
Abstract
A unique assemblage consisting of 113 pine samples collected from a submerged Mesolithic landscape in the Haväng area, southern Sweden, was examined to assess the presence of large herbivores, as well as changes in wild-game population density and composition. Bark-stripping damages on prehistoric trees is an extremely underutilized source of information about past game-population dynamics, yet our analyzes of wood samples – dated to around 10 500 cal. yr. BP – shows that such material can be successfully used to study the presence and activities of large herbivores, most likely ungulates. To evaluate our results, comparisons have been made with subfossil peatland trees that grew around 6000 years ago, as well as trees from two present day clearcut logging sites in southern Sweden. Furthermore, studies in a wild-game reserve were performed to recognize and understand different types of damages on trees caused by ungulates. Bark-stripping indicate the presence of ungulates, and the rate of damage is commonly associated with the density of the wild game. Bark-stripping was most frequently observed in the submerged wood material from the early Holocene, where damages were detected in 15% of the trees. In comparisons, 11% of the mid-Holocene trees show bark-stripping damages, whereas marks could be detected in the range between 0% and 6% of the trees from the two present-day clearcut logging sites. Our results show that tree-ring analyzes of prehistoric wood can generate information about wild-game dynamics of the past, and thereby being a valuable complement to more commonly used paleoecological and zooarcheological records.
Keywords
Introduction
In the southern Baltic Sea basin there are numerous submerged coastal landscapes containing organic archeological findings, vegetation remains, and rooted tree stumps from the early and mid-Holocene (Bailey et al., 2020; Rosentau et al., 2017). The relatively low salinity in the Baltic Sea means that the shipworm (Teredo navalis) is uncommon, which allows for exceptional preservation conditions for wooden remains (Björdal et al., 2012). Such findings can be extremely valuable for paleoecological and archeological studies. Numerous studies of submerged landscapes have been performed in Danish and German coastal waters (Fischer, 2007; Hartz et al., 2014), but so far surveys have not been carried out to the same extent along the Swedish coastline (Figure 1). In southernmost Sweden, submerged wooden remains have been documented since the early 20th century (Isberg, 1927), but, except from the Haväng site, in depth archeological and geological research has been lacking (Nilsson et al., 2018). The Haväng site is situated at the outlet of the Verkeå River on the eastern coast of the Skåne region. During the early Holocene, lower water levels in the region allowed for a pine forest to extend beyond the present coastline, which was subsequently submerged during the rapid shore level rise called the Ancylus transgression about 10 500 years ago (Hansson et al., 2018a). The Haväng site, which has been known since the 1960s (Nilsson, 1961), is one of the most important submerged landscapes in Sweden and has been the target of various paleoenvironmental research projects since the 1980s (Gaillard and Lemdahl, 1994; Hansen, 1995). Since 2009, surveys have intensified, with high-resolution bathymetric studies, surveillance, sample diving, and sediment coring being carried out. The many diving campaigns have resulted in an ever-increasing body of evidence, consisting of wood samples, faunal remains, and sediment samples from the submerged landscape (Hansson, 2018; Hansson et al., 2018a, 2019; Nilsson et al., 2018).
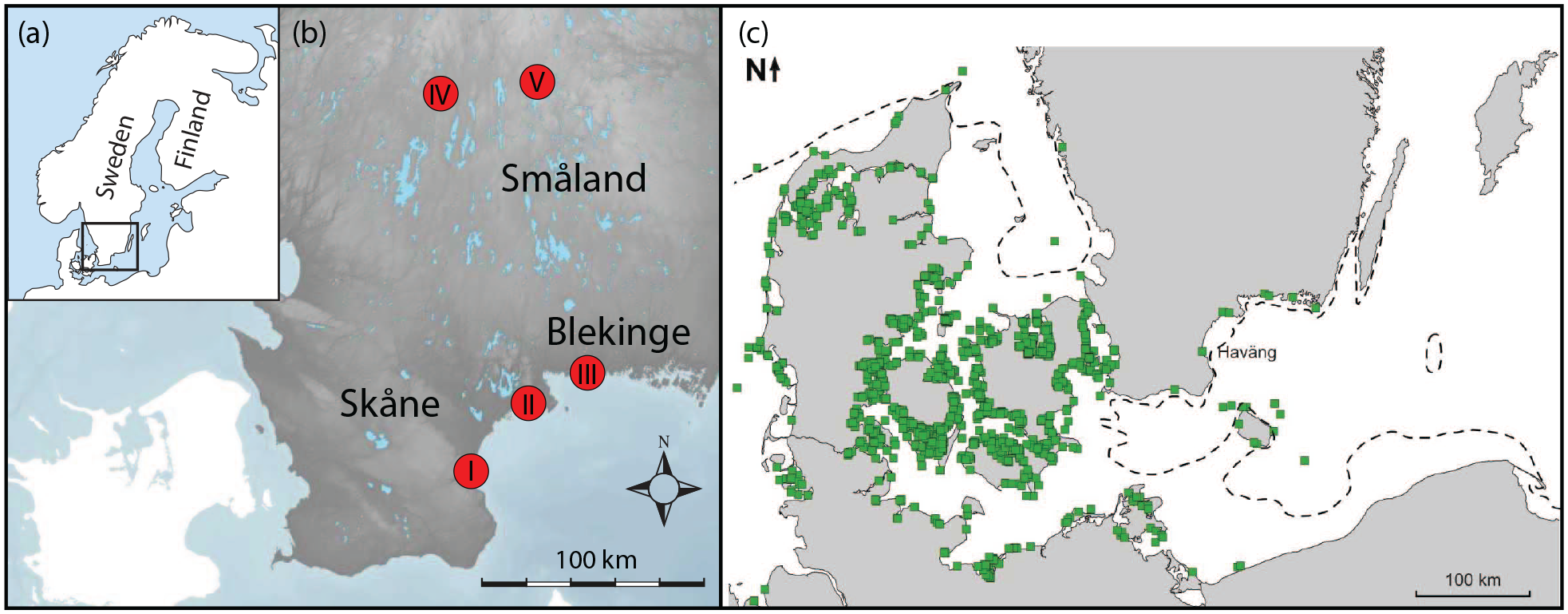
(a) Overview of the location of the study area. (b) Location of the study sites: (I) Haväng area, (II) Ljungaviken, (III) Eriksberg, (IV) Store Mosse National Park, and (V) Hällarydsmossen (topographical data ©Lantmäteriet). (c) The green squares show submerged study sites along the south Swedish and Danish coasts (SPLASHCOS-Network, 2016), whereas the dashed line shows the lowest coastline since the last deglaciation.
Radiocarbon dating and dendrochronological analyzes of trees from the submerged landscape in the Haväng area have been used previously to study landscape dynamics and shore-level displacement during the early Holocene (Gaillard and Lemdahl, 1994; Hansson, 2018; Hansson et al., 2018a, 2019; Nilsson et al., 2018). Besides detailed reconstructions of local coastline and environmental changes, the wood samples revealed damages probably caused by large herbivores, foremost ungulates, while the trees were still alive.
Marks and damages on trees caused by large ungulates is a natural phenomenon that can reveal information about the presence of different wild-game species, their distribution, population density, and variations in their behavior (Gill, 1992; Ueckermann, 1956; Verheyden et al., 2006). There is a plethora of reasons why different wild-game species cause such damages on trees. Common causes can be to access nutrients from the trees, to darken the horns with resin, or to counteract itching. The damages can be divided broadly into three categories: browsing, bark-stripping, and fraying damages (Gill, 1992). Browsing is usually defined as damages caused by the removal of twigs, shoots, leaves, needles, buds, and flowers, leaving no marks on the stem of the trees. Bark-stripping on the other hand, is when animals, such as deer (Cervidae), wild horse (Equus ferus), and brown bear (Ursus arctos) peel off the bark external to the cambium on the trees with their teeth (Kuiters et al., 2006; McIntyre, 1972; Sommer et al., 2011; Zyśk-Gorczyńska et al., 2016). Fraying damages occur due to the removal of bark from trees by the antlers of deer, the horns of bovids (Bovidae), or the tusks of wild boar (Sus scrofa) and another example of damages related to mating behaviors, marking of territories, or velvet removal of antlers of deer (Gill, 1992). Territorial markings by claws of large predators, such as and brown bear and Eurasian lynx (Lynx lynx), also cause damages on trees (Černe et al., 2019).
Dendrochronology is a well-established research discipline in which growth patterns of trees are used to study and date climate dynamics, environmental changes, and wooden artifacts, among other things (Cook and Kairiukstis, 1990; Fritts, 1976). Tree-ring data from construction timber and subfossil trees has, for example, been used in studies of cultural heritage (Haneca et al., 2009), archeology (Baillie, 2014; Kuniholm, 2002), paleoecology (Edvardsson et al., 2014; Lindbladh et al., 2013), and paleoclimatology (Edvardsson et al., 2016; Hughes, 2002). Dendrochronology has also been used to study submerged wooden material to reconstruct coastline shifts, land uplift, and vegetation changes during the early Holocene (Hansson et al., 2018a; Kaiser et al., 2018). Moreover, scars and damages on trees can be used to date and study natural hazards (Stoffel and Bollschweiler, 2008) and fire frequencies in natural forests (Drobyshev et al., 2015; Harley et al., 2018). However, even though wood samples do often show traces of animal behaviors (Gill, 1992; Verheyden et al., 2006), dendrochronological studies have been used to document wild-game population dynamics to a much lesser extent. For subfossil trees, the number is lesser still, with only a small handful of studies of insect fauna (Boswijk and Whitehouse, 2002), findings of wood gnawed by beaver (Castor fiber), and marks after woodpecker (Picidae) (Björkman, 1993). Furthermore, despite their potential, we note a complete absence of dendrochronological studies using prehistoric wood for the documentation of large ungulates such as red deer (Cervus elaphus), European bison (Bison bonasus), and Euroasian elk (Alces alces).
Reconstructions of wild-game density and dynamics can be done on three levels: (i) presence of the wild game, (ii) relative estimates of the wild-game population, and (iii) absolute estimates of the wild-game population. Most paleoecological and zooarcheological studies of past wild-game populations are based on analyzes of faunal remains to reconstruct the chronological and geographic distribution of different taxa. Additionally, quantification of faunal remains from a site or period is used to estimate the relative frequencies of different wild-game taxa (Grayson, 1979; Magnell, 2017; Tsahar et al., 2009). However, there are as yet no available zooarcheological or other methods to quantify the actual abundance and population density of wild game in the past. Damages on trees caused by large ungulates may have the potential to be used as a direct indicator of local presence of the animals, as well as a possible proxy to assess wild-game population density.
In this study, wood samples have been analyzed with the aim to (i) evaluate the possibilities to confirm the presence of large ungulates at a site (without preserved or recovered faunal remains) through studies of damage to the trees, to (ii) study early Holocene wild-game population dynamics in a submerged landscape in the Haväng area, southern Baltic basin, based on damage on trees, and (iii) to test which of the three levels (presence, relative estimates, or absolute estimates) of the wild-game population that can be achieved from studies of the submerged Haväng trees. To evaluate and put our results into context, we will also compare the wood material from the Haväng area with bark-stripping and fraying damage on trees from other sites and periods during the Holocene. Comparison with contemporary material may give us the opportunity to estimate the size and composition of the wild-game population in the Haväng area during the Early Mesolithic.
Methods and material
Site description and regional setting
This study is mainly based on subfossil trees from the submerged landscape in the Haväng area (Figure 1). For comparative studies, we analyzed subfossil peatland trees from the Hällarydsmossen peatland, as well as recently felled trees from a clearcut logging site adjacent to the Store Mosse National Park and another clearcut logging in the Ljungaviken area. To study and identify different types of wild-game damages on trees, we have also documented marks on trees made by ungulates in the Eriksberg wild-game reserve recently.
The Haväng area is situated on the coast of the Hanö Bay in southeastern Sweden (Figure 1). The bay is part of the Baltic Sea and constitutes the eastern coastline of the Skåne region and the southern coastline of the Blekinge region. The area is characterized by sandy sediments, which are cut through by a small meandering river called Verkeån, which has its outlet at the coastline at Haväng. The sandy sediments were deposited in local glacial lakes situated between the receding ice sheet to the northeast and the higher topography toward the west during the last deglaciation (Malmberg Persson, 2000). The sandy sediments overlying the diamicton seafloor extend about 10 km east from the present coastline (Björck et al., 1990).
Beyond the present coastline, remains of the relict Verkeån River extend about 2.5 km toward the east, down to a depth of around 25 m b.s.l. (Hansson, 2018). The size of this submerged river valley is around 500 m × 2500 m. In the Haväng area, episodes of relatively low water levels in the early Holocene have been recorded from the Yoldia Sea stage until the initial Littorina Sea stage (c. 11 700–8500 cal. yr. BP). During parts of this episode, the coastline was located several kilometers to the east of the present coastline, allowing forests to establish on the exposed land areas. Today fragments of this landscape can be found on the seafloor in the shape of organic deposits from and surrounding the ancient river channel, as well as 100s of rooted pine stumps and remains of fallen trees (Hansson, 2018). Several archeological findings have also been recovered from the organic sediments on the seafloor, including beaver bones with cut marks from butchering and worked bones and antlers from aurochs (Bos primigenius), red deer and Eurasian elk. Moreover, pine torches and stationary fishing constructions have been discovered in the organic sediments (Hansson et al., 2018a, 2018b).
Hällarydsmossen is a peatland in the central part of the region of Småland (Figures 1 and 2). The site is located 215 m a.s.l., and about 200 km north of the Hanö Bay area. Due to extensive peat extraction, an estimated 4 m of peat has been removed and as a result, the remains from 1000s of subfossil Mesolithic pine trees have been exposed (Edvardsson et al., 2012). There are no sites with faunal remains in the Hällarydsmossen area but based on the presence of wild-game fauna from Mesolithic sites in the region of Småland and nearby regions of south Sweden, red deer is expected to have been the most common wild game during the Middle Mesolithic, followed by wild boar, roe deer (Capreolus capreolus), and Eurasian elk (Gummesson et al., 2019; Magnell, 2017).

Photos from the five sites from which trees have been studied. (a) Tree sampling at the submerged landscape in the Haväng area. (b) Sampling of subfossil trees at the peatland Hällarydsmossen. (c) The clearcut logging site west of the Store Mosse National Park. (d) The timber from the clearcut logging site in Ljungaviken. (e) The Eriksberg wild-game reserve. Photos by A. Sjöström (a) and J. Edvardsson (b−e).
The Store Mosse National Park (Figures 1 and 2) is also located in central Småland and contains the largest raised bog complex in southern Sweden (Svensson, 1988). The studied trees, however, grew on mineral soil in a managed forest area adjacent to the National park and west of the peat bog. The trees were felled during the autumn of 2019. Eurasian elk is the main ungulate in the area expected to cause bark-stripping, but roe deer and to a lesser extent red deer and fallow deer (Dama dama) are also present.
Ljungaviken is an area defined by a small bay just outside the town of Sölvesborg, located in southwestern part of the Blekinge region (Figures 1 and 2). Since a residential area is being built along the eastern coast of the bay, trees have been felled in areas that were previously occupied by a mixed forest of spruce and pine. The trees were felled and studied during the spring of 2020. The wild-game population in the area is limited to roe deer and wild boar.
In order to study and document different kinds of marks and damages to trees caused by large ungulates, the Eriksberg wild-game reserve was visited (Figures 1 and 2). Eriksberg is a fenced reserve with forests and grasslands covering 915 ha situated by the archipelago in the Blekinge region. The reserve was established in 1938 and today (2020) continues to manage a free-roaming wild-game population of 55 European bison, 250–300 red deer, 700–800 fallow deer, 30 Père David’s deer (Elaphurus davidianus), 300–350 wild boar, and 150–200 mouflon (Ovis gmelini).
Material
From the submerged landscape in the Haväng area, a total of 127 wood samples were collected for dendrochronological analysis, shoreline displacement studies and paleoecological reconstructions (Gaillard and Lemdahl, 1994; Hansen, 1995; Hansson et al., 2018a, 2018b, 2019). The majority of this assemblage (i.e. 113 samples), are Scots pine (Pinus sylvestris L.). The remaining taxa are willow (Salix spp, 12 samples) and birch (Betula spp, 2 samples). In total, 28 of the pine samples have been radiocarbon (14C) dated (Gaillard and Lemdahl, 1994; Hansson et al., 2018a). Moreover, two pine tree-ring chronologies have been developed from cross-dated tree-ring series (Table 1; Hansson, 2018).
Information about the studied trees and the tree-ring chronologies.
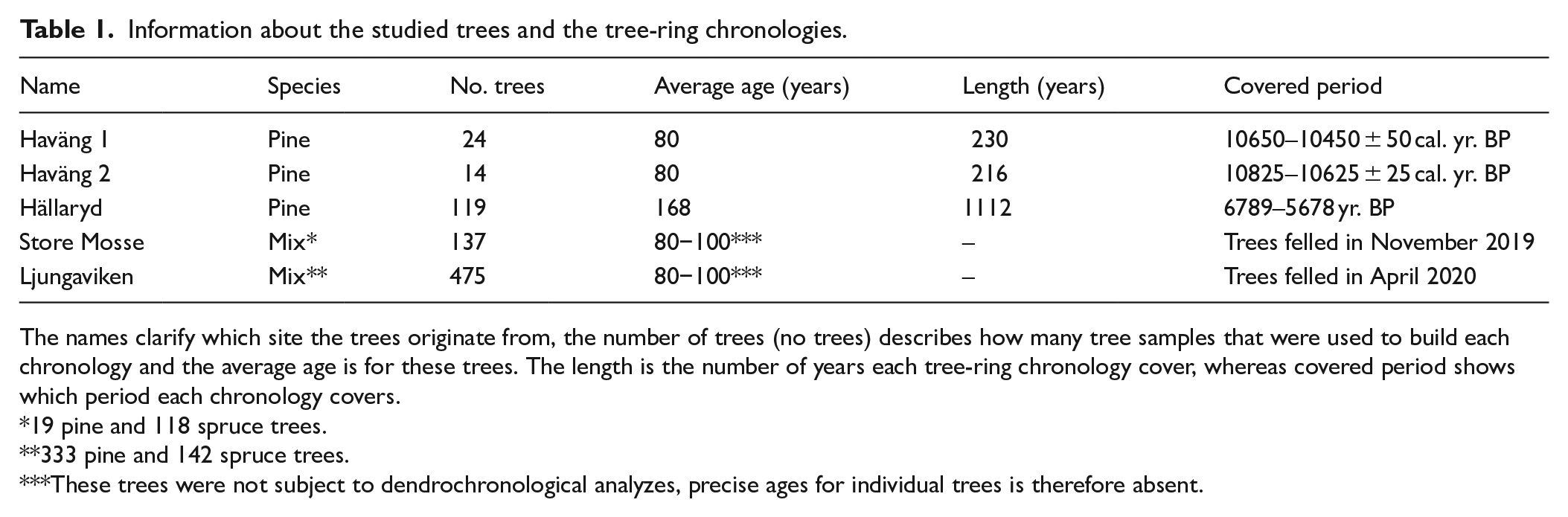
The names clarify which site the trees originate from, the number of trees (no trees) describes how many tree samples that were used to build each chronology and the average age is for these trees. The length is the number of years each tree-ring chronology cover, whereas covered period shows which period each chronology covers.
19 pine and 118 spruce trees.
333 pine and 142 spruce trees.
These trees were not subject to dendrochronological analyzes, precise ages for individual trees is therefore absent.
For comparison, an assemblage of 139 subfossil peatland trees from the Hällarydsmossen peatland (Edvardsson et al., 2012) was used. This assemblage is comprised exclusively of Scots pine; from 119 of the samples, a 1112-year long tree-ring chronology (cross-dated to 4839–3728 BCE using northwest German pine chronologies) has been developed (Edvardsson et al., 2012). To homogenize the time scales and facilitate comparisons to the radiocarbon dated records (and because exact calendrical years are of less relevance in this study), we will hereafter write 6789–5678 BP before present (yr. BP, years prior to 1950) when referring to the age of the Hällarydsmossen trees.
To enable a comparison with known, present-day conditions, stumps, and stems of conifers were studied at two clearcut logging sites, one at Ljungaviken close to the town of Sölvesborg and the other adjacent to the Store Mosse National Park. At the Ljungaviken site, visible surfaces, and tree trunks from 475 felled trees could be studied, whereas 137 visible stump surfaces and tree trunks could be recorded at the clearcut logging in the Store Mosse National Park. The trees that were studied had all been felled shortly before the visits to the sites in 2019 and 2020, respectively, which meant that cross-sections on the stumps and trunks were sufficiently clear, and that external effects and damages to the trees were detected in the field (Figure 2).
Methods
The wood material from the Haväng area has been analyzed using standard dendrochronological approaches, and 28 samples were selected for radiocarbon dating (Hansson et al., 2018a). For the study of wild-game damages using the submerged wood remains, only the pine trees were considered. The reasons for this are mainly because (i) most of the well-preserved wood samples from the Haväng area consist of pine (88% of the trees), (ii) the main part of the material available for comparative studies also consists of pine trees, and (iii) pine is still present in the region today, allowing modern analogies to be made.
The material has undergone a detailed visual examination, during which different types of damages and growth anomalies have been identified, counted, and measured. In this study we define damages as disturbances in the growth pattern of the tree-rings caused by external factors which, based on their morphology, have been divided into four categories: wild-game damage, fire scars, reaction wood, and undefined marks. Wild-game damage to trees can be further divided into three categories: browsing, bark-stripping, and fraying (Gill, 1992). Unlike browsing, bark-stripping and fraying are wild-game activities that include the removal of the bark from the trees, and it is these two types of damage that we expect to detect on the tree trunks and stumps studied. Other type of damage expected in the trees is fire scars. Fire scars can be separated from damage caused by wild game since fires leave (i) charred bark and wood, (ii) mainly scars at the base of tree, and (iii) triangular shape scares, often with an open and charred surface (Johnson and Gutsell, 1994). Furthermore, trees of small size or young age can usually not withstand large wildfires. Damage that has occurred during the trees’ first years can therefore often be attributed to causes other than wildfires. Furthermore, we can observe reaction wood, that is, when the stem of a tree recovers from being bent or tilted and reerects itself to grow vertically again (Scurfield, 1973). The reaction wood appears as an asymmetrical growth pattern, and can be considered a growth anomaly rather than actual damage. Damage can also occur if trees fall against each other during storms, or as a result of natural hazards (Stoffel and Bollschweiler, 2008). Where possible, the height of the damage above the root level was noted. On discs where damages were detected, the extent (width) of the damage was measured.
The subfossil pine trees from the Hällarydsmossen peatland have undergone the same procedure as the trees from the Haväng site to allow for comparative studies. As for the comparative material from the two clear cuts, the material was visually inspected, and photographed in the field. For each tree, the number of visual damages were inspected, counted, and interpreted. Furthermore, recent damages to trees in the Eriksberg reserve was studied. These damages could be studied and documented both on living and felled trees, and in parallel could be linked to various ungulates.
Results
After the visual examination of the 113 pine samples from the submerged landscape of Haväng, possible marks caused by large ungulates were recorded on 17 samples (15.0%), whereas reaction wood (i.e. indicating that the tree had started to tilt) was recorded on 14 samples (12.4%). Non-climatic disturbances were recorded in 27.4% of the samples (Table 2). As it is not possible to determine what has caused the growth anomalies on the samples (i.e. they only show reaction wood and no actual scars), we avoided classifying these features as wild-game damages in the result section, rather paying attention to these growth changes in the discussion.
Information about the compiled pine populations including sites the trees originate from, the number of trees the statistics are based on, and what percentage of the trees as different types of damage have been detected in.

With regard to the total number of damages, we have included damage interpreted to be caused by other sources than the wild game and fires (e.g. humans and hazards) are included.
Visible in unknown number of trees.
Marks interpreted as wild-game damage on the trunks were found from root level up to about 2 m above the ground. A large spread was also noted on the size of the damages on the studied material, ranging from few to 10s of centimeters in size (Figures 3 and 4). Furthermore, a large spatial spread among the damaged trees was noted: most of the damaged trees were found at depths of around 6 m b.s.l, but damaged trees occur throughout the area, from a few 100 m to 2.5 km out from the present shoreline.

Present day damages on trees. (a) Damages to the bark and tree trunk underneath from European bison horns. (b) A tree that has been used, over many years, by the wild game to rub their horns on. (c) Tree that show both marks from horns and that animals have rubbed their bodies against the trunk. The horns make scratches whereas the rubbing has resulted in large spots where the bark has disappeared completely. (d) Trees which, at the age of about 7, 11, and 17 years, have been damaged by deer, but thereafter allowed to re-grow relatively undisturbed. (e) Several generations of wild-game damage around the entire stem. Photos by J. Edvardsson.

(a) Submerged pine tree with two instances of wild-game damage from when the tree was about 5 and 15 years old, similar to what was observed in Figure 3d. (b) In situ sample from Tärnö, Blekinge (not included in this study), showing several instances of wild-game damage. (c) Subfossil pine tree with fire scar. (d) Submerged pine tree with several instances of wild-game damage around the stem. (e) Close-up of damage from the previous image. Note the similarities between the damage on this c. 10 500-year-old pine tree and the modern-day trees in Figure 3. (f) Example of reaction wood. The tree has initially had an upright position (inside the green circle), but thereafter started to tilt and produced reaction wood so as not to fall. Photos by J. Edvardsson (a, c−f), and A. Sjöström (b).
Damages cause by wild game were recorded less frequently in the material used for comparative studies. Damage interpreted as caused by large ungulates were noted in 10.8% of the trees from the Hällarydsmossen peatland, whereas fire scars were detected in 7.2% of the samples. Even less damage was recovered on the recently felled trees (Table 2). In total, 8.8% of the studied trees from the Store Mosse National Park showed damage, of which 5.5% were interpreted as being caused by ungulates and 1.5% by fires. The least damage, 3.2% in total, was observed on the trees from the Ljungaviken clearcut logging. Reaction wood was observed not only on tree samples from the Haväng area, but the documentation of this particular parameter was not accurate enough for a precise percentage to be calculated for the other sites. However, reaction wood was noted to occur frequently on the trees from Hällarydsmossen.
Discussion
Interpretation of the results
Bark-stripping and fraying damages
Based on the morphology, size, and placement of the marks and damages on the trees, we interpret most of the damage to the submerged wood from the Haväng area to be caused by large ungulates, resulting from bark-stripping. Bark-stripping is a common feeding behavior among large ungulates, and damages on tree stems are frequently found in areas that are rich in wild-game (Gill, 1992; Verheyden et al., 2006). Bark-stripping is also expected to have been a common phenomenon in the early and mid-Holocene forests in northern Europe, something we can see traces of in the Haväng material. However, fraying and territorial markings by antlers, for example, of red deer before the rut, are also a common feature on trees in areas with a large game population (Gill, 1992). By way of example, we observed that markings caused by the horns of European bison occurred frequently on the trees in the Eriksberg reserve (Figure 3a). Since it is likely that large bovids, such as aurochs present in the area during the early Holocene as shown by bone finds, had similar behaviors, some of the damage on the Haväng material are also assumed to have been caused by fraying. We can therefore expect such damage in the Haväng material, but it is difficult to distinguish fraying from bark-stripping with certainty. Regarding other causes of damage, fire scars could be detected, whereas damage often discussed in the literature, such as rock falls, landslides, and avalanches (Stoffel and Bollschweiler, 2008) can be excluded because the landscape around the surveyed sites is flat.
We interpret that the proportion of trees showing ungulate damage, associated foremost with bark-stripping, to be positively associated with the density of the wild-game populations at different sites and periods (Tables 2 and 3). A study of the bark-stripping frequencies on conifers by Verheyden et al. (2006) shows that the damage frequency is significantly larger in areas with high deer density than areas with medium or low deer density. This basic assumption has been confirmed in several studies (Gill, 1992; Kiffner et al., 2008; Ueckermann, 1956; Welch et al., 1991), but there are also examples of studies that do not show a significant relationship between deer density and bark stripping frequency (Volk, 1999; Welch et al., 1987). Moreover, the number of bark-stripping events is not evenly distributed between all the trees in a forest stand and is often clustered depending on the size and age of the trees, size of the forest block, food availability, tree taxa, ground vegetation, and topography (Gill, 1992; Verheyden et al., 2006; Zidar, 2011). Therefore, comparison between different regions often shows large spread even in contemporary contexts where knowledge of the wild-game population is relatively good (Table 3). It is therefore extremely difficult to make a quantitative estimate of the density of a wild-game population based on marks on trees alone, especially if we focus on periods far back in time. But nevertheless, it is possible to make comparative studies using wood from different sites and periods.
Recent quantifications of frequency of bark-stripping rate on conifers in 33 areas in Europe in relation to density of red deer and elk.
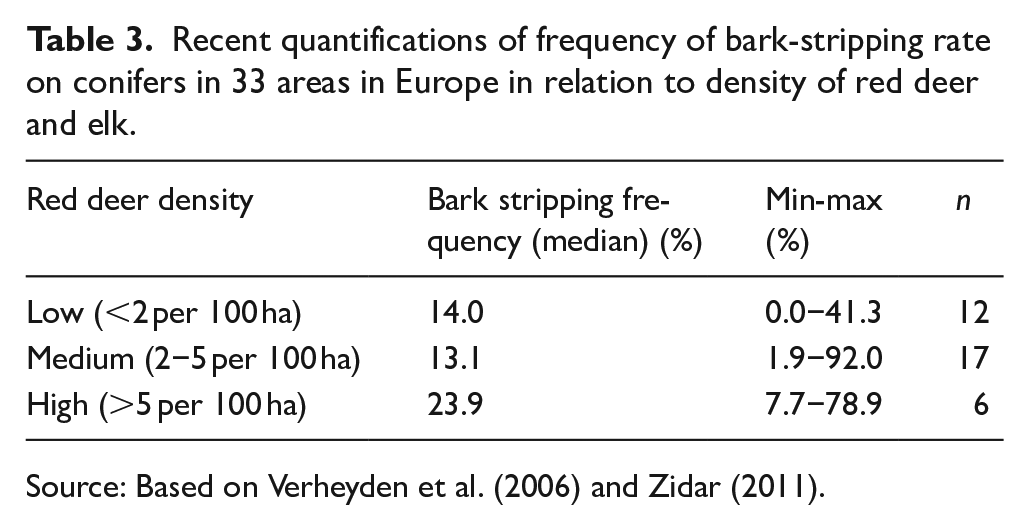
Source: Based on Verheyden et al. (2006) and Zidar (2011).
Comparisons between the Haväng material and the trees from the other sites show less wild-game damage toward the present day (Table 1). Damage was, for example, recorded in just 3.2% of the trees from Ljungaviken. Moreover, the location and shape of this damage makes them more likely to be caused by human activities in the area than by the wild game. Most of the damage we could observe on the trees from the Store Mosse area, however, were in line with the ungulate damage studied in the Eriksberg reserve. These observations are in good agreement with the presence of wild game in Ljungaviken and Store Mosse. There are hardly any large ungulates in the Ljungaviken area, while, for example, Eurasian elk are present in the Store Mosse National Park, which explains the greater proportion of bark-stripping damage (5.8% compared to 0%) on the trees adjacent to the National Park. The trees at Hällarydsmossen and Store Mosse grew in the same geographic region with a similar mosaic landscape composed of lakes, peatlands, fens, and forested hummocky moraine, although during different time periods. The larger proportion of bark-stripping marks on the trees from Hällarydsmossen can be interpreted as an indication that the wild-game population in the region was possibly larger during the mid-Holocene compared to present (Tables 2 and 3). In parallel, the decrease of fire scars in contemporary pine population points to the effect of contemporary forestry management with a reduced number of forest fires as a result. Similar trends have been observed in numerous tree-ring-based fire reconstructions (Dieterich and Swetnam, 1984; Pinto et al., 2020). Similarly, the two tree populations from the coastal regions, Haväng and Ljungaviken, can be compared. Once again, a decrease in bark-stripping damage is seen as the trees in the Ljungaviken clearcut logging site show no marks from wild-game while the early Holocene trees from the submerged Haväng forest show damage associated with ungulates on at least 15% of the wood samples (Table 2). To draw a parallel to contemporary studies (Table 3), we can state that the wild-game population in the region seems to have decreased even if we can not make a reliable estimate of how extensive the decrease has been.
The location of damage and marks
Given the varied size and distribution among the damage, it can be assumed that several wild-game species were present in the Haväng area during the early Holocene. Different ungulate taxa usually cause bark-stripping at different heights. For example, deer usually strip bark at heights of 0.5–1.0 m above ground while Eurasian elk mainly strip bark at heights between 1.0 and 1.7 m (Faber and Thorson, 1996; Gill 1992). Although the height of the bark-stripping damage on the trees could only be determined in a few cases, the variations show that ungulates of different sizes were present in the Haväng area. This assumption is supported by the faunal remains from Haväng showing the presence of aurochs, elk, and red deer, which are all large ungulates that were well adapted to the open pine forest surrounding the Verkeån River outlet during the early Holocene (Hansson et al., 2018a). It cannot be excluded that European bison and wild horse also have caused damages on the trees but finds of these species are relatively rare in relation to the mentioned three species or absent in the faunal record from south Sweden. Predators such as brown bear and Eurasian lynx can also cause damage to trees. However, the frequency of damages of carnivores on trees are low in relation to ungulates (Černe et al., 2019; Verheyden et al., 2006; Zyśk-Gorczyńska et al., 2016). Further, its most likely that the predator populations should have been small in relation to the ungulates that we can probably exclude their impact to the studied material.
Reaction wood
In over 12% of the material from the Haväng area, reaction wood could be observed. This indicates that the position of the trees has changed in such a way that the trees have been forced to change their growth pattern so as not to fall. Reaction wood is common in trees that grow on slopes and areas with ground movement (Gardiner et al., 2014), and can also be noted in trees affected by storms or in areas with a prevailing wind direction which causes the trees to tilt (Gaglioti et al., 2019). The submerged landscape in the Haväng area consists of loose, well-sorted sand and was relatively flat (Hansson et al., 2018a). Severe storms or soil movement due to water saturation are factors that may have resulted in the change of position of the tree and hence formation of reaction wood. However, another possible explanation for the trees to suddenly change position could have been caused by larger ungulates leaning or baring against the trees. We could study this phenomenon in detail in the Eriksberg reserve where large ungulates pushed or rubbed their bodies against young trees so that they began to lean and thus began to form reaction wood so as not to fall as they continued to grow. Many trees from Hällarydsmossen also show reaction wood. However, since these trees have grown in a soft and peaty substrate, hydrological changes may be the most likely cause of instability and subsequent formation of reaction wood among these trees (Edvardsson et al., 2016; Stoffel and Bollschweiler, 2008). Since there are many possible explanations for the formation of reaction wood, we have chosen not to use the occurrence of reaction wood in our argumentation for wild-game damage, even though large ungulates may have caused the growth anomalies in the trees in some cases.
Wild-game population dynamics
Our unique assemblage clearly shows the presence of large ungulates in the Haväng area during the early Holocene. Moreover, the comparison to wood material from additional sites and periods indicate a decrease in the large wild-game populations since the early Holocene. Additionally, changes over time, and the local composition of the fauna at the sites, may have caused trees to become more susceptible to damage by large ungulates. For example, aurochs occur relatively frequently in the early Holocene faunal assemblages from archeological sites in South Scandinavia. However, the population started to decrease around 8500, and by 7500 cal. yr. BP, aurochs were possibly extinct – or at least extremely rare – in Sweden (Magnell, 2017). Also, the frequency of Eurasian elk decreased during the same period. These two large ungulates seem to have been replaced by red deer as the primary, large herbivore in the region by 8000 cal. yr. BP (Magnell, 2017).
European bison have been shown to prefer tree bark during the winter and possibly the same could have been the case for the aurochs in the early Holocene (Zieleke et al., 2017). The higher frequency of bark-stripping at the Haväng site could possibly result from the presence of aurochs, which were probably rare or absent during the mid-Holocene. This statement is supported by data from Hällarydsmossen that show less bark-stripping than the Haväng material. The differences between the sites with recently felled forest, namely Ljungaviken and Store Mosse National Park, clearly reflect the variations in the wild-game populations between the two regions. In the same way, the less frequent damage from bark-stripping at these sites compared to Haväng and Hällarydsmossen also reflects how the large game population has decreased during the Holocene (Tables 2 and 3).
In a similar way, the differences between Hällarydsmossen and Store mosse could reflect changes in the fauna. At Hällarydsmossen red deer was probably the species responsible for the damage, and, since red deer form herds, it can result in relatively dense populations locally (Clutton-Brock et al., 1982). Eurasian elk – the most common large ungulate in the Store Mosse area – are generally solitary or live in smaller groups which may have resulted in less damage. The low-damage frequency on trees from the Ljungaviken area is most likely a reflection of a faunal assemblage with very few large ungulates present to cause bark-stripping.
To some extent, behavior and movement patterns can also be studied from the damage to the trees. Previously bark stripped trees were, for example, often reselected by the ungulates (Figure 3). This behavior can also be seen in 21% of the bark stripped trees at Haväng as they contain multiple scars (Figure 4a). The assumption that some trees are reselected by the ungulates is supported by results from several previous studies (Danell et al., 1985; Gill, 1992; Welch et al., 1991). We were also able to study this behavior in the Eriksberg reserve, where some trees seem to have been selected for bark-stripping or fraying, and to have been used repeatedly over many years (Figure 3b and c).
Possible error sources or biases
Studies based on submerged or subfossil wood will always have limitations owing to restrictions in the number of good samples available. However, the two examined tree populations from the early and mid-Holocene consist of more than 100 trees each. The information and data the trees generate may not give a complete picture of the study areas, but should be considered as relevant source of information about the environment the trees once grew in.
Another concern may be whether the studied trees are representative. It is possible that, due to increased resin content in the stems, damaged trees are better preserved during submersion than undamaged trees. But based on our observations of trees found submerged or in peatlands, we can conclude that the context in which the trees have been preserved is by far the most decisive factor in whether they become preserved or not. Over-representation of preserved resin-filled trees cannot be ruled out, but we do not have reason to assume that it is a significant factor for preservation.
The aims of a study will have a direct influence on the sampling strategy of the material. For example, if the purpose of the study is to develop tree-ring data for climate reconstructions, damaged samples will be excluded, whereas studies aiming to count damages from wild-game populations, fires, or slope processes will focus on damaged trees. The initial aim during the fieldwork campaigns for this project was to collect samples which could be used to reconstruct climate dynamics, environmental changes, and shoreline displacement. Since the primary goal was not to study wild-game population dynamics, damaged samples might have been excluded, which may have caused an under-representation of wild-game damaged samples.
The frequency of damage seen on the trees does not directly reflect the density of the game population; bark-stripping may depend on other factors and could be linked to local characteristics in the landscape. It is common that damage caused by ungulates is not evenly distributed and there are studies showing accumulations of damage in certain parts of forests (Gill, 1992; Welch et al., 1988). We could clearly observe similar patterns in the Eriksberg reserve, where the ungulates often selected certain trees and areas to which they repeatedly went back. During the early Holocene when the studied trees from the Haväng site grew, the area was characterized by a freshwater stream (Hansson et al., 2018a). This is an optimal environment that attracts the game for drinking and foraging. The relatively high amount of wild-game damage at the site does indicate a large game population. At the same time, the studied wood material may also show more bark-stripping than average for the time and region as the study area is assumed to represent a favorable environment for the animals. In parallel, the area was populated by humans as indicated by bone refuse and fishing constructions (Nilsson et al., 2018), so another possibility is that ungulates avoided staying too long near the watercourse owing to human activity.
The damage on the trees may also reflect environmental or climatic changes causing behavioral changes among the ungulates instead of the density in the wild-game population. In regions with harsh winters and extensive snow cover, for example, bark can comprise up to 10% of the red deer diet (Verheyden et al., 2006). In contrast, bark is not a part of the diet at all in regions where deer can find alternative food year-round. The interaction between food availability and wild-game density is complex and not straightforward to estimate. Since most of the compared trees are mature – 80–200 years in age – and the main periods compared (Preboreal, the Holocene thermal maximum, and present) are considered to be three relatively warm periods (Edvardsson et al., 2016; Wanner et al., 2008), we can assume that it is not likely an over-representation of severe winters resulting in limited access to food during one of the periods in comparison to the others. Moreover, tree-ring data from the sites do not show any correlation between periods with bark-stripping and severe conditions indicated by unusually narrow growth rings. We thus assume that food availability has a negligible influence when comparing the different periods.
The trees from the submerged landscape in the Haväng area have been sampled to make dendrochronological analysis aiming to study environmental and climatic changes. Most of the collected samples are therefore about 10 cm in thickness. In rare cases, meter-long segments of the trunks were sampled. For such samples, however, damages could be observed that could not be noted on thinner cross discs which were used during the dendrochronological analysis. Since the original purpose of the sampling was not to study the wild game, the sampling has not been done in an optimal way for that purpose. Damage may therefore have been missed if they were in a different area of the trunk than the part which was sampled. The difference between detecting and over-looking damage can be small, and in some samples, we could see the damage on the upper side of the cross section but not on the lower side of the disc even though the sample was <5 cm in thickness. Damage by wild game in the studied trees is therefore possibly larger than what was observed in the analyzed wood material.
In summary, we can see factors that may involve both over-representation and under-representation of wild-game damages. The material should therefore, when all these factors are taken into account, be considered as representative and give a reasonable picture of the presence of different ungulates in the Haväng areas during the early Holocene.
Conclusion and prospects
This study shows that damage and marks from bark-stripping and fraying on subfossil and submerged trees is an extremely underutilized source of valuable information about past wild-game populations. We therefore encourage the use of tree-ring analysis of such wood material as an indicator of ungulates, while simultaneously acknowledging that there are shortcomings that practitioners must be aware of. Such shortcomings may be, (i) the wood material that can be studied is usually very limited, (ii) the distribution of damaged trees within a tree population is not even, and (iii) the degree of conservation of damaged and undamaged trees may vary. The method is therefore insufficient for precise estimations of the wild-game density, but the presence of marks from ungulates is a solid indicator for wild-game activities at the study site. Dendrochronological analysis of prehistoric bark stripped wood should thus be regarded as a valuable source of information that can be complementary to archeology, osteology, species-specific or environmental DNA, and other sources of knowledge of prehistoric wild-game distribution, activities, and composition.
Among the objectives for the study to test to what level the wild-game density and dynamics could be reconstructed: (i) presence of the wild game, (ii) relative estimates of the wild-game population, or (iii) absolute estimates of the wild-game population. The fact that bark-stripping damage is observed on the wood indicates presence of wild game, and the fact that variations between different sites and periods can be made enables relative comparisons between different wild-game populations. Our study has reached the first two of these three levels, but the third has not as we have not been able to calculate with certainty how large the ungulate populations have been. It remains uncertain as to whether this step can be achieved.
Our results show that damage interpreted as being the result of bark-stripping by large ungulates were observed more frequently in the early Holocene material compared to the mid-Holocene material and present-day trees. This observation indicates larger wild-game populations during the early Holocene coastal site of Haväng than the inland mid-Holocene population represented by the Hällarydsmossen material. Furthermore, the submerged and subfossil tree samples indicate significantly higher frequencies of damage associated with ungulates in the past than in the region today. We do not intend to speculate on the reasons for this, but want to shed light on the material and methods we have used so that they are useful for future studies. When it comes to future studies, we want to draw users’ attention to there being a significant difference in the wild-game information that can be obtained from meter-long tree trunks instead of thin discs. This became especially clear during our studies in the Eriksberg reserve where we had unlimited access to material of different types and sizes. If the purpose is to detect and study wild-game damage, we recommend that meter-long segments of trunks are sampled, 3D-photogrammetrically documented, or at least carefully inspected before sampling so as to make sure that the desired parts are sampled.
Footnotes
Acknowledgements
We are grateful to Per-Arne Olsson, CEO at Eriksberg Nature Reserve, for his valuable knowledge, excursions, and help with the sampling of trees. We also thank Dr. Amber Hood for feedback and proofreading the text, as well as two anonymous reviewers for their valuable comments on the manuscript.
Author contributions
OM, JE, and BN designed the study together, but all authors contributed with ideas. JE, OM, AH and BN contributed to the writing of the manuscript, and HL and AS contributed with discussions and interpretations prior to and during the writing process. BN secured funding, AS and AH conducted the diving and main part of the fieldwork at Haväng. AH and HL did the dendrochronological analyses of the material from Haväng, and OM and JE later re-interpreted the damages on the trees. OM, BN, AS, and JE performed studies on trees in Ljungaviken and the Eriksberg reserve, and JE did the studies in Store Mosse National Park and with the material from Hällarydsmossen.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study is part of the project “Blue Archaeology. Assembling the cultural and natural heritage of submerged Stone-Age Seascapes in Sweden,” which was funded by the Swedish Research Council (Grant no: 2016:02027). Open access funding provided by Lund University.