
Abstract
The tropical forests of the Philippine Archipelago are some of the most threatened in the 21st century. Among the most prominent threats are the introduction of new plant and animal species, as well as new forms of land management (e.g. plantations), that have accompanied industrial expansion. Such threats have a potentially long-term history and prehistory in the Philippines, not just as a consequence of Spanish colonial administration and land-use changes from the 16th century, but also in the context of pre-colonial introductions of rice agriculture and domesticated animals. However, the impacts of such arrivals on local Philippine societies and ecologies have remained relatively unexplored, especially in comparison to contemporary exchanges between Europe and the Neotropics. Here, we evaluate archaeological and historical evidence for the integration of novel plants, animals and economic strategies into local Philippine cultures and economies from 4000 years ago to the 19th century AD. This includes material culture, archaeozoological and archaeobotanical analysis, as well as archival references to pre- and post-colonial urban settlements, the evolution of land management and rural settlements across the Archipelago. We argue that prehistoric land-use changes, as well as the colonial introduction of crops and domesticated animals, represent a potentially interesting contrast to other tropical regions that came under Spanish imperial control between the 15th and 19th centuries. Nevertheless, to determine the full extent of their impacts on social organisation and Philippine landscapes more detailed, long-term multidisciplinary investigation is required.
Introduction
The Philippines is a group of some 7100 islands in the western Pacific Ocean, 5° to 20° north of the Equator (Figure 1), and is uniquely rich in endemic fauna and flora (Sodhi et al., 2004). Today, the Archipelago is plagued by deforestation and soil erosion that have been argued to have a potentially deep history (Bankoff, 2013). The Philippine Archipelago is one of the largest combined landmasses in Island Southeast Asia to have witnessed three of the major pre-colonial Holocene land-use changes seen across Island Southeast Asia, the introduction of rice agriculture, the introduction of domesticated pigs and the formation of globalised ‘Metal Age’ and Protohistoric trading polities and port sites. It is also one of the few regions in Island Southeast Asia to have experienced Spanish conquest and the associated arrivals of new plants, animals and forms of political and social organisation. The Philippines thus has the potential to play a major role in pan-tropical discussions of changes in pre- and post-colonial subsistence, demographics and overall land-use – discussions that are becoming of increasing interest to climate and earth scientists attempting to determine the legacies left by prehistoric and historic societies on earth systems (Koch et al., 2019; Lewis and Maslin, 2015; Roberts et al., 2018). As an island system, like the Caribbean, the impact of prehistoric and historic translocations, particularly intensively-used domesticates, have the potential to induce major changes in landscapes and biodiversity (Fitzpatrick and Keegan, 2007). Meanwhile, access and use of large domesticated animals prior to European arrival may have given Southeast Asian populations more resistance against pandemic diseases, especially compared to the catastrophic mortality witnessed in the Neotropics (Koch et al., 2019; Newson, 2009). Nonetheless, while it was the Spanish conquest of the Philippines that united Manila, Mexico City and Madrid into the first truly globalised trade system, the majority of discussions of ‘Columbian Exchange’ translocations, their impacts and their predecessors have tended to almost entirely focus on the Atlantic sphere.
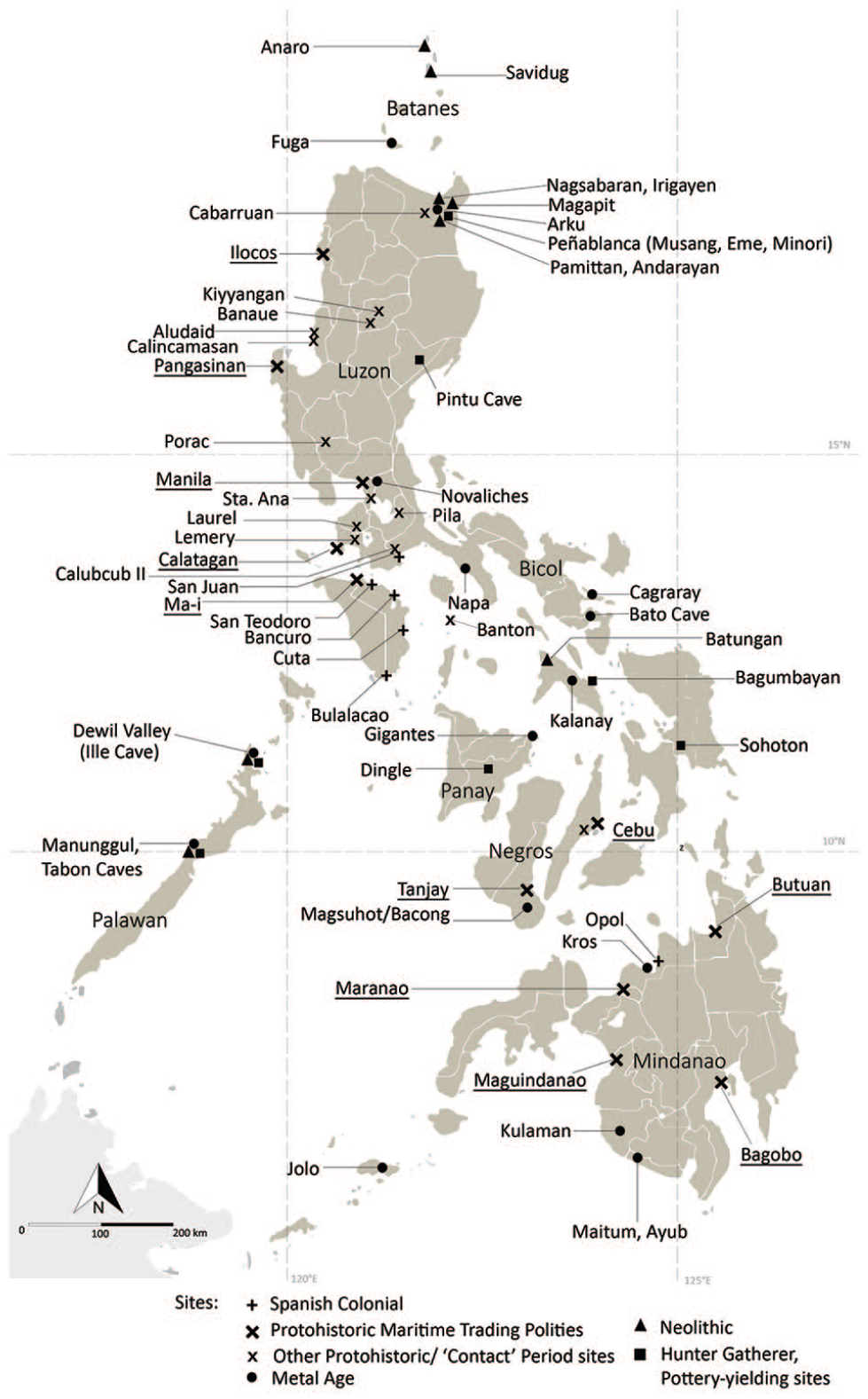
Location of key archaeological sites in the Philippine archipelago as well as known 10th–16th century maritime trading centres (underlined).
Almost a century of systematic archaeological research (see Paz, 2017; Ronquillo, 1985) has provided some understanding of the human past on the Philippine archipelago. However, discussions of prehistory have tended to focus on cultural typologies, centring on the ‘Three Age System’. The Palaeolithic period begins with earliest evidence of hominin presence ca. 700 thousand years ago (Ingicco et al., 2018), the Neolithic with the arrival of Austronesian speaking people ca. 2500–2000 BCE, together with agriculture and a suite of material culture including red-slipped pottery and polished stone tools (Bellwood, 1997, 2017; Bellwood and Dizon, 2013a), and the Metal Age ca. 500 BCE with the occurrence of metal working (Dizon, 1983, 1990), distinct pottery types (Solheim, 1970) and the emergence of socio-political ‘complexity’ (i.e. see Junker, 1994, 1999, 2002). The focus on material culture used to define these periods has often distracted from direct investigation into changes in subsistence and human impacts on diverse island landscapes in the region (Paz, 2017: 153), and changes in artefact types do not always appear to correspond to major changes in economy or diet (i.e. Amano et al., 2013). This is problematic given that, from 4000 years ago into the historic period, the Philippines witnessed the arrival of new modes of food production, new plants, new animals and, particularly with the arrival of the Spanish Empire, new ways of perceiving and managing land. A focus on culture-historical typologies has also meant that there has been a general lack of data with which to build or develop palaeoecological understandings of the longevity of trends in human environmental impacts or with which to construct land-use maps for inclusion within earth system models (as per Kay and Kaplan, 2015; Kay et al., 2019) until recently.
Here, we provide a review of the available prehistoric and historic evidence of animal and plant introductions to the Philippine Islands and the potential ecological impacts of these introductions and changes in land-use from 4000 years ago to the 19th century CE. We structure our evaluation using the Three Age system of Philippine prehistory (Figure 2), with full awareness of its limitations and a clear note that we use these terms in a purely regional context. Comparisons should not necessarily be made with similar terms used elsewhere without caution. Nonetheless, there is currently no practical alternative since most archaeological evidence has been reported using this framework. We analyse the archaeological evidence for the Neolithic introduction of domestic animals and possibly rice ca. 2500 years ago and archaeological and historical insights into the introduction of American crops and novel land-use and administration during the Spanish Colonial Period (1521–1898 CE). We suggest that the existing archaeological record demonstrates no ‘Neolithic Revolution’ in land-use following the introduction of domestic animals, and evidence for large-scale prehistoric rice agriculture remains elusive. There was also no apparent widespread intensification in animal husbandry until the Protohistoric period. The Spanish colonial period saw the introduction of New World plants to the archipelago, including crops, vegetables and fruit trees, although there is currently limited archaeological research looking at habitation sites from this period. In contrast to many areas of Latin America, however, it appears that the beginning of large-scale land conversion for plantations of New World crops only occurred following the Bourbon Reforms and the opening of the Port of Manila to world trade during the second half on the 19th century.
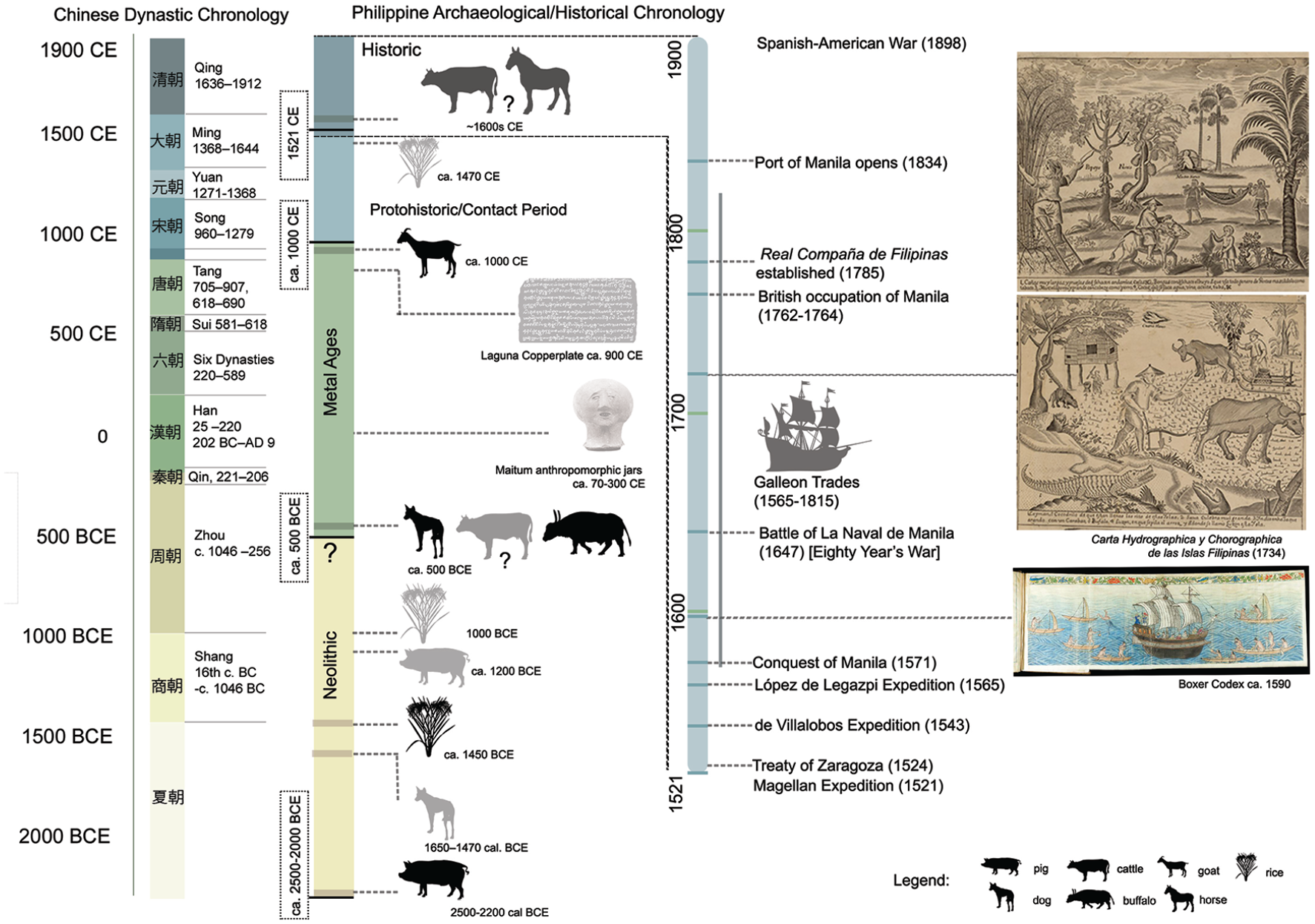
General timeline of Philippine prehistory/history, from ca. 2000 BCE to the 18th century CE.
The Arrival of Agriculture
Pawlik and Piper (2019) recently reviewed the ‘Pre-Neolithic’ archaeological record in the Philippines from ca. 12,000–2000 BCE, arguing that the region has played a central role in the movements of people, material culture and ideas across Island Southeast Asia as early as the Late Pleistocene. They note that the Philippine archipelago seems to have been part of a vast maritime network connecting Southeast Asia to Near Oceania, as evidenced by common shell and lithic technologies (i.e. Barton et al., 2009; Mijares, 2006; Neri et al., 2015; Pawlik, 2012; Pawlik et al., 2015) and shared complex burial traditions, including secondary inhumations and cremations (i.e. Detroit, 2006; Lara et al., 2013, 2016; Lewis et al., 2008; O’Connor et al., 2017; Szabó et al., 2007). This maritime network was apparently well in place when early farming, Austronesian-speaking communities arrived in the Philippine Islands between 2500 and 2000 cal. BCE from China via Taiwan (Bellwood, 1997, 2002, 2017), likely stimulating the movement of novel domesticated plants and animals. Nevertheless, this Out of Taiwan model for Austronesian expansion, as well as the accompanying subsistence changes, has been hotly debated over the course of the last forty years (i.e. Bellwood, 2005; Blench, 2010; Blust, 1984; Denham 2013, 2018; Hung et al., 2011; Simanjuntak, 2017; Spriggs, 2011).
Whatever their route and date of arrival, Austronesian communities are assumed to have introduced a new subsistence economy based on cereal agriculture and the raising of domestic animals to the Philippine Archipelago, as well as a distinct material culture (Bellwood, 2002, 2017; but see Barretto-Tesoro, 2016; Denham, 2018; Paz, 2002; Spriggs, 2011). Systematic research in the past three decades has produced clear evidence for the material culture associated with Austronesian communities. Excavations in Batanes Islands in the northern Philippines produced evidence for red slipped pottery from ca. 4000 BCE (Bellwood and Dizon, 2005, 2008, 2013a; Bellwood et al., 2013a, 2013b) alongside other material culture associated with the Austronesian expansion, including polished stone adzes and notched pebble sinkers (Bellwood and Dizon, 2013b; Duff, 1970) and baked clay spindle whorls (Cameron, 2013) dating to ca. 1500–500 BCE. The 30 or so shell midden sites along the Cagayan River in northern Luzon provide the best evidence for the Neolithic material culture in the Philippines (Aoyagi, 1977; Cabanilla, 1972; Hung, 2009; Ogawa, 1998). These sites, dated from ca. 3000 BCE (but see Hung, 2009) to the first millennium BCE, include Catugan (Tanaka, 1998), Bangag (Tanaka, 1997), Gaerlan (Garong, 2002), San Lorenzo (Tanaka, 2003), Magapit (Aoyagi et al., 1997; Thiel, 1980), Irigayen (de la Torre, 2000; Ogawa, 2005), Pamittan (Tanaka and Orogo, 2000), Andarayan (Snow et al., 1986) and Nagsabaran (Hung, 2009).
Sites are rare outside northern Luzon, with only Batangas in southern Luzon (Barretto-Tesoro, 2016), Batungan in Masbate (Solheim, 1954, 1968), Batang and Duyong (Fox, 1970) and Ille (Lewis et al., 2008) in Palawan providing evidence for Neolithic presence. The limited archaeological research conducted on Neolithic sites in other parts of the Philippines could be attributed to the difficulty of finding settlements rather than from deliberate omission. Bellwood et al. (2008), in their search for sites in the Ilocos region of western Luzon, noted that high sedimentation rates during the past 2000 years most likely buried Neolithic settlements, most of which would have been close to rivers and estuaries, making their detection difficult. A lack of settlement sites, particularly beyond Luzon, as well as a focus on material culture typologies, notably pottery sequences (see Ogawa, 2005; Tsang, 2007) and, in some cases, poor preservation of organic remains has meant that investigations into the nature of the subsistence economy, and understanding of the various importance of new domesticates in the diets and land-use, of Neolithic communities are limited. In-depth studies looking at subsistence strategies have been restricted to sites in Batanes Islands (Bellwood and Dizon, 2013a) and a couple of sites along the Cagayan River Valley (Hung, 2009), while the number of archaeobotanical and archaeozoological practitioners operating in the Philippine Archipelago has been limited to a handful of dedicated researchers.
The agriculture of early Austronesian communities is assumed to have centred on rice (Oryza) and millet (Panicum/Setaria) cultivation (Bellwood, 1997). However, there is no archaeological evidence for early millet cultivation in the Philippines or, for that matter, anywhere in Island Southeast Asia (Denham, 2018). There is also a scarcity of hard evidence for early rice agriculture in the Philippine Archipelago, owing to the limited amount of archaeobotanical investigations conducted in the region and poor preservation of botanical remains in sites dating to the early Neolithic period. The site of Andarayan in northern Luzon has provided the earliest evidence for the presence of rice in the Philippines (Snow et al., 1986). Rice husks and stem fragments embedded in a single earthenware sherd from the site returned direct radiocarbon dates of 1500–1400 BCE. Snow et al. (1986: 4) noted that the spikelet morphology of the specimens is ‘intermediate between cultivated rice (O. sativa) and its immediate wild relative (O. rufipogon or O. nivara)’. Recently, more conclusive evidence for local rice cultivation in northern Luzon was reported by Deng et al. (2018). Charred rice grains from Magapit site were directly dated by AMS radiocarbon to ca. 1000 BCE. The rarity of rice remains, not only in the Philippines but also in other early sites in Island Southeast Asia, has led researchers such as Hayden (2003, 2009), Barton (2009, 2012) and Acabado (2012; 2018) to hypothesise that rice was initially cultivated as a prestige crop and widespread cultivation as a staple occurred only during the last 2000 years with the onset of the Metal Age. Instead of rice, taro (Colocasia esculenta) and yam (Dioscorea) were presumed to have been the staple crop for early communities in the Philippine islands (i.e. Acabado, 2012), as in other parts of Island Southeast Asia and Oceania (Barton and Denham, 2011, 2018; Spriggs and Matthews, 2012), though the remains of these soft root crops can be difficult to find. Charred taro remains were reported by Tsang (1995) from Lal-lo site in northern Luzon. These remains yielded 14C dates of ca. 3900–3300 BCE.
Direct evidence for the presence of domestic animals in the Neolithic Philippines is more robust. Three domestic animals, the dog (Canis lupus familiaris), pig (Sus scrofa) and chicken (Gallus gallus), as well as commensal rats (Rattus exulans), are traditionally considered as part of this introduced Neolithic package, all of which have been argued to have had potential ramifications for local landscapes and endemic floral and faunal diversity in the Pacific (see Spriggs, 2001; Swift et al., 2017). The earliest evidence for domestic pigs in the Philippines comes from Nagsabaran, where a mandibular premolar yielded direct radiocarbon dates of 2500–2200 cal. BCE (Amano et al., 2013; Piper et al., 2009a). Domestic pigs were also present in the early Neolithic levels of sites on the islands of Itbayat and Sabtang in Batanes from ca. 1200 BCE (Piper et al., 2013). Elsewhere in the Philippines, possible domestic pigs were identified in ca. 500 BCE–1000 CE layers of Sohoton I (Mudar, 1997) as well as in excavations in Bohol Island (Alba, 1994). Possible domestic pigs were also identified in the Metal Period layers of Ille Caves (Amano, 2011), though these are currently tentative identifications because of the difficulty of differentiating domestic pigs from the wild Palawan bearded pig (S. ahoenobarbus) based on dental measurements and morphology (see Ingicco et al., 2017).
The earliest evidence of dogs in the Philippine archaeological record was reported from the Terminal Pleistocene and Early Holocene layers of Ille Cave in northern Palawan (Lewis et al., 2008; Piper et al., 2011). The remains most likely represent local extinct Asiatic wild dogs (Cuon alpinus) that reached the island during periods of low sea levels (Piper, 2017; Piper et al., 2011). Evidence for the presence of dogs were also reported from Callao Caves in Peñablanca, northern Luzon in layers with associated charcoal 14C dates of 1650–1470 cal BCE (Mijares, 2006; Piper et al., 2013). However, Piper et al. (2013: 197) noted that the association of the dog remains with the date is rather tentative and the second millennium BC presence of dogs in the Philippines remains uncertain. Indisputable evidence for the presence of domestic dogs in the region comes from Savidug and Anaro in Batanes (Piper et al., 2013) and Nagsabaran in northern Luzon (Amano et al., 2013; Piper et al., 2009b) from ca. 500 BCE. At both Savidug and Nagsabaran, numerous dog bone fragments exhibited butchery marks suggesting that dogs were most likely consumed. A dog burial was recorded at Nagsabaran, also dating to ca. 500 BCE or slightly earlier (Amano et al., 2013), hinting at a complex dog-human relationship. Dogs are common in the Metal Age and Protohistoric periods, with dog burials reported in sites such as Ille in Palawan (Ochoa, 2009) and Sta. Ana in Manila (Vitales, 2018).
Evidence for chicken in the Philippine archaeological record currently remains elusive. A possible chicken remain was reported by Piper et al. (2013) at Savidug site from layers dated to ca. 500–300 BCE. In Nagsabaran, not a single chicken remain was identified even though a number of well-preserved bird bones were recovered (Amano et al., 2013). Mudar (1997) also failed to identify any chicken remains in the different prehistoric sites she looked at, save for two fragments from Tanjay site in layers dating to ca. 1300–1600 CE. However, genetic evidence potentially hints towards the Philippines being the possible origin of chickens introduced to Remote Oceania. Thomson et al. (2014a), looking at mitochondrial DNA from archaeological and modern chicken samples from Polynesia and Island Southeast Asia, identified a unique genetic signature (the ancestral ‘Polynesian motif’) in archaeological samples from Niue, Hawai’i and Rapa Nui. This unique genetic signature was identified in modern samples from the Philippines, pointing to the archipelago as the possible homeland for these Polynesian chickens.
Evidence for the presence of the commensal Pacific rat (Rattus exulans) also remains elusive in the Philippine archaeological record, mostly due to the difficulty of identifying the taxon based on morphology alone. Piper et al. (2013) identified a Rattus mandible from the Savidug site in the Batanes Island suggesting that a ‘commensal species inhabited the islands during prehistory’ (p. 198). Rat-size murid bone that cannot be specifically assigned to taxon were identified by Amano et al. (2013) and Piper et al. (2009b) from Nagsabaran site. Genetic evidence paints a rather complicated history of R. exulans introduction to the Pacific. Modern R. exulans specimens from the Philippines were shown to be part of two haplogroups, one consisting of samples from the Philippines, Borneo and Sulawesi and the other distributed from the Philippines to New Guinea, the Bismarck Archipelago and the Solomon Islands (Matisoo-Smith and Robins, 2004). The second haplogroup has been linked to the maritime exchange network connecting the Philippine archipelago to Near Oceania. Subsequent genetic analyses conducted by Thomson et al. (2014b) suggest the island of Flores in Indonesia as the possible homeland of R. exulans introduced to other parts of Island Southeast Asia and Oceania. Given that rats have been shown to have dramatic impacts on island ecosystems elsewhere in the Pacific (Swift et al., 2017), further investigation of the environmental impacts of these taxa across the varied islands of the Philippines warrants further research.
Overall, like rice, it appears that domesticates, specifically pigs, were raised by early farming communities as prestige or ritual animals rather than the main protein source, and there is no evidence of a concerted pastoral economy. At Nagsabaran, although domestic animals were present, the faunal assemblage was dominated by wild taxa suggesting heavy reliance on hunting for subsistence (Amano et al., 2013; Piper et al., 2009b). There is one domestic pig for every three or four wild pigs in the Nagsabaran assemblage. As Amano et al. (2013) point out, there appears to be an implicit assumption that domestic animals played a vital economic role among early farming communities in the Philippines, perhaps because of associations drawn with early Neolithic settlement sites in South China and Mainland Southeast Asia. However, as Hayden (2001, 2014) and Barker (2007) argue, the role of domestic animals in the subsistence of early agriculturalists might have been over-emphasized. Instead they argue that domestic animals might have played a role in rituals, or as indicators of social status in community interactions, as ethnographic data suggest they do within contemporary indigenous societies in Southeast Asia (i.e. Ayalew et al., 2011; Jenks, 1905; Lemonnier, 2002). Overall, the arrival of domestic plants and animals from outside the Philippines seems to have been limited and gradual. Instead of transforming land-use, these arrivals seem to have rather been fit within indigenous models of food productions that centred on the exploitation of root crops, the hunting of wild animals and fishing in freshwater and marine contexts.
In addition to domestic and commensal taxa, several wild animal species are hypothesised to have been introduced either deliberately or unintentionally within the Philippine archipelago perhaps during the Late Holocene. These include the Asian house shrew (Suncus murinus), the Malayan civet (Viverra tangalunga), the Asian palm civet (Paradoxurus hermaphroditus) and the long-tailed macaque (Macaca fascicularis) (see Piper et al., 2011). The exact timing of these introductions, however, remains to be fully elucidated because of the scarce zooarchaeological information on these species outside the island of Palawan where they are native. For example, although long-tailed macaques are currently found in all islands in the Philippines (Abegg and Thierry, 2002; Fooden, 1991), no macaque remains were identified in the faunal assemblage of Nagsabaran (Amano et al., 2013; Piper et al., 2009a, 2009b) or in any Neolithic sites along the Cagayan River in northern Luzon. In her analysis of faunal remains from the Pintu/Busibus rockshelter site in northern Luzon, Mudar (1997) found macaque remains only on the upper (undated) layers of the site. Similarly, the Asian palm civet is currently found in the Philippine archipelago but in-depth analyses of several sites in Luzon did not yield evidence for their presence during the Early or Mid-Holocene (Amano et al., 2013; Mudar, 1997; Piper et al., 2009a, 2009b). Interestingly, Piper et al. (2013) identified several civet cat remains in Anaro site in the Batanes Islands from layers dated to 1000 BCE to recent history. More research is needed to determine the timing of these introductions, and their impacts on the environment. Nonetheless, translocations of small to intermediate-size animals, including macaques and civet cats into Wallacean islands have been well documented (i.e. Heinsohn, 2001, 2003; van den Bergh et al., 2009).
Domestic Animals and Crops in the Metal and Protohistoric Periods
The Metal Age in the Philippines commenced ca. 500 BCE with the first appearance of metal artefacts in the archaeological record (Dizon, 1983), though the exact timing of the arrival of metallurgy or metal artefacts in the Philippines remains to be precisely elucidated. While in other tropical regions, such as West and Central Africa, the introduction of metal artefacts, namely iron, has been linked to significant deforestation as a product of mining and smelting (e.g. Bayon et al., 2012; Garcin et al., 2018; Kay et al., 2019) there is currently no clear evidence for such activities in the Philippines dating to this period. Rather, iron working is hypothesized to have been limited to processing local lateritic ores in small furnaces, as evidenced by recovery of slags in several archaeological sites (see Dizon, 1983; Junker, 1999; Paz, 2017). Bronze and gold working have also been reported (Dizon, 1983; Fox, 1968, 1970), but mostly for the manufacture of weapons and ornaments rather than agricultural implements or house/boat-building tools (Junker, 1999: 271). As pointed out by Paz (2017), the majority of Metal Age sites are defined not by the presence of metal artefacts but instead by distinct pottery styles and forms (i.e. Solheim, 2002). The Metal Age was followed by the Protohistoric or Tradeware Ceramics period which is characterised by the presence of high-fired ceramics from China and Mainland Southeast Asia (Fox, 1970). Commencing around the 9th–10th century CE, this period is particularly marked by late Tang dynasty tradeware ceramics. The Protohistoric period is also characterized by the rise of chiefdoms linked to trade networks with Chinese dynasties as well as the thalassocratic empires of Srivijaya and Majapahit based in Sumatra and Java, respectively (Bacus, 1996; Hutterer, 1981; Junker, 1994, 1999; Paz, 2017).
A considerable amount of research on Metal Age and Protohistoric sites has been done, although excavations have mostly focussed on burial sites. Junker (1999: 43) attributes this to the difficulty of locating habitation sites given a lack of monumental constructions in stone, unlike those seen in contemporaneous societies in mainland Southeast Asia. Key Metal Age sites include Ayub Cave in southern Mindanao (Dizon, 1996) and Manunggul Cave in Palawan (Fox, 1970), which are notable for their anthropomorphic earthenware vessels which were used as burial jars. Other sites include Kalanay in Masbate (Solheim, 2002), Napa Site (Paz et al., 2016) and Bato Cave (Fox and Evangelista, 1957) in southern Luzon, Magsuhot/Bacong in Negros (Tenazas, 1974) and Seminoho Cave (Kurjack and Sheldon, 1970; Kurjack et al., 1971) and Kulaman Plateau (Maceda, 1965, 1966) in Mindanao. Research in the Protohistoric period has also focussed on large cemeteries including the 14th–15th century site of Calatagan in Batangas (Barretto-Tesoro, 2008; Fox, 1959) and Sta. Ana in Manila (Fox and Legaspi, 1977; Peralta and Salazar, 1974). Nevertheless, centres of historically known polities such as Butuan in Mindanao (Brown, 1989; Burton, 1977; Lacsina, 2016; Ronquillo, 1987), Cebu (Hutterer, 1973, Nishimura, 1988, 1992), Tanjay in Negros (Hutterer, 1981; Junker, 1993a, 1993b, 1996) and Manila (Peralta and Salazar, 1974) have been subject to long-term, systematic excavations and have provided important data on the nature of pre-Hispanic urban centres.
While industrial activities seem to have been relatively limited, and focus on cemeteries has limited insights into the livelihoods of Metal Age and Protohistoric communities, archaeological evidence does point to the introduction of some new domestic animals, including the water buffalo, goat and possibly cattle. Excavation on the islands of Sabtang and Itbayat in Batanes produced evidence for the presence of goats in northern Philippines by the 12th century CE (Piper et al., 2013). Early Spanish archival records emphasised the role of goats as trade and food items in Philippine chiefdoms (Scott, 1990: 302). Pigafetta (1550–1559) in his chronicle mentioned goats being offered by the local chief as a gift to Magellan:
‘On Friday, April twenty-six, Zula, a chief of the island of Matan [Mactan], sent one of his sons to present two goats to the captain-general, and to say that he would send him all that he had promised. . .’ (in Blair and Robertson, 1906, vol. XXXIII: 175)
Although in other parts of Island Southeast Asia, particularly at the sites of Sembiran and Pacung in Bali ca. 700 BCE–70 CE (Fenner et al., 2018), goats have been hypothesised to have arrived from India or Sri Lanka as part of wide-ranging maritime networks, in the Philippine case, goats were most likely introduced to the Philippines from China just after the 11th century CE. Indeed, their presence in the archaeological record coincides with increased occurrence of Chinese tradeware ceramics – for instance, in 12th–15th century deposits in the polity of Butuan in Mindanao (Bautista, 1990).
The timing and nature of the introduction of domestic water buffalo to the Philippines remains something of an open question. Historical accounts suggest that water buffalo populations, both maintained and wild (possibly feral), were well established in the Philippine islands by the time of Spanish arrival. de Morga in 1609 wrote:
‘There are many buffaloes, which are called carabaos, which are raised in the fields and are very spirited; others are brought tame from China; these are very numerous, and very handsome. These last are used only for milking, and their milk is thicker and more palatable than that of cows’. (in Blair and Robertson, vol. XVI, 1907:90).
Early Spanish accounts also note the hunting of wild buffaloes which most likely represent feral populations. Miguel de Loarca in his Relacion de las Yslas Filipinas (1582) noted:
This province [Pangasinan] abounds in food supplies, such as rice, goats, and swine; and many buffaloes are hunted.
In these islands are very many swine, and goats of excellent quality. There are also a great many wild buffaloes, which, if caught when young, can be easily tamed’. (in Blair and Robertson, 1906, vol. V:105).
Mudar (1997) reported remains of large bovids, which she identified as water buffaloes, from Sohoton I Cave site in southern Luzon, in layers dated to 10,500 BCE. These Early Holocene bovid remains most likely represent an extinct wild endemic species. Piper and Mijares (2007) also recorded an extinct bovine in 67,000 year old deposits from Callao Cave in northern Luzon. Two endemic bovids, one extant and another extinct, are known from the archaeological/palaeontological record of the Philippines. The extant Bubalus mindorensis endemic to the island of Mindoro has been recorded in Late Pleistocene/Early Holocene hunter-gatherer sites (i.e. Boulanger et al., 2019; Pawlik et al., 2015). A dwarf buffalo (Bubalus cebuensis) has been described by Croft et al. (2006) from Cebu Island and is presumed to have gone extinct during the Holocene. Interestingly, Cuevas et al. (2010), in their excavation of a pre-Hispanic settlement site in Central Cebu, retrieved a complete skeleton of a dwarf bovid, possibly suggesting that endemic bovids were present in the island as late as the 13th century CE. The earliest possible domestic buffalo remains were reported by Mudar (1997) from the Pintu/Busibus rockshelter site in northern Luzon in layers dated to the first millennium BCE and by Amano et al. (2013) from Nagsabaran site from layers dated to ca. 500 BCE. These remains show evidence of butchery suggestive of consumption. Early ethnographic accounts also highlight the role domestic buffaloes played in Philippine chiefdoms as a prestige and ritual animal, especially in feasting, as well as for traction in agriculture (i.e. Boxer Codex 1590 in Cole, 1913, 1922; Jenks, 1905; Jocano, 1975).
The first presence of cattle in the Philippine archaeological record was reported in the Metal Age deposits of Nagsabaran site in northern Luzon from layers dated to ca. 500 BCE (Amano, 2011). The specimen, an upper second premolar, is the only cattle skeletal element recorded in Nagsabaran, or indeed in any other pre-Hispanic sites in the Philippines, so the identification should be treated cautiously at present. Mudar (1997) recorded cattle remains only within the Spanish occupation levels of Cebu and she did not identify any cattle elements in the Metal and Protohistoric sites she looked at. No cattle remains were recorded in the precolonial and colonial layers of Kiyyangan in northern Philippines (Acabado et al., 2019) and likewise in the 12th–15th century deposits of Butuan in Mindanao (Bautista, 1990). These conform to the available secondary historical literature which indicate that the introduction of domestic cattle occurred in the 16th century (Scott, 1990: 302). de Morga mentioned (in Blair and Robertson, vol. XVI, 1907: 89) that:
‘Beef is eaten, cattle being raised abundantly in stock-farms in many different parts of the islands. The cattle are bred from those of China and Nueva España’.
While there has been very limited research done on archaeobotanical remains from the Metal Age and Protohistoric periods, existing historical records suggest that rice, taro, banana, coconut and sugarcane were also increasingly integrated into the subsistence economies of the Philippines by the time of Spanish arrival, with potential widespread modification to Philippine landscapes (Alcina, 1668; Blanco, 1837; de Morga, 1609). Pigafetta (1550–59) noted of the people he encountered from central Philippines, ‘They eat coconuts . . . figs one palm in length [bananas], sugarcane, and flying fish, besides other things’. (in Blair and Robertson, 1906, vol. XXXIII: 99). These crops were planted in swidden fields irrigated with natural drainage (i.e. Murillo Velarde, 1734 illustration; see Figure 2). de Morga (1609) observed:
‘. . . their settlements; for they always build them on the shores of the sea, between rivers and creeks. The natives generally gather in districts or settlements where they sow their rice, and possess their palm trees, nipa and banana groves, and other trees (in Blair and Robertson, vol. XVI, 1907:117)’.
In particular, the Protohistoric period has been hypothesised to have witnessed large-scale land transformation for terracing and irrigation. This has often been linked to increasingly intensive rice cultivation but may also have been associated with root crops (i.e. Acabado, 2012; Peterson et al., 2005). Rice had become the preferred staple in many regions, although in several areas Spanish chroniclers observed a heavy reliance on root crops, such as taro and yam for subsistence, stating that rice production was insufficient (Scott, 1990). Junker (1999: 330–331) argued that because of the high amount of labour required for production, rice was considered as a high-status food by many groups, for instance as suggested by archaeological investigations in northern Luzon (Acabado et al., 2016). Gunn (1995, 1996, in Junker, 1999) in her study of charred macrobotanical remains from settlements sites in Tanjay noted the relatively higher predominance of rice remains in areas presumed to have been occupied by elites or members of the ruling classes in comparison to non-elite habitation zones. Historical accounts emphasize that dry-land rice cultivation in hillside swidden fields was a common practice (Scott, 1990, 1994). Large scale wet-rice cultivation appears to have been limited to certain regions, particularly in river valleys with high population densities such as the Pangasinan region in Eastern Luzon, the Tagalog region near Manila, and the Bicol Peninsula (Alcina, 1668; Scott, 1990, 1994). Lowland terraces for wet-rice agriculture have been reported in the historical record in Tayabas in southern Luzon (Manuel, 1994) and the Bicol region (Scott, 1994), although their exact age remains to be ascertained.
The extensive wet-rice terraces in the cordillera region of northern Luzon represent one of the best examples of pre-colonial landscape modifications for agriculture in the Philippine Islands. First mentioned in the Spanish historical record in the late 18th century (Antolin, 1789; Scott, 1974), radiocarbon dates showed that, contrary to previous arguments, the terraces were built between 1486 and 1788 CE, after the arrival of the Spanish, perhaps by people moving inland away from Spanish conquest (Acabado, 2009, 2017). Investigations of terraces in the nearby Kiyyangan site show a marked shift to wet-rice agriculture from taro cultivation, with intensification occurring in response to Spanish colonisation (Acabado, 2012, 2015; Acabado et al., 2019). Research at the site provided evidence for wet-rice cultivation, in the form of phytoliths and starch from terrace sediments, from as early as 13th century CE (Horrocks et al., 2018). By contrast, investigations in Carcar in Cebu Island by Peterson (2005) of presumed wet agriculture terraces did not yield any evidence for rice cultivation. The terraces, dated between 1000 and 1300 CE, instead indicate the cultivation of sugarcane and taro, showing the variation in pre-Spanish Philippine economies. Sugarcane (Saccharum sp.) appears to have been well established in many parts of the Philippines by the time the Spanish arrived. Initially domesticated in New Guinea, sugarcane (S. officinarum) was introduced to Indonesia, China and India where they were hybridised with other species (i.e. S. spontaneum; Grivet et al., 2004; Moore et al., 2013). Donohue and Denham (2010) noted that linguistic reconstruction points to sugarcane being present in Taiwan before the Austronesian dispersal and therefore suggesting a pre-Neolithic westward movement of the crop from New Guinea. By around 500 CE, sugarcane has been introduced to southern Arabia and East Africa (Paterson, 2012).
A lack of obvious settlement structures in many pre-colonial sites has often led focus on the Spanish period when considering major historical changes in Philippines settlement patterns and the way people occupied tropical Philippine landscapes. Nevertheless, work has explored major changes in settlement patterns between the 9th and 14th centuries CE in the Archipelago. The pioneering research of Hutterer and Nishimura (i.e Hutterer, 1973, 1974; Nishimura, 1988, 1992) at Cebu, for example, shows how this site developed into a major urban centre with increasingly specialized areas for socioeconomic and political power, specialized craft production including ironworking, and agricultural activities. Research at Manila (i.e. Peralta and Salazar, 1974) Butuan (Bolunia, 2013; Burton, 1977; Lacsina, 2016) and Sulu (Spoehr, 1973) have provided similarly important insights into pre-Hispanic growth of trading centres during the Protohistoric period. A long-term, regional-scale research project at Tanjay in Negros Oriental provides perhaps the most detailed information on changes in landscape use and settlement patterns associated with an emerging maritime chiefdom in the Philippines between the Metal Age and Protohistoric period. Excavations of eight settlement areas, including primary regional centres and upland homesteads, and surveys of more than 500 archaeological sites occupied between 500 and 1600 CE (Hutterer, 1981; Hutterer and Macdonald, 1979, 1982; Junker, 1994, 1996; Junker et al., 1994, 1996) document how, from a two-level settlement hierarchy in the first millennium CE, by 1600 CE, Tanjay had become a 50 ha centre, with several secondary settlement centres of 4 to 7 ha (Junker, 1999: 113–118). This example of a dendritic system extending political and economic from a single primary centre over radiating settlements seems to be repeated in several island southeast Asian chiefdoms from the first millennium and early second millennium CE (Bronson, 1977; Junker, 2000, 2006; Santley and Alexander, 1992).
The Arrival of New World Plants and Animals
The Spanish colonial period in the Philippines can be divided into three broad periods (i.e. Skowronek, 1998), each with a different likely imprint on Philippines economies, societies and landscapes. The earliest is defined by the initial Spanish expeditions to the Philippine islands, the first led by Ferdinand Magellan (1521), then by Ruy López de Villalobos (1543) and Miguel López de Legazpi (1565), up to the conquest of Manila in 1571. While in Latin America, the initial forays of Spanish colonisers led to massive reductions of population through genocide, warfare and the transmission of lethal diseases (i.e. Abad et al., 2012; Lovell, 2019), Spanish conquest appears to not have resulted in the same scale of demographic collapse in the Philippines although abuses of the local population were certainly frequent (Newson, 2006, 2009). The second period of the Spanish colonial period in the region is known as the Galleon Trade period, which lasted from 1573 to 1815. The Galleon Trade is seen by many as heralding the start of a globalised world economy (e.g. Cushner, 1971, 1976; Flynn and Giráldez, 2008; Peterson, 2014; TePaske, 1983). Two galleon ships sailed from the port of Manila to Acapulco in Mexico (then New Spain) in most years, bringing with them products from Asia such as precious stones, ivory and cotton from India and Sri Lanka, spice from Indonesia, and silk, jade, sandalwood, copper, iron and porcelain from China, and returning to Manila carrying American silver (e.g. Chaunu, 1960; Mehl, 2016; Schurz, 1959; Skowronek, 1998; Wang, 2019; Wu et al., 2019).
The Galleon Trade period saw the introduction of a wide array of plants native to the New World to the Philippines, plants that are known to have had significant repercussions for farming and populations in other parts of Europe and Asia (Flynn and Giráldez, 2017; Machuca, 2014). Nevertheless, Machuca (2014: 236) has pointed out that the introduction of American plants to the Philippines was ‘neither coercive nor deliberate’ since, unlike in New Spain, the Spanish government did not seek to implement a European agroeconomic system on the islands, nor did they have the presence or administrative control to do so had they so wished. Rather, New World plants were slowly introduced by individuals to be planted in the estates of religious orders that were the main drivers of administrative, social and economic change in the Archipelago prior to the 18th century CE (Roth, 1977). By the late 18th century CE, several New World plants had been successfully introduced to the Philippine Islands. In his Flora de Filipina (1837), the botanist Fransico Manuel Blanco provided a comprehensive list of plants present in the Philippines that, in addition to endemic species, included introduced flora. Figure 3 illustrates some of the New World plants present in the Philippines by the late 18th century, many with their Nahuatl (Aztec language) name preserved.

American plants in the Philippines ca. 16th–18th century introduced through the Galleon Trade; Latin name (English common name/common name in Mexico/common name in the Philippines). A. Bixa orellana (achiote /achiote/atsuete), B. Indigofera suffruticosa (anil/añil/anyil), C. Paubrasilia echinata (sappanwood/palo brasil/palo colorado), D. Mirabilis jalapa (marvel of Peru/maravilla/a las cuatro), E. Cosmos sulphureus (yellow cosmos/chochopali/cosmos), F. Plumeria rubra (frangipani/sacalasúchil/kalatsutsi), G. Senna alata (candle bush/arbusto de la tiña/akapulko), H. Nicotiana tabacum (tobacco/tobacco/tabako), I. Arachis hypogaea (peanut/cacahuate/mani), J. Theobroma cacao (cocoa tree/cacao/kakaw), K. Carica papaya (papaya/papaya/papaya), L. Cucurbita spp. (squash/calabaza/kalabasa), M. Ananas comosus (pineapple/piña/pinya), N. Solanum tuberosum (potato/papa/patatas), O. Ipomoea batata (sweet potato/camote/kamote), P. Zea mays (corn/maiz/mais), Q. Manihot esculenta (cassava/mandioca/kamoteng kahoy), R. Annona muricata (soursop/guanbána/guyabano), S. Annona squamosa (sugar-apple/cherimoya/atis), T. Spondias purpurea (jocote/ciruela/sinigwelas), U. Sechium edule (mirliton/chayote/sayote), V. Diospyros nigra (black soapapple/zapote negro/sapote), W. Agave spp. (agave/maguey/maguey), X. Muntingia calabura (calabur tree/capulín/aratiles), Y. Capsicum spp. (chili/chile/sili), Z. Solanum lycopersicum (tomato/tomate/kamatis), AB. Manilkara zapota (sapodilla/chicosapote/tsiko), AC. Pithecellobium dulce (monkeypod/guamúchil/kamatsile), AD. Phaseolus lunatus (lima bean/frijol/patani), AF. Psidium guajava (guava/guayaba/bayabas).
The primary role of Indigenous incorporation is highlighted by the fact that one of the most successful crops arriving during this time was the sweet potato (Ipomoea batatas), a root crop that fit within the existing prevalent swidden cultivation of taro and yams. Genetic analysis has confirmed the Mesoamerican origin of sweet potatoes in the Philippines (Roullier et al., 2013), in contrast to the Portuguese introduction to Indonesia/Moluccas around the same time via the Caribbean and Africa, (Boomgaard, 2003; Clarke, 2009; Green, 2005) or the pre-Colombian introduction to Polynesia (Hather and Kirch, 1991; Ladefoged et al., 2005). Historical accounts hint that sweet potato was introduced in the Philippines by the mid-16th century CE, perhaps during the Villalobos expedition. Accounts after 1543 mention sweet potatoes, called camotes by the locals, the Nahuatl word for the crop. Loarca reported that people in central Philippines eat ‘beans, and a kind of root resembling the potatoes of Santo Domingo, called by the natives camotes’ (1582 in Blair and Robertson, 1906, vol. V: 45). de Morga (1609) recorded that people in Zambales in central Luzon ‘eat boiled camotes (which are sweet potatoes), beans, quilites and other vegetables; all kinds of bananas. . .’. Pretty in 1588 (1904: 40) mentioned that while the English privateer Thomas Cavendish was off coast of the island of Capul in a bid to intercept a galleon ship, a local cacique brought them ‘potato rootes, which they call camotas’. Pigafetta, much earlier, (1550–59) also mentioned that people in the central Philippines consume ‘camotes [batate]’ (in Blair and Robertson, 1906, vol. XXXIII: 99), although it is very likely he mistook, taro or yams as sweet potato (see Scott, 1994: 42–43). Spanish records (i.e. de Plasencia, 1589; San Buenaventura, 1613) mentioned that sweet potato, and other root crops, were grown in hillside swiddens. In Benguet Province in northern Luzon, 17th century records mentioned that such swiddens were used to cultivate root crops for 3 years before being abandoned (Newson, 2009: 231, see also Scott, 1974).
As well as facilitating the arrival New World domesticates, the Spanish did, on occasion, actively import and cultivate Old-World plants in the Philippines, wheat being a noteworthy example. Wheat was indispensable to imperialism, religion and trade. The host taken with each Mass in Catholicism, the religion the Spanish spread across the Archipelago as part of their imperial mission (Phelan, 1959a), required wheat. Likewise, wheat was the key ingredient in bizcocho, an unleavened and twice-cooked bread that was the primary ration aboard the Manila Galleons. Wheat was also the staple grain of the Spanish colonizers. Since the Philippine climate was (and is) not amenable to wheat, the Spanish imported it from China. However, when trade with China faltered in the mid-17th century, the alcalde-mayor of Laguna de Bay province, Luis de Matienzo, reported successfully cultivating wheat through indigenous labour in 1664 and 1665, much to the pleasure of clergymen and civil administrators (AGI, Filipinas 193, N. 20). Unlike sweet potatoes and other New World plants, though, wheat did not become integrated into indigenous diets. Wheat was too difficult to cultivate. Its costly, labour-intensive, limited production was only sporadically justifiable, but nonetheless could momentarily transform land-use in select provinces.
In addition to crops, livestock was introduced to the Philippine islands from New Spain. For instance, de Morga (1609) in Sucesos de las Islas Filipinas described how traditional goat farming was combined with attempted Spanish introduction of sheep to the Philippines.
‘Goats and kids are raised, although their flesh is not savory, because of the humidity of the country. These animals sicken and die for that reason, and because they eat certain poisonous herbs. Ewes and rams, although often brought from Nueva España, never multiply. Consequently there are none of these animals, for the climate and pasturage has not as yet seemed suitable for them’. (in Blair and Robertson, vol. XVI, 1907:90).
As noted above, there is also currently no concrete evidence for cattle introduction in the Philippines prior to Spanish arrival, although there are tantalizing hints. Even in the early Spanish period, archaeological evidence for the appearance of cattle remains limited. For example, habitation sites with deposits dating to the early Spanish period did not yield evidence for the presence of cattle (i.e. Bersales and de Leon, 2011; Ledesma et al., 2015; Mudar, 1997; Peterson et al., 2005). Historical sources maintained that cattle were introduced to the Philippine islands late in the 16th century, from China and Mexico (in Scott, 1994: 50) and, by the early 17th century, cattle estates had been established in many parts of the Philippines.
More important than cattle, especially for indigenous producers, was the introduction of the horse to the Spanish controlled areas of the archipelago. The horse is not native to the Philippines and its presence in the islands forms a key part of the Spanish conquest after 1565. Linguistic reconstruction points to the possible pre-Hispanic presence of horses in the Philippines, particularly in the island of Mindanao (see Scott, 1994: 278), although there is currently limited archaeological evidence to support this. Tenazas (1968), in her excavation of 15th century deposits in Pila in southern Luzon, reported the presence of horses. More notably, Locsin et al. (2008) identified horse remains (based on skull fragments and cervical vertebrae) from layers dated to 8000–6000 BCE in their excavation of Lemery Site also in southern Luzon. This early date is questionable and therefore the pre-Hispanic presence of horses remains to be elucidated by archaeological evidence. Spaniards certainly considered that they had introduced the animal as it was not present in those areas of the archipelago where they first settled. The religious orders played a key role in expanding the number of horses in the colony and already by 1604 Chirino mentions the existence of ‘great stock-farms’ in relation to horses (in Blair and Robertson vol. XII: 191). Numbers had already increased to such an extent by 1689 that William Dampier reported the animal plentiful on Luzon and feral on Mindanao (in Blair and Robertson vol. XXXIII: 87). Half a century later, horses are even mentioned in connection with the archipelago’s commerce (in Blair and Robertson vol. XLVII: 302), and adopted widely by native farmers both as beast of burden and as means of land transportation (Bankoff, 2004).
Beyond particular fauna and flora, the Spanish also introduced a land tenure system to the Philippine Archipelago that likely had significant implications for patterns and intensities of land-use in different areas (Cushner, 1973, 1976; Merchant, 2012; Phelan, 1959a; Reed, 1967; Sobritchea, 1981). Most pre-Hispanic societies practiced communal land management under the administration of a chieftain (Cushner, 1976; Merchant, 2012). The Spanish introduced ‘the notion of land ownership as opposed to land use’ (Phelan, 1959a: 117) and in doing so paved the way for the use of land for agricultural enterprises (Cushner, 1976; Merchant, 2012). Merchant (2012: 54) noted that 1571 to 1626 marked the initial growth phase of the colonial estates, with the Spanish crown granting around 200 land deeds to private individuals and church orders. The Spanish crown also granted allotments to Spanish officials (‘caballería’). The Laws of the Indies stated that such allotments were
‘composed of a ground lot of 100 x 200 feet, 799 acres for wheat or barley, 79 for corn, 63.6 for orchards and enough pasture for 50 swine, 100 cows, 20 mares, 500 sheep and 100 goats’ (Recopilación de las Leyes. . . Book IV, Title XII in Cushner, 1973: 33).
By the early 17th century the system of land estates had been established for agriculture, mostly for rice production but also tobacco and sugar, as well as ranching, mostly cattle but also horses and sheep (Cushner, 1976). Orders and records (i.e. Colin, 1663; de San Agustín, 1720) from the period noted that livestock were imported from China, Japan and Mexico. Livestock ranches were established in many parts of the country. For instance, 24 such ranches were established around Manila by 1606, some of which had more than 4000 cattle (Diaz Guiral, 1606, in Blair and Robertson, vol. XIV, pp. 156–157). In Cebu, a Jesuit estate reportedly had 14,000 cattle (Costa, 1961). However, cattle ranching appears to have been very limited and, unlike in New Spain, did not cause large scale land transformations and human population displacements (Phelan, 1959b). Treasury records show that for the years 1632–1633 and 1634–1635, cattle ranches in the archdiocese of Manila only paid 300 and 211 pesos in tithe, respectively (Manila Treasury Accounts, 1638 in Blair and Robertson, vol. XIV, pp. 81–84).
The trend for cattle ranching occurred alongside an upturn in maize farming. Maize, presumably introduced to the Philippines in the mid-16th century (see Phelan, 1959a, 1959b; Spencer, 1975), was planted in many ranches but did not replace rice as a staple. Spencer (1975: 9) argued that this was due to the fact that the milling technology for the processing of whole grains into an edible staple was not introduced by the Spanish. Instead, maize was consumed as a green vegetable. Only during the latter part of the Spanish colonial period was maize incorporated as an important part of the diet. Demographic growth on the island of Cebu during the 18th (Cullinane and Xenos, 1998: 98–99) or 19th centuries (VanderMeer, 1967: 328) may be in a large part attributable to maize farming (see also Spencer, 1975).
Beyond historical records, actual archaeological or palaeoecological evidence for subsistence, settlement and land-use changes during the first two periods of Spanish occupation of the Philippines has been limited away from the intensification of terracing systems in the Cordillera discussed in the previous section. The majority of archaeology from this time period has focussed on the construction of church buildings, as well as changes in local material culture and burial goods as a product of cultural contact (e.g. Barretto-Tesoro, 2015; Bautista and de la Torre, 1994; Bersales and De Leon, 2011; Dizon, 1994; Neri et al., 2010; Paz, 2006; Peterson, 2003). Studies of settlement patterns and archival records show that the Spanish did have an impact on settlement organisation, particularly through the practice of reducción, which forced Filipino communities away from dispersed and dendritic settlement networks and into more consolidated towns, often based around a town hall and church, potentially placing more intensive demands on certain areas of the landscape (i.e. Acabado and Barretto-Tesoro, 2020; Barretto-Tesoro, 2015). However, its widespread effectiveness and environmental consequences remain obscure. Remarkably, very little has been done to look at archaeobotanical and zooarchaeological evidence of changing plant and animal management practices, dietary reliance on different resources and landscape impacts following the introduction of New World crops and animals to the Philippine Archipelago. This is similar to the scenario in Central and South America, as well as the Caribbean, where most archaeological and palaeoecological attention on shifting human impacts on tropical environments has centred on the pre-Spanish phases of occupation, with historical research dominating the later period, with a few notable exceptions (e.g. Andrade et al., 2019; Maezumi et al., 2018). Overall, however, the current evidence suggests that the Galleon Trade did not stimulate widespread production for external trade within the Philippine Archipelago and while settlement patterns and ways of using the land changed, their overall impact on Philippine landscapes remains to be properly ascertained.
The third and final stage of the Spanish Colonial period in the Philippine Archipelago is defined by the Bourbon Reforms with end of the Galleon Trade and subsequent establishment of the Royal Philippine Company (Real Compaña de Filipinas) in 1785 which had the monopoly on tobacco, cotton, indigo, abaca, coffee and sugar plantation (e.g. Díaz-Trechuelo, 1965; Fradera, 2004; Schurz, 1959). Shortly thereafter, in 1834 the Port of Manila was opened to international commerce followed, in 1855, by other ports in the archipelago. At this point, non-Spanish Europeans were allowed to own land in the Philippines, transforming the local Philippine economy and, as a consequence, land. Huge areas were transformed into plantations to produce sugar, coffee, abaca and tobacco for export (Fradera, 2004; Legarda, 1999; Legarda y Fernandez, 1967; McCoy and de Jesus, 1982; Mehl, 2016). Prior to this period, plant cultivation in the Philippines was limited to subsistence agriculture (e.g. Cushner, 1971; Rafael, 1988; Skowronek, 1998) and small land estates (Cushner, 1976). A landmark example is the large-scale transformation of land in the island of Negros in the Central Philippines to sugar cane plantations. Precipitated by the opening of the nearby port of Iloilo in 1855, the island became virtually a monoculture of sugarcane (see Aguilar, 2017; Quirino, 1974). Detailed archival research is needed to estimate how much land was converted to plantations but sugarcane exports in the port of Iloilo overtook that of the main port in Manila by 1873. In 1885, the Port of Iloilo exported more than 105,000 metric ton of sugar compared to around 65,000 metric ton exported from the port of Manila and ca. 27,000 metric ton from the nearby port of Cebu (Foreman, 1899: 295–296). Similarly, Bankoff (2013) has noted that prior to the 19th century, both pre-Spanish and Spanish logging was selective for particular tasks (e.g. ship building) and often culturally mediated. However, from the mid- to late-19th century commercial logging enterprises expanded, including less-desirable hardwoods. By the time American imperial rule started, the new administrators were disappointed by the lack of availability of high-quality lumber for sale on global markets in Luzon and the Visayas, though large areas of forest remained in Mindanao and Palawan (Bankoff, 2007, 2013).
Discussion
The Philippine Archipelago is an often-neglected but critical region for studying how past movements of plants, animals and ways of using the land have impacted tropical environments. This is especially the case as archaeologists, palaeoecologists and earth systems scientists posit the major role changes in tropical land-use, among pre- and post-colonial societies, might have had on the climate and even the atmosphere on regional and global scales (Fuller et al., 2011a; Lewis and Maslin, 2015). The Philippines is one of the few land-masses hosting endemic Island Southeast Asian biodiversity to experience both major prehistoric and historic changes in subsistence activities and land-use stimulated by trade resulting from integration within a pan-tropical European Empire. However, as Figure 1 shows, most work has been focussed in northern Luzon and the central Philippine Islands with limited research done in other areas to date. There is also a certain temporal bias in archaeobotanical and zooarchaeological analyses, and much work has focussed on Neolithic, and some Metal Age, contexts, even despite the problems there have been with finding occupation sties. Archaeobotanical and zooarchaeological research has been notably lacking for protohistory and the Spanish period, at least outside major urban centres such as Manila and Cebu. There has also been a general absence of the direct radiocarbon dating of animals and plants, using novel pre-treatment methods, from all time periods, making it difficult to build detailed reconstructions of introductions and subsistence changes in different parts of the Archipelago beyond the ‘Three Age System’ used here. Nor have previous historical studies included a detailed analysis of the rich archival sources on land cover, agriculture and livestock to be found in the archives of the Philippines, Spain and Mexico. Finally, stable isotope analysis to look at changing dietary reliance on incoming plants and animals is yet to be applied in the region.
This, alongside a general lack of palaeoecological records, has meant that reconstructing the exact impacts of novel translocations on subsistence, land-use and environments is currently challenging. Nevertheless, our review reveals some interesting patterns worthy of, albeit broad, comparison to other tropical areas that have been hypothesised as witnessing major changes in pre- and post-colonial land-use. Firstly, pre-colonial translocations into the Philippines in the Neolithic and Metal Ages were apparently fit within Indigenous swidden cultivation focussed on root crops and hunting, gathering and fishing This pattern perhaps has some similarities with the arrival of maize in the Amazon Basin as it was incorporated within a mixed subsistence strategy that relied upon wild forest resources, freshwater animals and ‘garden’ plots of indigenous tubers such as manioc (Heckenberger and Neves, 2009; Hermenegildo et al., 2017; Roosevelt, 1999) – although this region has a deep, growing record of significant human landscape and soil modifications that is thus far absent from the Philippines (e.g. de Souza et al., 2018; Lombardo et al., 2020). In the case of the Caribbean islands, although a relatively late arrival of humans, especially when compared to the Philippine Archipelago, has been argued to have perhaps had dramatic impacts on ecosystems as a product of megafaunal hunting and forest clearance (Cooke et al., 2016). However, more detailed palaeoecological research suggests that Archaic inhabitants of the Caribbean combined the introduction of novel domesticates, such as maize, sweet potato and chili peppers, with the promotion of endemic economically useful wild plants such as wild avocado and marine resources (Pagán-Jiménez et al., 2015; Rivera-Collazo, 2015), in a way not too dissimilar to Neolithic and Metal Age Philippine communities. While these forms of land-use may have had a shifting impact on tropical forest environments, and the addition of rice, pigs, buffaloes and commensals such as rats may also have had ramifications for biodiversity and soil erosion, there is certainly not the same dramatic land-use change argued by Fuller et al. (2011b) for the spread of rice and water buffaloes across mainland Southeast Asia which they argued may even have impacted global methane emissions.
By the end of the Metal Age period, and particularly during the Protohistoric period, there is a clear intensification of rice agriculture and the keeping of domesticated animals such as goats and water buffaloes, as chiefdoms and more urbanised settlement networks grew in certain areas of the Philippines. This is recorded in archaeological and palaeoecological records of terracing, notably in certain parts of Luzon, and in historical records made by the Portuguese and Spanish upon arrival in the region. There may even have been a pre-colonial arrival of cattle and horses, though this remains tentative. There is clearly an expansion of more permanent swidden systems for the growth of dry rice, as well as more extensive terrace systems in upland areas for either the growth of wet rice or root crops. Water buffalo also seems to have been integrated within these field systems as a form of traction. This would likely have begun to have more significant impacts on forest clearance, as documented from one of the few palaeoecological records available from the Cagayan River Valley (Stevenson et al., 2010), although it also seems that rice and water buffalo remained ‘elite’ food sources, perhaps limiting the scale of these impacts. Indeed, unlike mainland Southeast Asia, the Amazon Basin and North and Central America, there is no clear evidence for extensive urban settlements and large populations across the Philippines prior to the 15th century AD, despite the growth of more urban nodes, such as Butuan and Manila, as part of expanding maritime Southeast Asian networks. The pattern is perhaps more akin to the growth of ‘Ceramic-age’ chiefdoms of the Caribbean, which began to have more widespread impacts on biodiversity and soils in this region (e.g. Fitzpatrick and Keegan, 2007). Nevertheless, there is no clear evidence for changing settlement patterns or field systems having a widespread impact on pre-colonial Philippine environments.
The Philippines certainly potentially provides an important counterpoint to the Neotropics in the context of demographic change following the arrival of Europeans in the region. While the movement of smallpox, measles and the bubonic plague as part of the ‘Columbian Exchange’ across the Pacific may have impacted indigenous Filipino communities, alongside abuses of Spanish conquerors and clergy (De Bevoise, 1995; Newson, 2006, 2009), there does not seem to have been anywhere near the 90% indigenous mortality recorded across the Neotropics (e.g. Koch et al., 2019). Unlike in the Neotropics, there were no big cities and relatively few urban centres that would have hastened the spread of diseases. Likewise, protohistoric communities in the Philippines most likely had some level of acquired immunity thanks to their increasingly close proximity to domestic animals. This likely shaped the initial uptake of novel plants and animals brought by the Spanish, as well as the effectiveness of Spanish governance across what was to become the centre of the Spanish East Indies. Sweet potato, the most effective early arrival, fit neatly within indigenous swidden cultivation practices, meanwhile the horse was utilised as a means of transportation to move goods to market and people between communities (Bankoff, 2011). Widespread pastoralism only occurred in selected regions and, even then, mass ranching introduced by the Spanish only really expanded in the 19th century. The Spanish practice of reducción may, as in the Neotropics, have had impacts on how land-use was dispersed across the Philippines, though the overall application of this approach across the Archipelago seems to have been less in comparison to other regions of the pan-tropical empire (Acabado and Barretto-Tesoro, 2020; Barretto-Tesoro, 2015). Indeed, difficulty of extending control, even across Luzon, particularly between the 16th and early 19th centuries, is highlighted in the remaining independence of populations in the Cordillera and eastern Luzon (Acabado, 2015, 2017). In the case of the former, more intensive terracing also occurred, but primarily in response and resistance to Spanish imperial forces. It was only from the mid-19th century, that certain provinces specialized in certain land-use and logging, ranching and other forms of plantation use took on a more commercial element with corresponding impacts on the scale and nature of deforestation.
This resistance to Iberian rule perhaps has some similarities to the majority of the Amazon Basin that only began to feel direct Portuguese rule from the early 19th century, though even there the widespread arrival of European diseases decimated Indigenous populations and their land-use practices, as they did across the Caribbean, North and Central America and the high Andes. Although there are many local exceptions (Alexander and Álvarez, 2018), across much of these areas the Spanish rapidly sought to expand plantation agriculture, pastoralism and particularly European models of towns and cities (Dumire, 2004; Melville, 1994; Miller, 2007), either by co-opting existing prominent capitals, such as Tenochtitlan, or by building their own new centres (Miller, 2007). It has been estimated that by 1600, nearly half of the population lived in cities across the Spanish-occupied portions of the Neotropics (Miller, 2007), something not seen in the Philippine Archipelago until the latter half of the 19th century. Furthermore, unlike North, Central and South America, the Philippines proved to have relatively few rich ores for mining, limiting, at least after initial investigation, disastrous deforestation, massive population relocation and mortality that occurred around such extractive enterprises in the Neotropics (Brown, 2012). In fact, one of the most evident changes in land-use in the Philippines following Spanish arrival was the fort and shipbuilding enterprises that highlighted the tenuous imperial grip on much of the Archipelago. By the mid-19th century, however, following the Bourbon reforms of the Spanish Empire and the arrival of commercial interests, the same extensive land-use changes as a result of tropical plantation agriculture and the beginnings of commercial lumber production, began to impact the Philippines as they did much of the former Spanish Empire, likely leaving a legacy on the forest cover, landscape stability and perhaps even the climate of the Philippine Archipelago to this day (Bankoff, 2007, 2013).
Although these contrasts and patterns are interesting, for the most part they remain to be definitively proven, at least in the context of widespread pre- and post-colonial land-use changes across the Philippine Archipelago. Elsewhere in the tropics, the incorporation of archaeologically and palaeoecologically informed land-use changes into climate models has been argued to demonstrate that both pre- and post-colonial communities could have significant impacts on tropical forest environments, change precipitation patterns and even global emissions (e.g. Cook et al., 2012; Fuller et al., 2011a; Koch et al., 2019; Lewis and Maslin, 2015; Stephens et al., 2019). Meanwhile, on more local or regional scales, changes in forest burning and the introduction of different plants and animals, has been shown to have drastically altered forest composition, vulnerability to catastrophic forest fires and plant and animal communities on prehistoric and historic timescales (Fitzpatrick and Keegan, 2007; Levis et al., 2018; Maezumi et al., 2018; Roberts et al., 2017). Such work has been crucially informed by the application of multidisciplinary palaeoecological coring of lake and geomorphological sequences (Maezumi et al., 2018), LiDAR remote sensing to map settlement patterns and extents through areas with dense vegetation cover (Iriarte et al., 2020), and more thorough estimates of dietary reliance based on zooarchaeology, archaeobotany and biochemical analyses. This has been lacking from the Philippines and, alongside renewed archival work and archaeological survey of settlement sites, has much promise to develop and test some of the hypotheses outlined above in relation to the impacts of translocations and land-use shifts in the last 6000 years of the region’s history. For the time being, however, we would suggest that the Philippine record should temper assumptions of the sweeping effects of the ‘Neolithic’, the arrival of metal, and even colonialism and the ‘Columbian Exchange’ on tropical landscapes that have been hypothesised, and in some cases shown, elsewhere (i.e. Bain et al., 2018; Castilla-Beltrán et al., 2018; Scarborough et al., 2019).
Conclusion
The Philippine Archipelago is a key point of comparison for other tropical regions where significant plant and animal introductions have been argued to have had significant impacts on deforestation, soil erosion and regional climate over the course of the Holocene. Not only did it experience the three major changes in Southeast Asian land-use in prehistory, including the arrival of rice agriculture, domesticated livestock and metal technology, but it also experienced subsequent Spanish colonisation from the 16th century CE onwards. Nevertheless, the Philippines has been largely neglected in discussions of pre- and post- tropical land-use and translocations in favour of the Atlantic sphere of the Neotropics. Our review demonstrates that archaeological investigations conducted in the Philippines provide important information on the timing of the introduction of various domestic animals and crops into the Philippines, as well as shifts in land tenure and organisation. These introductions potentially represented major turning points in land-use, endemic biodiversity and environmental change although, currently, evidence for significant human-associated environmental degradation is largely limited to the late period of Spanish colonisation in contrast to many other areas of Spanish conquest in the tropics. We argue that more multidisciplinary research, including detailed palaeoecological reconstruction and more regular archaeobotanical and zooarchaeological investigation, as well as renewed archival appreciation, are essential in order to gain insights into how extreme and widespread the impacts of translocations and hypothesized land-use organisational changes were in the Philippines, and whether they have left legacies lasting into 21st century deforestation, soil erosion and threatened biodiversity.
Footnotes
Acknowledgements
The authors would also like to thank three anonymous reviewers whose comments and recommendations have greatly improved the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This project is funded by the European Research Council Starter Grant PANTROPOCENE (no. 850709) awarded to P.R., P.R. and N.A. would also like to thank the Max Planck Society for financial support.