
Abstract
Like any other living being, humans constantly influence their environment, be it intentionally or unintentionally. By extracting natural resources, they shape their environment and also that of plants and other animals. A great difference setting people apart from all other living beings is the ability to construct and develop their own niche intentionally, and the unique tool for this is cultural behaviour. Here, we discuss anthropogenic environmental changes of hunter-gatherers and present new palaeoecological and palynological data. The studies are framed with ethnoarchaeological data from Western Siberia to gain a better understanding of how different triggers lead to coping mechanisms. For archaeological implication, we use two Mesolithic case studies from Germany: One of them focuses on hazelnut economy around ancient Lake Duvensee, and the other broaches the issue of selective roe deer hunt and its consequences at the site of Friesack. We address the archaeological evidence from the perspective of active alteration and its consequences, starting our argumentation from a perspective of niche construction theory. This approach has rarely been applied to early Holocene hunter-gatherers in Northern Europe even though the available data render possible to discuss human–environment interaction from such a perspective. It is demonstrated that archaeological research has tools at hand that enables to detect anthropogenic niche construction. However, the ethnoarchaeological example shows limitations and archaeologically invisible triggers and consequent results of human adaptations. The critical revision of such perspectives based on empirical data provides a better understanding of social and environmental transformations in the early- and mid-Holocene.
Keywords
Introduction
Through their behaviour, humans have been constantly influencing their environment even before the dawn of agriculture and livestock breeding. But this impact depended on many different factors such as season or habitat as well as sizes and intentions of the human groups. For the Mesolithic, the entanglement of long-lived cultural systems and man-made environmental alterations has been proven in several studies that exemplify hunting strategies which targeted only selected age classes or the manipulation of wild plant resources to increase their productivity and potentially also to attract game for hunting (e.g. Dietze et al., 2018; Innes and Blackford, 2003; Johnson and Miyanishi, 2012; Magnell, 2005; Mertens, 2000; Rowley-Conwy and Layton, 2011; Ryan and Blackford, 2010; Simmons and Innes, 1985, 1987; Zvelebil, 2003; but see the argumentation in Brown, 1997). These manipulations had different degrees consequences for the ecological, economic and also social spheres, that is, by creating habitats for particular plant or animal species or by the development of (semi-)sedentary lifestyles.
Increasingly, it has become a focus in archaeology to understand the discussed processes as functions of human–non-human cohabitation and coevolution beyond the culture–nature dualism rooted in Western scientific traditions and deciphering the associated economic, ecologic and social practices (cf. Bird and O’Connell, 2006; Brittain and Overton, 2013; Codding et al., 2016; Fitzhugh et al., 2018; Pilaar Birch, 2018; Rowley-Conwy and Layton, 2011; Stewart et al., 2004; Wieckowska-Lüth et al., 2018; Zeder, 2015). As many hunter-gatherer practices are embedded within a culturally and spiritually charged environment, they can at best be partially grasped by archaeological methods (e.g. Arnold, 1996; Brinch Petersen, 2012; Grøn, 2012; Jordan, 2003; Schmölcke et al., 2017). In addition, changes and transformations in the status of organisms or non-biotic parts of the landscape can only be empirically traced if data can be addressed from a sequential perspective, that is, with samples or observations that reflect different but related points in time. While much archaeological data rather reflect accumulated situations with no internal spatial stratification, the detection of transformation processes or the creation of new ecological niches can often only be discerned in the medium term. However, as we will show in the following, given archaeological records do provide the possibility to detect active alteration of the environment under specific circumstances. While many aspects of niche construction in the past are difficult to detect with conventional archaeological methods, ecological, ethnological and ethnoarchaeological observations can give impressions of possible developments and processes which may have played a role during prehistory.
Approach
To discuss possibilities and difficulties for finding intentionally and unintentionally modified environments of hunter-gatherers, we provide examples of how these can be investigated and which challenges are present with respect to the topic in focus. We address the archaeological evidence from the perspective of active alteration and its consequences, which is based on the concept of niche construction theory (Odling-Smee et al., 2003). In archaeology, niche construction theory is of utility because it allows to quantify and incorporate human activities as active variables in environmental change and cultural development (Laland and O’Brien, 2010). It has widely been used in archaeology (e.g. Banks et al., 2006; Benito et al., 2017; Boivin et al., 2016; d’Errico et al., 2017; Gerbault et al., 2011; Riede, 2011; Rowley-Conwy and Layton, 2011; Slim, 2016; Smith, 2013, 2015; Weninger, 2017); however, so far, it has rarely been applied to early Holocene hunter-gatherers in Northern Europe (cf. Wieckowska-Lüth et al., 2018). Our objective is to provide examples of empirical studies and their application, rather than discussing niche construction theory for the sites under consideration in its entirety. Hence, we provide data-driven applications of it to exemplify research designs for this approach.
We aim for combining ethnoarchaeological and palaeoecological perspectives to widen the theoretical and empirical basis for understanding processes of transformations in the past. Archaeological and palaeoecological data will be evaluated for indications of niche construction in the Mesolithic, followed by a critical review of the possibilities and limitations for detecting anthropogenic niche construction in archaeological data. As will be shown, archaeological sciences have several tools and opportunities for discussing such processes.
In this study, we use palynology for testing the hypotheses of human improvement of growth and productivity of hazel (Corylus avellana) which has been a commonly used staple during the Mesolithic in Europe. A zooarchaeological perspective will shed light upon hunting pressure and prey selection at the site of Friesack where selective hunt shaped the composition of the deer populations. Eventually, ethnoarchaeological evidence is used as a basis for discussing the archaeological visibility of various types of interaction with the environment in hunter-gatherer communities. Introducing such a perspective underlines that niche construction mechanisms can involve agents and interrelations between environmental, economic and social spheres that are difficult to be detected by current archaeological methodologies. Respectively, we evaluate archaeological and palaeoecological data for preserved evidence of anthropogenic environmental transformations in the past and also critically review the possibilities and limitations of this approach against a background of ethnoarchaeological studies.
Active alterations by hunter-gatherer communities in temperate environments: Archaeological and ethnoarchaeological case studies
Following the theory, niche construction is an omnipresent phenomenon in ecosystems and a basic process of evolution. However, it is not unpretentious to detect in the past, be it caused by biotic or abiotic agents. For archaeological visibility, at least one of the following two prerequisites must be met: first, a well-dated archaeological site or microregion with several specifiable occupation phases and preservation of different organic find groups to reconstruct developments in the succession of vegetation and fauna as well as transformations in human exploitation methods. Such a case is given with the Mesolithic site Friesack in Northern Germany (Gramsch, 2000; Groß, 2017). Second, a microregion with several precisely dated sites of short-time occupations. Thereby, relationships and interplays between human and environment can be investigated, given again that data from different palaeoecological disciplines are available. This exceptional case is available at ancient Lake Duvensee in Northern Germany with its numerous early Mesolithic sites (Bokelmann, 2012; Groß et al., 2018). In order to reach a better understanding of human–environment interaction in hunter-gatherer communities of temperate regions in general, we will analyse archaeological evidence from these two regions on the Northern European Lowlands. In addition, these prehistoric studies are accompanied by and linked with ethnoarchaeological information from modern hunter-gatherer communities in Western Siberia. Today, this region reflects to some degree the ecological conditions (e.g. climate, vegetation and fauna) of the early Mesolithic in Northern Germany and also the people’s similar forager lifestyle make them predestined for comparisons.
Archaeological case studies
Due to the characteristic of the record, which is by far patchier than the ethnoarchaeological data presented below, the archaeological case studies are subjected to circumstantial evidences. Today, both regions under consideration are situated in extensive fen landscapes and thus provide good preservation of organic remains and accordingly rich information on prehistoric ecology. The main difference between both regions is their specific economic foci: most of the Duvensee sites were used during the early Mesolithic as short-term or specialized camps, where at some sites, especially during the Boreal, hazelnut exploitation and roasting was one of the primary activities (Bokelmann, 2012; Groß et al., 2018). In contrast, the contemporaneous Friesack sites (Gramsch, 2000, 2016; Groß, 2017) were seasonal camps which had been repeatedly occupied over longer periods. Here, hazel was present as well (Wolters, 2016) but hunting played the major role for human subsistence. The differences between the two selected study regions highlight the diverse appearance of activity areas within the early Holocene landscape and provide concrete insights into the variability in usage, exploitation and shaping of environments during that period.
Ancient Lake Duvensee
During the first millennia of the Holocene, the present-day fen Duvenseer Moor in Northern Germany was a lake which western shore was frequently visited by Mesolithic foragers. During the last decades, many of their campsites have been excavated and most of them date between the late Preboreal and the early Atlantic (see Groß et al., 2018 for a summary). The sites were located on small islets close to the former edge of the lake (Bokelmann, 2012). With regard to the topic of the present paper, it is significant that the Duvensee campsites can be connected to short-term or specialized events, often in connection with hazelnut exploitation and roasting (Bokelmann, 2012; Groß et al., 2018). Particularly, the sheer quantity of hazelnut macro-remains at some sites and high pollen percentages in local sedimentary sequences appear to suggest that hazel was an important component of the landscape. Hence, it is reasonable to explore the idea that hazel presence at Duvensee might have been favoured by direct human intervention. In the present paper, we address this hypothesis by evaluating whether the Duvensee region is in fact anomalous compared to other sites.
To interpret the activities, it is fundamental to know the former topography and consequently the exact location and extension of the occupied islets. Therefore, based on ground penetrating radar (GPR) measurements, the palaeotopography was reconstructed (Corradini et al., in preparation; Figure 1a). The stratigraphy of the basin was documented through a series of cores along a transect and then compared with GPR measurements, showing how the latter consistently succeeds in identifying a transition between sand and more markedly clay layers (Figure 1b). We associate this transition with the end of local glacial and fluvioglacial activities, and with the onset of lacustrine sedimentation. This stratigraphic discontinuity was then followed throughout the whole GPR-mapped area and interpolated between the profiles in order to create a palaeotopographic map (Figure 2).
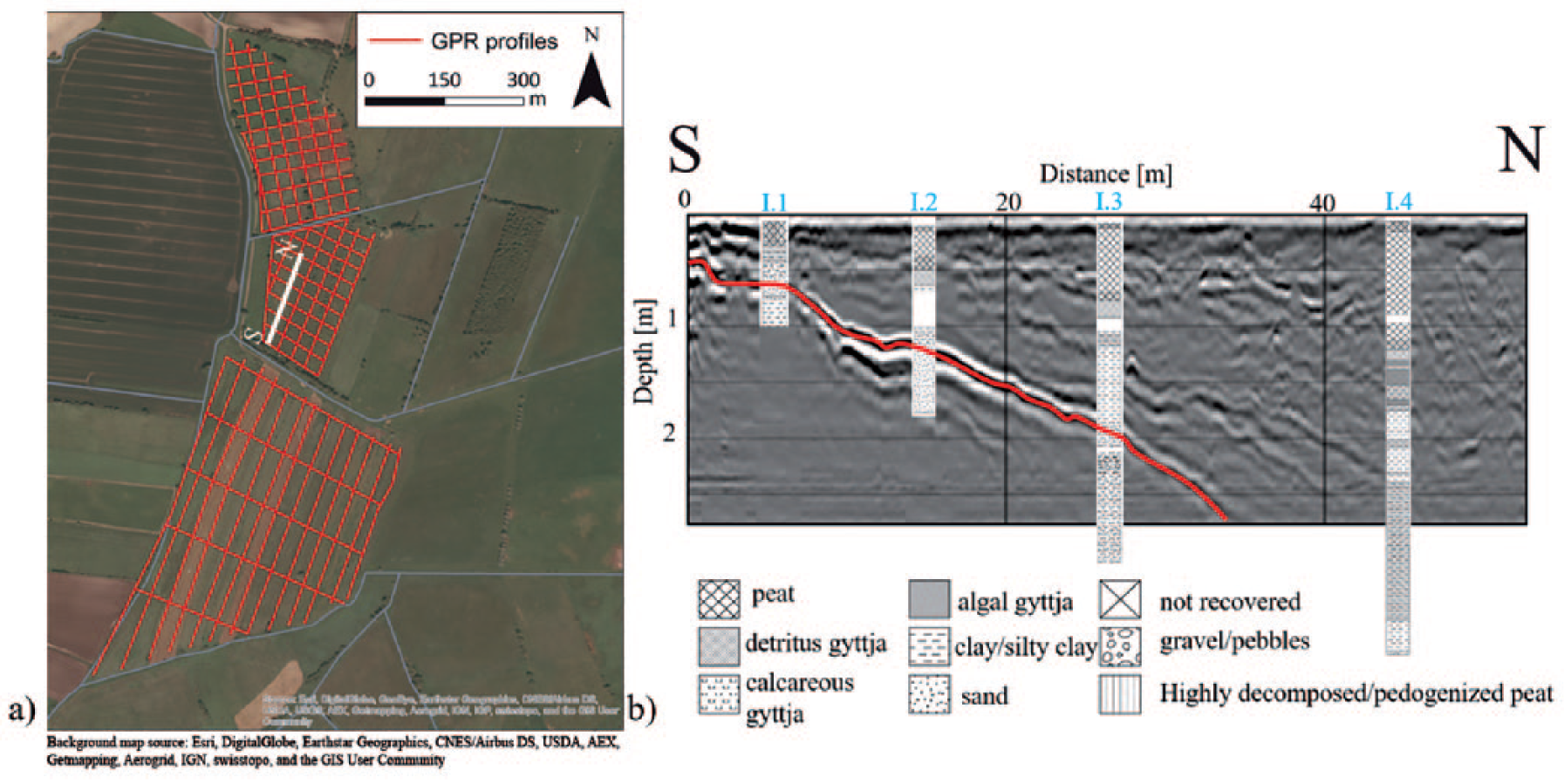
(a) The investigated area at Duvensee with red lines indicating the ground penetrating radar (GPR) profiles surveyed. (b) Comparison between a radargram and the stratigraphy from the corings I.1, I.2, I.3 and I.4. The red line indicates the reflection coming from the bottom of the lake visible in the cores with the transition between clay and sand. We used a GSSI GPR antenna with 200 MHz frequency on a grid of 30 m in the northern part of investigated area and a profile spacing of 30 m (WE direction) and 100 m (NS direction) in the southern part of the site. For the location of the site, see Figure 3.
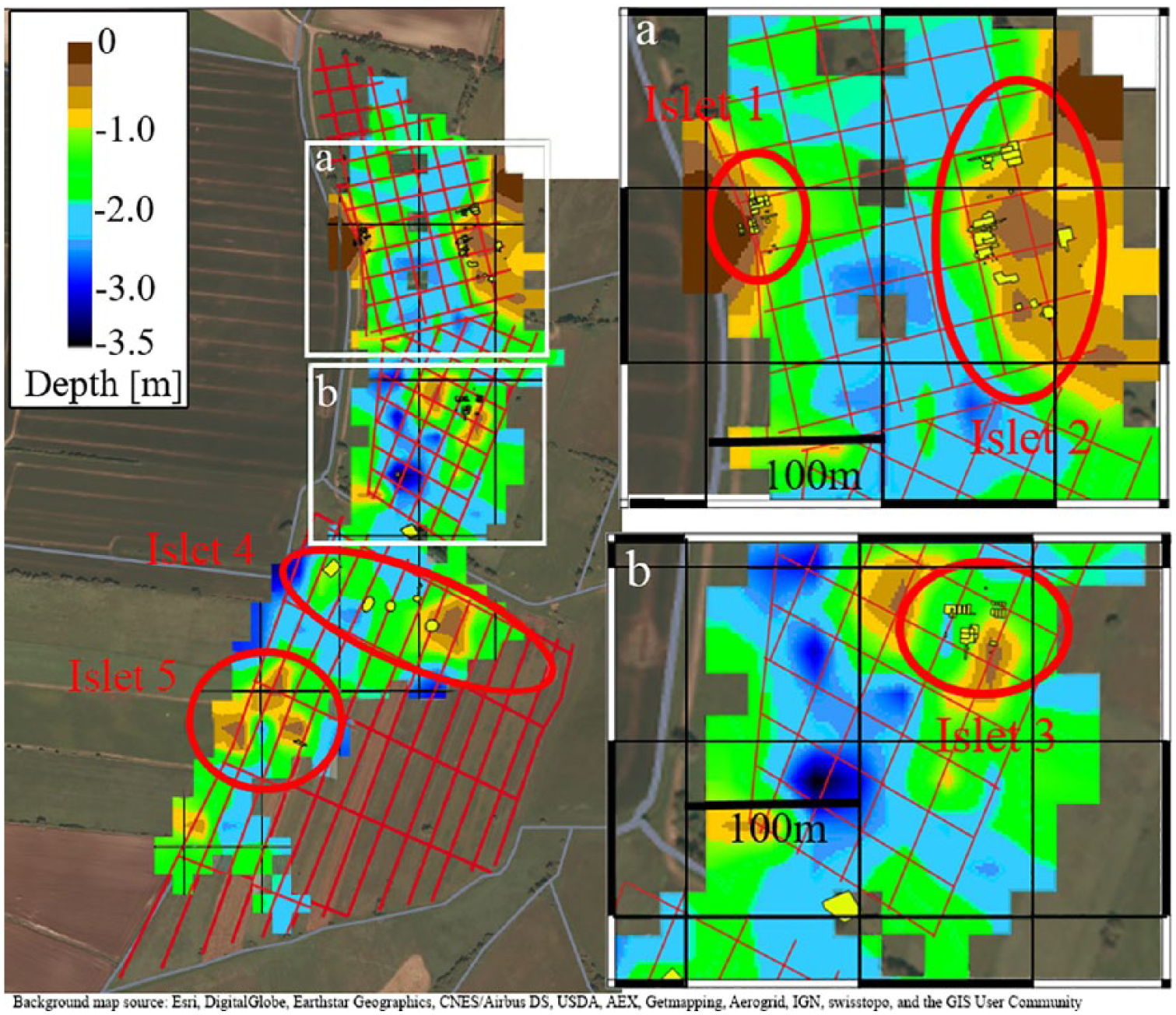
The brown areas show where the bottom of the lake is closer to the surface in contraposition to the blue areas which represent lower areas. The Mesolithic sites (yellow polygons) are located in the higher areas (islets 1–5 after Bokelmann, 2012). The conversion from time to depth has been done by calculating an average velocity of 5.5 cm/ns from the diffraction hyperbolas in the radargrams of the entire survey.
On islet 1, large numbers of hazelnuts have been found at a late Preboreal campsite. However, most sites date slightly younger and provide Boreal ages, especially those on islet 2 (Bokelmann, 2012). A three-dimensional (3D) reconstruction of the lake bottom (Figure 3) shows that islets 1 and 2 were probably the first that emerged after a drop in lake level during the Preboreal. With the regression of the water, the other islets were exposed and thus provided more potential settlement areas as well as potential stands for hazel bushes and trees.
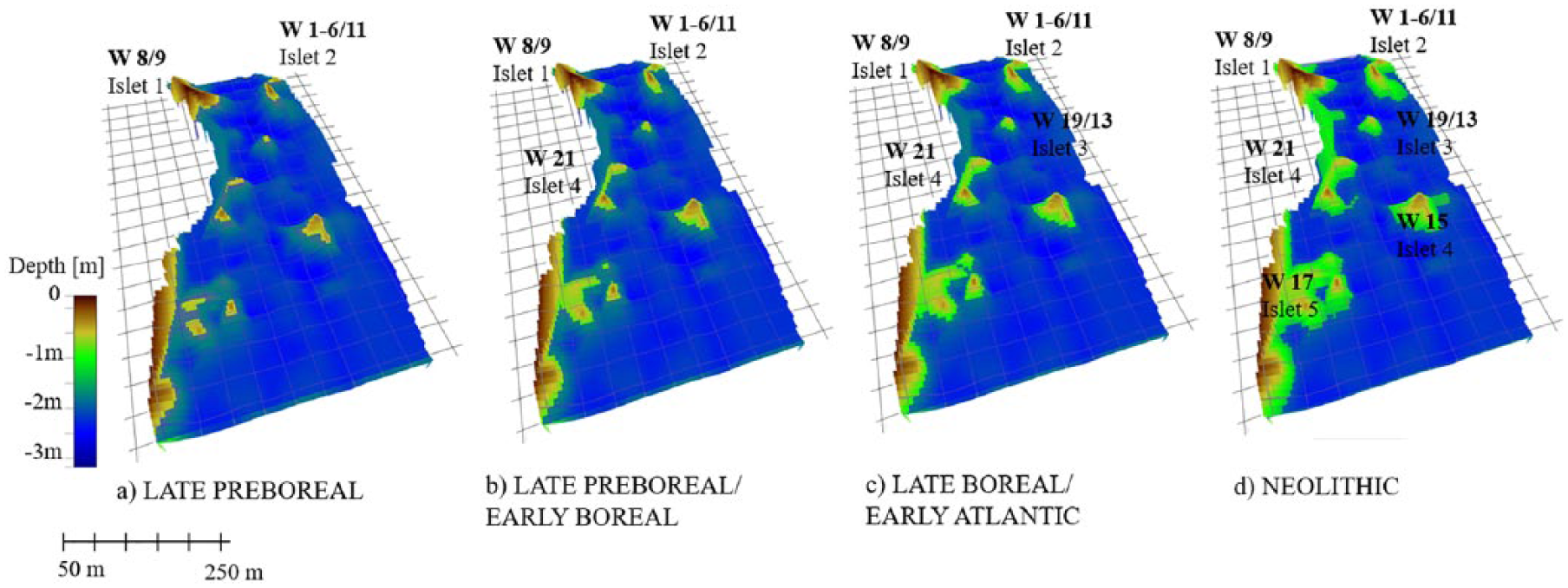
3D reconstruction of the Duvensee area (cf. Figure 2). The sites are indicated by W and the respective site number (cf. Groß et al., 2018).
Early Holocene hazel (Corylus avellana) expansion and exploitation
Hazel (Corylus avellana) is a fast-growing shrub, which prefers nutrient-rich and loamy soils and thrives along forest edges. When growing as an understorey component, it is favoured by an open canopy, as excessive shaded conditions will impede its flowering (Vera, 2000). In prehistory, it has been exploited for a variety of purposes. Its wood and especially shoots are well known to have been used in crafting (e.g. for fish weirs or handles), and the energy-rich nuts provided a good and durable staple. The pathways of the extraordinarily rapid expansion of hazel after the last Glacial have been discussed controversially: while Hagerup (1942) accounted European nuthatches (Sitta europaea) as important components of the process, Firbas (1949) saw more potential in fluvial or even anthropogenic distribution. A compilation by Giesecke et al. (2011) shows that the rapid expansion of hazel pollen in the early Holocene took place to a great degree contemporaneously over large parts of Europe. Pre-existing and widespread Corylus communities, whose flowering had been previously suppressed under unfavourable climatic conditions, likely played a role in this seemingly synchronous expansion (Tallantire, 2002).
The high pollen percentages of hazel during the early Holocene are generally interpreted as representing large hazel forests (Firbas, 1949), or as an established presence of this shrub along forest edges and in park-like landscapes (Vera, 2000). It has also been discussed whether such hazel presence could be the result of human cultivation (Bishop et al., 2015; Holst, 2010; Smith, 1970). A partial relevance of hazelnuts in the subsistence of early Mesolithic people has been proven (e.g. Bishop et al., 2015; Bokelmann, 2012; Holst, 2010; Mithen et al., 2001; Regnell, 2012). However, it is yet unclear to what extent hazelnuts played a role for nutrition as their relevance for subsistence is often overestimated (cf. Leesch et al., 2017).
Hazel bushes are resistant to pruning. This way the bush is encouraged to produce shoots or suckers which have been used for fish weirs in the younger Mesolithic (Bishop et al., 2015; Klooß, 2014, 2015; Pedersen, 1997). Since the crop yield is also improved by such processes (Regnell, 2012), managing the bushes can work in favour of food production.
Evaluating the promotion of hazel at ancient Lake Duvensee
The continent-wide expansion of Corylus in pollen diagrams might primarily depend on environmental factors, yet changes in the local abundance of this species might still potentially reflect an intentional human intervention. If the expansion of Corylus was supported by humans, they would have had to influence its natural reproductive capabilities, for example, by burning or removing other species competing for the same ecological space, or by promoting plant renewal through pruning. Yet, before evaluating the role of Mesolithic people in actively supporting the expansion of Corylus, it is necessary to reasonably exclude the influence of other factors independent from human intervention.
Given the visible differences in the abundance of hazelnuts finds between Duvensee and Friesack, we focus on the broad area surrounding these two Mesolithic sites (Figure 4). Pollen data from the study region are used to assess any qualitative differences in the sub-regional distribution of Corylus.
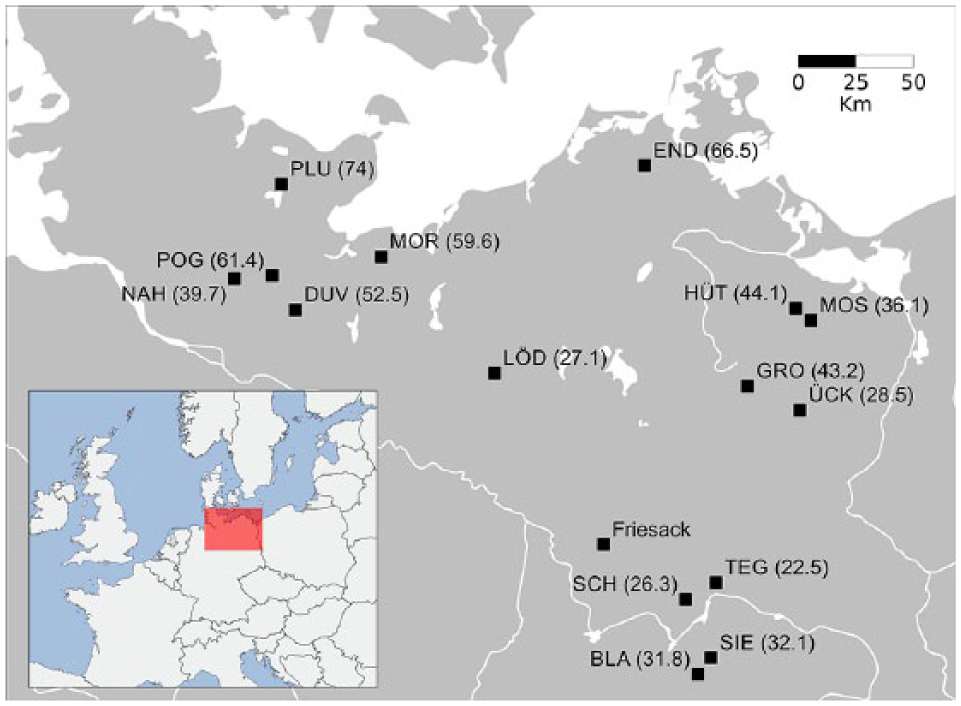
Map of the study area displaying the locations of the sites involved in the pollen–soil comparison. The numbers in brackets show the average Corylus percentage for each site (mean of three samples with the highest Corylus values; Figure S1, Supplementary Material). The position of Friesack is added for contextualization. The complete list of the sites and abbreviations is presented in Table 1.
Materials and method
The pollen data analysed in this study have been extracted from the European Pollen Database (EPD; europeanpollendatabase.net; version used: EPD_Postgres 2017-10-31). Among all the pollen archives available within the study area, we selected only those where the early Holocene Corylus rise and maximum expansion was clearly recognizable, that is, where the Corylus pollen reaches its highest percentage values at the expenses of birch (Betula) and pine (Pinus) and subsequently declines concurrently with a growing presence of alder (Alnus). Alnus was selected as a biostratigraphic marker due to its fairly synchronous expansion across Northwestern Europe (Giesecke et al., 2011). Due to a limited availability of sites in the surroundings of Duvensee, we integrate data from unpublished records (Nahe, Poggensee; see Figure S1, Supplementary Material). The Duvensee data have been partly discussed in previous publications (Averdieck, 1986; Bokelmann, 2012). The calculation of pollen percentages is based on the sum of all terrestrial species. We opted to use the average of the three highest samples (position shown in Figure S1) for the early Holocene at every site in order to mitigate the influence of possible isolated spikes.
Pollen load and fruit production depend on a series on environmental factors including interspecies competition, landscape openness, climate, soil quality, fauna grazing habits and human management (e.g. Baldwin, 2008). Any of the comparisons and statistical analyses presented within this case study are based on the assumption that the peak percentage values reached by Corylus in each pollen diagram are a direct result of the maximum flowering capabilities (and thus, arguably, hazelnut yield) locally achievable by this species. Of all the variables that may affect presence, spatial coverage and nut yield of Corylus, we focus on soil composition since high-resolution data for the study area are readily available (BÜK200, 2015). Soil data for each pollen archive were extracted by placing a circular search window around each site. The radius of the search window was limited to 25 km, since larger values would have exceeded the area covered by the soil map of Germany. The data extracted from each circular search window were divided into two main levels: soil type and parent material. The information contained within each level was then simplified into a limited number of variables for comparative purposes (Table S1, Supplementary Material). Recently developed soils (raw soils) and soils of anthropic origin (anthrosols) were excluded from further analysis. Pearson’s correlation coefficient (r) was used to measure the linear correlation between Corylus percentages and soil variables. The aim of this test is to isolate a single environmental variable and evaluate to which extent it can explain the different pollen percentages observed across the study area.
Results and discussion
A visual comparison between pollen percentages reveals that sites in the larger Duvensee area do reach higher maximum Corylus values than most other sites in the study region, arguably pointing to a wider Corylus coverage and/or to more intense flowering (Figure 4). Nonetheless, this simplistic comparison does not take into account the different distribution and pollen productivity of other dominant species, primarily Pinus. Sites with relatively low Corylus percentages (<40%) are characterized by higher Pinus values in comparison with sites where Corylus is more abundant (>40%; Figure S1, Supplementary Material). This situation reflects indeed a more widespread presence of Pinus, yet the generally lower pollen productivity of Corylus compared to Pinus might also dampen its relative proportion in the pollen diagrams. Assuming that differential pollen productivity does not invalidate a comparison of Corylus percentages across multiple sites, we focus our analysis on comparing pollen and soil data.
As changes in vegetation cover, climate and groundwater distribution undoubtedly have affected soil formation processes during the Holocene, any comparison between soil types and pollen data should be interpreted with the due caution. In addition, time itself plays an important role in soil formation (Jenny, 1980). Because of the humidity-driven leaching in Northern Central Europe, natural soils develop towards so-called climax stages (luvisols on loamy sites and podzols on sandy sites). Thus, since our study focuses on early Holocene landscapes, the recent soil cover does not reveal the Mesolithic conditions but merely shows the differences in soil types. More solid conclusions can be drawn from parent material data, since it can be assumed that their general texture properties have remained fairly constant since the early Holocene. While there are processes that result in changed contents of the finer soil components (i.e. clay mineral formation, addition of fine material by aeolian processes), the absolute amount of such changes is small (Dreibrodt et al., 2013).
The results of the soil versus pollen comparison are summarized in Table S1, Supplementary Material. Most notably, this comparison reveals a strong negative correlation between coarse parent materials and pollen data (r = −0.85, p = 0.0001) and a comparably strong positive correlation with fine parent materials (r = 0.76, p = 0.001).
Concerning soil types, a positive correlation exists between pollen and stagnosols (r = 0.63), while a negative correlation of similar magnitude is registered for poor cambisols (r = −0.64). Yet, neither of these two comparisons (stagnosols and cambisols) are statistically significant after Bonferroni correction. Regardless of significance, these results appear to successfully replicate the conclusions of early Holocene land cover models based on comparable data sets, which reconstruct abundant Corylus populations on stagnosols and a near-absence on poor sandy soils (Theuerkauf et al., 2014) (Table 1).
List of sites for the pollen–soil comparison sorted by longitude (Reference system: WGS84).
Coarse parent material data were used to further explore the relationship between pollen and soil composition, since this variable displays the most notable correlation coefficient with Corylus percentages (r = −85, Figure 5a; Table S1, Supplementary Material). The Corylus pollen dataset was divided into two parts using 40% as a threshold value. Sites with >40% Corylus display a significantly lower presence of coarse parent material compared to sites with <40% Corylus (t = 4.58, p < 0.05, Figure 5b). The use of a 40% threshold represents an arbitrary choice motivated by the decision to divide the dataset into two parts of approximately equal size. Considering this clear divide, a correspondence analysis (CA) was carried out using all parent material data for all pollen archives (Figure 5c). For reference, soil data for Friesack were extracted using the same procedure and included in the CA. The CA highlights the paedological similarities between Friesack and sites with low Corylus percentages, as also suggested by its position in Figure 4.
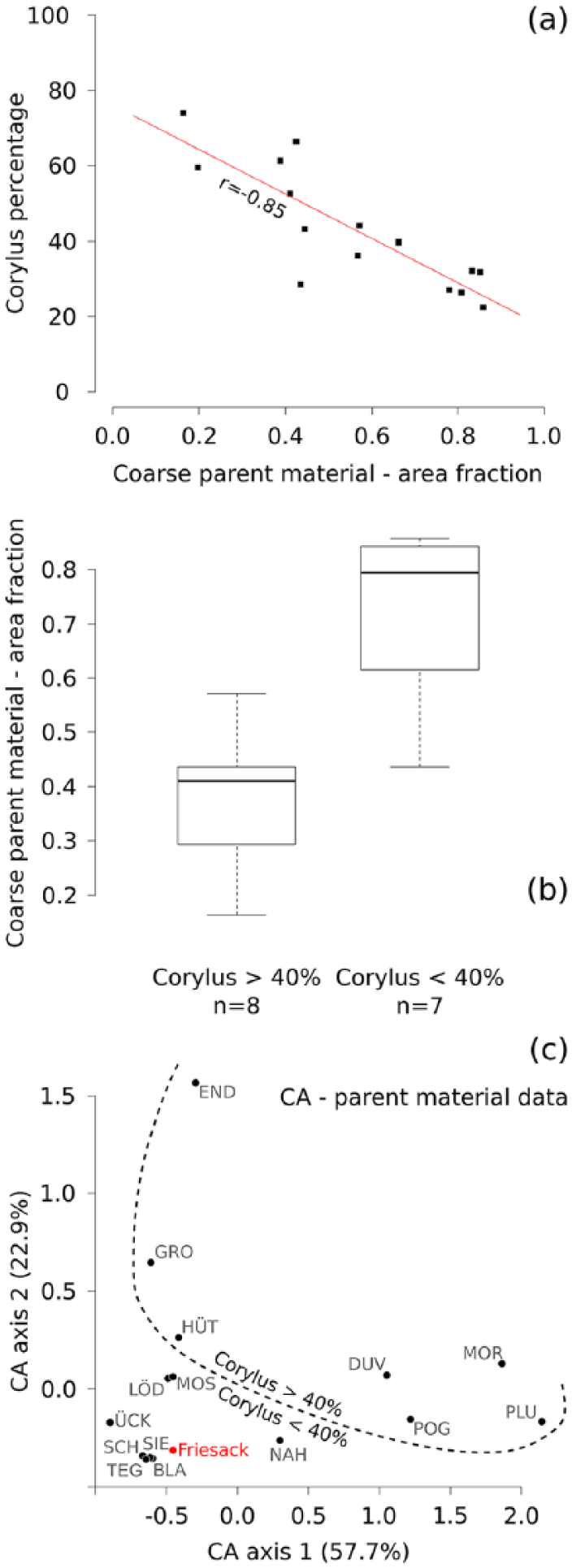
Comparison between Corylus pollen and soil composition for selected data. (a) Linear correlation between Corylus percentages and areas covered by coarse parent material. (b) Differences in soil composition (area with coarse parent material) between sites with <40% and >40% Corylus. (c) Correspondence analysis including all parent material variables. The proportion of variance explained for each axis is reported in brackets. The dashed line was added to provide a visual aid with separating sites with <40% and >40% Corylus.
Conclusion
For future studies, a higher number of sites with minimally overlapping pollen source areas would constitute a desirable condition to draw more solid conclusions. Yet, the currently available data appear to show a strong environmental control upon Corylus in pollen diagrams, suggesting that the humans’ persistent strategy of harvesting and procuring hazelnuts in the Duvensee area is very likely connected to good regional growing conditions for hazel over centuries. A direct human intervention on the landscape aimed specifically at increasing hazelnut yields does not appear to be inferable from the available data. This does not mean that people did not support Corylus intentionally or unintentionally where it naturally grows, for example, by hunting browsers (herbivore mammals feeding on woody plants), but they most probably did not influence the habitat of the shrub to such an extent that it would be detectable in the archaeological and environmental records.
While indications are missing for Duvensee that specific plants were intentionally promoted during the early Holocene, such evidence has been detected in other areas or phases of the Mesolithic (e.g. Divišováa and Šída, 2015; Innes and Blackford, 2003; Innes et al., 2013; Klooß, 2014, 2015; Law, 1998; Mithen et al., 2001; Regnell, 2012; Sjöström, 2009; Woldring et al., 2012). A plausible example is, for instance, reported from Mesolithic Scotland, where hunter-gatherers systematically used hazel and oak for food and fuel and it seems that they supported them in particular through the maintenance or creation of woodland clearings – potentially a case of an early vegetation management strategy which included the construction of new ecological niches (Bishop et al., 2015).
Strategies of active niche construction: Hunting female roe deer at Friesack
Contrary to the extensive evidence for hazelnut collecting in Duvensee, the sites near Friesack highlight the importance of animal protein in the Mesolithic diet. Friesack has provided one of the best early Holocene zooarchaeological assemblages on the Northern European Lowlands (Gramsch, 2000; Schmölcke, 2016). Here, the oldest three occupational phases of the Preboreal and Boreal are of special interest. While the Duvensee sites give clear seasonal indications for settlements during autumn, the sites at Friesack were repeatedly used at least during summer, but most likely over most of the year (cf. Schmölcke, 2016).
During the Mesolithic, human hunting strategies could influence niche construction in different ways. Among them is the displacement of competitor species living in the same habitat and using similar resources, or the massive reduction or even extinction of over-hunted species (Rowley-Conwy and Layton, 2011 with examples). In addition, different species could have profited from human activities as well. In Friesack, for instance, it is apparent from the osteological evidence that the number and proportion of hunted roe deer (Capreolus capreolus) increased from the earliest to the latest phase under consideration, even though Capreolus is, in contrast to aurochs (Bos primigenius), elk (Alces alces) or red deer (Cervus elaphus), a relatively small, energetically less productive prey (Schmölcke, 2016). According to Optimal Foraging Theory (Pyke, 1984; cf. Mohlenhoff et al., 2015), in hunter-gatherer communities, the exploited resources should be ranked in line to their energetic return per time expended in their acquisition. Contrary to this, the human strategy was different in Friesack where people more relied on a successful hunt than getting a maximum of food resource per expedition. Since, for hunters, it is comparatively easy to track down Capreolus and hunting them is not dangerous.
This conclusion can be drawn based on demographic trends in this species such as individual age and sex ratio. Animals with an individual age between 1 and 2 years were by far the most extensively hunted age-group in Friesack with between 38% and 53% in the three occupation phases, while these age-groups represent only ca. 15% in natural populations (Schmölcke, 2019). Furthermore, there is a clear prevalence of female roe deer in the archaeological material (Schmölcke, 2016). As Capreolus is rather stationary and prefers stands where they have a good overview of the surroundings (especially) during breeding season, they are easily spottable by hunters during that time. The combination of notable demographic patterns and increasing significance as hunting prey in the course of the Mesolithic indicates that, triggered by the human behaviour, Capreolus started to behave in a more r-selected manner, that is, to reproduce at higher rates. Comparative studies on modern populations have shown that roe deer is generally more r-selected than other European game species and reacts immediately and flexible to population density changes (Hartl and Reimoser, 1988; Putman et al., 1996). Similar reactions to hunting pressure are also known from other species such as beaver (Castor fiber), where adults get more youngs the next year if many juveniles were culled (Dods, 2002). In Friesack, a similar picture emerges for wild boar (Sus scrofa) with young adult females predominantly hunted by humans as well as young boars (Schmölcke, 2016). Sus scrofa can be considered a typical r-selected larger mammal with very high reproductive potential. Hypothetically, the observed increasing proportions of juveniles and young adults in both Capreolus and Sus could show overhunting of the local populations as was argued for Swiss red deer populations during the Neolithic (Schibler and Steppan, 1999). However, it seems to be unlikely that small forager groups had the same direct influence on game populations in a completely forested landscape as farmers in an artificially opened environment. In addition and independent from human activities, it should be noted that the increasing population density of Capreolus in the surroundings of Friesack over time was probably also due to the early Holocene climate warming, because midwinter temperatures have a strong effect on roe deer populations (Putman et al., 1996).
These patterns are of further interest when other human exploitation practices as well as stable isotope analyses of human subsistence are included: as has been shown by Meadows et al. (2018), the diet of the Mesolithic people at Friesack has changed over time from a more aquatic to more terrestrial food (cf. Benecke, 2016; Robson, 2016). This might be reflected in the need for a more resilient hunting strategy which is possibly documented by the described kill-off patterns.
However, it must be considered that Friesack has been occupied over a long period. Detailed stratigraphic analyses (Gramsch, 2000) make it possible to differentiate approximately 120 occupation events. These are summarized in four main occupation phases, and while the bone assemblages have been analysed according to these, the single phases reproduce several events occurring at different times of the year. As a consequence, the overall picture for each phase may be blurred – a general methodological problem in the analysis of long-term or repeatedly occupied archaeological sites. However, investigations on the age and sex distributions in Sus scrofa remains from Mesolithic settlements in Southern Sweden also reflect fundamental differences in hunting strategies by optionally promoting younger or strong old animals (Magnell, 2005). It is very likely that such selective prey choices influence the local animal population, for example, in terms of behavioural changes. In addition, it remains to be investigated whether the humans increased intentionally and purposefully the carrying capacity of their preferred prey species by, for instance, vegetation management strategies.
Ethnoarchaeological case study
In order to better understand patterns of human–environment interactions in forager communities, ethnoarchaeological observations can provide new perspectives on archaeological data. For understanding the role of human and non-human agency in shaping landscape, biosphere and culture, investigations among indigenous hunter-fisher groups can add valuable insights. The aim, however, is not to detect direct analogies and hence explanations for certain patterns in the archaeological record. Such an approach would be jeopardized by the general problem of comparability of prehistoric and contemporary societies, since the latter are an integral part of the modern increasingly globalized world (Clarke, 1972; Lane, 2014; Wylie, 1985; see also Gosselain, 2016). Thus, there is no continuum between historically singular formations on one hand and anthropological universals on the other.
Ethnoarchaeological information rather can be used to trace dimensions of variability in the reasons for and mechanisms of human behaviour and coping strategies. Another important point concerns the possibility to assess the potential archaeological visibility (or disguise) of certain ecological, economic and cultural features shaped by such mechanisms and to integrate indigenous specialist knowledge on technologies, meanings and even interspecies interaction (e.g. Grøn, 2012).
People, reindeer and mosquitoes
Studies among Western Siberian indigenous groups in the taiga provide insights into contemporary forager societies of the temperate climatic zone. Comparisons to early Mesolithic living conditions can be drawn on ecological and economic grounds. Here, we focus on the Selkup who have preserved their nomadic lifestyle in a forest environment until today (cf. Poshekhonova et al., 2017).
About 300 years ago, some of the Selkup people migrated 500 km to the north to the area of the Taz River and unintendedly extended their original hunter-gatherer-fisher subsistence model by the uptake of a kind of symbiosis with local reindeer (Rangifer tarandus; Golovnev and Tuchkova, 2005). Over time, reindeer became confiding to the people by learning that the smoke of the human fireplaces effectively protects against the myriads of mosquitoes occurring there every summer. The humans responded as they started to provide special smoke fires around their camps where the reindeer assemble to be relieved from the biting vermins. These fires are either located under the open sky or at both ends of specially constructed reindeer houses, in which they take shelter (Figures 6 and 7). Hence, the deer roam freely in the forests and wetlands around the human summer settlements but return to the camps to rest and find relieve from the insects on a daily basis. In autumn, at the end of the mosquito season, the reindeer start to wander off and have to be recollected in winter to be used as draught animals for the sledges (Kenig, 2010). Therefore, the humans feed them dried or frozen fish to lure the animals close to the settlements in that season. Consequently, the Selkup’s intense winter fishing with stationary constructions does not only provide food for the people but is also an integral part of their cohabitation with reindeers.

Open-air smoke oven providing relief for the reindeer from the mosquitoes. Selkup summer settlement, upper Taz region, Western Siberia, summer 2017 (photograph: C. Engel).

Reindeer house with two smoke ovens, with post construction incorporating an in situ tree (in the foreground) that would not be recognizable archaeologically as part of the ground plan. Selkup summer settlement, upper Taz region, Western Siberia, summer 2016 (photograph: H. Piezonka).
The symbiosis of reindeer and the Northern Selkup has led to an adjustment of the people’s seasonal cycle as well as the associated settlement system and building structures. Further south in their former settlement area, the Selkup’s hunting and fishing activities followed a recurring seasonal round based on the predictable abundance of various resources with specific locations for the respective seasonal settlements. Contrary to this, the uptake of reindeer husbandry in the north made the seasonal movements less predictable because the best summer grazing spots for the herds vary from year to year, and hence new locations have to be chosen more frequently for the settlements. As a consequence, also the houses of the Selkup have changed: Formerly, the main winter dwellings were sunken-floor earth houses with wall and roof constructions made from wood, birchbark and earth and clay-built ovens on one of the side walls. They could be reused over several seasons. Nowadays, further north, the winter houses became less stable: the floor is no longer sunken (apparently to spare the additional labour-intense work; Adayev and Zimina, 2016), and the interior arrangement with a central hearth or metal oven resembles the more simple, symmetrical layout of the conical tents that are used during summer and as light winter dwellings for small hunting parties.
Material indications
There are several material indications for the Northern Selkup’s adjusted economic and settlement system that could be detectable by archaeological methods, including the change in winter house building traditions, the appearance of the post-built reindeer houses with the two smoke ovens as a new building type, the construction of open-air smoke facilities that are lined by stake fences to prevent the reindeer from getting too close and evidence for reindeer husbandry such as bridle parts. It is questionable though whether the niche construction for the newly established reindeer–human cohabitation by providing relief from the mosquitoes could actually be recognized, as the insects themselves are invisible archaeologically. Drawing a connection between this relevant but undetectable factor and the observable material changes would hardly be decipherable by archaeological means alone, without ethnographic knowledge of such practices. As an additional challenge, naturally rooted trees are frequently incorporated into the layout of the post-built structures of the reindeer houses, leading to ‘incomplete’ ground plans in the archaeological record (Figure 7). From a long-term perspective, it might be possible to trace the beginning of reindeer herding by archaeometric means, as the Selkup’s practice of feeding fish to the generally herbivorous animals in winter might be recognizable through the isotopic (13 C/15 N) signature of the animals’ bones.
Discussion: On evidences and absences
To deduce prehistoric hunter-gatherer behaviour from archaeological evidence, it is necessary to take into account how humans have inhabited and appropriated their natural surroundings. As they lived in their own ecological niche all along, their ability to recognize the potential of the landscape and its specific resources has been shown in manifold ways. For instance, practices involving intentional burning, irrigation, pruning, sowing, tilling, transplanting and weeding are reported from hunter-gatherers from California where humans temporarily influenced some plant communities, but without permanent changes of the whole biotope (Anderson, 2005). Further examples show similar modifications of the natural surroundings (e.g. Arnold, 1996; Bang-Andersen, 1996; Binford, 1980; Brinch Petersen, 2012; Edwards, 1996; Grøn, 2012; Ingold, 1983, 2000; Tipping, 1996; Wiessner, 1982). However, the general assumption which is underlying many studies on the topic is that people with a ‘Mesolithic lifestyle’ intuitively understood what we call the Concept of Sustainability: they recognized the importance of sustainably utilizing the resources provided by nature and had daily practical experience of the fact that they are dependent on functioning natural systems for their own survival (Hill and Bowen, 1997; Ingold, 2000; van der Post and Taylor, 1984). As a result, they have been able to maintain an equilibrium relationship with the natural environment over long periods of time, and at the same time had to be able to recognize potentials of purposeful changes of restricted aspects of the surrounding landscape. However, this preconception is difficult because many – sometimes quite different – groups of people are compared and hence quite diverse ways of dealing with the environment are usually simplified for summary or analogy (cf. Warren, 2018). Nonetheless, the application of niche construction theory to archaeological and ethnographic contexts can help to better understand these entanglements, whereas ethnoarchaeological studies can also highlight aspects of their archaeological footprint.
A good archaeological example for this is provided by another Northern German Mesolithic site, Hohen Viecheln, where the animal bone assemblage shows a significant lack of fish remains (Wundsch, 1961). Since the site was located at a lakeshore and finds of over 300 bone points (and fragments) made extensive fishing activities very likely, it has been assumed that the fish carcasses were fed to dogs, leaving no archaeological traces. Even though the excavation strategy was not adequate for a Mesolithic site (Schuldt, 2005) and therefore the lack of fish remains may also be due to insufficient documentation and sampling, recent stable isotope analyses of dog bones from Friesack confirm that Mesolithic dogs’ subsistence could in fact have relied to large extends on aquatic resources (Ewersen and Schmölcke, 2013; Ewersen and Ziegler, 2014 cf. Fischer et al., 2007 with similar results for Danish Mesolithic dogs). Likely, leftovers from fishing were disposed as fodder to the dogs (cf. Schmölcke, 2016). Fish is widely regarded to have been a basic staple for Mesolithic communities (e.g. Boethius et al., 2017; Meadows et al., 2018; Ritchie et al., 2016; Robson, 2016; Robson et al., 2018; Schmölcke, 2016; Solheim and Persson, 2016; Terberger et al., 2012, 2018; Zabilska-Kunek et al., 2015), though bone remains are sometimes scarce for several reasons (cf. Dark, 2003; Hartz et al., 2011), some of which become clear from ethnoarchaeological observations (cf. Lovis and Hart, 2015).
Examples for the extensive use of fish can be found in contemporary Siberian hunter-fisher-herder communities as well, where fish plays a crucial role for securing nutrition of humans and also of dogs and reindeer in winter (Adaev, 2007; Golovnev, 1995). The Northern Selkup use different ways of securing a sufficient and durable supply of fish and fish parts as food for humans and fodder for animals. One way is smoking in open sheds over a continuously smouldering fire. While certain species are prepared as human food (e.g. salmonids), the less valuable or tasty species are prepared for the dogs the same way. In addition, the carcasses of filleted fish are also smoked for dog fodder. Another strategy is based on the autumn fishing season when, at the time of the first frosts, large amounts of fish are caught in stationary fishing constructions and stored frozen in specific log-built storage boxes next to the winter dwellings. This stored fish then serves as winter supply for the humans, dogs and reindeer. As the latter are also fed from this supply, these stores do not contain perch, as the stingy fins are dangerous for the soft reindeer mouths (V. Irikov, Pokalky, recordings by V. I. Adaev and H. Piezonka during field work, 2017). In the Selkup settlements, these practices lead to an almost complete disappearance of fish parts from the material remains, as has also been proven by excavations of Selkup winter houses in the Taz region (Poshekhonova et al., 2017).
Similar mechanisms can be considered for archaeological contexts such as Hohen Viecheln, if the extensive and complete use of all parts of fish as human food and animal fodder was practised (cf. Robson et al., 2018). Tracing this will only be possible through coprolites or isotope analysis (e.g. Reinhard et al., 2007; Zhilin et al., 2014). The various structures connected with catching, preparation and storage of fish have different chances of being detected in the archaeological record. While remains of stationary fishing structures such as fish weirs and fish traps can be preserved under waterlogged conditions (for the study region, see Klooß, 2015; for other parts of temperate Mesolithic Europe, see Fischer, 2007; Koivisto and Nurminen, 2015; Lozovskaya, 2013; McQuade and O’Donnell, 2007), the structures for processing and storage situated within the settlements on dry land are harder to find and recognize (a possible Mesolithic example for a Mesolithic storage pit is discussed in Lübke, 2009; cf. Brinch Petersen, 2012), although they must have been essential for Mesolithic people (Ingold, 1983). Hearth-pits and fireplaces leave traces of the (potential) smoke fire, but actually connecting them to the preparation of fish is difficult (Groenendijk, 2015). Log-built storage boxes like the ones used by the Selkups have no subterranean parts and would therefore disappear without a trace after the disintegration of the wood, while fishing gear such as hooks and harpoons made from bone would be recognizable in the archaeological record if deposited in preserving environments.
Furthermore, it must be considered that the detected particularities in the archaeological record may have been caused by other, ‘irrational’ (Adaev, 2005) factors such as cultural practices or transformed perceptions of the environment. An example is the meticulously regulated ritual disposal of bones of different prey species in predestined places outside the settlements, as known from, for example, Evenk and Khanty communities in the Siberian taiga (Grøn and Kuznetsov, 2003, 2004; Jordan, 2003). The extensive hunting practised by these groups would not be recognizable in the archaeological record from osteological material of the settlements alone, as no bones are found.
Moreover, within the archaeological record, we are faced with the challenge of dissimilar contemporaneity: what appears to have happened synchronously in the past may be a result of accumulated activities (e.g. Cziesla, 1990) and vice versa: what might appear as different structures today might have belonged to the same settlement or event (cf. Brinch Petersen, 2012). This is of special relevance when dealing with processes that actively transform the environment or create niches, because at hunter-gatherer sites with long-term occupation and a more or less unstratified find layer, it is hardly possible to detect direct anthropogenic influence on the landscape or fauna in and around the settlement. The same is true for short-lived sites but long-term proxies, like in our example from ancient Lake Duvensee. Here, the palynological record reflects longer timeframes than the sites actually cover. So, a direct contemporaneity of the pollen signal and the settlements is hardly traceable. Vegetational changes could have appeared before, after or during the use of the area and hence might have been the result of other triggers than human influence. Furthermore, it has been shown that an anthropocentric perspective easily leads to misinterpretations of the record. Due to the large amount of hazelnuts at ancient Lake Duvensee, the possibility of human manipulation of the bushes was already indirectly suggested by Schwantes et al. (1925) when they proposed for the first excavated site that ‘hazel played the role as the later wheat’ (own translation). As has been shown above, it is more likely that the area has provided hazel with rather good growing conditions. Therefore, it is yet more likely that the ecologically favourable conditions for the bushes have played a more important role for their exploitation than human manipulation of the plants.
Conclusion
It is yet to discuss to what extend humans’ impact on their environment during the early Holocene might have caused long-lasting effects. Traceable changes in landscapes or faunal composition are hard to investigate due to the specific character of hunter-gatherer campsites. Therefore, addressing such aspects will have to rely mostly on circumstantial evidence. However, studies have shown that people’s coping strategies have adjusted to specific environmental niches or have been developed to overcome shortages and environmental changes (e.g. Adaev, 2005; Anderson, 2005; Boethius, 2018; Regnell, 2012; Schmölcke et al., 2006; Woldring et al., 2012). Even aspects like perishability of food were sophistically faced, by, for instance, fermenting and thereby preserving fish as a study from southern Sweden recently showed for the mid-Mesolithic (Boethius, 2016).
By critically examining the available data, it becomes clear that it is possible to find indications that argue for and against active niche constructions in prehistoric hunter-gatherer societies. Since we merely see the material results of such processes, niche construction needs to be addressed with an inductive approach. As has been shown by the ethnoarchaeological example, that even tiny, archaeologically undetectable agents like mosquitoes can contribute, in the long run, to substantial changes in the economic and settlement systems. But only the latter leave a multitude of archaeological traces such as new building types, zooarchaeological and biometric data, and different seasonality evidence. The investigation of such interrelated developments can lead to a better understanding of human–environment interaction and social transformation in the past.
However, as our case study from ancient Lake Duvensee has shown, it is possible to investigate probabilities with the given data for active alteration of the environment by prehistoric hunter-gatherers. Applying a theoretical framework and approaching it empirically can help to better untangle ecology and economy and thus help to understand human behaviour. It can even show that often propagated assumptions and convictions become more unlikely, like we have shown with respect to the growing conditions of hazel. This also stresses the advantage of incorporating multi-species perspectives into our considerations for understanding parts of ecosystems before anthropocentrically judging the archaeological evidence. The example from Friesack showed potential for addressing anthropogenic niche construction from a rather pin-pointed zooarchaeological perspective. As a consequence, it can be said that approaching prehistoric data from a process-oriented perspective like niche construction theory provide good interpretational possibilities for understanding anthropogenic transformation of the environment, the impact of people and societies as well as subsequent results of their behaviour.
Supplemental Material
Gross_etal-FigS1 – Supplemental material for Adaptations and transformations of hunter-gatherers in forest environments: New archaeological and anthropological insights
Supplemental material, Gross_etal-FigS1 for Adaptations and transformations of hunter-gatherers in forest environments: New archaeological and anthropological insights by Daniel Groß, Henny Piezonka, Erica Corradini, Ulrich Schmölcke, Marco Zanon, Walter Dörfler, Stefan Dreibrodt, Ingo Feeser, Sascha Krüger, Harald Lübke, Diana Panning and Dennis Wilken in The Holocene
Supplemental Material
Gross_etal-table_s1 – Supplemental material for Adaptations and transformations of hunter-gatherers in forest environments: New archaeological and anthropological insights
Supplemental material, Gross_etal-table_s1 for Adaptations and transformations of hunter-gatherers in forest environments: New archaeological and anthropological insights by Daniel Groß, Henny Piezonka, Erica Corradini, Ulrich Schmölcke, Marco Zanon, Walter Dörfler, Stefan Dreibrodt, Ingo Feeser, Sascha Krüger, Harald Lübke, Diana Panning and Dennis Wilken in The Holocene
Footnotes
Acknowledgements
This article is part of the Collaborative Research Centre 1266 ‘Scales of Transformation – Human-Environmental Interaction in Prehistoric and Archaic Societies’ funded by the Deutsche Forschungsgemeinschaft (DFG). The research has been undertaken within the Interlinking Group ‘Human-environmental Interaction’. The work of the European Pollen Database data contributors and community is gratefully acknowledged. Furthermore, the authors thank two anonymous reviewers for their valuable comments and suggestions for improving the text.
Funding
The authors disclose receipt of the following financial support for the research, authorship, and publication of this article: This study was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation, Projektnummer 2901391021 – SFB1266).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.