
Abstract
This is the first time an archaeobotanical analysis based on macroremains, both charred and desiccated, from Cambodia is reported. The archaeobotanical samples are rich and provide evidence of rice processing, consumption of non-indigenous pulses, and the use of economic crops. The evidence is supported by data from inscriptions, texts and historical ethnography. This study demonstrates that the city of Angkor in the 14th and 15th centuries CE, despite its decline, was still occupied. Angkor’s inhabitants continued their everyday lives cultivating and consuming their staple food, rice, with a suite of pulses, and also used the harvests in the performance of rituals.
Introduction
From its vast urban centre on the northern flood plain of the Tonle Sap lake, Angkor is recognised by its unique monumental and artistic artefacts, its dynamic cultural influence and its enduring significance to Cambodian identity and world heritage. Until recently, archaeobotanical research at Angkorian archaeological sites has been limited. Analysis of plant consumption at Angkor Thom started with palynology commissioned by French archaeologist Bernard-Philippe Groslier and conducted by Madeleine Van Campo of the Muséum national d’Histoire naturelle (Paris) using materials from two excavation campaigns inside the Royal Palace enclosure (Groslier, 1959, 1960, see also Pottier, 2014). The focus has been rice agriculture because of its importance to the population of Angkor and links to hydrological infrastructure (e.g. Groslier, 1974, 1979; Hawken, 2011, 2013; Pottier, 2000; Van Liere, 1980, 1989). Since Groslier’s pioneering work, archaeobotanical research has concentrated on pollen analysis (Penny et al., 2005, 2006, 2014), offering a broad record of past vegetation. However, there has been no elaboration or additional detail of the plant species used in the Angkorian kingdom (c. 800–c. 1450 CE). Recent fieldwork in Cambodia has yielded botanical remains of rice and other economic crops dated by accelerator mass spectrometry (AMS) 14C analysis to the 14th and 15th centuries.
At the core of Yaśodharapura (Angkor) is the 9-km2 walled urban enclosure of Angkor Thom. The Royal Palace precinct in the northwest quadrant of Angkor Thom covering an area larger than 1 hectare is characterised by temples, stone-lined ponds, embankments, enclosure walls, and an extensive array of wooden buildings which have now perished but are recognised by roof tile remains and postholes. The first phase of the Royal Palace probably dates to the late 9th or early 10th century, and its enclosure wall from the late 10th or early 11th century. Angkorian occupation at the site is commonly accepted until the first half of the 15th century (e.g. see Gaucher, 2013; Groslier, 2014 [1958]; Pottier, 2014). During excavations north of the Royal Palace, which focussed on an 11th–12th century copper-base alloy foundry and sculpture workshop (Polkinghorne et al., 2014), soil samples were collected for botanical remains analysis.
While the collapse of Angkor has been of interest since the beginnings of modern scholarship of the city and ancient Khmer kingdom, continued occupation has been little studied. Colonial savants initially recognised an Ayutthayan invasion in the Royal Cambodian and Royal Ayutthayan Chronicles as the most straightforward explanation for a catastrophic episode that supposedly involved the relocation of many hundreds of thousands of people (Aymonier, 1904; Garnier, 1871; Leclère, 1914; Moura, 1883). Additional causes were supposedly the weakening of power structures associated with the introduction of Theravāda Buddhism, and exhaustion of the population through overwork (see Briggs, 1951; Cœdès, 1963; Finot, 1908; Glaize, 1993 [1944]; Kulke, 1993; Leclère, 1899; Mabbett, 1978). Recent studies have made great advances in providing alternative understandings to the why of an Angkorian demise. New economic opportunities (Vickery, 1977), over-commitment to massive infrastructure (Fletcher et al., 2008), unsustainable modes of habitation and subsistence (Evans et al., 2013) and severe climatic events (Buckley et al., 2010, 2014) are all viewed as cumulative factors precipitating decline. However, these studies offer limited or no evidence for endurance of occupation at Angkor between the late 13th and early 16th centuries. The archaeobotanical remains provide new information on the agricultural activity, food consumption and ritual during the decline of Angkor.
Excavation methods and materials
Excavations were conducted adjacent to the Royal Palace of Angkor Thom to investigate associated craft production (Polkinghorne et al., 2014). From these excavations, one group of archaeobotanical samples was extracted from an in-filled pond (Khmer sraḥ), another from a ritual deposit in a lead vessel (Figure 1). The central nature of this location and the richness of its archaeological record are represented in these samples and illuminate life in Angkor at its most famous urban complex.

(a) Angkor Thom, location of the excavation north of the Royal Palace. LiDAR image courtesy the Khmer Archaeological LiDAR Consortium. (b) Angkor Thom (detail) location of the relevant excavation trenches. École française d’Extrême-Orient (EFEO) courtesy Christophe Pottier, annotated by Martin Polkinghorne.
In-filled pond (Trench 4)
Between 1918 and 1934, French archaeologist Henri Marchal conducted surveys and excavations to demarcate the extent of an in-filled pond north of the Royal Palace walls (Commaille and Marchal, 1909–1918; Marchal, 1918, 1918–1921, 1919; Marchal and Trouvé, 1934, 1934–1935). He identified a small sandstone-block wall decorated with nautical reliefs corresponding in style with another in-filled pond (Sraḥ Srei) inside the Royal Palace enclosure. Five excavation trenches were opened, one at each corner, and one at the eastern end. Marchal discovered descending laterite steps that designated a pond bordered in stone (Marchal and Trouvé, 1934: croquis no. 2). The foundations of the decorated wall, which relate directly to the basin, and holes in the laterite pavement, suggest a series of associated wooden structures.
In 2012, an excavation trench (Trench 4) was opened to establish the relative chronology of the sraḥ to the craft production situated to its east. Trench 4 straddles the sraḥ being both inside and outside and avoids Marchal’s excavations. Excavations revealed 14 laterite steps leading to the original floor of the pond. The sraḥ is an exiguous catchment of secondary fill which contains charcoal and organic matter from the surrounding area, and its sediments are rich in macrobotanical remains. Observed in cross-section, the stratigraphy was superpositioned in smooth and wavy layers. The absence of tree-roots, termite bioturbation, and modern artefacts suggests the excavation trench is an undisturbed archaeological record (Appendix 1). Furthermore, context security of soil samples for macroremains analysis was established by examining the proportion of roots, modern seeds and fungal sclerotia. That proportion was low for all samples analysed. Layers, herewith described as contexts, were interpreted and grouped in the field by horizon boundaries defined by matrix colour with a Munsell colour chart, and estimation-by-feel of consistence and texture class (Munsell Color, 2010; Schoeneberger et al., 2002; Thien, 1979).
In the western section of Trench 4, excavators recognised 31 contexts that can be broadly grouped into five phases (Figure 2a). Phase 1 (contexts #31 to #28, 21.44–22.11 m a.s.l., measured from the corner of the western and southern walls) is primarily silt clay, dark yellowish brown (10YR 3/4) to very dark grey (10YR 3/1), and contains ceramic sherds, sandstone and laterite fragments, and charcoal pieces. Phase 2 (contexts #27 to #24, 22.11–22.52 m a.s.l.) is predominantly loamy sand/sand, dark yellowish brown (10YR 4/4) to brown (10YR 4/3), and contains ceramic sherds, and sandstone and laterite fragments. Phase 3 (contexts #23 to #21, 22.52–22.71 m a.s.l.) is chiefly a silt clay/clay, black (10YR 2/1) to very dark greyish brown (10YR 3/2) with very few artefacts. Phase 4 (contexts #20 to #3, 22.71–23.98 m a.s.l.) is predominately loamy sand, yellowish brown (10YR 5/6) to brown (7.5YR 4/3), interbedded with two brown (7.5YR 4/4) sand layers (contexts #17 and #11, 22.71–22.98 and 22.94–23.17 m a.s.l., respectively) and a very dark greyish brown silt clay layer (10YR 3/2) (22.68–22.74 m a.s.l.), and contains ceramics, sandstone and laterite fragments. Finally, Phase 5 (contexts #2 and #1, 23.98–24.18 m a.s.l.) is a black (10R 2/1) to very dark brown (10YR 2/2) silt loam topsoil containing ceramics, sandstone and laterite fragments.
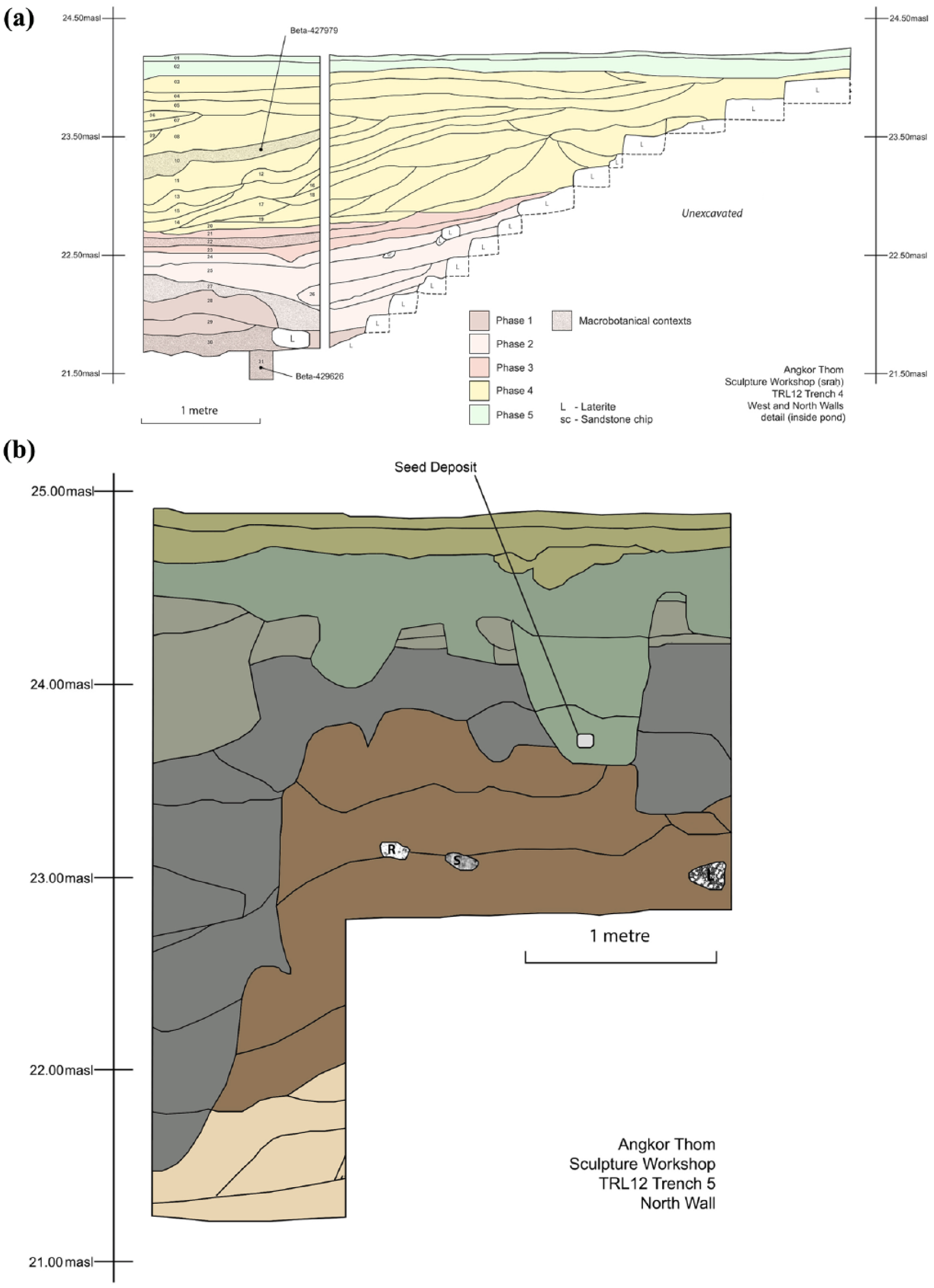
(a) Stratigraphic drawing, inside and outside pond (sraḥ), north of the Royal Palace (Trench 4), illustrating the location of archaeological contexts containing macrobotanical remains and samples presented for AMS 14C dating. (b) Stratigraphic drawing, Angkor Thom foundry and sculpture workshop (Trench 5), illustrating location of seed deposit containing macrobotanical remains and sample presented for AMS 14C dating. Drawings: Martin Polkinghorne and Lugh Lay.
Outside the sraḥ at a depth of 22.52 m a.s.l., the excavation exposed east-west aligned wooden planks set into sandy clay. The addition of postholes in the laterite steps suggests that the sraḥ was surrounded by a wooden structure similar to the galleries known in many Angkorian temples and illustrated on bas-reliefs (Parmentier, 1914). Based on the associated architectural evidence and the centrality of this location, we presume the pond contained water and was used by members of an Angkorian elite who likely inhabited this area. We do not know the date of construction, but a logical assumption situates the sraḥ in the end of the 12th century by linking the structure with stylistically dated nautical reliefs at Sraḥ Srei (Massin, 1997) and known modifications to the Royal Palace supposedly made under the reign of Jayavarman VII (r. 1182/1183 to c. 1220) (Groslier, 2014 [1958]; Pottier, 2014). Evidence for re-excavation and what is possibly deliberate cleaning of water features in the Angkorian period suggests the pond was free of sediment during its operational life (Penny et al., 2005, 2006; Pottier and Bolle, 2009). Therefore, the sraḥ may well have become disused when it began to accumulate sediment that was not cleared.
Based on AMS 14C analysis of charred plant materials and ceramic finds in associated contexts, the archaeobotanical materials and sedimentation of the pond are tentatively dated to the 14th century (Figures 2a and 3). Two samples of charred rice grains (Oryza sativa L.) were presented for AMS 14C analysis. One sample originates from the middle of the stratigraphy (Beta-427979, context #10, 23.17 m a.s.l.), another from the bottom (Beta-429626, context #31, 21.44 m a.s.l.). The AMS 14C age of the bottommost sample (Beta-429626) is marginally later than the upper sample (Beta-427979). The presence of imported diagnostic ceramics at the bottom of the stratigraphy (context #30, 21.70 m a.s.l.) provides complementary evidence for a 14th-century infill of the sraḥ. This context includes a total of 19 sherds dated to the 14th century from China (Dehua porcelain, Yuan green glaze and Longquan celadon) and Vietnam (Trần Dynasty sgraffito with brown glaze inlay) (Dupoizat, 1999; Medley, 1974; Stevenson and Guy, 1997).
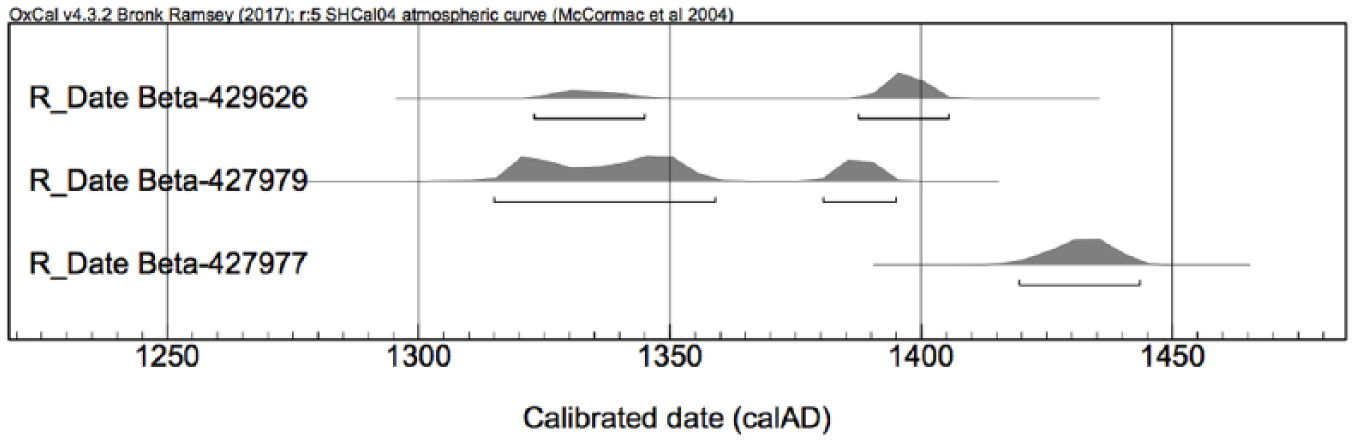
Calibrated radiocarbon dates from macrobotanical remains. SHCal04 calibration data with a small offset of−21 ± 6 years because of monsoonal air-mass mixing (on these methods, see Hendrickson et al. (2013: 41, 43, Table 1)).
We are cautious to date sedimentation of the pond based on two AMS 14C samples. Without additional dates or other evidence, the rate of pond sedimentation cannot be estimated. Furthermore, the calibrated date ranges for the samples are distributed over nearly a century because of interception with a 14C calibration curve plateau in the 14th century (SHCal04, McCormac et al., 2004; on calibration method, see Hendrickson et al., 2013). Because the trench did not present any disturbed contexts, the reason for the inverted dates is unknown. We suppose at least one of the dated samples was displaced from its original stratigraphic position by a process that has not been recognised. In any case, despite an undetermined sedimentation rate, absolute AMS 14C ages of two charred rice grains date the outputs of agricultural activity inside Angkor Thom.
Seed deposit (Trench 5)
Another excavation trench (Trench 5) was located to consider the easterly extent of metallurgical activity at the foundry. It contained an abundance of metallurgical artefacts. Contexts of Trench 5 are broadly grouped into five phases and are described in Polkinghorne et al. (2014). By Phase 4, the foundry and sculpture workshop had ceased operations. Phase 4 had predominantly sandy loam, yellowish brown (10YR 5/6) to very dark greyish brown (10YR 3/2) layers, and excavators discovered a small metal vessel at 23.70 m a.s.l. containing seed remains (Figure 2b). Observation of the trench stratigraphy suggests a small pit was dug to bury the vessel deposit that contained plant materials (Figure 4). The vessel’s body and lid are comparable with Khmer covered box ceramic forms of the danlap type (e.g. see Chhay et al., 2010). A vessel fragment was identified as lead from analysis with a Fourier-transform infrared spectrometer (Douglas, 2012). A desiccated rice husk fragment (Oryza sativa L.) from inside the lead vessel was dated by AMS 14C (Beta-427977). The plant materials, vessel deposit and interment are dated to the early 15th century consistent with interpretation of site stratigraphy (Polkinghorne et al., 2014).

Seed deposit in situ. Photo: Martin Polkinghorne.
Archaeobotanical methods
Soil samples were retrieved for each stratigraphic context from the western section of Trench 4. Not all the soil collected was floated and not all the floated contexts were sorted. The focus of this paper is four samples from different stratigraphic contexts in Trench 4 (context #22, #27, #30 and #31), sorted and analysed to 250 µm, and one sample from Trench 5 of desiccated plant remains found in the lead vessel. All contexts were checked for bioturbation by examining the amount of roots and modern seed contaminants (after Castillo, 2013; see Appendix 1). For context security, the Trench 4 samples were selected on the basis of their stratigraphic position below the topmost AMS 14C sample (Beta-427979) and low bioturbation index.
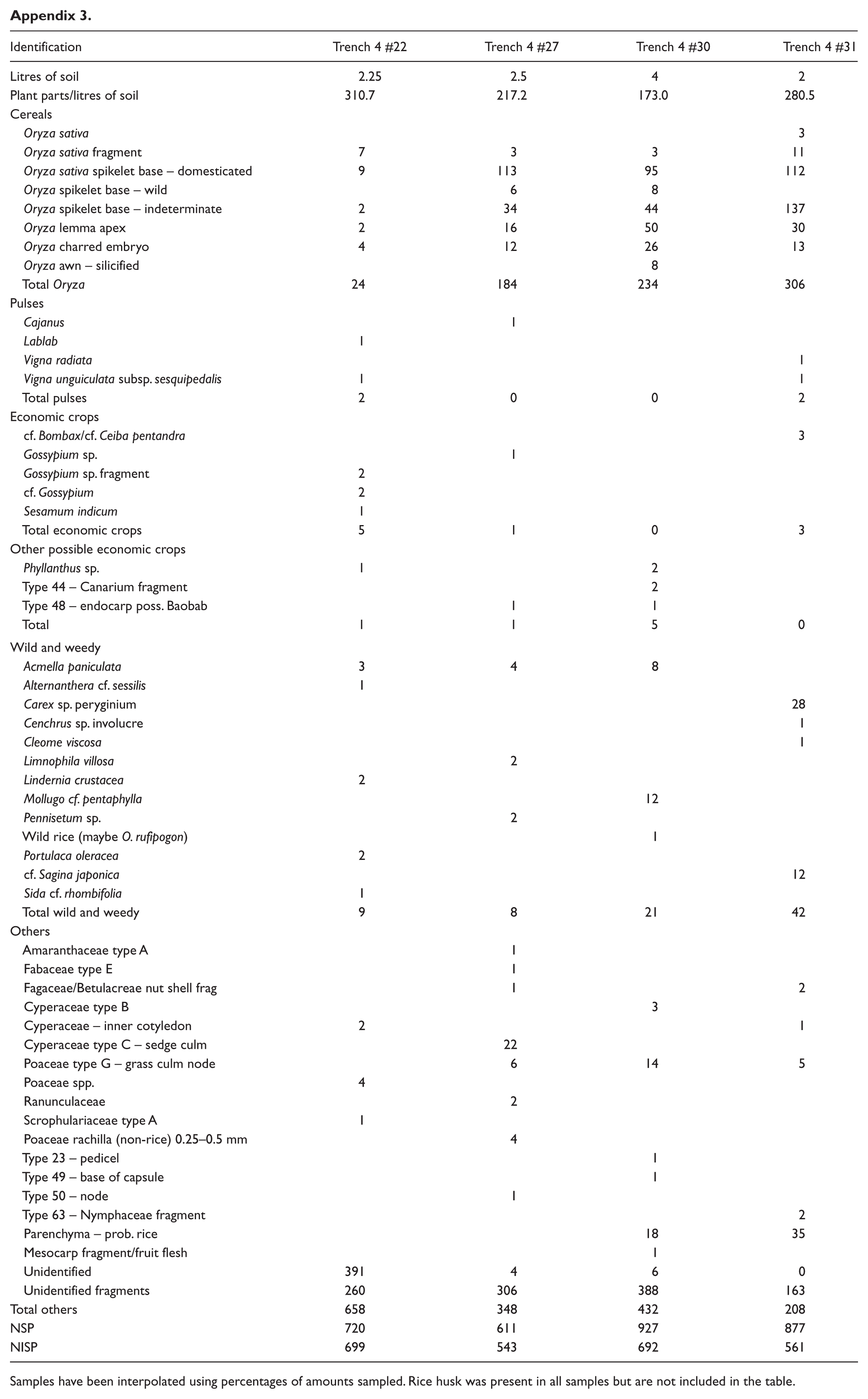
The botanical macroremains were processed using the washover bucket flotation technique (Pearsall, 2000) and 250 µm mesh-sized bags. A total of 48.5 L of soil was floated, an average of 4.85 L of soil per context (see Appendix 2).The heavy fraction was wet-sieved after flotation but it only yielded wood charcoal, which was pulled out but not identified. Of the 10 contexts, four (#22, #27, #30 and #31) were sorted and identified. Half of the contents from contexts #22, #27 and #31 were sorted, whereas context #30 was fully sorted (see Appendix 2). The average number of plant parts per litre of soil is 245 (see Appendix 3 for details), not including rice husk fragments which were estimated as proportions.
To measure the soil collected before flotation, a bucket marked every 5 L was used. To measure the volume of samples after flotation, graduated cylinders were used.
The high average number of plant parts per litre of soil in the excavation samples indicates rich archaeobotanical deposits compared with other sites in Southeast Asia (Castillo, 2013; Castillo et al., 2017). This high average might be the result of low bioturbation, better preservation because the site is relatively recent, dating to the 14th and 15th centuries, or that the samples were recovered from a primary midden.
Seed identifications were conducted in the laboratories of University College London (UCL), Institute of Archaeology with the low-powered microscope Leica EZ4D. The modern reference collection housed in UCL, Institute of Archaeology was used to identify the samples. Botanical identification guides were also used (e.g. Digital Atlas of Economic Plants, Flora of Thailand, GRIN website). Micrographs of the testa and the crops were taken with the Scanning Electron Microscope Hitachi S-3400N at UCL, Insititute of Archaeology to confirm their identifications. Specimens were mounted on studs with double-sided conductive adhesive discs and silver paint.
Rice length and width were measured and compared with other relevant archaeobotanical datasets. To convert the measurements of archaeological spikelets to equivalent grain metrics, we compiled available measurements of unhusked modern spikelets that have been compared with dehusked grains from the same specimens or the sample populations. The sources of such studies include the methodology in Castillo et al. (2016b) and measurements found in Katayama et al. (1972), Zheng et al. (2004), Song et al. (2007), Wan et al. (2008) and Fuller et al. (2010). The resulting formulae used for estimating grain measurements from spikelets is Length: Ls × 0.5347 + 1.3668 = L; and Width: Ws × 0.7004 + 0.3203 = W (Supplementary Material, available online).
Results
The archaeobotanical assemblage in Trench 4 has a high percentage of unidentifiable fragments from the 0.25- to 1-mm fraction. Of the identified plant parts, rice has the highest representation across all samples analysed (Figure 5a). Other economic crops found in Trench 4 are sesame (Sesamum indicum L.), mungbean (Vigna radiata), yardlong bean (Vigna unguiculata subsp. sesquipedalis), hyacinth bean (Lablab purpureus), pigeon pea (Cajanus cajan), cotton (Gossypium sp.) and Indian kapok or silk cotton tree (cf. Bombax/cf. Ceiba). None of these crops are native to Southeast Asia with many from India or via India (Castillo et al., 2016a).
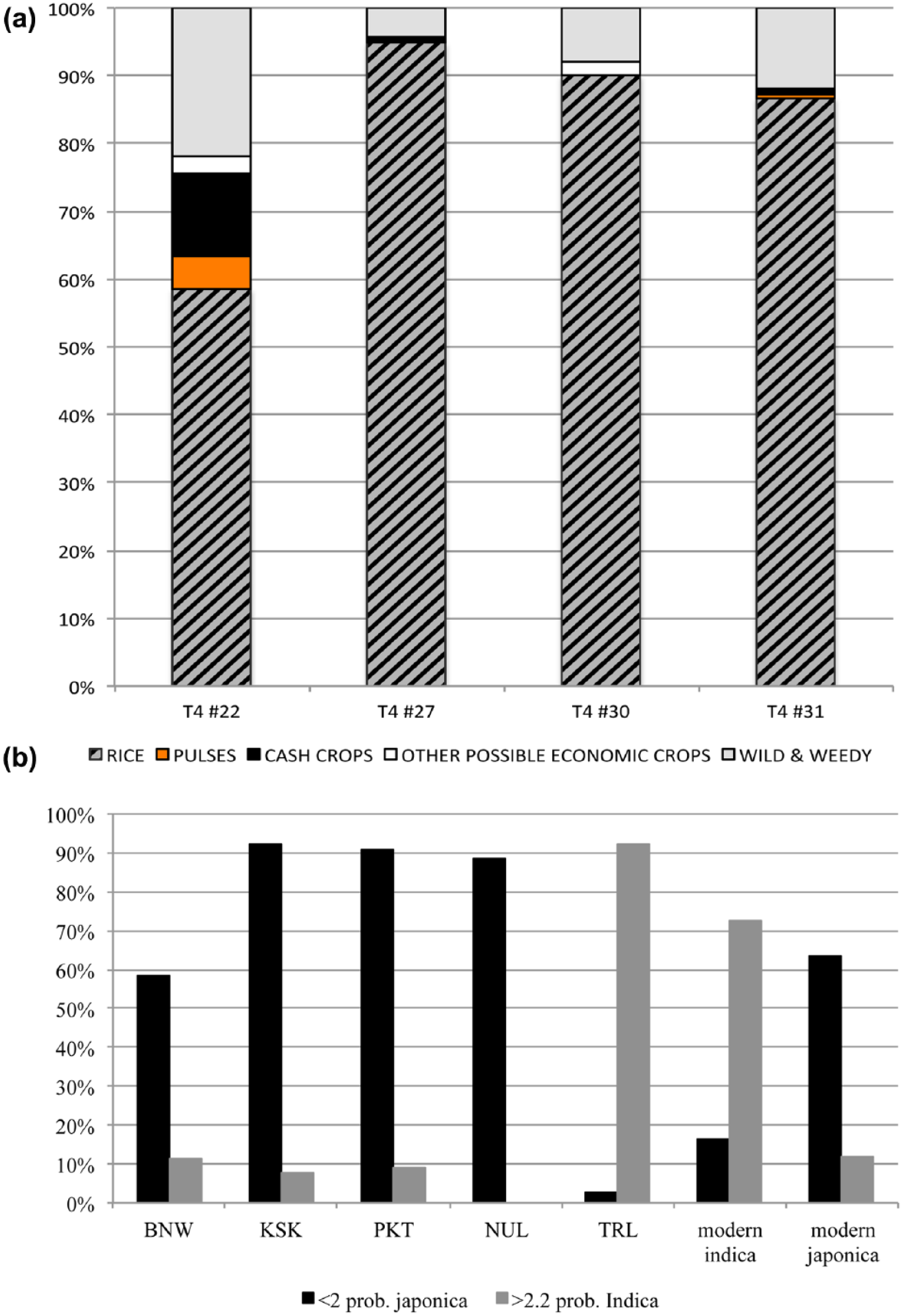
(a) Proportion of plant remains excluding unidentified remains. The rice remains does not include husk. (b) Proportion of indica and [japonica] designation based on the L/W ratio of archaeological rice grains from Ban Non Wat (BNW) belonging to the transitional period between the Bronze and Iron Ages, Khao Sam Kaeo (KSK) and Phu Khao Thong (PKT) belonging to the Late Metal Age, Noen U-Loke (NUL) belonging to the Iron Age, and the Angkor Thom sculpture workshop and foundry (TRL) belonging to the c. 14th–15th CE.
The lead vessel deposit found in Trench 5 is composed of desiccated seed remains. This deposit principally comprises three crops: rice, sesame and mungbeans. All three crops were found in great abundance and more than a thousand seeds of each were identified. The weight of the sample is 55 g and the volume is 125 mL. Morphological features of the seeds were conserved and therefore examination of rice husk and the testa, or seed coat, of sesame and mungbeans was possible. The hilum was also present in whole mungbeans with testa. On the other hand, most of the rice caryopses within the husk were degraded and shrivelled.
Rice (Oryza sativa L.)
Rice dominates the archaeobotanical assemblage from Trench 4 in the form of grains, spikelet bases, awns, husk and embryos. Waste in the form of lemma apices, spikelet bases, awns and husk were more abundant than rice grains and fragments in all samples. The most abundant rice plant part is husk, both silicified and charred (approximately 1% of the 250 µm fraction), indicating crop processing was taking place. Spikelet bases are also abundant in Trench 4 possibly because of the hardy nature of the rachillas, as has been previously noted in charring experiments (Castillo, 2013). The rice is domesticated with a very small percentage (~0–8%) of wild rice (probably O. rufipogon, and weedy rice), and the samples contain both awned and awnless types of rice (Figure 6a and b).
Rice from the deposit of Trench 5 is preserved mainly as dessicated unhusked spikelets. There is a correlation between length and width measurements of spikelets and dehusked grains. Desiccated unhusked whole rice grains from Trench 5 were measured, and 93% of the grains had a length to width ratio (L/W) of >2.2 (Figure 5b).
Sesame (Sesamum indicum L.)
Sesame seeds were found in Trenches 4 (context #22) and 5. The desiccated sesame seeds in Trench 5 are sometimes found in clusters (Figure 6c). Where present, the testa of the sesame seeds possessed a distinctive granular pattern, indicative of the domesticated type (Figure 6d).

Scanning electron micrographs of (a and b) desiccated rice spikelets, (c) a cluster of sesame seeds, (d) the granular surface pattern of a sesame seed coat, (e) a whole mungbean with testa and visible hilum, (f) the surface pattern of the testa of a desiccated mungbean, (g and h) and a split cotyledon displaying the plumule in red. All seeds come from Trench 5. Images: Cristina Castillo.
Mungbean (Vigna radiata)
Mungbeans found in Trench 5 are either whole with testa and hilum or split cotyledons with a visible plumule (Figure 6e and g). Scanning electron micrographs of the surface pattern of the testa show the distinctive elongated cells distributed in a wavy band (Figure 6f). When the hilum is present, they are flush with the testa and do not possess an encircling aril. Furthermore, the length of the plumules measures approximately three quarters of the entire length of the cotyledon (Figure 6h). This identifies the pulse as Vigna radiata and not Vigna mungo.
Cotton (Gossypium sp.)
Three contexts (Trench 4, context #22, #30 and #31) contained fragments of cotton. A complete cotton seed was identified in Trench 4 (context #27) (Figure 7a). Although identification to species is difficult based on morphology, identification to genus is possible because the cotton fragments show distinctive long cells in the testa, a spongy interior and a combination of irregular short cells followed by regular long cells in the transect of the seed fragments (Fuller, 2008; Vaughan, 1970).

(a) Cotton seed from Trench 4 context# 27. (b) Archaeological Vigna unguiculata subsp. sesquipedalis compared with (c) a modern charred immature seed. Images: Cristina Castillo.
Other pulses
Three other pulses of economic importance were identified at Angkor Thom, pigeon pea (Cajanus cajan) (Trench 4, context #27), hyacinth bean (Lablab purpureus) (Trench 4, context #22) and yardlong bean (Vigna unguiculata subsp. sesquipedalis) (Trench 4, context #31) (Figure 7b and c). The three pulses were identified by examining their shape and size and compared with modern reference collections. Pigeon pea is flat in the proximal view and kidney-shaped, round or squarish in the lateral view. Hyacinth beans are oblong or reniformed with a long linear hilum and yardlong bean is long and thin.
Indian kapok or silk cotton tree (cf. Bombax/cf. Ceiba)
Another economic crop identified in Trench 4 (context #31) is a form of kapok, a tree known for its large pods that contain soft fibres formed from seed arils. Kapok includes various local species of Bombax and the pan-tropical species Ceiba pentandra. The fragments exhibit a palisade layer of long cells similar to Gossypium but are thicker overall (Vaughan, 1968).
Discussion
Plant consumption in Angkor Thom
The first analyses of botanical macroremains from 14th and 15th century Cambodia can be supplemented with evidence from texts and ethnography. The paucity of historical period archaeobotanical studies in Southeast Asia means that some of the macroremains in this study are the next known after an early introduction in Peninsular Thailand. Therefore, the archaeobotanical evidence from Angkor Thom can contribute to the history of food and cultivation in mainland Southeast Asia.
During the Angkorian period, rice from irrigated ricefields (Old Khmer sre) was the most consumed cereal. Sesame (Old Khmer lṅo) and several species of beans and peas (Old Khmer santek (generic term)), all cultivated in dry fields (Old Khmer caṃkā), were complementary food products. These plants formed the basis of vegetal nutrition at Angkor (Dagens, 2003). Correspondingly, rice, sesame, mungbean, yardlong bean, hyacinth bean and pigeon pea were found in our excavation assemblage.
Rice
The present study identifies abundant domesticated rice. The most plentiful plant remains were processing waste. We suggest that whole unhusked rice spikelets or paddy (Old Khmer sru) were transported to the site, dehusked (Old Khmer raṅko) and eaten. Thirteenth-century Chinese emissary Zhou Daguan described rice processing with mortar and pestle (Pelliot, 1951 (30. Les ustensiles)). Rice weeds found in the assemblage were probably discarded during winnowing (Castillo, 2013).
The small quantity of wild rice found might be the result of cultivation and/or hybridisation. The cultivation of floating wild rice (Old Khmer sraṅe) in the Angkor region is attested by inscriptions (Pou, 2004; Pou and Martin, 1981) and the account of Zhou Daguan (Pelliot, 1951 (17. Agriculture); see also Delvert, 1961). Alternatively, wild rice is a persistent weed in cultivated rice, and genetic studies indicate wild rices have arisen from regional crops by re-evolving wild seed dispersal (with wild-type shattering spikelet bases) and adaptations to cultivation (Huang et al., 2017; Thurber et al., 2013; Zhang et al., 2012). In some cases, there may be introgression of regional old populations, but the presence of weedy rice indicates that permanent and intensive wet rice cultivation provided an ecological niche where weeds adapted.
The transition from japonica to indica rice in mainland Southeast Asia occurred after the prehistoric period, although details regarding this shift are unknown. It has been hypothesised that indica rice arrived in mainland Southeast Asia after the initial contact with South Asia (c. 7th–9th centuries CE) (Castillo, 2011; Castillo et al., 2016b; Fuller et al., 2010b). While there is a scarcity of Southeast Asian archaeobotanical rice from the 7th century onwards, the samples from Angkor Thom demonstrate that the indica subspecies was certainly established by at least by the 14th and 15th centuries.
Comparing grain sizes (L/W ratios) can clarify the japonica to indica shift (Figures 5b and 6a and b). Rice grains from Thai prehistoric sites of Khao Sam Kaeo, Phu Khao Thong, Ban Non Wat and Noen U-Loke (c. 1000 BCE to c. 500 CE) are short and plump, fitting the description of japonica-type rice (L/W ratios <2). Ancient DNA analysis of the rice from these four prehistoric sites confirmed japonica (Castillo et al., 2016b). Whereas the L/W ratios of the Angkor Thom samples are long and thin, corresponding with the indica subspecies. Conversely, we also observe awned rice in the samples. While awnless types have arisen in both japonica and indica subspecies, awned varieties are typically primitive and might indicate persistence of ancient japonica varieties.
Sesame
Sesame is an important component of ceremonial offerings to gods, spirits and ancestors during the Angkorian period (Cœdès, 1953 (K. 352, K. 451); Mikaelian, 2007; Pou, 2004, and below), but its consumption as a foodstuff is unclear. In contemporary Cambodia, sesame oil is extracted and used in the local food and pharmacopoeia; they serve, in particular, to coat cakes (Vidal et al., 1969).
Sesame samples from our excavations are the next known in Southeast Asia after introduction in Peninsular Thailand. Sesame was domesticated in the greater Indus region of South Asia (Bedigian, 2004; Weisskopf and Fuller, 2014). In Southeast Asia, the earliest documented sesame is from the site of Phu Khao Thong in Peninsular Thailand (c. 1000 BCE) (Castillo et al., 2016a). Faced with limited data, we cannot provide a hypothesis on the introduction of sesame into Cambodia.
Mungbean
While the mungbeans identified in Trench 5 are from a ritual context, they may have played a more important dietary role than the other pulses at Angkor. Today, mungbean is an important pulse grown in Cambodia (Modern Khmer saṇṭaek pāy) (Dy Phon, 2000; Vidal et al., 1969). Archaeologically, it is the most visible of all pulses analysed in Southeast Asia (Calo et al., 2015; Castillo, 2015; Castillo et al., 2016a). Mungbean is a domesticate that originated in South Asia (Fuller and Harvey, 2006). This pulse was first introduced to Southeast Asia in the late first millennium BCE through exchanges with India and has been found in several sites, including Khao Sam Kaeo and Phu Khao Thong in Thailand, and Bali in Indonesia (Calo et al., 2015; Castillo et al., 2016a). Complementary research has identified mungbean at the Angkorian site of Preah Khan of Kompong Svay (Bakan) (Castillo, 2015).
Cotton
Based on historical and ethnographic evidence, we propose the species of cotton from our excavations is Gossypium arboreum, or tree cotton. The use of cotton (Old Khmer krapās) in the Khmer kingdom is documented by inscriptions (Pou, 2004; Pou and Martin, 1981), and seeds might provide evidence of a textile industry. Cotton seeds could indicate on-site processing of the fibre into thread. However, we note Zhou Daguan’s observation that Angkorians did not know how to spin thread with a wheel and made their textile filaments by hand (Pelliot, 1951 (29. Les vers à soie et le mûrier); see also Harris, 2007). A feasible deduction is that Angkorians used tree cotton (Gossypium arboreum) that has short fibres and is challenging to spin. Zhou Daguan also recorded that ‘wild’, or ethnic minorities, living near to Angkor cultivated the cotton tree, and weaved products he deemed to be ‘coarse’ (Pelliot, 1951 (11. Les sauvages); see also Groslier, 1921). This activity may have continued for several centuries. At the end of the 19th century, Jean Moura observed cotton among the products cultivated by the Samre or Pear ethnic minority who once lived in the Angkor region (Moura, 1883 (I); see also Baradat, 1941). Also in the 19th century, Clovis Thorel (2001 [1873]) observed that Gossypium arboreum was planted in hedges and around the fields of the Mekong Basin.
Evidence of cotton in Southeast Asian sites is limited to a few seed and fibre fragments or a few complete seeds. In combination with samples recently found at the Angkorian site of Preah Khan of Kompong Svay (Bakan) (Castillo, 2015), these fragments are the first identified in mainland Southeast Asia during the historic period. Cotton tree originated in the Indus region and thought to have been widely grown during the Harappan period, eventually dispersing to other parts of India by c. 500 CE (Fuller, 2008). The earliest archaeological evidence of cotton in Southeast Asia dates to the first millennium BCE from Ban Don Ta Phet and Khao Sam Kaeo in Thailand (Cameron, 2010; Castillo et al., 2016a; Castillo and Fuller, 2010). Based on the strong links these sites have with India, the cotton found is identified as Gossypium arboreum.
Other pulses
Yardlong bean (Vigna unguiculata subsp. sesquipedalis), hyacinth bean (Lablab purpureus), and pigeon pea (Cajanus cajan) are widely grown and eaten in East and Southeast Asia. Yardlong or ‘asparagus bean’ (Modern Khmer saṇṭaek drīṅ) is primarily cultivated for its green pods, which are eaten as a vegetable in numerous dishes (Dy Phon, 2000). Similarly, the hyacinth bean is favoured for its pods as a green vegetable, throughout North and East India, China, Southeast Asia and Cambodia (Modern Khmer saṇṭaek hoḷān’ tāv) (Dy Phon, 2000; Smartt, 1990). Pigeon pea is eaten as a pulse and vegetable in Southeast Asia, including ripe and immature seeds, leaves, young pods and sprouts (Van Wyk, 2005), but roasted seeds are also eaten (Van der Maesen, 1995). In Cambodia, pigeon pea (Modern Khmer saṇṭaek kliṅṭ or saṇṭaek grāp’ sa) is often grown near houses to make fences. As well as food and fodder, it is used as a medicine (Dy Phon, 2000).
These pulses originated in India and Africa (Fuller and Harvey, 2006). Pigeon pea is a South Asian domesticate brought into Southeast Asia together with the East African domesticate, hyacinth bean, during the initial period of contact between India and Southeast Asia in the second half of the first millennium BCE (Castillo, 2013). However, this is the first evidence of the African Vigna unguiculata at an archaeological site in Southeast Asia.
Kapok
The kapok samples are the first published from an archaeological context in Southeast Asia, although we cannot be certain of the genus. It is possible that kapok specimens are a regional species of Bombax, such as Bombax ceiba (syn. Bombax malabaricum) attested in Angkorian epigraphy (Old Khmer rkā; Pou, 2004; Pou and Martin, 1981) and by Zhou Daguan (Pelliot, 1951 (25. Les légumes); see also Groslier, 1921; Harris, 2007). Bombax features in Angkorian pollen and spore assemblages of the Bakong temple moat and a Phnom Kulen Plateau reservoir (Penny et al., 2005, 2014). Another Cambodian archaeological site, Preah Khan of Kompong Svay (Bakan), yielded fragments tentatively identified as cf. Bombax/cf. Ceiba testa from mid-12th-century layers (Castillo, 2015).
In contemporary Southeast Asia, there are several Bombax species, which produce similar seedpods containing soft fibres. The most widespread is Bombax ceiba, known as the Indian silk cotton tree, which is native throughout India in moist deciduous zones and adjacent dry deciduous zones (Asouti and Fuller, 2008), and extending through mainland Southeast Asia and tropical South China (Robyns, 1963). Local to the region are Bombax albidum, reported as endemic to South Vietnam; Bombax valetonii found through Thailand, Malaysia and Indonesia; and Bombax anceps, which is found throughout mainland Southeast Asia, including Cambodia. The seedpods of Bombax species produce loose seed fibres or a kind of cotton suitable for stuffing of pillows or thick clothes.
Alternatively, Ceiba pentandra is a small tree planted all over Cambodia around villages. The light wood is occasionally used to make soles of sandals. Like Bombax, fibres that surround the seeds are used as stuffing. Additionally, the fibre is employed for rituals that honour the ancestors or spirits. The green seedpods of Ceiba pentandra are also occasionally consumed as a vegetable and the pods are employed in games for both children and adults (Vidal et al., 1969).
Consecration deposits at Angkor and plant materials
The botanical remains found in a lead container from Trench 5 probably had ritual significance. The association of rice with sesame and mungbean is particularly relevant in drawing this conclusion. These plants are listed together, usually in the same order, in a series of inscriptions in Old Khmer detailing offerings and provisions to Angkorian temples (Old Khmer sru raṅko lṅo santek, ‘paddy, husked rice, sesame, beans’; Pou, 2004). Mikaelian (2007) proposes that the association of these plants is a metaphoric representation of agrarian production in the Angkorian kingdom. The same plants also appear in Sanskrit epigraphy as essential elements of the pūjā or daily ritual. For example, in the late 12th-century Ta Prohm and Preah Khan steles, the plants are cited in the specific sequence as lists of donations (Sanskrit taṇḍula, ‘husked rice’; tila, ‘sesame’; mudga, ‘mungbean’; Cœdès, 1906, 1941). During the Early Modern and Modern periods, the same plants are prescribed for offerings in various ceremonies including those of royal consecration (Mikaelian, 2007), royal ploughing (Porée-Maspero and Commission des Mœurs et Coutumes du Cambodge, 1985a) and house consecration (Giteau, 1971; Porée-Maspero, 1961).
At Angkor, consecration deposits are typically associated with stone architecture (Boisselier, 1966; Pottier, 1997a, 1997b). Deposits were placed in foundations and cupola of central towers (Old Khmer prāsād), or below doorways and thresholds. Deposits of gold, silver, glass, carbonised or oxidised fragments of seeds were identified at the 11th-century Baphuon temple (Groslier, 1961; see also Pottier, 1997b). Another series of consecration deposits containing sesame seeds, leaves, gold, bronze, gems, silk and possibly resin were discovered at the nearby 12th- to 13th-century Terrace of the Elephants (Pottier, 1997a, 1997b, 1998)
It is problematic to directly link text and material culture, especially between disparate geographic regions and across time; however, a review of rituals described in Sanskrit technical manuals on temple building and iconography (the vāstu- and śilpa-śāstras, and certain purāṇa, c. 6th to c. 16th centuries CE) can provide useful analogues for the emplacement and contents of consecration deposits (Ślączka, 2007, 2011). Ślączka’s translation and analysis of the 11th- to 12th-century South Indian text, the Kāśyapaśilpa, may assist in appraising the character of the deposit. According to the ritual texts, the fundamental purpose of a consecration deposit is to bring prosperity to those who perform the ritual. The inclusion of life-giving seeds, the embryos and generators of wealth, underlines the association of consecration deposits with fertility. Rice and sesame are named numerous times in the Kāśyapaśilpa as requisite additions of fruitful deposits (Ślączka, 2007).
The consecration deposit found in Trench 5 may relate to a wooden building that has not been identified in the archaeological record. This building may have been part of a densely occupied area directly north of the Royal Palace, with a possible relation to the adjacent Buddhist monastery of Tep Pranam. The first construction phase of Tep Pranam may date to the reign of Jayavarman VII (r. 1182/1183 to c. 1220), or slightly after, however, the consecration deposit likely relates to a second phase beginning from the 14th century (Groslier, 1969, 1973 see also Tun, 2015). The deposit might have been interred in the performance of a ritual, perhaps like those observed by contemporary Cambodians, especially in association with the Kruṅ Bālī, owner of the land and water (Giteau, 1969, 1971; Porée-Maspero, 1961; Porée-Maspero and Commission des Mœurs et Coutumes du Cambodge, 1985b).
Implications and conclusions
Although the samples were collected during excavations to consider elite craft production, they relate to contexts deposited after the foundry and sculpture workshop had ceased operations (Polkinghorne et al., 2014). The study examined a small area within the enclosure of Angkor Thom and the number of sorted contexts is limited. However, the results are significant because they provide evidence of food items consumed by the inhabitants of the area, and plant materials used as fibre and ritual deposits. Low counts retain importance because macrobotanical remains in the tropics are typically poorly preserved because of extreme humidity, and exposure to weathering, erosion and bioturbation (Kealhofer and Grave, 1999; Piperno and Pearsall, 1998; Sobolik, 2003). Furthermore, it is unusual in Southeast Asia to find a consecration deposit rich in organic remains like that discovered in Trench 5.
While recent scholarship offers new evidence for demise (Buckley et al., 2010, 2014; Evans et al., 2013; Fletcher et al., 2008; Vickery, 1977) on the subject of continuity, there are only fragmentary studies. In the 14th and 15th centuries, there is evidence of limited modifications to architecture (Groslier, 2014 [1958], 1969; Polkinghorne et al., 2013; Pottier, 1997b; Thompson, 1998), trade with China and Southeast Asia (Dupoizat, 1999; Wade, 2011), sculpture production (Leroy et al., 2015; Polkinghorne et al., 2013; Tun, 2015) and partial occupation at Angkor Wat (Brotherson, 2015; Stark et al., 2015). The discovery and examination of seeds and grains from the 14th to the early 15th centuries indicate that Angkor was active and producing economic crops during the period of its decline.
Radiocarbon dates and diagnostic ceramics within the infill of the stone-lined and decorated sraḥ suggest that activities at this specific area associated with the Royal Palace of Angkor Thom changed terminus post quem from the 14th century. We hypothesise those responsible for its upkeep, logically the royal court and political administration who lived at or next to the Royal Palace, no longer commanded the resources necessary to maintain it or the need to use it. This may correspond to the movement or the emergence of a rival elite who had established their political centre in the region of Phnom Penh (Khin, 1988; Kitagawa, 2000; Vickery, 1977; Wolters, 1966). The proximity of the consecration deposit to the Buddhist monastery of Tep Pranam may explain the continuing significance of this area. Tep Pranam may be the first structure dedicated to Theravāda Buddhism at Angkor. Its location, size and composition indicate it was extremely well resourced (Groslier, 1973). Consecration deposits are a key feature of contemporary Cambodian Theravāda practice, and the deposit described above may relate to persistence of occupation and activity of the Tep Pranam saṅgha or lay community in the early 15th century.
A fall or shift in the political centre of Angkor does not necessarily mean abandonment by the local population. There was a change in the function of the area but people continued to live there, cultivate crops and perform rituals. Continuity of these activities and occupation is demonstrated by the discovery of the archaeobotanical remains. While rice remained the most important cereal, Angkorians also consumed a suite of pulses, and employed plant materials in the function of ceremonies with historical and contemporary analogy. Our study has combined archaeological evidence with inscriptions, texts and ethnography to understand the character of 14th- to 15-century Angkorian life.
Footnotes
Appendix
| Identification | Trench 4 #22 | Trench 4 #27 | Trench 4 #30 | Trench 4 #31 |
|---|---|---|---|---|
| Litres of soil | 2.25 | 2.5 | 4 | 2 |
| Plant parts/litres of soil | 310.7 | 217.2 | 173.0 | 280.5 |
| Cereals | ||||
| Oryza sativa | 3 | |||
| Oryza sativa fragment | 7 | 3 | 3 | 11 |
| Oryza sativa spikelet base – domesticated | 9 | 113 | 95 | 112 |
| Oryza spikelet base – wild | 6 | 8 | ||
| Oryza spikelet base – indeterminate | 2 | 34 | 44 | 137 |
| Oryza lemma apex | 2 | 16 | 50 | 30 |
| Oryza charred embryo | 4 | 12 | 26 | 13 |
| Oryza awn – silicified | 8 | |||
| Total Oryza | 24 | 184 | 234 | 306 |
| Pulses | ||||
| Cajanus | 1 | |||
| Lablab | 1 | |||
| Vigna radiata | 1 | |||
| Vigna unguiculata subsp. sesquipedalis | 1 | 1 | ||
| Total pulses | 2 | 0 | 0 | 2 |
| Economic crops | ||||
| cf. Bombax/cf. Ceiba pentandra | 3 | |||
| Gossypium sp. | 1 | |||
| Gossypium sp. fragment | 2 | |||
| cf. Gossypium | 2 | |||
| Sesamum indicum | 1 | |||
| Total economic crops | 5 | 1 | 0 | 3 |
| Other possible economic crops | ||||
| Phyllanthus sp. | 1 | 2 | ||
| Type 44 – Canarium fragment | 2 | |||
| Type 48 – endocarp poss. Baobab | 1 | 1 | ||
| Total | 1 | 1 | 5 | 0 |
| Wild and weedy | ||||
| Acmella paniculata | 3 | 4 | 8 | |
| Alternanthera cf. sessilis | 1 | |||
| Carex sp. peryginium | 28 | |||
| Cenchrus sp. involucre | 1 | |||
| Cleome viscosa | 1 | |||
| Limnophila villosa | 2 | |||
| Lindernia crustacea | 2 | |||
| Mollugo cf. pentaphylla | 12 | |||
| Pennisetum sp. | 2 | |||
| Wild rice (maybe O. rufipogon) | 1 | |||
| Portulaca oleracea | 2 | |||
| cf. Sagina japonica | 12 | |||
| Sida cf. rhombifolia | 1 | |||
| Total wild and weedy | 9 | 8 | 21 | 42 |
| Others | ||||
| Amaranthaceae type A | 1 | |||
| Fabaceae type E | 1 | |||
| Fagaceae/Betulacreae nut shell frag | 1 | 2 | ||
| Cyperaceae type B | 3 | |||
| Cyperaceae – inner cotyledon | 2 | 1 | ||
| Cyperaceae type C – sedge culm | 22 | |||
| Poaceae type G – grass culm node | 6 | 14 | 5 | |
| Poaceae spp. | 4 | |||
| Ranunculaceae | 2 | |||
| Scrophulariaceae type A | 1 | |||
| Poaceae rachilla (non-rice) 0.25–0.5 mm | 4 | |||
| Type 23 – pedicel | 1 | |||
| Type 49 – base of capsule | 1 | |||
| Type 50 – node | 1 | |||
| Type 63 – Nymphaceae fragment | 2 | |||
| Parenchyma – prob. rice | 18 | 35 | ||
| Mesocarp fragment/fruit flesh | 1 | |||
| Unidentified | 391 | 4 | 6 | 0 |
| Unidentified fragments | 260 | 306 | 388 | 163 |
| Total others | 658 | 348 | 432 | 208 |
| NSP | 720 | 611 | 927 | 877 |
| NISP | 699 | 543 | 692 | 561 |
Samples have been interpolated using percentages of amounts sampled. Rice husk was present in all samples but are not included in the table.
Acknowledgements
Thanks to HE Bun Narith, HE Ros Borath, Mao Lor, Tin Tina, An Sopheap, Chhay Rachna, Tho Thon, So Malay, Janet Douglas, Zuzana Faltyskova, Gill Green, Christophe Pottier, Miriam Stark, and Mark Thomas. Special thanks to the fieldwork team: Chunn Sambor, Moul Komnet, Vitou Phirom, Kang Samouen, Suy Pov, Voeurn Vann, Hen Chenda, Lugh Lay, Shaun Mackey, David Brotherson, Sally MacLennan, students from the Royal University of Fine Arts, Phnom Penh, and dedicated workers. Excavations at the Royal Palace foundry continues under a joint initiative of the APSARA National Authority and the École française d’Extrême-Orient. Martin Polkinghorne is an Honorary Associate of the Department of Asian Studies, University of Sydney. We would also like to thank John Watson for editing the manuscript.
Funding
This research was funded by an Australian Research Council Discovery Project Grant (DP110101968) and the National Geographic Society/Waitt Grants Program (#W122-10). The archaeobotanical research was supported by the Natural Environment Research Council (NERC) Grant # NE/N010957/1. Funding for the 14C accelerator mass spectrometry (AMS) analysis was provided by the École française d’Extrême-Orient. Project collaborators included The University of Sydney; University College London (UCL), Institute of Archaeology; the APSARA National Authority; the École française d’Extrême-Orient; the National Museum of Cambodia; the Freer and Sackler Galleries; Smithsonian Institution; the Metropolitan Museum of Art; the Centre de recherche et de restauration des musées de France; and the Institut national de recherches archéologiques préventives.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.