
Abstract
Various studies using pollen stratigraphies have demonstrated significant correlations between Holocene plant diversity, climate, and human activities. Studies that have analyzed longer Holocene timescales tend to discuss cultural data very superficially. This is remarkable because detailed insights into past human activities may be key to gain an understanding of the observed trends in biodiversity. This study aims to reconstruct and explain spatio-temporal trends in past plant diversity (alpha, temporal, and spatial beta diversity) by integrating data on vegetation dynamics, human subsistence economy, and land-use patterns. The landscape of Northwest France during the greater part of the Iron Age and the start of the Roman period (600 BC–AD 100) is selected as a case study. In total, 30 high-quality pollen-stratigraphical sequences allow for the reconstruction of the main long-term trends in plant diversity and more generally of the changing fabric of the landscape. Additionally, increasingly detailed images of the Iron Age rural landscape are available because of a steep increase in archaeological data (aerial photography, surveys, and excavations). These different types of data are integrated and used as input for a wider discussion on the relation between human activities and plant diversity. In general, the taxonomic richness increases steadily during the period under study. Some spatio-temporal differences are observed. The increasing richness values correspond with the growing impact of human activities on the landscape. Archaeologically documented land-use changes on smaller timescales are less clearly reflected in the richness values and vegetation dynamics, which might result from the (large-scale) research design.
Keywords
Introduction
Since the early 1990s, the preservation of biodiversity has been one of the top priorities of the global environmental agenda (e.g. Blondel, 2005; Brown and Caseldine, 1999). Paleoecology has made an important contribution to the understanding of biodiversity at several levels (Odgaard, 1999). Various studies using pollen stratigraphies have demonstrated significant correlations between Holocene plant diversity, climate, and human activities. It is widely accepted that, to a certain extent, biodiversity and human interference are positively linked (e.g. Birks and Line, 1992; Grime, 2001; Paillet et al., 2010; Peet et al., 1983). Recent research into these relations mostly focuses on the last 200 years (Birks et al., 2016; McGill et al., 2015). Studies that have analyzed longer Holocene timescales (e.g. Berglund et al., 2008; Birks, 2007; Birks and Line, 1992; Birks et al., 1988, 2015, 2016; Colombaroli et al., 2013; Felde et al., 2016; Goring et al., 2013; Lindbladh, 1999; Lindbladh and Bradshaw, 1998; Matthias et al., 2015; Meltsov et al., 2011, 2013; Odgaard, 1994, 1999, 2001, 2013; Weng et al., 2006) tend to discuss cultural data very superficially – if they are even introduced at all. This is remarkable, because these data in particular may be key to gaining an understanding of the observed trends in biodiversity. The currently dominant monodisciplinary approach hampers detailed assessments of the links between past human behavior and biodiversity.
This study aims to reconstruct and explain spatio-temporal trends in past plant diversity by integrating data on vegetation dynamics, human subsistence economy, and land-use patterns. The landscape of Northwest France during the greater part of the Iron Age and the initial phase of the Roman period (600 BC–AD 100) is selected as a case study. Compared to many other parts of Northwest Europe, this area is rich in paleobotanical data. These allow for the reconstruction of the main long-term trends in plant diversity and more generally of the changing fabric of the landscape. At the same time, increasingly detailed images of the Iron Age rural landscape are available because of a steep increase in archaeological data (aerial photography, surveys, and excavations). These point to significant changes in land-use history during the period under study. The different types of data are integrated and used as input for a wider discussion on the relation between human agency and plant diversity.
The research questions addressed in the paper are as follows:
What are the spatio-temporal trends in plant diversity and vegetation dynamics in Northwest France between 600 BC and AD 100?
How do these trends relate to habitation development, land-use patterns, and subsistence economy?
What bearing do these results have on wider discussions on the relation between human activities and plant diversity?
Context and problem definition
Climate change and human impact are essential in the development of ecosystems and their biodiversity over different temporal and spatial scales (Berglund et al., 2008; Ricklefs, 1987). Reasons for maintaining and enhancing biodiversity include its value as an indicator of environmental quality and the preservation of a gene base. Systems with higher biodiversity may also be more stable and resilient and have a higher productivity (Brown and Caseldine, 1999: 3; Chapin et al., 2000; Tilman et al., 2006). According to Odgaard (1999), paleoecology can contribute to the understanding of biodiversity by (1) providing estimates of ‘base-line’ levels of species richness under ‘natural’ conditions, (2) describing diversity aspects of long-term succession, (3) using paired study sites to reveal effects of long-term land-use changes, and (4) supplying historical backgrounds to understand diversity variation. In short, the study of pollen records may lead to better insights into the processes influencing long-term trends in plant diversity (cf. Berglund et al., 2008; Brown and Caseldine, 1999; Birks and Line, 1992; Lindbladh, 1999).
The basic component of biodiversity is species richness, which is the number of species (Odgaard, 1999). Another important aspect of diversity is evenness or equitability, describing the distribution of individuals among the taxa (the relative frequency of taxa; Birks and Line, 1992; Meltsov et al., 2013). Palynological richness in a pollen count is one proxy for reconstructing the alpha diversity trend through time (McGill et al., 2015; Weng et al., 2006). In this study, we are also interested in the changes in assemblage composition across space for one time period (spatial beta diversity; Felde et al., 2016). Finally, the time series of reconstructions clearly reflect the biodiversity at landscape level (Gamma diversity; Odgaard, 2013).
Studies in this field have significantly increased after the introduction of the rarefaction technique in past plant diversity research by Birks and Line (1992; see below). Even though the species richness in pollen records is biased by various factors (see below), rarefaction estimates are generally accepted as roughly reliable reflections of trends in past floristic richness. So far, most studies have centered on Scandinavia (e.g. Berglund et al., 2008; Lindbladh, 1999; Odgaard, 1994; Seppä, 1998), Great Britain (e.g. Bunting, 1996; Fossitt, 1996; Skinner and Brown, 1999), and Ireland (e.g. Brown, 1999), but other regions are receiving attention as well (e.g. Colombaroli et al., 2013: Switzerland; Meltsov et al., 2013: Estonia). A first study into the palynological richness of Northwest France has been undertaken by Marguerie (2009).
Various studies of pollen records indicate a positive correlation between human impact and taxa richness (e.g. Balée, 2006; Birks and Line, 1992; Birks et al., 2015, 2016; Huston, 1994; Peet et al., 1983). An increase in pollen percentages of anthropogenic indicators is in most cases closely connected with a rise in the number of taxa recorded (Lindbladh, 1999). The ‘intermediate disturbance’ hypothesis (Connell, 1978; Rosenzweig, 1997; Wilkinson, 1999) states that diversity is higher when disturbances are intermediate on the scales of frequency and intensity. It is based on the assumption that moderate disturbances interrupt and set back the process of interspecific competition and, therefore, maintain a nonequilibrium state. This disturbance level is sufficient to prevent the development of dominance by any single component but insufficient to cause extinction of components at a landscape scale (Birks and Line, 1992). At a wider spatial scale, human interferences may create new landscape units and, therefore, increase landscape, plant community, and species diversity (Berglund et al., 2008; Marguerie, 2009; Matthias et al., 2015; Meltsov et al., 2011, 2013; Miras et al., 2015; Odgaard, 1999). Pollen records generally demonstrate a steady increase in floristic richness after the introduction of agriculture in the Neolithic, but shorter periods of expansion or contraction can be detected. Berglund et al. (2008) describe significant shifts as ‘diversity events’. Birks and Line (1992) note that floristic richness today is often greatest at boundaries, particularly ecocline boundaries between vegetation types. A floristically rich landscape is thus often one with many component vegetation types and hence with many boundaries. This is also referred to as a highly heterogeneous landscape. Most pollen records demonstrate a decrease in floristic richness since the 18th/19th century AD, because of agricultural intensification and an increasing homogeneity in landscape structure (e.g. Birks et al., 2016; McGill et al., 2015).
So far, studies into the correlation between past human behavior and plant diversity have been rather one-dimensional. Fossil pollen records form the analytic starting point and are used to deduce human influence on past ecosystems. When cultural (archaeological and historical) data are brought into the picture, these discussions remain superficial and undetailed, especially if we go back further in time than the last few centuries. The heavy reliance on paleobotanical sources in reconstructing the degree of human disturbance is remarkable if we take into account the important role that is generally ascribed to this factor. This certainly applies to what in ecological research is referred to as the ‘Homo sapiens’ or ‘oligocratic’ phase, which is characterized by increasing late prehistoric and early historic human forest clearances and land-use changes (e.g. Birks et al., 2016). The lack of integration of high-resolution cultural data brings the danger of circular logic and hampers detailed assessments of the link between human interference and past plant diversity dynamics.
In this study, we analyze pollen records from 30 sites from four different regions within Northwest France and compare the obtained trends in floristic richness with archaeological data on a well-documented protohistoric period: the Iron Age and initial stage of the Roman period (600 BC–AD 100). During this time span, the cultural landscape underwent a series of changes linked to important socioeconomic transformations.
Research area
Northwest France protrudes into the Atlantic Ocean and forms the westernmost tip of the European continent (Figure 1). The historical rural habitation pattern is dispersed, consisting of isolated farmsteads and small hamlets. Productivist agriculture was introduced in the second half of the 20th century, replacing the ancient system of polyculture and cattle breeding (Menez and Lorho, 2013).
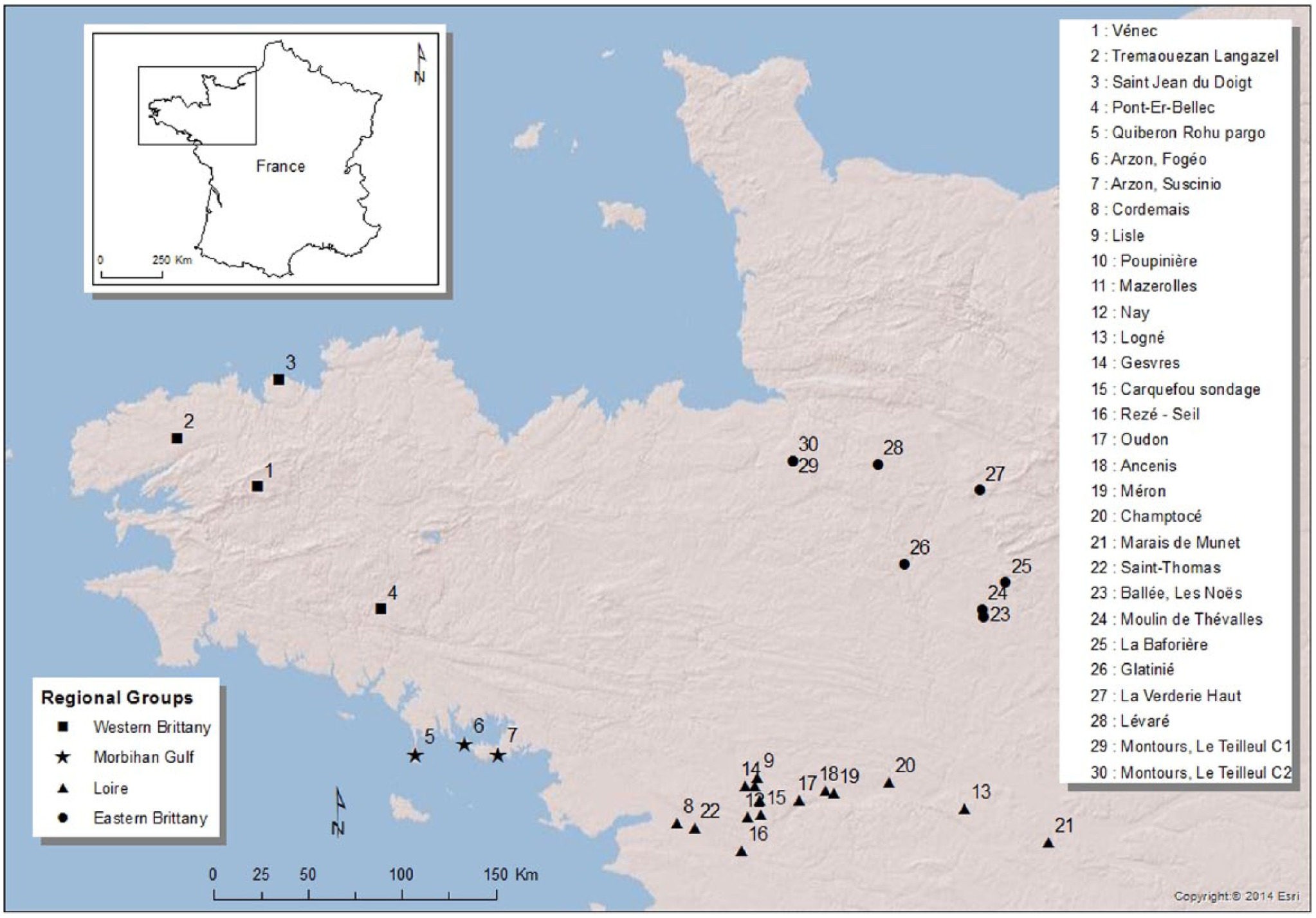
Location of the study area and distribution of the analyzed palynological datasets, clustered in four regions (GIS processing: Y. Rantier).
The landscape of western Gaul, on the eve of the Roman conquest, did not conform to the ‘mythical’ image of vast and impenetrable forests (Marguerie, 1990, 1992). The opposite is true: the later Iron Age landscape was relatively open, heavily influenced by man, and densely settled. Important overview studies of paleoecological research in Northwest France have been published by Marguerie (1992, 2009; Marguerie and Hunot, 2007), Gaudin (2004), and David (2014).
The numerous pollen data obtained at the Armorican Massif scale were used by David (2014) to estimate the regional vegetation by the REVEALS model of Sugita (2007). Sugita introduced the possibility of grouping multiple small-sized sites (radius = 20 m) as well, which is useful for the Northwest French situation where most basins are relatively small (Table 1).
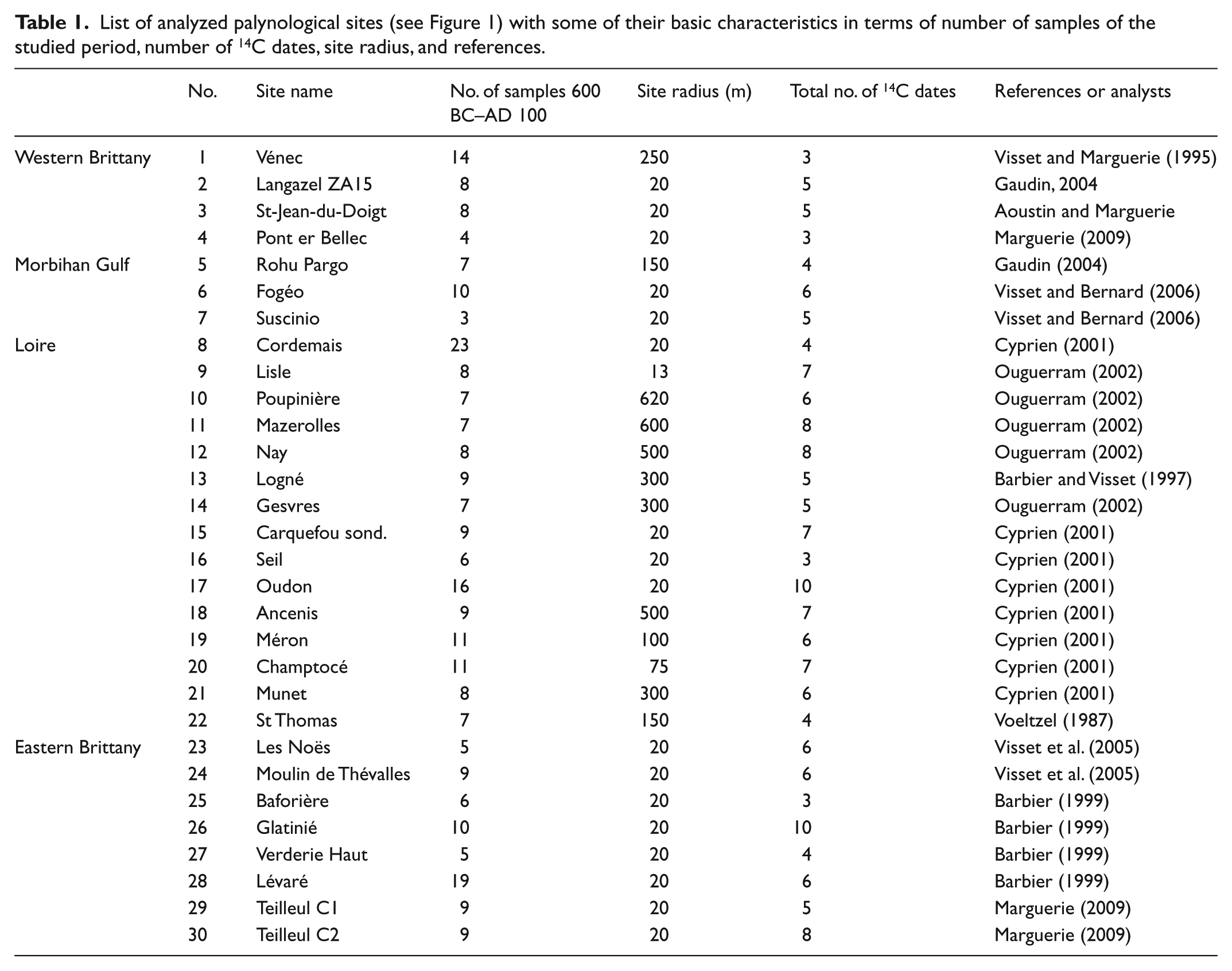
List of analyzed palynological sites (see Figure 1) with some of their basic characteristics in terms of number of samples of the studied period, number of 14C dates, site radius, and references.
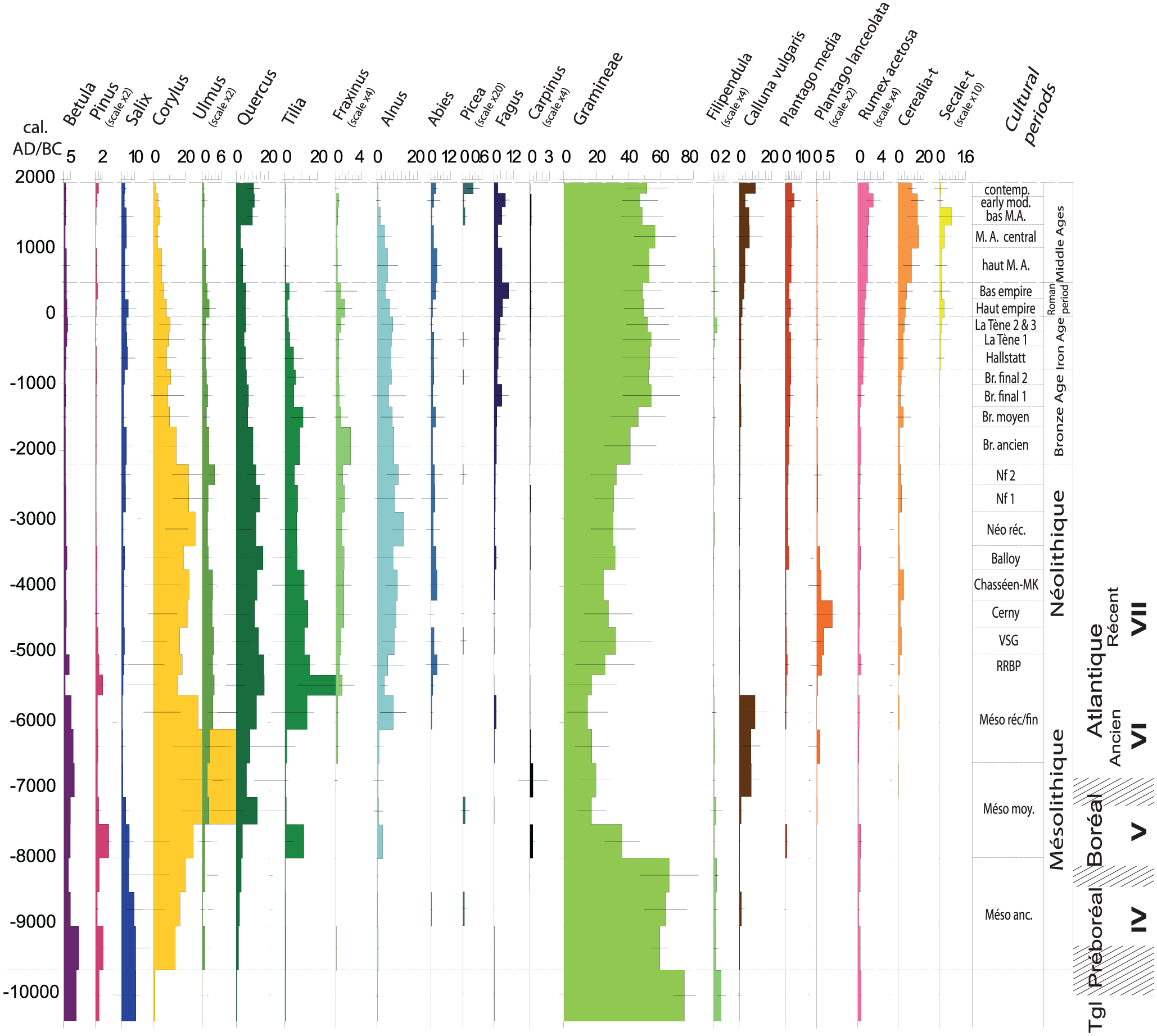
The sixth Regional Pollen Assemblage Zone (RPAZ VI) defined by David (2014) for Northwest France covers the period between the end of the Bronze Age and the end of Antiquity (1350 BC–AD 475). This period is characterized by a decline of the main ligneous taxa, except the beech tree, whereas Poaceae increase clearly (Figure 2). During the subperiod (RPAZ VIa) from the Late Bronze Age to La Tène 1 (1350–250 BC), Corylus and Alnus are in constant proportion whereas Tilia, Quercus, Ulmus, and Fraxinus decrease gradually. Fagus and Salix increase slightly. Poaceae are very well represented. The subzone (RPAZ VIb) from La Tène 2–3 to the end of the Roman period (250 BC–AD 475) is marked by some minor changes such as a small decrease in Corylus and Alnus. Ulmus, Fraxinus, and Fagus demonstrate a modest increase, whereas the decline of Quercus ends. Poaceae decrease slightly.
Estimated vegetation cover between the Bronze Age and Modern period in the Armorican Massif based on palynological analyses (REVEALS model). Modified after David (2014): (a) enclosures, (b) souterrains, (c) stelae, (d) tribes, (e) fortifications, and (f) salt production.
Historical sources indicate that later Iron Age Brittany was inhabited by the tribes of the Osismii, Coriosolites, Veneti, Redones, and Namnetes (e.g. Menez and Lorho, 2013). The introduction of systematic aerial photography to the region in the 1970s led to the discovery of thousands of protohistoric and early historic sites, which are generally recognized by their characteristic ditch systems (Langouët, 1991; Leroux et al., 1999; Figure 3). Detailed (micro-)regional studies have assessed the links between archaeological field data and aerial photographic information (e.g. Leroux et al., 1999; Naas, 1999). Since the early 1980s, the region has witnessed a steep increase in Iron Age settlement excavations (Menez and Lorho, 2013). These focused both on ‘inland’ areas (e.g. Le Bihan et al., 1990; Menez, 1993) and coastal regions (e.g. Daire, 2009; Daire et al., 2011; Hyvert and Le Bihan, 1990). The latter also harbor numerous salt production sites scattered along the coastline (Daire, 2003). As more attention was also given to other site types (e.g. cemeteries, cult sites, and infrastructure), a detailed image of the Iron Age cultural landscape has emerged.
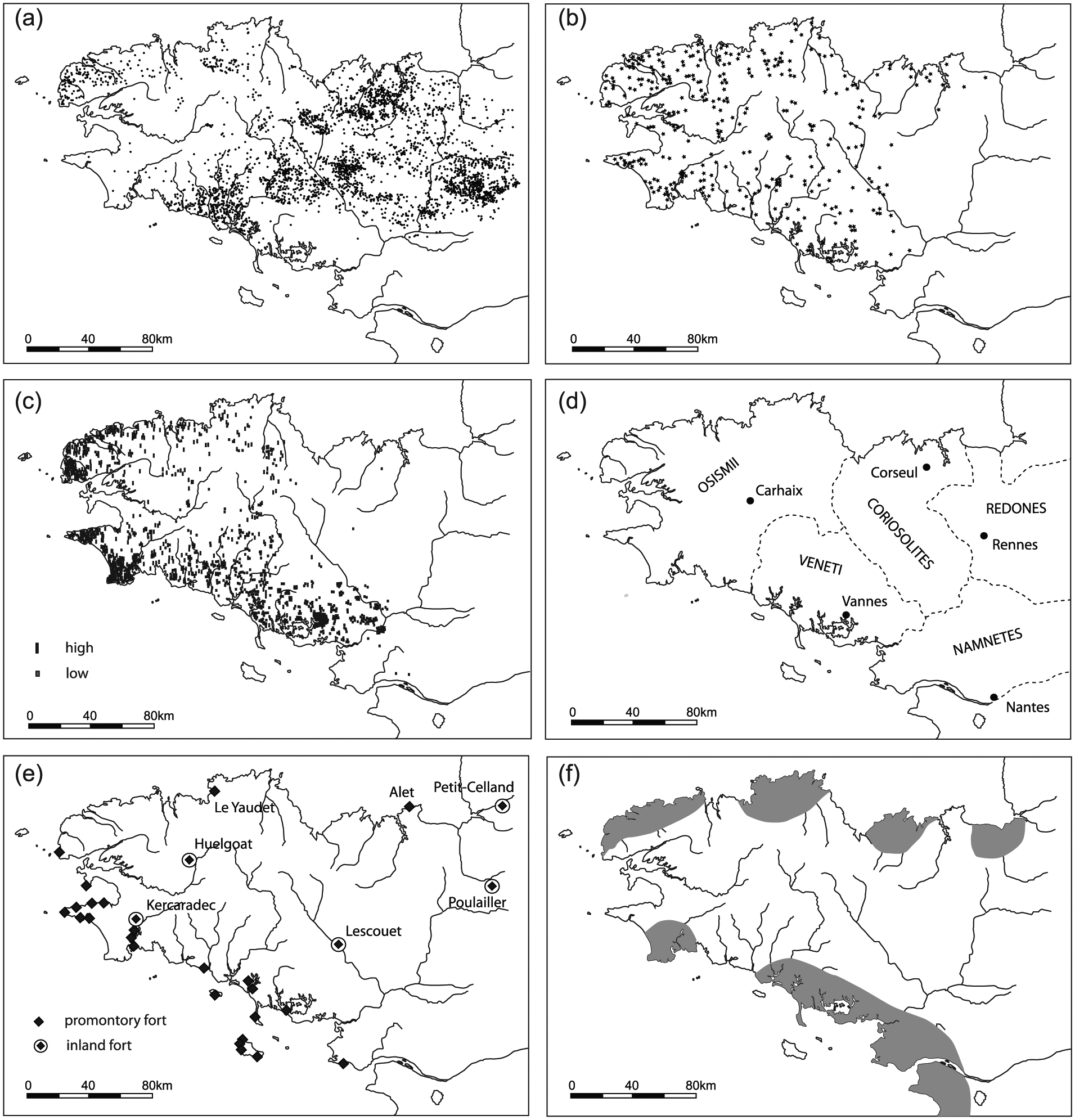
Distribution patterns of a selection of elements characterizing Iron Age Brittany: (a) enclosures (partly also dated to the Roman period, or undated; after Gautier, 2002), (b) souterrains (after Le Goffic, 2002a), (c) stelae (after Le Goffic, 2002b), (d) ‘tribal’ division according to historical sources (modified after Pape, 2002), (e) coastal promontory forts and large (>25 ha) inland fortifications (partly based on Henderson, 2007), and (f) primary salt production regions (after Daire, 2003).
Methodology
Contrary to previous research, our study takes human land-use patterns as a starting point. We are mainly interested in the changing ‘fabric’ of the Northwest French landscape between 600 BC and AD 100. Therefore, the most important trends in land use, landscape organization, and subsistence economy were reconstructed by means of a detailed survey of archaeological literature. A wide variety of sources was consulted, covering different components of the Iron Age cultural landscape (e.g. settlements, cemeteries, cult sites, fortifications, field systems, and infrastructure). The compiled data allow a division in three successive phases with markedly different characteristics. There are currently no indications for significant spatial differences in habitation history on a regional scale (e.g. habitation density) that could be of major influence on taxonomic richness estimates, even though the subsistence economy of coastal and inland settlements may have differed to some extent.
Of all palynological analyses in the Armorican peninsula, a selection of 30 well-dated and high-quality pollen-stratigraphical sequences was made. The Iron Age period is included in each of them. Most span larger parts of the Holocene. The sites cluster in four regions: 4 in Western Brittany, 3 in the Morbihan Gulf region, 15 in the Loire region, and 8 in Eastern Brittany (Figure 1). This regional division allows us to trace possible spatial variation in palynological richness. At this point, not enough high-quality data are available for Central Brittany. Therefore, this region has been left out of the research.
All existing data for the 30 sites were reanalyzed for the present research and studied integrally. The number of palynologically analyzed levels dating between BC 600 and AD 100 varies between 3 and 23 per site, with an average of about 9 and a total number of 272. All sampling contexts consist of small peat bogs with an average diameter of 140 m (Table 1). Samples from small basins have the advantage of recording vegetation at both local and regional scales (e.g. Broström, 2002). Pollen landing on the peat surface was not transported further. These kinds of sites have a chance of recording pollen from herbaceous taxa including anthropogenic indicators (Lindbladh, 1999).
A total of 172 radiocarbon dates, generally on seeds or twig fragments, were obtained for all the pollen sequences used in this study. The mean number of radiocarbon dates for each core is 5.7 (minimum = 3; maximum = 10; Table 1). Each 14C date was calibrated to calendar years BC/AD using CALIB4.3 (Stuiver and Reimer, 1993) and the IntCal98 calibration dataset (Stuiver et al., 1998). Problematic for this study is the so-called Hallstatt-plateau in the radiocarbon calibration curve between c. 800 and 400 BC, decreasing the precision of dates (e.g. Hamilton et al., 2015). Therefore, some caution is needed when links are assessed between vegetation trends and archaeological data.
Age–depth models were constructed for each sequence based on the available datings distributed throughout the core. When the pollen stratigraphy only had the minimum number of 14C datings (= 3), the model used was CLAM (Blaauw, 2010) with a linear regression interpolation. If the number of datings was higher than three, RenCurve was used (Lanos, 2004; Lanos and Philippe, in press) with a cubic spline interpolation and Bayesian approach (David, 2014).
Rarefaction analyses were performed on all pollen sequences. This technique permits to detect temporal changes in palynological richness (the number of pollen types per sample) which can be interpreted as a measure of floristic and landscape diversity (Berglund et al., 2008).
Palynological richness (among others) depends on the number of grains counted. The rarefaction analysis puts forward an exactly equal pollen count which is usually the lowest value of all sequences. With the ‘expected number’ E(Tn) of taxa independent of the number of counted pollen grains obtained, the comparison between levels of a set of samples is meaningful (Birks and Line, 1992). For the first time, rarefaction analysis was performed for a large number of western French Holocene pollen sequences. The software used was ‘Past’ (diversity/individual rarefaction).
Additionally, an analysis was made of the development of a selection of vegetation groups represented in Armorican pollen records during the period under study. These data serve to contextualize the results of the rarefaction analyses and to assess the links with human activity as reflected in archaeological data.
In total, five of the most common and relevant habitats were selected, based on the pollen-based vegetation community types (Leroyer et al., 2009). The first two represent forests. Closed forest is a mixed temperate broad-leaved forest, characterized by the presence of Quercus, Fagus, Carpinus, Tilia, Fraxinus, and Ilex. Open forest results from woodland clearance or use for agro-pastoralism and is characterized by heliophilous taxa such as Corylus, Betula, Acer, and Ulmus.
Among the open communities, three anthropogenic groups of light-demanding plants are defined. They indicate agro-pastoral activities and are associated with apophytes, native plants colonizing disturbed land or abandoned fields, and anthropochores, nonnative plants related to cultivation (e.g. Behre, 1986; Gaillard et al., 1992). Ruderals grow on disturbed and phosphorus-enriched soils in inhabited areas and pastures and include apophyte taxa such as Plantago, Convolvulus arvensis, Polygonum aviculare, Cuscuta, Brassicaceae, Chenopodiaceae, Caryophyllaceae, Artemisia, Ranunculaceae, Potentilla, Trifolium, Urticaceae, Lamiaceae, Alchemilla, and Centaureae. Adventive taxa grow within the confines of cultivated plants in arable fields: Centaurea cyanus, Papaver, Melampyrum, Rumex, and Mercurialis annua. The anthropochore ligneous and herbaceous taxa of the crops group include Castanea, Juglans, Cerealia, Secale, Linum, Fagopyrum, Vicia, and Cannabis. Pollen values were calculated as the percentage of total pollen sum excluding Alnus pollen and fern spores.
Pitfalls and premises
Archaeological biases
Despite the steep increase in Iron Age data, there is still a shortage of high-quality information on land-use patterns on different spatial scales. The main focus has been on settlements and less on the cultural landscape. This study integrates different types of archaeological data to sketch a holistic image. Because of a poor preservation of organic material, not much is known yet about agricultural practices.
Pollen analysis biases
As discussed at length in previous research, the interpretation of pollen data in terms of past floristic richness is not unproblematic (Brown, 1999; Brown and Caseldine, 1999; Birks et al., 2015; Felde et al., 2016; Goring et al., 2013; Matthias et al., 2015; Lindbladh, 1999; Odgaard, 1999, 2001, 2013; Weng et al., 2006). The relationship between the number of pollen types in an assemblage and the number of plant species around a site is complex and biased by many factors, such as the lack of taxonomic precision, differential pollen production and dispersal, and the sample size.
Some pollen types can only be identified at a genus or even family level. As families may consist of a large number of species, this leads to a considerable reduction in pollen richness values. However, the taxonomic precision might not limit the pollen-based diversity approach (Matthias et al., 2015). The pollen studies used here come from two laboratories (POLEN and ArcheoSciences) which use the same referential of taxonomy, follow identical published illustrations and keys (Beug, 2004; Faegri and Iversen, 1989; Reille, 1992), and share their reference collection.
According to Matthias et al. (2015), the bias related to production and dispersal may not inhibit a correspondence between landscape diversity around a site and pollen diversity in a surface sample.
Finally, when various pollen samples are combined or compared, it is almost inevitable that different pollen counts are represented. This is corrected for by rarefaction analysis (Birks and Line, 1992; see above).
Results: Iron Age habitation and land-use patterns
Macroregional trends
Iron Age Northwest France had a distinct character, marked by the distribution of specific cultural elements (e.g. promontory forts, stone stelae, and so-called souterrains used for storage) and artifact styles (Henderson, 2007; Figure 3). The Iron Age is characterized by a significant increase in settlement density (e.g. Giot, 1995; Le Bihan et al., 1990; Menez and Lorho, 2013). This observation is based on the dense distribution patterns of souterrains, stone stelae, and settlement sites. Giot (1995) estimates the population of Brittany in the last centuries BC at between 150.000 and 300.000 inhabitants, corresponding with a density of about 3–10 persons/km2. Small rural farmsteads, generally enclosed by ditches, were the dominant settlement type.
Diachronic patterns
Many archaeologists note marked differences between the First (800–450 BC) and Second (450–50 BC) Iron Age (e.g. Giot, 1995; Henderson, 2007). Remains of First Iron Age settlements are rare and generally consist of diffuse distributions of settlement refuse, reflecting a dynamic settlement system (see below; Menez and Lorho, 2013). By contrast, Second Iron Age settlements are abundant and easily recognizable. The main diachronic trends in settlement foundation and abandonment have recently been analyzed by Menez and Lorho (2013; Figure 4). A first small peak in settlement foundations appeared around 550 BC (Figure 5). In the late 5th and early 4th century, various settlements and cemeteries were abandoned. In the late 4th and early 3rd century, a recovery sets in, which accelerated greatly until the middle of the 1st century BC. Many settlements were abandoned in the decades following the Gallic Wars (58–51 BC), followed by a renewed expansion in the 1st century AD.
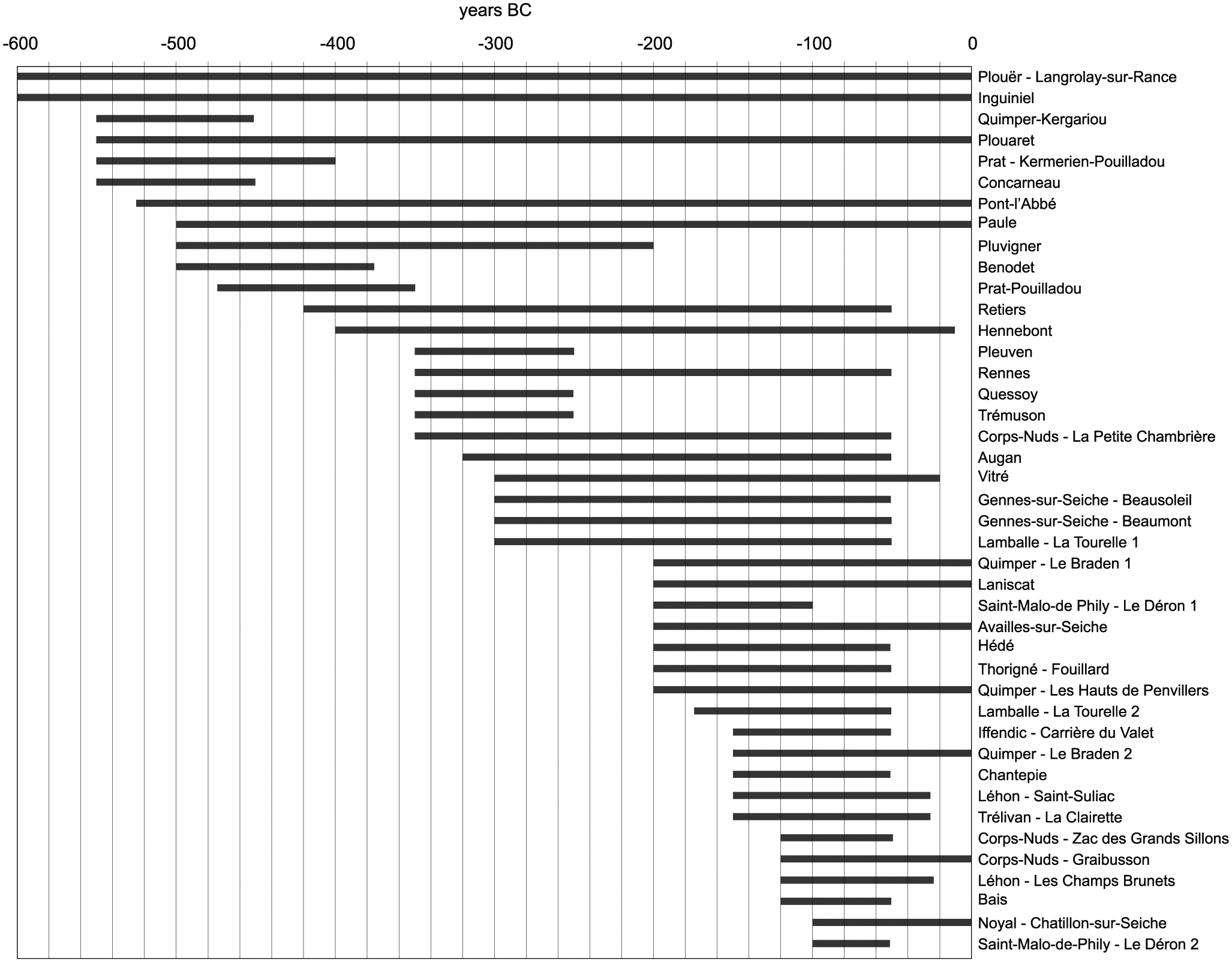
‘Life spans’ of Iron Age settlements in Brittany with high-resolution excavation data (e.g. large-scale research, 14C dates, detailed studies of find materials; after Menez and Lorho, 2013).
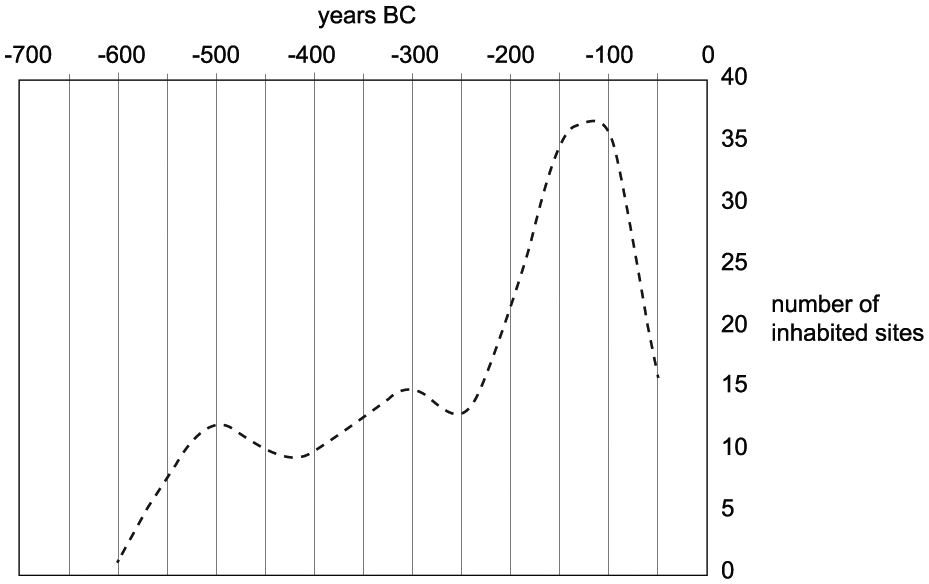
Generalized trends in Iron Age settlement foundation and abandonment in Brittany, divided in ‘time slices’ of 50 years. Graph based on the dataset of well-investigated sites depicted in Figure 4 (after Menez and Lorho, 2013).
Settlement patterns
Iron Age settlements mainly appear at the edges of plateaus and promontories and on slopes nearby stream valleys (Giot, 1995; Le Bihan et al., 1990; Menez and Lorho, 2013; Figure 6). Various sites are situated in coastal areas (e.g. Baudry and Daire, 2013; Daire et al., 2011; Menez et al., 1990). The average distance between settlements is 500–2000 m, but in some microregions, contemporary sites from the last two centuries BC are only a few hundred meters apart (Le Bihan et al., 1990; Menez and Lorho, 2013). These generally have a small territory and may demonstrate that a certain level of ‘saturation’ had been reached. In the late 1st century BC and 1st century AD, the landscape was restructured. Some later Iron Age sites remained inhabited, but overall, the network of Roman settlements was less dense and consisted of sites of different sizes and characters – varying from small farmsteads to Gallo-Roman villa sites and early agglomerations (Henderson, 2007; Menez and Lorho, 2013).
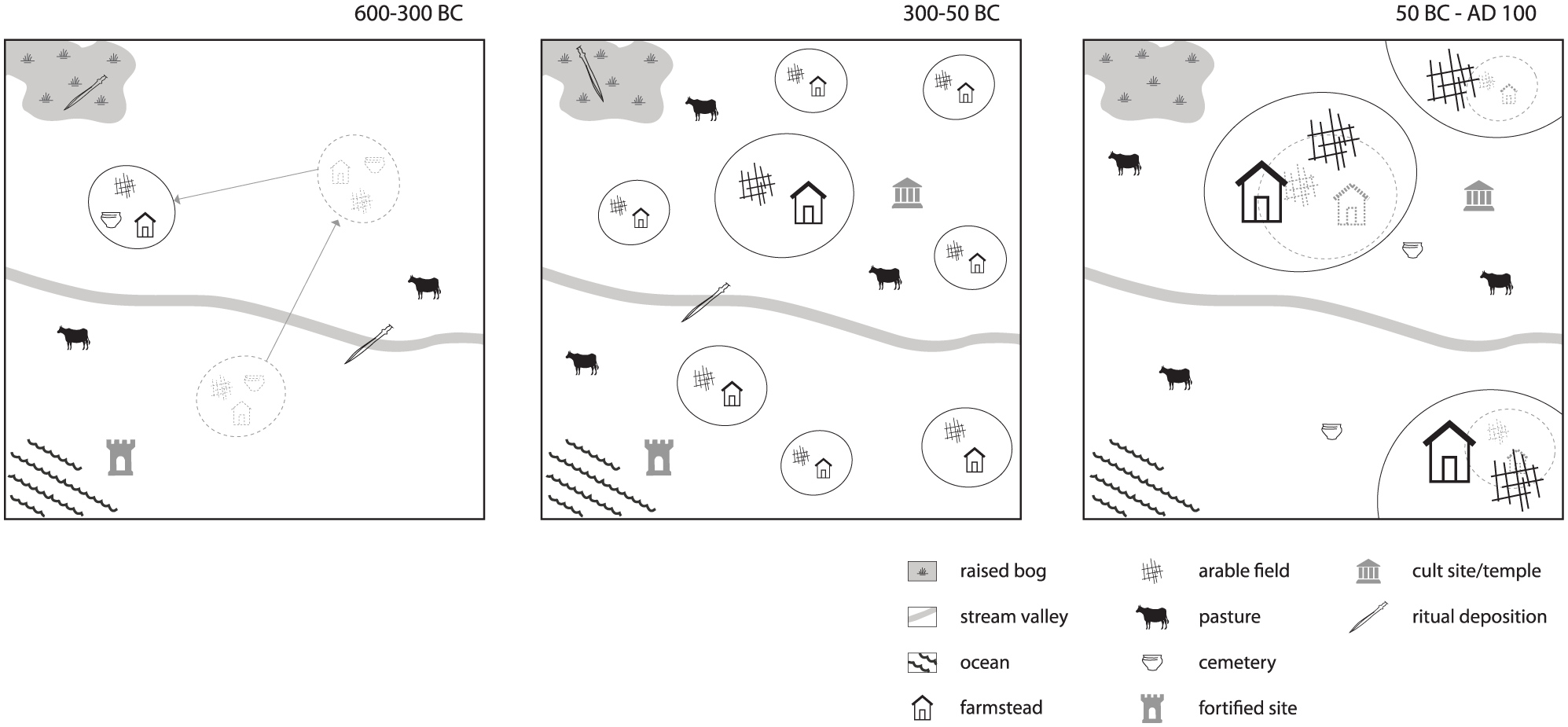
Schematic overview of some of the main changes in landscape organization in Brittany between the 6th century BC and 1st century AD. The first phase is characterized by a relatively low habitation density and dynamic settlement system. This is followed by a strong densification of the settlement pattern in the second phase, combined with a more ‘stable’ landscape organization and economic diversification. After an initial steep drop in habitation, the third phase is characterized by agricultural scale enlargement and increasing Roman influence. Some settlements are continuations of later Iron Age sites.
Landscape texture
Besides settlements, the Iron Age landscape structure was determined by fortifications, cemeteries, and ritual sites. Iron Age fortifications can broadly be divided into promontory forts (French: éperons barrés) and fortified inland sites (e.g. Henderson, 2007; Maguer, 1996; Wheeler and Richardson, 1957). Promontory forts are coastal promontories with defense systems. Fortified inland sites generally appear on hilltops near the confluence of streams. Both categories may have fulfilled a variety of functions such as refuges, temporary domestic residences, trading sites, and symbolic centers (e.g. Buchsenschutz, 1984; Henderson, 2007). Compared to other parts of France, the forts in our study area are small (<30 ha) and lack evidence of ‘urbanized’ elements (Giot, 1995; Henderson, 2007).
Small, dispersed cemeteries were dominant between the 6th and 4th century BC. Their life span was maximally 100–150 years (Menez, 1993; Menez and Lorho, 2013). Hardly anything is known about burial customs between the 3rd and 1st century BC (Menez and Lorho, 2013; Tanguy et al., 1990). Iron Age ritual practices were varied (Bouvet et al., 2003; Giot, 1995). Some cult sites were later overbuilt with Gallo-Roman temples (Bouvet et al., 2003). Various ritual offerings (e.g. weapons and pottery) have been retrieved from bogs, lakes, and streams (e.g. Allard et al., 1971; Léjars, 2014). Cult sites probably formed structuring elements in the densely settled later Iron Age landscape (Le Bihan, 2002).
Subsistence economy, agricultural expansion, and diversification
The Iron Age economy was essentially based on crop cultivation and cattle husbandry. For Northern France, Zech-Matterne (2015; cf. Zech-Matterne et al., 2009, 2013) recognizes three successive agricultural regimes. In the Bronze Age and First Iron Age, polyculture was practiced. Different cereals (e.g. barley, emmer wheat, spelt, and millet) were cultivated on small, well-maintained fields. The Second Iron Age witnessed specialization on specific species. Field systems were larger and maintained less intensively and appeared on a wider diversity of soils. The Roman period demonstrated another scale enlargement and further specialization. Specific to Northwest France are an early appearance of buckwheat and rye and a relatively high proportion of leguminous species (Malrain et al., 2013; Marguerie, 1992). The rare bone assemblages retrieved from settlements generally consist of cattle, pig, sheep, and goat, complemented with dog, horse, birds, and deer (Baudry, 2012; Giot, 1995; Méniel et al., 2009). At various coastal sites, many remains of seasonally exploited sea food are found, such as mussels, scallops, sea snails, oysters, and sea urchins (e.g. Dupont and Mougne, 2015; Giot et al., 1986; Mougne et al., 2014). Other coastal settlements fully depended on agriculture (Hyvert, 1990; Le Bihan et al., 1990).
The last three centuries BC witnessed economic expansion and diversification, probably linked to the expanding Roman consumer market (e.g. Galliou, 1990; Henderson, 2007; Menez and Lorho, 2013). The settlement density increased, field systems expanded, and well-maintained road networks were created. Local iron production increased, leading to the improvement and diversification of agricultural tools such as plowshares, sickles, and scythes (Giot, 1995; Langouët, 1990). Numerous salt production sites date to this phase (e.g. Daire, 2003; Gouletquer, 1970). Some settlements developed into regional production, storage, and trading centers (Henderson, 2007; Menez, 2009). Local coins were produced from the middle of the 2nd century BC onward (De Jersey, 1994). The abandonment of many farmsteads after the Gallic Wars may be attributable to their economic fragility and the loss of life in the war (Menez and Lorho, 2013).
Generalized phases
To conclude, the Iron Age was a period of rapid societal and landscape change. Based on archaeological data, three generalized phases in landscape development can be distinguished.
Phase 1 (600–300 BC): Characteristic is a dynamic settlement system, consisting of self-sufficient farmsteads practicing a system of polyculture. Settlements and cemeteries were relatively short-lived.
Phase 2 (300–50 BC): A dense, more ‘stable’ settlement pattern emerged. Human influence on the landscape increased greatly. Agricultural production was on a larger scale and more specialized. Simultaneously, an economic diversification took place.
Phase 3 (50 BC–AD 100): After an initial drop in settlement density, the region became fully incorporated in the Roman Empire. The settlement pattern was somewhat less dense than before and comprised a variety of settlement types. Agricultural production was upscaled. Larger trading networks developed.
Results: Trends in vegetal diversity and some vegetation groups
Palynological richness
The palynological richness expressed by the expected number of taxa E(Tn) and calculated as a mean value of all the sites demonstrates a regular increase during the three time periods. The highest diversity is reached during the beginning of the Roman period (50 BC–AD 100): numbers increase from E(Tn) value 28.25 in phase 1 (600–300 BC) to 30.08 in phase 2 (300–50 BC) and 32.34 in phase 3 (50 BC–AD 100; Figure 7).
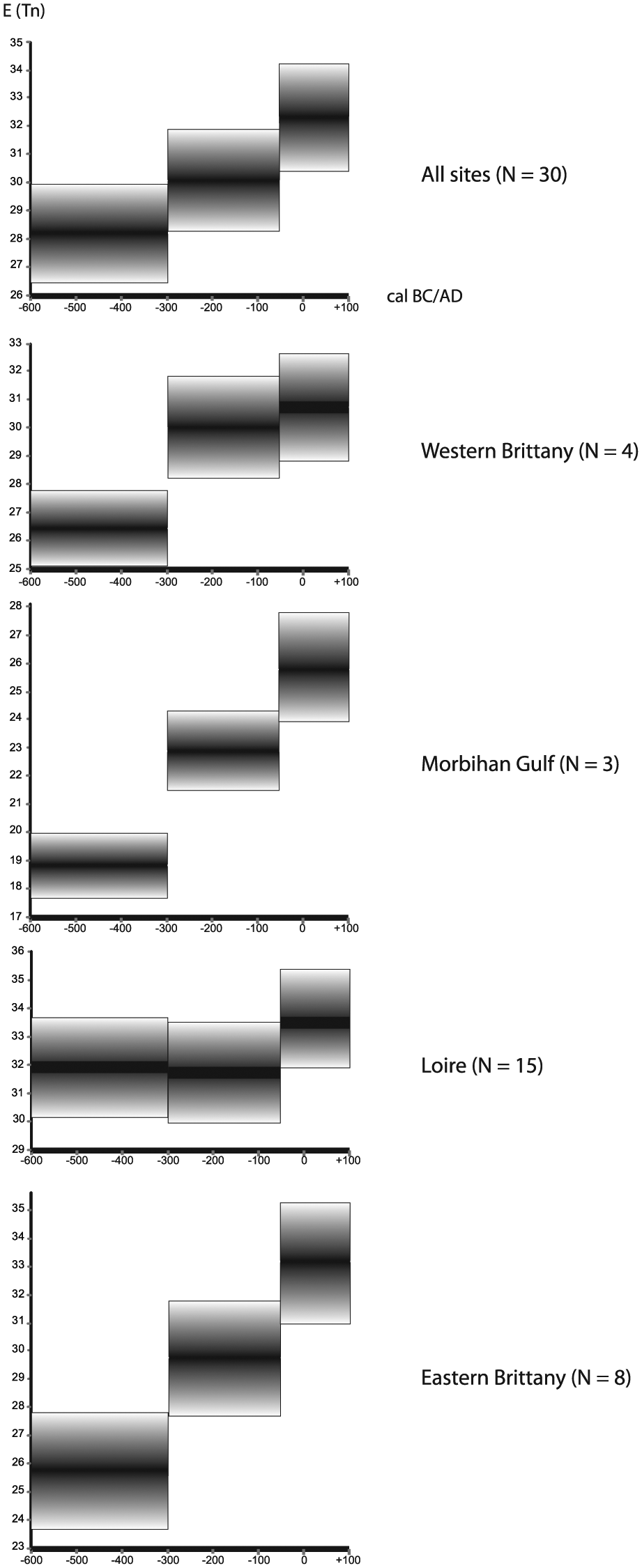
Combined results of the palynological richness (expected number of pollen taxa E(Tn) by rarefaction analysis) for all sites (top) and grouped per region, with graphical representation of the average and the SD of E(Tn) per phase.
In western Brittany an increase in palynological richness is observed (26.45 in phase 1, 30.02 in phase 2, and 30.70 in phase 3), with a strong rise between phases 1 and 2. In the Morbihan Gulf (southern Brittany), the E(Tn) values steadily increase: 18.86 (phase 1), 22.91 (phase 2), and 25.81 (phase 3). Again, the strongest increase occurs between phases 1 and 2. In the Loire area (southern part of the Armorican Massif), palynological richness is more or less equal in phases 1 (31.97) and 2 (31.72). Then, a period of expansion is identified between phases 2 and 3 (33.65). In eastern Brittany (Coglais-Mayenne), a regular increase in E(Tn) is observed during the three periods as well: from 25.70 in phase 1 to 29.75 in phase 2 and 33.15 in phase 3. The increase in the expected number between the phases is significant, especially between phases 1 and 2.
All over the study area, the taxomic richness increases steadily during the three periods under study, except for the Loire area (transition phases 1–2). At the beginning of the Roman period, the diversity of the taxa is systematically higher than during the Iron Age.
Dynamics of the vegetation structure
Among the five pollen-based community groups selected to analyze the vegetation dynamics and human impact through time, the ‘closed forest’ presents the most notable development (Figure 8). The average relative percentage of this group within all counted pollens (minus alder pollens and fern spores) ranges from 16% to 19%. On average, the representation of this group progresses over time. While the values in phases 1 and 2 are almost equal (16% and 16.4%, respectively), a significant increase is witnessed between phases 2 and 3 (from 16.4% to 19.3%). This general increase of closed forest in a period of increasing human environmental impact is remarkable. Within this trend, some regional differences are observed. The Loire region matches the general trend: a small rise between phases 1 and 2, followed by a steep increase in phase 3 (13.4%, 14.6%, and 20.2%). In the Morbihan Gulf, the landscape of phase 1 exhibits relatively few closed forests (7.8%). Phases 2 and 3 are distinguished by higher percentages (10.5%). The trends in eastern Brittany are diametrically opposite, with less closed forest over time and the most significant decrease between phases 2 and 3 (21.9%, 21.6%, and 19.4%). In western Brittany, phase 2 demonstrates a clear drop in closed forest pollen, before a marked extension in phase 3 (18.2%, 15.1%, and 20.3%).

Combined diagrams showing the dynamics of a selection of important vegetation types/communities throughout the three phases, for all sites (top) and grouped per region.
The average trends in the ‘open forest’ group, composed of four ligneous heliophilous taxa indicating woodland clearance, differ from the closed forest dynamics. A small decrease in open forest percentages is visible between phases 1 and 2, whereas those in phases 2 and 3 are about equal (15.1%, 13.9%, and 14.2%). Again, regional differences are encountered. The open forest trends in western Brittany resemble the closed forest developments (11.9%, 9.1%, and 14%). In the Morbihan Gulf, phase 2 is characterized by a strong increase in open forests, followed by a decrease in phase 3 (4.5%, 9.8%, and 6.4%). In the Loire region, phase 3 clearly demonstrates the highest open forest percentages (9.7%, 9.3%, and 12.4%). In eastern Brittany, the open forest group significantly decreases from phases 1 to 3 (28.7%, 24.6%, and 18%).
Concerning open areas, and more precisely light-demanding apophyte herbs and grasses which grow on disturbed soils, the ‘ruderals’ group decreases gradually when all sites are considered (7.1%, 6.5%, and 6%). Similar trends are encountered in western Brittany (4.7%, 4.6%, and 4.1%) and the Loire area (9.1%, 7.4%, and 5.9%). On the contrary, the Morbihan Gulf (3.1%, 8.2%, and 8.8%) and eastern Brittany (5.7%, 5.5%, and 6.3%) demonstrate an increase, with an especially significant rise occurring during phases 2 and 3 in the Morbihan (Figure 8).
Both in the entire study area and in each region, the representation of crops shows a small increase from the first phase to the third. The percentages are always low, around 0.5–2%. Phase 3 often demonstrates a clear rise in this anthropochore group and thus in agricultural practices. As expected, taxa included in the ‘adventives’ group, which grow on agricultural fields, demonstrate similar trends. However, in the Morbihan Gulf, adventives are more numerous in phase 2 (0.8%, 1.15%, and 0.6%).
Discussion and conclusion
Diversity
Biodiversity was tackled here by species richness (alpha diversity) through time (temporal beta diversity) and space (spatial beta diversity). As expected based on the archaeologically documented growing human environmental impact, beta diversity increases during the study period (Figure 7). This trend is common in all subareas, except in the Loire region where the values for phases 1 and 2 are similar. At the beginning of the Roman period, the taxonomic richness is systematically higher than before.
Gaudin (2004) studied the development of the number of ‘vegetation groups’ on well-drained terrains represented in Armorican pollen records. The distinguished vegetation groups are (1) arable fields, (2) wasteland and fallow land, (3) meadows and pastures, (4) heathland, (5) fallow land, (6) open or pastured forests, and (7) inhabited areas. Even though the number of vegetation groups represented at a regional scale cannot be equated directly with the degree of floristic diversity, it does provide insights into the level of landscape heterogeneity and the character of the landscape mosaic (Marguerie, 2009). Therefore, a comparison of these trends with those in taxonomic richness can indicate to which extent both are linked. The development of specific vegetation groups can either be of natural origin (e.g. caused by climate change) or, more frequently, result from human activity. The number of vegetation groups increases from 3.5 as a mean value in the Neolithic to 5.5 in the later Middle Ages. During the Iron Age (750–50 BC) and Roman period (50 BC–AD 500), 4.8 and 5.5 groups are recorded. It can be concluded that the trends in species richness and landscape heterogeneity during the period under study appear to match quite well.
Vegetation dynamics
While diversity increases gradually as expected by the archaeological data and thus reflects growing human environmental impact, the vegetation dynamics change. While we anticipated an increasing openness during the three phases under observation, the ‘closed forest’ group is constant or on the rise. As Quercus is the dominant taxon, the dynamics of this group mainly reflect trends in the representation of this tree.
The overrepresentation of ‘closed forest’ in phase 3 in Western Brittany is due to Quercus reaching a remarkable peak during this phase at the Vénec site. The other sites show a regular decrease in this taxon from phases 1 to 3. Among the ‘open forest’ taxa, Corylus is well represented in the first phase at three sites in this region. In the Morbihan Gulf, the increase in Quercus from phases 1 to 3 ends with a peak in the third phase at Suscinio. In the Loire area, the Quercus rise is regular from phases 1 to 3 at seven sites, and Fagus demonstrates the same behavior at five sites with an occasional overrepresentation in phase 3. Finally, in Eastern Brittany, the decrease in ‘open forest’ is due to the decline of Corylus through time at six sites and an overrepresentation of this taxon during phase 1 at Ballée Les Noës.
Irrespective of the area, the wooded taxa are always between 15% and 20% of the pollen assemblages. When these pollen values are transformed into estimated vegetation cover by the REVEALS model, forest vegetation forms about 10–15% of all ecosystems in the period under study (Figure 2). These values indicate a relatively open landscape. An overview study of palynological research in Iron Age France indicates two clear thresholds in woodland clearances: the first between the early and middle La Tène and the second between the middle and late La Tène. The first threshold is also characterized by an increase in the representation of ruderal herbs. The western part of France appears to have been more wooded than most other regions (Leroyer et al., 2009). In the French Massif Central, the 2nd century AD is a period of widespread woodland exploitation for agro-pastoral purposes at local and regional scales (Miras et al., 2015).
Relating to the process of forest clearance, a strong significant relationship exists between the percentage of herbaceous pollen and palynological richness (Goring et al., 2013; Meltsov et al., 2013; Morales-Molino et al., 2011). Increasing openness provides habitats for more species than completely forested landscapes do (Honnay et al., 2003; Meltsov et al., 2013; Söderström et al., 2001). However, semi-open landscapes exhibit greater floristic richness than completely open or completely forested landscapes (Billeter et al., 2008). The period and the region under examination here correspond to such a semi-open landscape. Since the Second Iron Age in Northwest France, human pressure on the woodlands, including deforestation practices such as slash and burn farming, was strong and induced the development of heathlands (Gebhardt and Marguerie, 2006; Vuorela, 1976). This expansion is particularly clear during the third phase at the sites of Langazel (western Brittany), Cordemais, Logné, Mazerolles (Loire), and Ballée-Noës and Baforière (eastern Brittany).
Light-demanding, apophyte and anthropochore herbs from the ruderal and adventive groups attest to the presence of meadows, pastures, livestock, and cultivated fields. The pollen values of these vegetations become a significant part of the pollen diagrams from the Second Iron Age onward. While the adventives are more or less rising in all areas, the ruderals appear to decrease except at the Morbihan Gulf and eastern Brittany. Plantago forms the basis of the ruderal group, whereas Rumex is often seen as representative of the adventives. In all areas, weeds and herbaceous plants favored by grazing are predominantly concentrated in phase 3. However, some counter-examples are found. In western Brittany, for example, Rumex and Plantago are registered in phase 1 at Saint Jean du Doigts and Vénec. In the Loire area, phase 1 demonstrates Plantago and Ranunculaceae at Ancenis. In eastern Brittany, ruderals are present from the very beginning of the analyzed period at Thévalles, Lévaré, Ballée-Noës, and Baforière. The results may be interpreted as reflecting an agricultural system at least partly based on livestock raising, as several indicators of grazing became frequent in the diagrams, for example, Plantago, Potentilla, and Rumex. This observation is corroborated by the rare bone assemblages, mainly consisting of cattle, pig, sheep, and goat, found at archaeological sites.
Evenness
More taxa are detected when fewer dominant taxa are present and then the pollen evenness is greater (Colombaroli et al., 2013). Most of the study sites in Northwest France correspond to marshes that developed on valley floors. During their development, the marshes frequently host a riparian vegetation mainly comprising Alnus. This ligneous hygrophilous vegetation can influence pollen dissemination. This type of local vegetation developed around the marshes declines as a result of clearances. Compared to the previous Sub-Boreal and Late Atlantic, Alnus declines during the studied period, and a positive correlation is noticed between palynological richness and evenness at that time. The ligneous riparian vegetation generally gives way to a reedy marsh. Nevertheless, Alnus is still dominant at Vénec (phases 2 and 3) in western Brittany, at Suscinio (phase 1) in the Morbihan Gulf, at eight sites in the Loire area, and at six sites in eastern Brittany.
In some cases, other taxa are very dominant in the pollen diagrams. This applies to Quercus in Langazel (phase 1) and Vénec in western Brittany, in Suscinio (phase 3) in the Morbihan Gulf, and at Poupinière (phase 2) in the Loire region. Corylus is dominant at Cordemais (phase 2) in the Loire area and at Thévalles, Lévaré (phases 1 and 2), and Glatinié (phase 1) in eastern Brittany. Finally, Betula and Salix are dominant at Saint Thomas in the Loire area. Low pollen evenness could either reflect a poor diversity of plant communities or result from a local dense woody vegetation composed of Alnus and/or Corylus getting in the way of pollen in the catchment area.
At a wider spatial scale, increasing human interferences may create a more fragmented landscape and new habitats with an enrichment of the flora by new species of apophytes and anthropochores (Berglund et al., 2008; Marguerie, 2009). Small-scale clearances in the landscape are considered to be an important factor influencing floristic richness in the northern temperate zone (Billeter et al., 2008; Meltsov et al., 2011). As noted above, Birks and Line (1992) argue that floristic richness in a fragmented landscape is greatest at ecotonal boundaries between vegetation types. From the first phase onward, our study area was influenced by the expansion of clearings, grazing, and farming which led to a patchy cultural landscape. The Iron Age and Roman landscapes of Northwest France seem to have been complex half-open landscape mosaics with an alternating mixture of forests, dry and wet meadows, heathland, pastures, settlements, and cultivated fields.
Land use–biodiversity relations
As mentioned, various authors have already stressed a positive correlation between human impact and taxa richness. In southern Estonia, for example, the pollen dataset of nine small lakes shows that pollen richness corresponds to land use and that this pollen richness has a strong correlation with nonforested areas (Meltsov et al., 2011). In southern Sweden, Berglund et al. (2008) correlated low diversity values with agrarian and settlement crises. In the same area, Gaillard et al. (1998) found a significant positive correlation between nonarboreal pollen percentages and the percentage of open herb vegetation cover. This correlation has been clearly demonstrated in our study as well. The increasing richness values in our research area correspond with the growing impact of human activities on the landscape. This applies to the general trends during the 700 years under study.
The paleo-vegetation richness increase can be interpreted as the combined effect of four phenomena (Marguerie, 2009):
Expansion of new groups of (heliophilic) vegetation in a more open landscape because of deforestation. This leads to less competition between species, creation of new landscape units, and compositional turnover (Balée, 2006; Berglund et al., 2008);
Heterogeneity of human pressures across an area leads to several stages of vegetation successions being present at the same time;
Introduction of nonnative plants, not only consumables and crops but also above all anthropochores such as neophytes that appear due to agricultural practices (plowing and manuring) and trade;
As shown by Odgaard (1999), the increase in pollen richness also results from an increase in total pollen production.
In the study area, the forests were opened up from the Neolithic to improve grazing and cultivation (Marguerie, 2009). During the Iron Age, arable farming is clearly well established. The crops, in particular cereals, are frequently registered in the pollen assemblages. However, they rarely display constant values during the three phases. Most crops are registered in phase 3, but earlier recordings are frequent as well. For instance, Cannabis was cultivated in western Brittany at phase 1 in Saint Jean du Doigts. In the Loire area (Cordemais, Gesvres, and Mazerolles), and eastern Brittany (Thévalles and Ballée-Noës), cereals are also present from phase 1 onward. The relatively low and discontinuous cereal values may point to an extensive agricultural system with nonpermanent fields. The cereal pollen may originate from a kind of shifting cultivation where new lands were continuously cleared for cultivation and grazing (Lindbladh, 1999). Based on archaeological evidence and especially the steep increase in settlement sites (often with clearly visible field systems), one might expect a more spectacular rise in cereal values during the period under study. However, we have noted an economic diversification in phases 2 and 3, which means that cereal farming was only one component of the late Iron Age and early Roman subsistence economy. Unfortunately, current knowledge on Iron Age and Early Roman agricultural practices in Northwest France leave a lot to be desired.
The well-known ‘intermediate’-disturbance hypothesis by Connell argues that biodiversity is higher at intermediate levels of disturbance on the scales of frequency and intensity (Connell, 1978; Rosenzweig, 1997; Wilkinson, 1999). This is corroborated by this study and by previous research in Northwest France (Gaudin, 2004; Marguerie, 2009). Before this period, the level of disturbance was too low to produce diversity maxima. In general, most pollen records demonstrate a decrease in floristic richness since the 18th/19th century AD (e.g. Birks et al., 2016; McGill et al., 2015). According to Berglund et al. (2008), human impact in southern Sweden represented an ‘intermediate’ level of disturbance from the Late Bronze Age to AD 1800. Comparable results are recorded in Denmark by Odgaard (1994). The small degree of vegetation openness produced by agriculture during this period is important in promoting the coexistence of species (Colombaroli et al., 2013). The cultural landscapes resulting from pre-intensive agrarian systems clearly contribute to biodiversity (Marty, 2005). Human pressure on the landscape prevents the expansion of only one vegetation component. In this case, human impact leads to a heterogeneity of dynamics levels. If the pressure decreases for a long time (e.g. because of population decrease or desertion of an area), the levels become more homogeneous (Marty, 2005).
In western France, a decrease in taxonomic richness is observed since the 18th century AD too. This is the time of the expansion of a more intensive, single-species, modern agriculture, and an increasing homogeneity in landscape structure. This cannot be a coincidence, and these kinds of practices led to the loss or deep alteration of inherited landscapes (Marty, 2005). The anthropogenic pressure exerted then exceeded the ‘intermediate’ level and had a negative effect (Berglund et al., 2008). A biodiversity reduction will affect the system resilience ultimately resulting in a loss of capacity to positively react to climatic or human pressures.
Implications and considerations
The large geographical area studied in this paper was selected to signal and explain large-scale changes in floristic richness and in the dynamics of some vegetation groups. Integration of a large number of pollen sites allows to assess biodiversity trends in different geographical areas at different spatial scales. Furthermore, by comparing pollen data with archaeological information, different land-use changes could be distinguished such as the conversion from forests to pastures and croplands (cf. Birks et al., 2016; Newbold et al., 2015). This method can also be applied in other areas and time frames, provided high-resolution palynological and cultural (archaeological and historical) data are available.
Even though the incorporation of different types of data in this study has yielded some promising results, their integration is not unproblematical. Whatever parameter is taken into account (richness, vegetation dynamics, and anthropogenic indicators), no marked breaks are apparent in the pollen data during the studied period. By contrast, the archaeological evidence points to a period of rapid and intensive societal change, in various respects. Archaeologically, the intensifying land use and agricultural intensification in our study area and period is most clearly visible between 200 and 50 BC and in the 1st century AD. This probably went hand in hand with further woodland clearances. It is difficult to tell whether the abandonment of farmsteads in the turbulent decades around the Gallic Wars (58–51 BC) led to some degree of forest regeneration and might account for part of the observed increase in closed forest. No clear peak in closed forest can be discerned in this specific period. This might result from an insufficient dating resolution of the pollen data. However, this phase of unrest may have been too short to leave a clear mark in the paleoecological record.
Future research could focus on exactly the latter topic: are archaeologically or historically documented land-use changes on small timescales not influential enough to be reflected in richness values, or do we not recognize them because the resolution of the available dating and palynological data are not sufficient at this moment? Detailed interdisciplinary research on a local/microregional level using high-resolution archaeological and environmental data, including well-dated pollen samples, should be able to take research in this field to another level.
Footnotes
Acknowledgements
We are grateful to Marie-Yvane Daire (CNRS-CReAAH-Université Rennes 1, France) and Yves Ménez (Direction régionale des affaires culturelles Bretagne, France) for providing information on the Iron Age habitation of the research area and for commenting on an earlier draft of this paper. Florence Mazier (CNRS-GEODE-Université Toulouse 2, France) and Yannick Miras (CNRS-GEOLAB-Université Blaise Pascal, Clermont-Ferrand, France) advised on and assisted with the rarefaction analysis software. Yann Rantier (CNRS-ECOBIO-Université Rennes 1, France) gave precious help in creating Figure 1. Alistair Bright (Woubrugge, Netherlands) edited the final English draft.
Funding
This work was supported by the Institute Écologie et Environnement (INEE) of the Centre National de la Recherce Scientifique (CNRS) within the framework of the postdoctoral award DIPEE Paysages-Rennes (les dispositifs de partenariat en écologie et environnement).