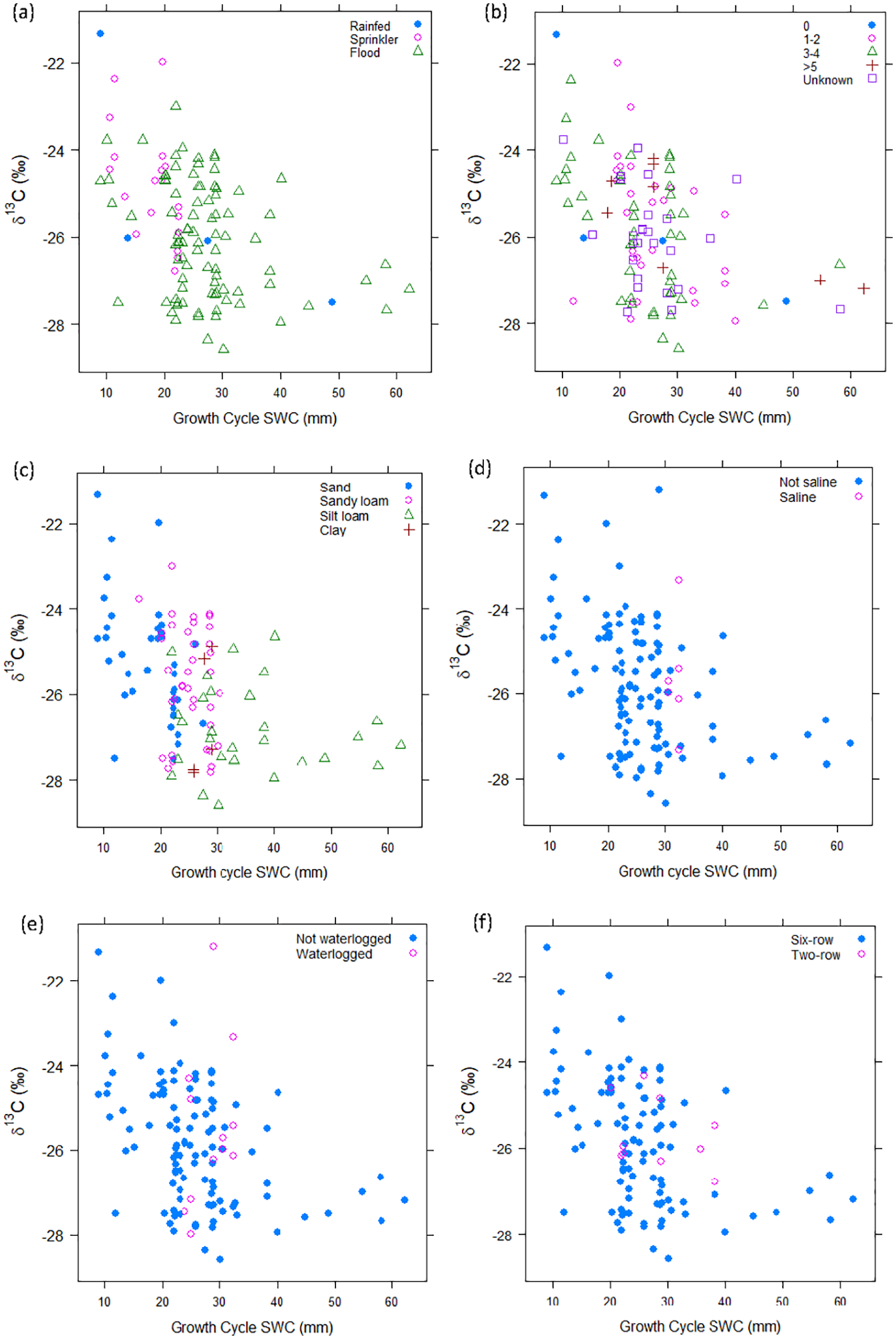
Modern NW Indian H. vulgare δ13C (‰), plotted against October to March Soil Water Content (SWC, mm) and distinguished by: (a) irrigation type, (b) irrigation frequency (the number of times irrigated over the growing season), (c) soil texture, (d) salinity status, (e) waterlogging status, and (f) six row versus two-row subspecies. Note that although the SWC data were log transformed for analysis, here we present the untransformed data in order to show the shape of the relationship and inflection point.
Results of regression models used to evaluate the effects of climatic water availability, irrigation and soil type on modern NW Indian H. vulgare δ13C.
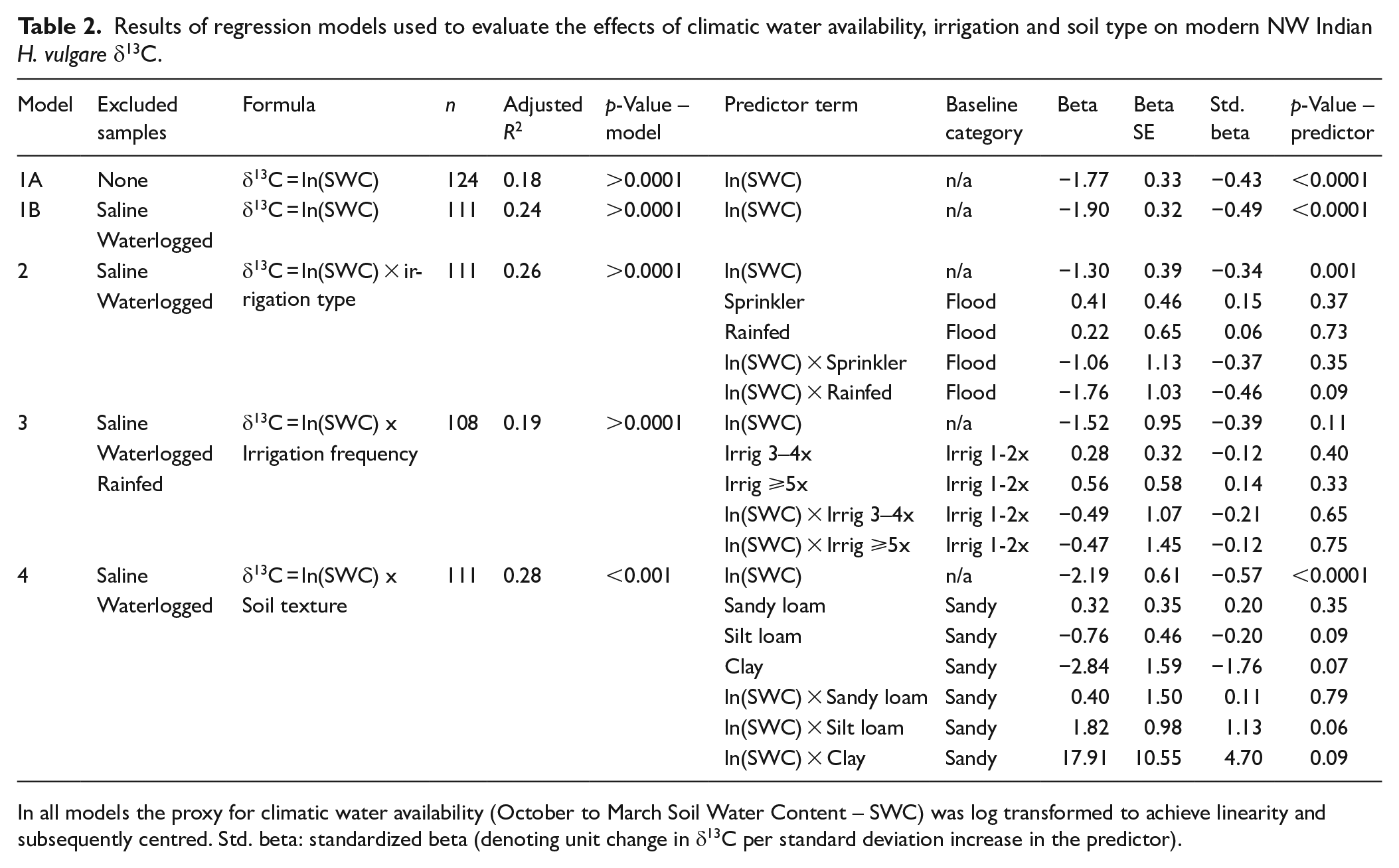
In all models the proxy for climatic water availability (October to March Soil Water Content – SWC) was log transformed to achieve linearity and subsequently centred. Std. beta: standardized beta (denoting unit change in δ13C per standard deviation increase in the predictor).
Thirteen of the 124 samples were from saline and/or waterlogged patches. As noted, salinity and waterlogging can affect Δ13C, and thus δ13C grain, independent of water availability (Cernusak et al., 2013). After removing these 13 samples, there was a tighter relationship between δ13Cgrain values and SWC (adjusted R2 = 0.24, full details in Table 2, Model H1B). On this basis, samples affected by salinity and waterlogging were removed from all further regression analyses. Their impact was investigated separately via targeted comparisons (see below).
Our sensitivity analysis with 2014 data only returned virtually identical results to the all-samples analysis; we therefore included both 2014 and 2015 data in all further analyses.
Effects of irrigation, soil texture, waterlogging, salinity and sub-species
Our second aim was to test the effect of irrigation, soil texture, waterlogging, salinity and sub-species (six-row vs two-row) on the relationship between climatic water availability and δ13Cgrain. We assessed irrigation effects in terms of: (a) irrigation type, and (b) irrigation frequency, as it was not possible to obtain quantitative irrigation input estimations.
Irrigation type
We found no marked or consistent offset between rainfed, sprinkler and flood irrigated samples within any climatic water availability band. The data show a negative logarithmic relationship between δ13Cgrain and SWC regardless of irrigation type (Figure 2a). Adding irrigation type to the regression model as an interaction term did not improve the model fit (adjusted R2 0.26 compared to 0.24 for the baseline model, ANOVA comparing δ13Cgrain = ln(SWC*IrrigType) to δ13Cgrain = ln(SWC): F = 1.71, p = 0.15). SWC remained the dominant predictor of δ13Cgrain in the irrigation type model (beta = −1.30 ± 0.39, standardised beta = −0.34). Sprinkler and rainfed irrigation had a minimal independent effect on δ13Cgrain (large standard errors and small standardized betas, for full details see Table 2, Model 2).
A supplementary analysis controlling for the effect of soil type (by using sandy-soil samples only) returned very similar results to the whole-of-dataset analysis, indicating that these results are not an artefact of soil-type effects. Full details of this analysis are in Supplemental Table S1.5.1 and Figure S1.5.5).
Irrigation frequency
As shown in Figure 2b, there was no consistent offset between more and less frequent irrigation within any given climatic water availability band. Adding irrigation frequency to the regression model as an interaction term did not improve the model fit (adjusted R2 was 0.19 for δ13Cgrain = ln(SWC)*irrigfreq compared to 0.24 for the baseline model, ANOVA comparing δ13Cgrain = ln(SWC*Irrigfreq to δ13Cgrain = ln(SWC): F = 0.72, p = 0.58). As for irrigation type, SWC remained the dominant predictor in the irrigation frequency model (beta = −1.52 ± 0.95, standardized beta = −0.39). The coefficients for all irrigation frequency groups and their interactions had large standard errors and p-values >0.3 (full details in Table 2, Model 3). A supplementary analysis controlling for the effect of soil type returned very similar results to the whole-of-dataset analysis, for full details see Supplemental Table S1.5.1 and Figure S1.5.5.
Soil properties
Soil types were distributed unevenly across the climatic water availability gradient (Supplemental Figure S1.2.1), but where they overlap there was no consistent offset between the soil texture categories (Figure 2c). The power of the regression model including soil type was limited by the uneven soil type distribution, however the results confirm that regardless of soil type, ln(SWC) remains a strong predictor of δ13Cgrain (beta = −2.19 ± 0.98, standardised beta = −0.57, for full results see Table 2, Model 4). There are possible but inconclusive indications of subtle soil texture effects: silt loam and clay both had negative coefficients – the direction that is consistent with greater water availability compared to the baseline of sand – with p-values between 0.05 and 0.10. The interactions between silt loam and clay with SWC also had p-values between 0.05 and 0.10 (details in Table 2, Model 4). Overall, adding soil type to the regression model marginally improved model fit to an adjusted R2 of 0.28, compared with 0.24 for the baseline model (ANOVA comparing this regression with the baseline model, F = 1.8, p = 0.10).
Results from a reduced data-set analysis controlling for irrigation type mirror those of this overall analysis, indicating that the results are not due to confounding. Full results of the reduced data-set analysis are in Supplemental Table S1.5.1 and Figure S1.5.6.
Waterlogging and salinity
Waterlogging and/or salinity were visibly evident at 13 sites (11 of which were waterlogged; five of which were saline). As illustrated in Figure 2d and e, neither the waterlogged nor the saline sites are consistently offset compared to others in the same climatic band. This distribution indicates that the impacts of salinity and waterlogging are not clearly discernible from other sources of variability on an inter-site basis.
The impact of these variables on δ13Cgrain was more apparent on an intra-site basis (illustrated in Supplemental Figure S1.5.3). In the fields subject to salinity, we found intra-site ranges between more and less saline patches of 0.3‰–2.2‰. The largest range was in the field with the sample with the least negative δ13Cgrain value (suggesting greatest salinity stress). With respect to the fields with more and less waterlogged patches, one displays an extreme range of >6‰, with a very high δ13Cgrain (−21.1‰) for the severely waterlogged area. In all cases the direction of difference in δ13Cgrain accords with theoretical expectations that the more stressed patch should display the highest (least negative) δ13Cgrain.
Six-row versus two-row sub-species
There is no systematic disparity between two-row and six-row barley samples across sites once climatic water availability is taken into account (although no two-row samples have values >−27.0‰, see Figure 2f).
At seven sampling sites we had the opportunity to compare δ13Cgrain directly across co-cultivated varieties. In some cases, these included both two-row and six-row varieties, in others all co-occurring varieties were six-row but were clearly phenotypically distinct. The intra-site, inter-variety range was <1.6‰ in all but one field in which there was also a difference in irrigation regime (Supplemental Figure S1.4.4). Where both two- and six- row barley co-occurred, there was no systematic trend as to which form had the higher δ13Cgrain.
Conversion to Δ13C
When converted into Δ13C for comparison with archaeological and other modern baseline studies, the Δ13C values of our samples range between 13.0‰ and 20.7‰ (Table 1, see also Figure 3). The range in converted values is slightly larger (7.7‰) than the δ13Cgrain range, due to the slight difference in δ13Cair between our two sampling years, 2014 and 2015. As noted above, we applied an uncertainty of ±0.4‰ onto our converted values due to uncertainty in δ13Cair. This is shown as error bars in Figure 3.
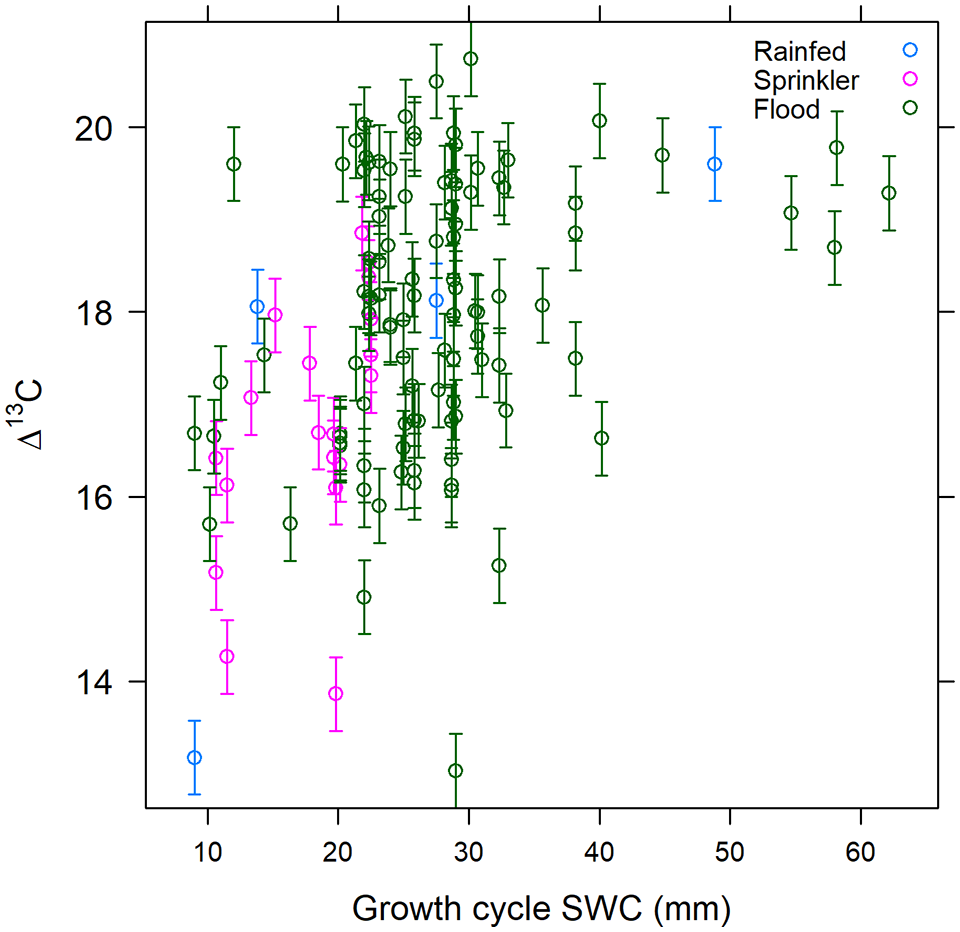
Converted modern NW Indian H. vulgare Δ13C (‰), plotted against October to March Soil Water Content (SWC, mm) and irrigation type. Δ13C conversions used the 2014 and 2015 global average estimates of δ13Cair derived from Graven et al. (2017). We estimate an uncertainty of ±0.4‰ on these δ13Cair estimates due to spatial and seasonal variation in δ13Cair, shown as error bars. Climatic water availability is measured by the growing cycle (October to March) Soil Water Content (SWC, mm).
Discussion
What is the relationship between δ13Cgrain and climatic water availability?
Our results show a clear negative logarithmic relationship between H. vulgare δ13Cgrain and soil water content (SWC), our chosen proxy for climatic water availability, across monsoonal northwest India. The logarithmic shape of the relationship indicates that climatic water availability more strongly influences δ13Cgrain in the drier parts of our study region and less so in moister areas. This pattern reflects the tendency for water to limit stomatal opening (and thus carbon isotope discrimination) more strongly in arid zones. At the upper end of the water availability spectrum, water becomes less limiting, and we see this reflected via a marked flattening of the δ13Cgrain water availability relationship in moister areas. This trend is in line with both theoretical expectations, other H. vulgare baseline studies, and global-scale, multi-taxon studies, where Δ13C appears nearly constant in humid climates (e.g. Diefendorf et al., 2010; Kohn, 2010). In this study, we found the most pronounced flattening of the carbon isotopic/climatic water availability relationship at values around 30–40 mm in October to March SWC (see Figure 2), which equates to approximately 550–600 mm mean annual precipitation.
Importantly, however, the relationship between soil water content and δ13Cgrain was characterised by significant δ13Cgrain variation amongst samples from sites experiencing similar climatic conditions. Indeed, across the whole study, the proportion of δ13Cgrain variance explained by climatic water availability was low at just R2 = 0.24 even after samples from saline and waterlogged sites were excluded. This relationship is much weaker than that found by some Mediterranean studies in which rainfall and/or water inputs were precisely known – for example, Araus et al. (1997) found a relationship of R2 = 0.75 between rainfall and H. vulgare Δ13C across a range of Mediterranean countries, while Flohr et al. (2019) found a relationship of R2 = 0.61 between H. vulgare Δ13C and water inputs in Jordan. Some of the scatter in our data thus likely reflects our use of modelled rather than measured climatic water availability values. Nevertheless, the degree of residual variation is too substantial to fully ascribe to imprecise climatic water availability estimation: indicating that numerous other factors must be impacting δ13Cgrain. The next section discusses the extent to which the residual variation can be explained by irrigation, soil texture, waterlogging, salinity and sub-species.
How do irrigation, soil texture, waterlogging, salinity and sub-species affect this relationship?
Irrigation
Contemporary irrigation regimes appear to have minimal impact on the relationship between climatic water availability and δ13Cgrain in H. vulgare grown in northwest India. There was considerable overlap between climatically-equivalent samples experiencing different irrigation regimes. Indeed in all but the very driest area, there is even overlap between irrigated and rainfed samples, although unfortunately, too few rainfed samples were available for a robust evaluation of the overall impact of irrigated versus rainfed watering regimes. Our regression analysis found that neither adding irrigation type nor irrigation frequency to the regression substantially improved model fit.
Our failure to find strong differentiation between more and less intensive irrigation regimes is consistent with findings from studies in the Mediterranean and south-west Asia (Flohr et al., 2019; Wallace et al. 2013), and most likely represents a ‘saturation’ effect, where in moister areas, even less intensive irrigation regimes may bring water availability to levels where it ceases to strongly influence stomatal opening (and hence Δ13C). This explanation is also consistent with field observations: with the exception of rainfed or minimally irrigated fields in the driest areas, all fields appeared to support healthy, well-watered barley, and almost all farmers interviewed stated that they had watered their barley to a level where water did not present a limit to growth. This ‘saturation effect’ is a key issue to consider in archaeological interpretation and is discussed further below.
Soil texture
While a study with greater power is needed to confirm conclusions, our data suggest that soil texture may have a minor mediating effect on the relationship between climatic water availability and H. vulgare δ13Cgrain in northwest India. For example, we found indications of a possible tendency for silt loam and clay soils to be associated with slightly more negative δ13Cgrain than sandy soils, given similar moisture regimes. This pattern is in line with theoretical expectations: coarse (sandy) soils retain water poorly, whereas silt loam soils are generally considered good providers of plant-available moisture because they balance water retention with a reasonable degree of plant water availability (Saxton and Rawls, 2006; Sperry and Hacke, 2002). Overall, however, the δ13Cgrain/climatic water availability trajectory is clearly dominant over any soil effects, and within any narrow climatic range, we found just as much variation amongst samples from the same soil types as amongst those across multiple soil types. On this basis, in an archaeological context, soil texture appears unlikely to add significant noise to the relationship between climatic water availability and δ13Cgrain.
Salinity and waterlogging
We found no clear offset between waterlogged and saline samples and those from unaffected sites, although two samples – one waterlogged and one that was both waterlogged and saline – appear as outliers for their climatic range with values >−23.5‰. These are both samples from particularly severely waterlogged and/or saline patches, which returned much higher δ13Cgrain values than less severely affected patches from the same site. Our data thus suggest that severe, but only severe, salinity and waterlogging significantly affect the relationship between water availability and H. vulgare δ13Cgrain.
Six-row versus two-row sub-species
We found no isotopic difference between two-row and six-row H. vulgare grown under similar conditions. This contrasts with experimental field studies in the Mediterranean and Canada which have found higher Δ13C (generally by 0.5‰–1.0‰, equating to more negative δ13Cgrain) in six-row compared to two-row forms under a range of water regimes (e.g. Anyia et al., 2007; Jiang et al., 2006; Voltas et al., 1999). This previously reported distinction has been attributed to a tendency for six-row forms to translocate more carbohydrates from the flag leaf and stem to the grain during the grain filling period: the rationale is that this translocated carbon would have been fixed at an earlier date, when water availability is assumed to have been greater (Brugnoli and Farquhar, 2000). It is not clear why a two-row/six-row offset is not evident in this study, but here, even amongst varieties from the same field, two-row varieties did not always return higher δ13Cgrain values (the direction expected if two-row varieties discriminate less). We note that although we found no difference in the overall δ13Cgrain/climatic water availability relationship, it is possible that the mediating effects of irrigation, soil texture, salinity and water-logging vary between the sub-species. We did not have the statistical power to test this here; but we note this as a worthy topic for further study.
What are the implications for archaeological application in monsoonal NW India?
These data provide a detailed empirical platform upon which to develop principles for archaeological applications of stable carbon isotope analysis in monsoonal NW India and potentially in other monsoonal areas. Overall, our results support the dominant influence of water availability on H. vulgare stable isotope values: the climatic water availability gradient clearly emerges as the dominant driver of δ13Cgrain, with soil type, salinity and waterlogging appearing to have little to negligible confounding effect. This pattern supports the capacity of stable carbon isotopes to act as a broad indicator of past moisture availability trends.
However, our data also present some important caveats and cautions. First and foremost, our results highlight that archaeological applications must account for a significant degree of noise in the relationship between water availability and stable carbon isotope values. We found differences frequently up to 2‰, in some cases 5‰–6‰, amongst samples from sites with similar SWC values (i.e. across sites experiencing similar amounts of precipitation and evaporation). We also found considerable overlap in samples experiencing different irrigation regimes, at both the lower and upper ends of our climatic water availability gradient. On this basis, we propose that it is neither possible to quantify climatic parameters, nor to determine irrigation regime on the basis of δ13Cgrain – or the inferred Δ13C values necessary for archaeological applications, which have additional uncertainty attached. Our data also show that much of monsoonal NW India – as a broad approximation, areas receiving ~>550 mm/year – is moist enough to render δ/Δ13C insensitive as an indicator for changes or differences in water availability: the flattening of the logarithmic relationship between δ13Cgrain and climatic water availability at ~550 mm/year implies that beyond this point water no longer presents a substantial enough limitation to photosynthesis to strongly impact Δ13C.
Nevertheless, within these constraints, there are patterns in our data that are consistent enough to support some very coarse-grained inferences, particularly from more extreme values of Δ13C. On this basis, we propose some conservative principles for archaeological Δ13C interpretation in our study region and climatically analogous monsoonal areas. These principles could be used to infer water availability (i.e. environmental conditions), water status (i.e. plant physiological status) or both, depending on the study purpose. Although our study only explicitly considered the relationship between Δ13C and water availability, both inferences are valid given that water availability impacts Δ13C via its effect on plant water status (and thus degree to which stomata are closed). Specifically, we propose that: +++++
Δ13C > ~19‰–20‰ can be interpreted as likely to indicate very well-watered conditions (or very well-watered barley), where water no longer presents a limit to the ratio of inter-cellular to atmospheric CO2 concentrations. This guideline is proposed on the basis that all sites receiving over 700 mm/year display values >19‰; and with one exception, all of the 35 sites with values >19‰ were either flood irrigated or were in very high rainfall areas (>1000 mm/year). This threshold is also consistent with the findings of studies from other regions (see Supplemental Table S1.6.1), where mean values of >19‰ are only, and consistently, reported for fully-irrigated barley in medium-high rainfall zones.
Δ13C < ~15‰–16‰ can be interpreted as likely to indicate poorly-watered conditions and/or severe salinity or waterlogging stress. In this study, other than highly saline/waterlogged patches, with one exception, only samples from rainfed or close-to-rainfed contexts (poorly-watered field margins) in areas receiving <450 mm rainfall returned values in this range. Again, this pattern is consistent with findings from other regions, where mean Δ13C < 16.0‰ is only reported for rainfed barley from low rainfall areas (Supplemental Table S1.6.1).
Interpreting values between these extremes is more challenging, and we note that Flohr et al. (2019) also concluded that interpreting mid-range Δ13C values is difficult. However, on the basis of our study, the range ~16‰–17‰ could be characterised as likely representative of ‘less well to moderately-watered’ barley, on the basis that >90% of samples with values of 16‰–17‰ came from sites at the lower end of the climatic water availability gradient (receiving <510 mm/year). It should be noted, however, that all samples in this range were irrigated (including some flood irrigated >5 times), and that values in this range therefore do not necessarily indicate a lack of irrigation or conditions of severe water constraint.
The range ~17‰–19‰ could be characterised as representing ‘at least moderately, and probably well-watered’ barley. Samples within this range were distributed across a wide combination of climatic and irrigation regimes, but overwhelmingly represent barley from irrigated fields in moderate to high rainfall areas, where farmer interviews and/or visual assessment suggest optimum or, at least near optimum water status. In modern northwest India, this isotopic range therefore appears to represent generally well-watered barley that experiences minimal or no water constraint. Finer distinctions within this range are not possible given the degree of noise from other factors.
Given (a) the degree of scatter in our data and (b) the uncertainty introduced by δ13Cgrain to Δ13C conversions, we suggest these ranges as diffuse brackets along a gradient of probabilities of water status – rather than as definitive thresholds above or below which barley should be considered well or poorly watered. In this context, the precise cut-points of each band should not be given undue emphasis. The degree of scatter in our data also emphasises that inferences made on the basis of single grain values or small sample sizes will not be robust. With these caveats, however, these principles (displayed in Figure 4) provide a basis for making useful inferences from archaeobotanical barley Δ13C in monsoonal contexts, without exaggerating the precision with which it is possible to infer past moisture availability, water inputs or irrigation regimes.
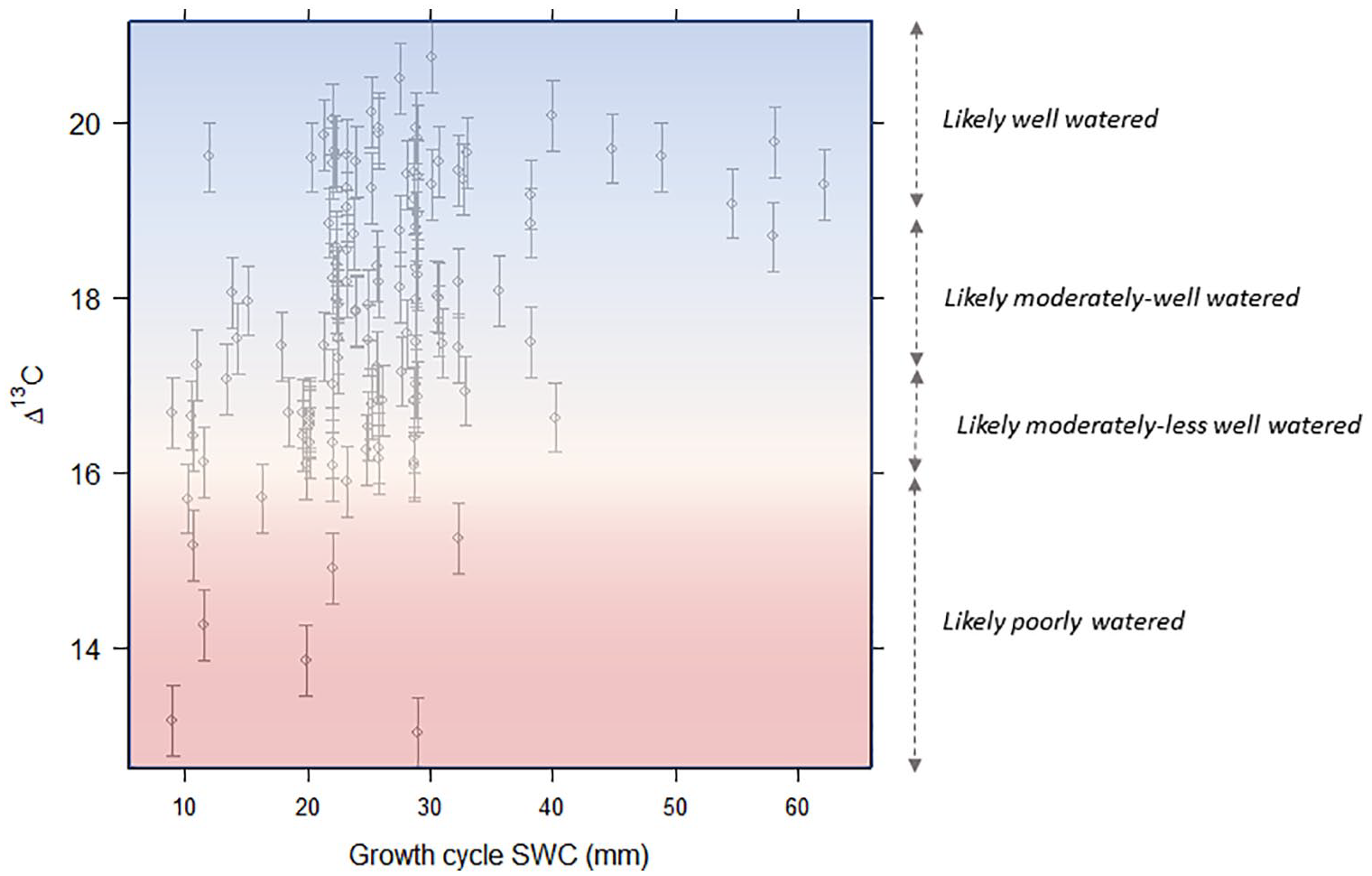
Proposed framework for the interpretation of archaeobotanical barley Δ13C in the greater Indus region, overlaid on a plot of modern barley Δ13C against climatic water availability and irrigation status. Climatic water availability is measured by the growing cycle (October to March) Soil Water Content (SWC, mm). The dotted lines and colour gradient emphasise that the values for demarcations between coarse bands of water status should not be treated as thresholds but simply useful marker points along a diffuse gradient of probabilities that a crop has been more or less well watered.
What are the broader implications for archaeological stable carbon isotope interpretation?
A key driver of this study was the need for empirical evidence to support archaeological applications of barley stable carbon isotope analysis interpretation in the greater Indus River Basin and analogous monsoonal climatic contexts. In addition to fulfilling this goal, our findings have important broader implications for archaeological Δ13C applications.
First, our findings support the broad transferability of water availability/Δ13C frameworks across a range of climatic regimes. Our proposed principles are significantly more conservative than, but not inconsistent with, frameworks previously proposed for Mediterranean regions. For example, on the basis of data from the Mediterranean and Levant, Wallace et al. (2013) proposed that barley Δ13C values >18.5‰ can be interpreted as indicative of well-watered barley, 17.5‰–18.5‰ as moderately watered, and <17.5‰ as poorly watered plants. We note that Wallace et al.’s (2013) values are approximately 0.4‰ offset (upwards) compared to ours, due to differences in the values used for modern δ13Cair - Wallace et al. (2013) used −8.00‰, whereas we used −8.42‰ and −8.44‰ as we had access to the more up-to-date δ13Cair estimates from Graven et al. (2017). Even after taking this difference into account, the thresholds proposed by Wallace et al. (2013) are broadly consistent with, even if much less conservative, than those proposed here. This consistency is not unexpected, but nevertheless represents an important explicit test of the transferability of principles across climatic contexts.
Second, this study adds to growing evidence that the degree of uncertainty inherent in crop Δ13C interpretation is much greater than commonly assumed. Our findings support the argument previously made by Wallace et al. (2013) and Flohr et al. (2011, 2019) that quantitative models relating Δ13C to water inputs are inappropriate, and demonstrate that in archaeological contexts, small differences in Δ13C either between sites or across periods cannot be confidently interpreted in terms of differences in moisture availability. In the context of barley, Wallace et al. (2013) suggested that any differences <0.5‰ should be disregarded. Even if some of the scatter in our data reflects our use of estimated rather than measured climatic water availability values, the sheer degree of scatter in the data presented here strongly suggest that an even more conservative approach is required. We further highlight the need to account for uncertainty introduced by δ13Cgrain to Δ13C conversions (which we suggest are at least ±0.4‰), in addition to possible impacts from changes in atmospheric CO2 concentration (Mora-González et al., 2019).
Third, our data provide further evidence that plant carbon isotope analysis has limited capacity to distinguish between intensities of irrigation in the archaeological record. We found considerable overlap in Δ13C in samples experiencing very different types and frequencies of irrigation. This observation adds to the findings of Flohr et al. (2019), who found overlapping Δ13C in barley receiving irrigation ranging from 40% to 120% of optimal water inputs in Jordan. Across a range of Mediterranean contexts, Wallace et al (2013) found that irrigation regimes recognised as distinct by farmers could not always be differentiated using Δ13C. Overall, this finding highlights that Δ13C cannot determine the detail of past water management regimes. An exception may be broad rainfed versus irrigated distinctions in arid zones – for example, Styring et al., 2016 found a marked difference between rainfed and oasis-irrigated barley in an arid (MAP <200 mm) area of Morocco, and in Jordan, Flohr et al. (2011) found a significant difference between rainfed and irrigated samples at sites receiving MAP <300 mm. Our data suggest that the ability to distinguish between rainfed/poorly-watered versus optimally-irrigated samples is likely to be strongest in areas receiving <400 mm rainfall/year: in this climatic band, rainfed or close-to-rainfed barley (low frequency sprinkler irrigation) has values typically <14.5‰, whereas all moderately to intensively irrigated barley has values over >16‰. In areas receiving > 400–450 mm/year, distinctions between un-irrigated, less intensively irrigated and more intensively irrigated barley appear difficult. This limitation has important implications for the ability of Δ13C analysis to contribute to debates about the existence and nature of irrigation in many archaeological contexts, including the Indus Civilisation.
Fourth, soil texture and mild to moderate salinity and water-logging appear unlikely to significantly confound the relationship between climatic water availability and Δ13C in an archaeological context. This observation is an important finding given that these soil qualities are often unknown and therefore difficult to control for analytically. Severe salinity and water-logging do result in very low Δ13C that could be mistakenly interpreted as a result of water shortage. However, other markers such as weed ecology and geoarchaeological proxies may assist in assessing the likelihood of any such confounding.
Finally, our study raises significant questions about the offset of 1‰ between two-row and six-row barley that has been applied in archaeological contexts (e.g. Styring et al., 2016; Wallace et al., 2015). We found no systematic difference between these two types and the ranges we propose above for broad categories of water status appear to apply equally to two- and six-row barley in NW India. Further investigation of whether and when any offset may be applicable is therefore required to support robust archaeological interpretation.
Conclusions
This study of modern barley stable carbon isotope values provides an empirically-supported platform for making inferences – however cautious – about past water status in NW India and climatically analogous monsoonal areas. Despite the high degree of noise, these data show that δ13Cgrain is sensitive to the general climatic water balance, and they support water availability as the dominant single driver of δ13Cgrain across the region. They suggest that inferred Δ13C values have capacity to identify both highly water-stressed and very well-watered conditions with a reasonable degree of confidence, even if inferences regarding irrigation regime, or moderate water status gradations appear difficult. Finally, within and beyond our study region, we highlight the significant uncertainty inherent in Δ13C interpretation, and recommend that isotopic interpretations are supported wherever possible by complementary evidence from weed ecology, geoarchaeological and palaeoecological data.
Supplemental Material
sj-pdf-1-hol-10.1177_0959683621994649 – Supplemental material for Crop water status from plant stable carbon isotope values: A test case for monsoonal climates
Supplemental material, sj-pdf-1-hol-10.1177_0959683621994649 for Crop water status from plant stable carbon isotope values: A test case for monsoonal climates by Penelope J Jones, Tamsin C O’Connell, Martin K Jones, RN Singh and Cameron A Petrie in The Holocene
Supplemental Material
sj-xlsx-1-hol-10.1177_0959683621994649 – Supplemental material for Crop water status from plant stable carbon isotope values: A test case for monsoonal climates
Supplemental material, sj-xlsx-1-hol-10.1177_0959683621994649 for Crop water status from plant stable carbon isotope values: A test case for monsoonal climates by Penelope J Jones, Tamsin C O’Connell, Martin K Jones, RN Singh and Cameron A Petrie in The Holocene
Supplemental Material
sj-xlsx-2-hol-10.1177_0959683621994649 – Supplemental material for Crop water status from plant stable carbon isotope values: A test case for monsoonal climates
Supplemental material, sj-xlsx-2-hol-10.1177_0959683621994649 for Crop water status from plant stable carbon isotope values: A test case for monsoonal climates by Penelope J Jones, Tamsin C O’Connell, Martin K Jones, RN Singh and Cameron A Petrie in The Holocene
Footnotes
Acknowledgements
The authors thank the barley producers of Haryana and Rajasthan for permission to access their fields and for providing cultivation histories. Appu Singh (MD University Rohtak) and Amit Ranjan (Banaras Hindu University) made this study possible as translators and field guides. We would also like to thank the Department of AIHC and Archaeology at Banaras Hindu University for their logistical support. At the University of Cambridge, we are grateful to Catherine Kneale and James Rolfe for their help with isotopic analysis, Chris Rolfe for allowing access to facilities and Louise Butterworth and Samantha Smith for technical support. We would like to thank Howard Griffiths, Nick Owen, Emma Lightfoot and Jennifer Bates for useful discussions and advice. AK Pandey provided valuable support in the field and we thank David Redhouse and Ian Hitchman for IT support. No aspect of the broader archaeological study that this work helped inform would have been possible without excavation and export permission from the Archaeological Survey of India.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This paper developed out of research conducted for the Land, Water and Settlement and TwoRains projects, which are both collaborations between Banaras Hindu University and the University of Cambridge investigating human–environment relations in northwest India. The Land, Water and Settlement project was primarily funded by a Standard Award from the UK India Education and Research Initiative (UKIERI) under the title ‘From the collapse of Harappan urbanism to the rise of the great Early Historic cities: Investigating the cultural and geographical transformation of northwest India between 2000 and 300 BC’. The TwoRains project is funded by the European Research Council under the European Union’s Horizon 2020 research and innovation program [grant number 648609]. This specific study received additional funding from the Geographical Club, INTACH-UK and the European Research Council project Food Globalization in Prehistory [grant number GA249642]. PJ Jones’ overall doctoral study was funded by the Rae and Edith Bennett Travelling Scholarship.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.