
Abstract
The ombrotrophic peat bogs of Tierra del Fuego are located within the southern westerly wind belt (SWWB), which dominates climate variability in this region. We have reconstructed late-Holocene water-table depths from three peat bogs and aimed to relate these records to shifts in regional climate. Water-table depths were quantified by the analysis of testate amoeba assemblages, and a regional transfer function was used to infer past water-table depths. During the last 2000 years, testate amoeba assemblages have been relatively stable, with a dominance of Difflugia pulex and Difflugia pristis type, and an increase in Assulina muscorum and other Euglyphida at the top of each section. Multivariate analyses show that water-table depth remained the main environmental variable explaining assemblages along the TiA12 core, but reconstructions were not significant for the two other cores. In line with the low variability in assemblages, water tables were relatively stable during the last 2000 years. Slightly wetter conditions were found between ~1400 and 900 cal. BP and a pronounced recent dry shift was reconstructed in all of the three peat profiles. Considering the regional climatic context, this recent shift may have been forced by a decrease in precipitation and warmer conditions linked to an increase in the importance of the SWWB. Nevertheless, we cannot exclude the influence of higher UV-B radiation resulting from the local degradation of the ozone layer since the late 1970s, which may have had an additional effect on the relative presence of A. muscorum in the southern Patagonian region.
Introduction
The climate of southern Patagonia is strongly influenced by the Southern Annular Mode (SAM), defined as the zonal mean pressure difference between the Southern Hemisphere mid-latitudes and Antarctica (Marshall, 2003). Although late-Holocene variations in climate have been identified in southern Patagonia (Chambers et al., 2014; Villalba et al., 2012), most reconstructions are qualitative and this region remains less documented than many regions in the Northern Hemisphere. For instance, the latest IPCC report states that there is ‘medium confidence that the positive trend in SAM since 1950 may be anomalous compared with the last 400 years’ (Masson-Delmotte et al., 2013). A positive SAM represents a large pressure difference between the mid-latitudes and Antarctica, which results in an intensification and poleward shift of the Southern Hemisphere westerly jet stream (Archer and Caldeira, 2008), which is strongly associated with the position and intensity of the southern westerly wind belt (SWWB). As the position of the SWWB varies at seasonal to millennial timescales (Lamy et al., 2010), SWWB dynamics exert a strong influence on the climate of southern Patagonia. At decadal timescales, the position of the SWWB may also have been affected by stratospheric ozone depletion since the 1970s (Thompson et al., 2011), because ozone depletion is associated with climate conditions similar to those of the positive phase of the SAM.
A range of proxies from various archives have been employed to reconstruct past changes in the southern Patagonian climate linked to SAM and SWWB dynamics, including tree ring, glacier, lake level and sediment composition, peatland and speleothem records (Boucher et al., 2011; Chambers et al., 2014; Mauquoy et al., 2004; Schimpf et al., 2011; Villalba et al., 2012; Waldmann et al., 2010). Many, if not most of these SAM/SWWB reconstructions, are based upon improved understanding of the link between westerly wind intensity and precipitation. The location of southern Patagonia near the core of the SWWB, combined with the orographic effect of the Andean Cordillera, results in an extreme regional rainfall gradient. Recent analyses suggest the SWWB brings high precipitation and mild temperatures to the western side of the Andean Cordillera, yet mild and very dry conditions are prevalent at the eastern side (Garreaud et al., 2013), with current mean annual rainfall decreasing about 20-fold from the main divide to the eastern slopes of the Andes near Punta Arenas (Schneider et al., 2003). The positive phase of the SAM and increased SWWB intensity are also associated with warmer conditions, concentrated in the summer season, and higher wind velocity (Abram et al., 2014; Moreno et al., 2014; Villalba et al., 2012).
In this paper, we have used testate amoeba analyses, novel for this region, to reconstruct regional climate dynamics from peatland records. Cores from ombrotrophic peatlands may be used as records of past climate change because these ecosystems are mainly fed by precipitation (Belyea and Baird, 2006; Swindles et al., 2012). As a result, reconstructions of water-table variability are generally interpreted as records of precipitation-driven summer water deficit (Booth, 2010; Charman et al., 2009). Because of the influence of local and autogenic factors, a series of peatland records is often crucial to identify changes which may be driven by climate variability. Peat records are often continuous and they generally allow for high-resolution chronologies because of the important presence of well-preserved organic material, notably Sphagnum mosses. A gradient of peatland types occurs in southern Patagonia related to the strong precipitation gradient and topographic setting of the peatlands. On the western, hyperhumid flanks of the Andes, blanket bogs are common, while the eastern parts of Tierra del Fuego are characterised by dry fens located within a flat steppe landscape (Grootjans et al., 2010). The western parts of Tierra del Fuego, as well as the peatlands around Punta Arenas, are generally ombrotrophic with a high dominance of Sphagnum magellanicum (Kleinebecker et al., 2007) and are therefore optimal for climate reconstructions. The studied bogs are located in the rain shadow of the Andean Cordillera and therefore mean annual precipitation is limited to 400–500 mm yr−1 (IAEA/WMO, 2013). These relatively dry conditions, together with a mild climate and low seasonality, suggest the peatlands develop at the warm-dry margin of the peatland climate envelope when projected onto the Northern Hemisphere peatland ‘climate space’ (Loisel and Yu, 2013).
The identification and quantification of testate amoeba assemblages may permit the quantification of decadal-scale variations in water-table depth (WTD) from various peatland types (e.g. Booth and Jackson, 2003; Mitchell et al., 2001; Payne et al., 2008; Swindles et al., 2015b; Warner and Charman, 1994). These unicellular organisms, which are most abundant at the surface of humid ecosystems, create proteinaceous, biosiliceous or xenogenic shells (‘tests’) that are generally well preserved in peat. In ombrotrophic bogs, testate amoeba assemblages often show a good relationship with local humidity conditions (Booth, 2002). These characteristics have allowed the development of transfer functions and application of these to peat records to reconstruct variability in local WTDs (Amesbury et al., 2013; Booth, 2008; Charman et al., 2007; Lamarre et al., 2013; Qin et al., 2013). A transfer function for ombrotrophic peatlands in southern Patagonia has recently been developed (van Bellen et al., 2014). Here, we apply this transfer function to late-Holocene testate amoeba assemblages from three peat cores to create high-resolution records of quantified WTDs in the region. Given the late-Holocene dominance of S. magellanicum in southern Patagonian bogs (Mauquoy et al., 2004), testate amoebae may be particularly useful as a proxy for past WTD in these ecosystems.
The main objective of this research is to reconstruct regional climate changes during the last 2000 years and to identify possible linkages with previously established SAM/SWWB records using the S. magellanicum raised peat bog archives in Tierra del Fuego. The region east of the main Andean divide currently shows a weak, negative relationship between local precipitation and SWWB intensity because only Atlantic, easterly air masses are able to bring humid air to the region (Garreaud et al., 2013). However, it is difficult to reconstruct late-Holocene SWWB and SAM dynamics because the sign and amplitude of the relationship between SAM and climate are likely to vary on decadal to millennial timescales (Moreno et al., 2014; Silvestri and Vera, 2009). In addition, our study region is located just east of the main Andean divide within a complex orography. Just east of the Andean crest, including the region around Punta Arenas, the correlation between westerly wind strength and precipitation may be only weakly negative and high westerly wind speeds may be associated with important rates of evaporation (Kilian and Lamy, 2012). As a result, an increase in large-scale westerly wind intensity over this region may not have a clear effect on peatland water balance. Moreover, rather than a north–south orientation, the Andean divide in Tierra del Fuego has a northwest–southeast orientation which may attenuate the rain shadow effect to the northeast. Considering the orographic complexity of southern Patagonia, Daley et al. (2012) suggested relatively important intra-regional differences in moisture source for precipitation between Punta Arenas and Ushuaia at decadal timescales.
Study region
The three studied peatlands (Tierra Australis, Andorra and Karukinka) are located between 55 and 53°S in Tierra del Fuego (Figure 1). All sites are S. magellanicum–dominated ombrotrophic peat bogs with mean pH around 4 (van Bellen et al., 2014). Previous research showed Andorra bog has low nitrogen deposition levels of 0.1–0.2 g N m2 yr−1 (Fritz et al., 2012) and given their remote location, nitrogen levels are likely to be as low in Tierra Australis and Karukinka peatlands as well. Although S. magellanicum tends to dominate along the entire microtopographical gradient, Sphagnum falcatulum, bordering pools and wet hollows, and Sphagnum fimbriatum are present in some locations as well. In the drier sections, Empetrum rubrum Vahl ex Willd. and Nothofagus antarctica (Forster) Oerst. are found, while some of the wetter lawns and hollows are characterised by Tetroncium magellanicum Willd. herbs. Marsippospermum grandiflorum (L.f.) Hook. is found in both drier and wetter locations. Coring locations were selected in pristine, ombrotrophic sections of each bog and cores were visually inspected to ensure continuous records of ombrotrophic peat were sampled. Andorra bog (200 m a.s.l.) is situated in a relatively narrow, NW-SE oriented valley near the Beagle Channel 7 km north of the city of Ushuaia. It is an S. magellanicum–dominated ombrotrophic bog, intersected and bordered by a few small streams. Core AND12 was sampled in the main part, about 200 m from any of the streams. Tierra Australis bog (130 m a.s.l.) is located about 45 km ENE of Ushuaia and 3 km south of Lago Fagnano, northeast of the Andean cordillera. Similar to Andorra bog, it is largely dominated by S. magellanicum although its microtopography is more pronounced (van Bellen et al., 2014). Core TiA12 was sampled 150 m north of a sector of the bog that has recently been drained for block cutting. Karukinka bog (220 m a.s.l.) is located 125 km NW of Andorra bog on the Chilean side of Tierra del Fuego and is part of Karukinka Natural Park. It is a pristine, S. magellanicum–dominated bog, located in a wide river valley characterised by some minerotrophic sectors. Core KAR13 was extracted from an area perched on a wide terrace adjacent to the valley.
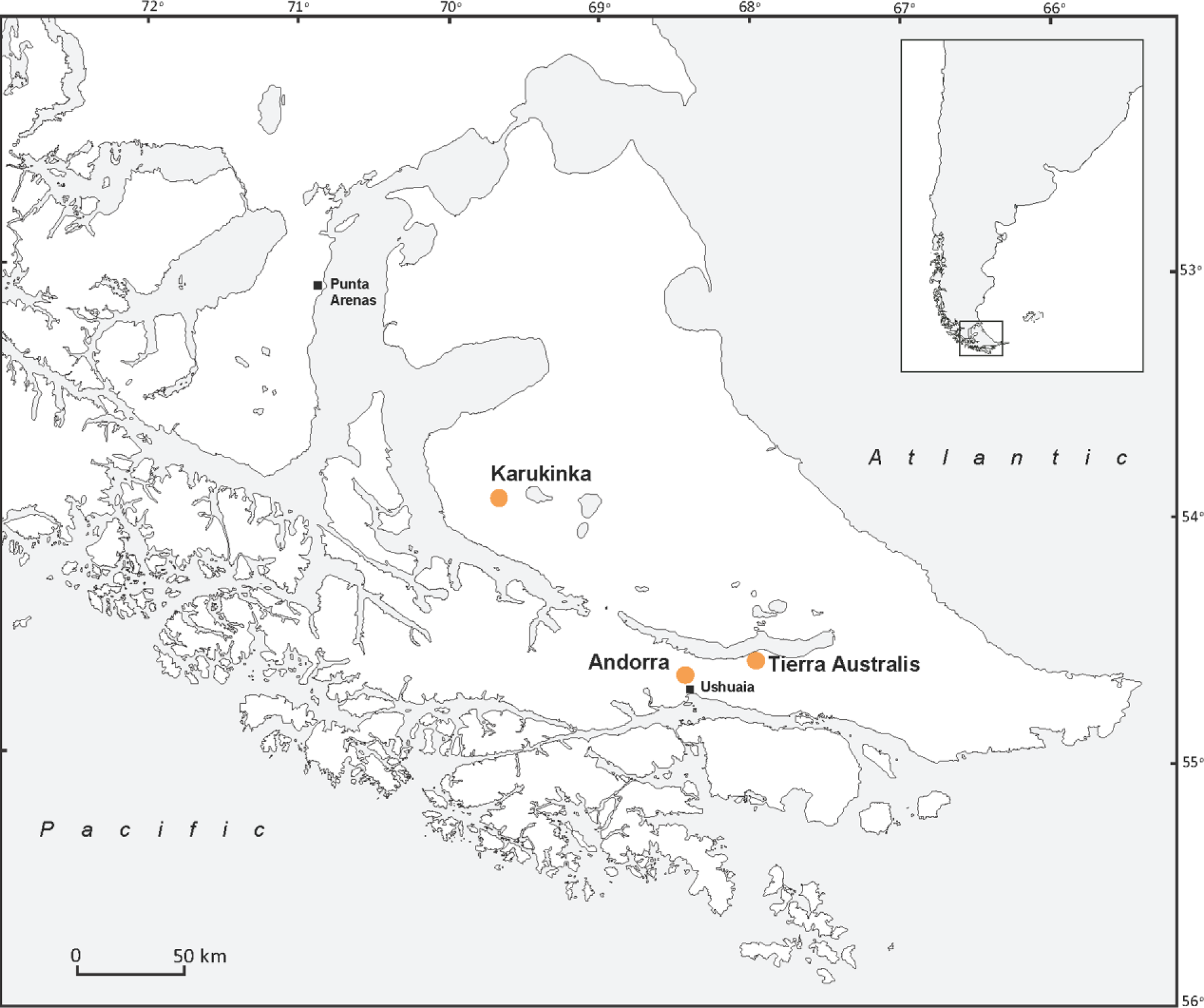
Location of the three studied peatlands in Tierra del Fuego.
The mean annual temperature and precipitation in Ushuaia are 6.2°C and 492 mm. Averages in Punta Arenas, 250 km NW of Ushuaia, are 6.1°C and 433 mm (IAEA/WMO, 2013). Although these values are similar and both regions are likely to be subjected to the same large-scale trend in SWWB dynamics, Punta Arenas and Ushuaia have different trends in precipitation isotopic composition (Daley et al., 2012). This may be related to differences in precipitation water sources and/or their topographic setting.
Methods
Fieldwork
Three complete cores were sampled in the austral summers of 2012 at Andorra and Tierra Australis bogs and 2013 at Karukinka bog. All cores were extracted from lawn microforms using a 50-cm-long, 10-cm diameter Russian peat sampler. Sections were wrapped in plastic film and aluminium foil and stored in a 50-cm PVC tube. Cores were stored horizontally in a cooler where possible before transport and storage in a cold chamber at 4°C.
Laboratory work
In the laboratory, each core was cut into 1-cm-thick slices. Subsamples of 1 cm3 were taken from the centre of each slice for testate amoeba analysis. These samples were prepared following the protocol of Hendon and Charman (1997), retaining the size fraction <355 and >15 µm. For identification of taxa, the key by Charman et al. (2000) was used and in some cases the adaptations by Booth (2008) were followed. Taxa not included in these keys were identified using publications by Vucetich (1978), Smith et al. (2008) and Jung (1942). For each sample, a minimum of 150 specimens was identified using a transmitted light microscope at 400× magnification and taxon abundance was expressed as a percentage of the total count.
WTD reconstructions and data exploration
WTDs were inferred using the weighted averaging–partial least-squares transfer function trained on samples from Tierra Australis, Andorra and Karukinka bogs as well as Parrillar bog near Punta Arenas (for details, see van Bellen et al., 2014). Prior to WTD reconstruction, fossil assemblages were adjusted in order to comply with the taxa included in the transfer function, because some taxa were not represented in the transfer function. To achieve harmonisation of fossil and training set assemblages, fossil taxa not present in the training set were eliminated and relative presences were re-calculated to attain a sum of 100%. WTDs were expressed with high, positive values corresponding to low water tables.
Multivariate analyses of square-root transformed fossil assemblages were performed using Canoco 5 (Ter Braak and Šmilauer, 2012) for each core separately to explore relationships between taxa. These analyses were further used to summarise datasets and evaluate the potential influence of environmental variables. Preliminary detrended correspondence analyses (DCA) of the fossil data showed gradient lengths of 3.6 standard-deviation units (SD) for TiA12, 2.5 SD for AND12 and 2.7 SD for KAR13. Generally, only gradient lengths exceeding 3 standard deviations justify the use of a unimodal response model implemented in DCA (Ter Braak and Prentice, 1988). Therefore, we applied the linear model of principal component analysis (PCA) in all analyses. Species richness was quantified by rarefaction to 100 specimens using Vegan 2.2-1 package (Oksanen et al., 2015) in R (R Core Team, 2013) to correct for any differences in count totals. Finally, zonation by stratigraphically constrained incremental sum of squares cluster analysis was performed on square-root transformed data with Psimpoll 4.27 (Bennett, 2007).
WTD reconstructions were tested for statistical significance using two methods included in the palaeoSig package in R (Telford, 2015). The ‘randomTF’ method compares the proportion of variance in the species data explained by the reconstruction with that explained by reconstructions based on random environmental variables (999). The rationale for this is that, in order for a WTD reconstruction to be considered significant, it should explain more of the variance in assemblages than 95% of the reconstructions from transfer functions trained on random data. A second test of the accordance of training set and fossil assemblages was performed using palaeoSig and the ‘obs.cor’ command. Taxon WTD optima were quantified by weighted averaging of the training set and these values correlated with Axis 1 species scores from redundancy analyses (RDA) constrained by the reconstruction. The strength and significance of this correlation indicates the degree of consistency in the relative position of taxa in modern and fossil assemblages. It should be noted that neither of these tests assesses whether the reconstruction is ‘correct’; that could only be assessed by correlation against a ‘reliable’ independent proxy.
Dating and chronologies
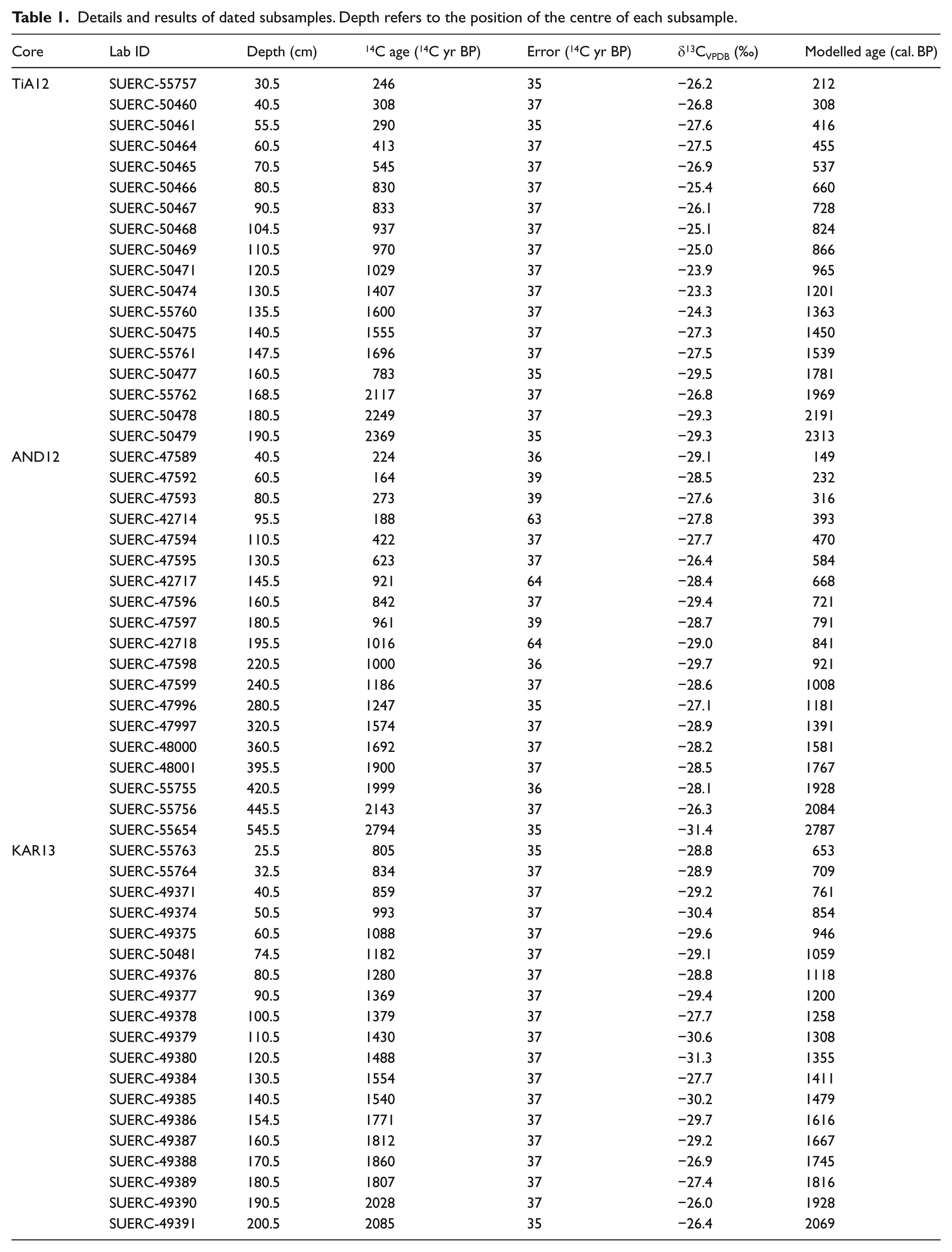
The age/depth models are based upon a total of 56 14C AMS dates (Table 1), which were obtained from 1-cm slices of peat. Fresh subsamples for dating were preferentially composed of Sphagnum stems and branches and ages were expressed as calendar years before present (BP, referring to 1950 CE) after calibration with the SHCal13 calibration curve (Table 1; Hogg et al., 2013). Chronologies were modelled using a Bayesian approach implemented in the Bacon version 2.2 package in R (Blaauw and Christen, 2011). After calibration, the modelling procedure of Bacon takes account of the entire probability distribution of each dated level while creating smooth chronologies including estimations of age uncertainties.
Details and results of dated subsamples. Depth refers to the position of the centre of each subsample.
Results
Testate amoeba assemblages
Testate amoeba assemblages from the three studied bogs are largely dominated by Difflugia pulex and Difflugia pristis type for most of the late-Holocene (Figure 2). This includes zones TiA-1 and TiA-5, AND-2, AND-3 and AND-5, and KAR-4 and KAR-5 in Figure 2. A shift in assemblages towards the top of the sequence is observed in TiA-6, AND-6 and KAR-6, with dominance of Assulina muscorum, Corythion-Trinema type and to a lesser extent, Euglypha tuberculata type. Core TiA12 shows a presence of some hydrophilous taxa, including Amphitrema wrightianum and Difflugia globulosa type, between 140- and 118-cm depth in zone TiA-2. The three records show that assemblages are relatively species-poor, with mean species richness values of 8.1, 8.8 and 9.9 taxa for TiA12, AND12 and KAR13, respectively.

Testate amoeba assemblages as a function of depth and age for each of the three peatlands. Amoeba abundance is expressed as percentage of the total count. Taxa included in the transfer function of van Bellen et al. (2014) are sorted by weighted-averaging optima from xerophilous (left) to hydrophilous (right).
Most of the frequently encountered fossil taxa correspond to the taxa well represented in the training set of van Bellen et al. (2014). PCA species plots show similar taxon distributions for each of the three cores (Figure 3). The dominant D. pulex and D. pristis-type are separated at the lower extreme of Axis 1. A second group is formed by the xerophilous Euglyphidae A. muscorum, Corythion-Trinema type, Trinema lineare, E. tuberculata type and Euglypha strigosa type, which are located at the higher end of Axis 2. Remaining taxa, including most of the hydrophilous taxa, are roughly separated in a third group. The absence of clear hydrological gradients along any of the axes suggests taxon assemblages in these cores may reflect a range of environmental variables.
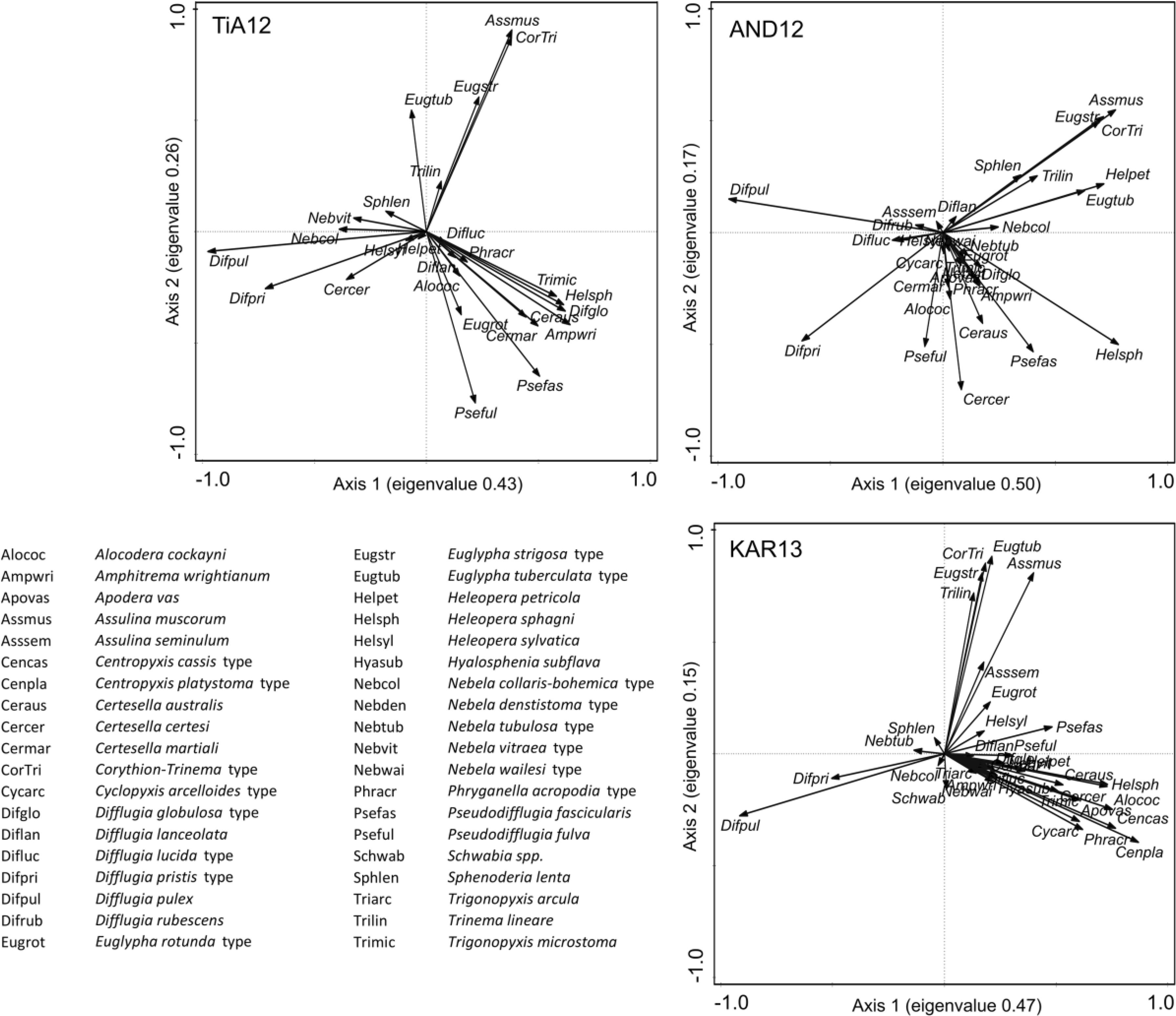
PCA species plots for each core. Taxa are abbreviated by the first three letters of genus and species as listed.
Analyses of the fossil and modern datasets with palaeoSig demonstrates that the weighted averaging–partial least-squares reconstruction did not explain any significant variance at 23.0% (p = 0.56), 36.4% (p = 0.22) and 23.9% (p = 0.65) for TiA12, AND12 and KAR13, respectively. Abundance-weighted correlations between RDA Axis 1 scores and weighted-average optima from the transfer function show a significant correlation for TiA12 (p = 0.02, r2 = 0.89), but AND12 and KAR13 had p-values of 0.43 (r2 = 0.64) and 0.21 (r2 = 0.64), respectively (Figure 4).
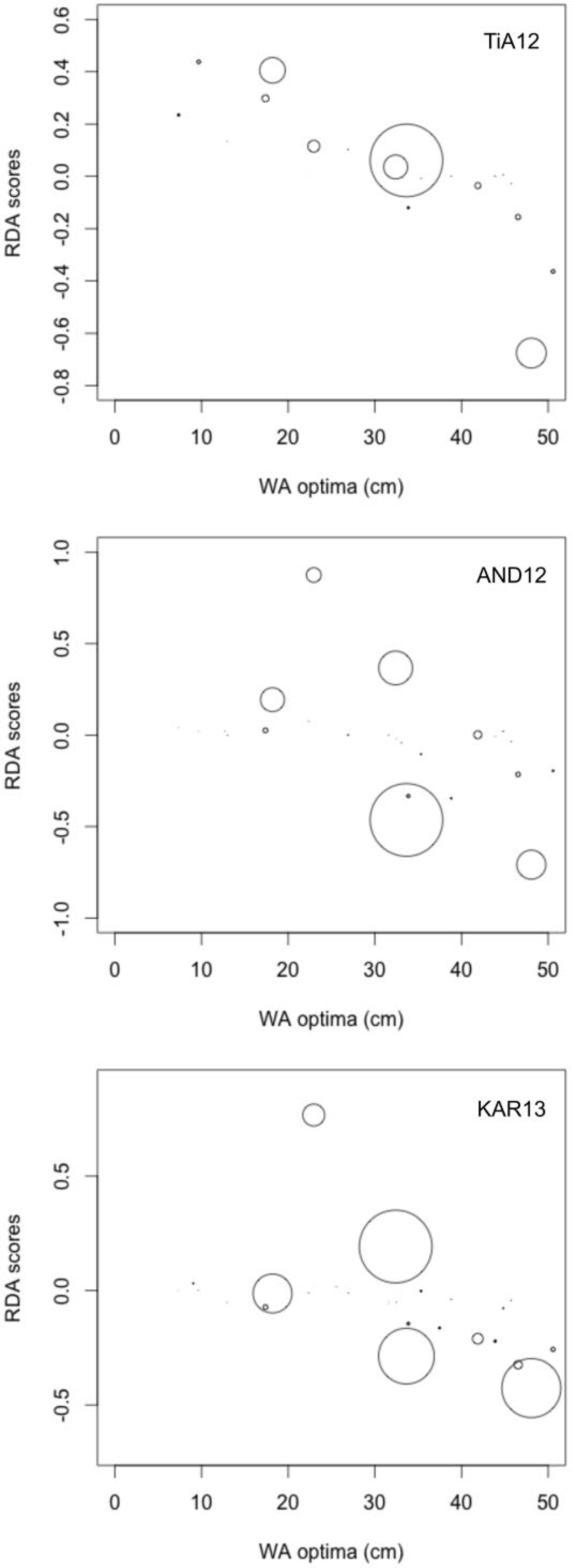
Abundance-weighted correlation between weighted-average optima for taxa observed in each core and RDA species scores of an ordination constrained by the WA reconstruction. TiA12: p = 0.022 (r2 = 0.89); AND12: p = 0.426 (r2 = 0.64); KAR13: p = 0.207.
Chronologies
Of the 56 samples prepared for 14C dating, one resulted in a clear outlier (core TiA12; 160.5-cm depth), which caused an age inversion (Table 1; Figure 5). This date was therefore omitted in the modelling. The high median accumulation rate of AND12 of 2.33 mm yr−1 may result from melting snowpacks delivering water to the river in whose valley AND12 sits. This higher river discharge could impede efflux from the bog, resulting in a higher stability of local water tables and stimulation of peat accumulation (Chambers et al., 2014).
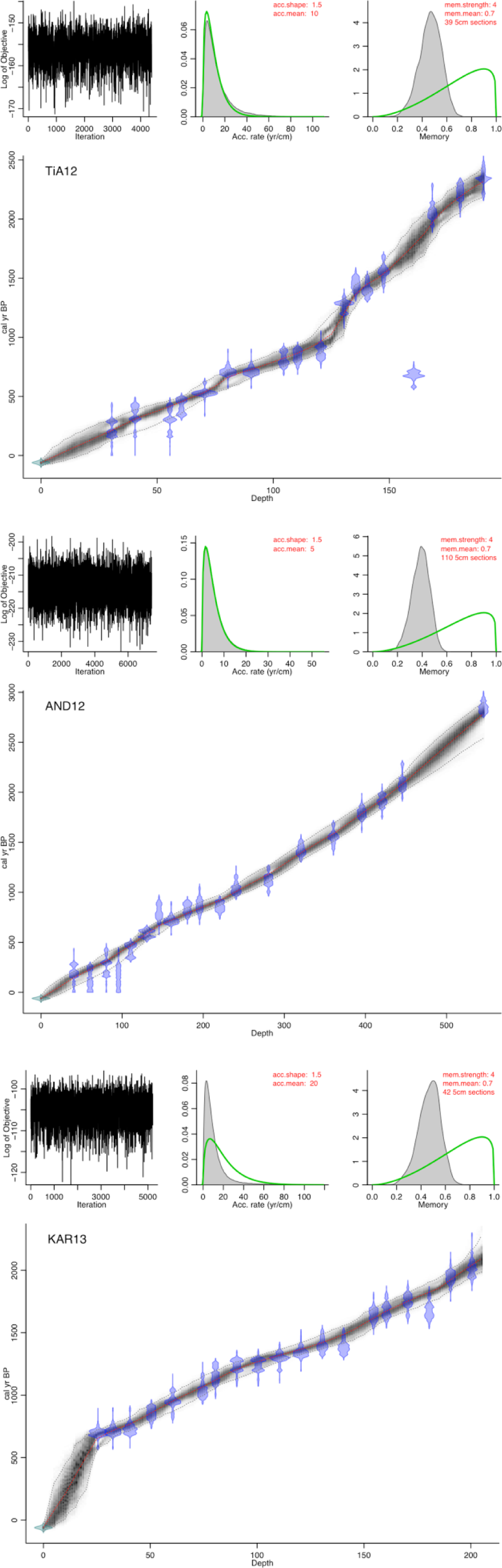
Age–depth models for each core. Top left panel shows the stability of Markov Chain Monte Carlo iterations; middle panel shows the prior and posterior (grey) histogram distributions for the accumulation rate. Right hand panel: distributions for autocorrelation between neighbouring depths.
WTD reconstructions
Application of the transfer function to the testate amoeba assemblages shows that during the last 2000 years, inferred WTD variations have been relatively minor, with the exception of the uppermost section of each of the cores, where a sharp drop in water tables is reconstructed (Figure 6). Whereas water tables were generally between 0 and 20 cm from the surface during the last 2000 years, 20th-century values decreased from 20- to 60-cm depth in all cores. Given the associated errors in the order of ±15 cm for the major part of each sequence, and the fact that the WTD reconstructions did not explain any significant variance in amoeba assemblages, most minor fluctuations reconstructed, that is, those in the order of 5–10 cm may be unreliable reflections of real change. However, some shifts may have been captured by our reconstructions. TiA12 shows a shift to lower water tables ~1450 cal. BP, a wet shift ~1380 cal. BP, a dry shift ~930 cal. BP and a sharp dry shift near the top of the sequence. None of the fluctuations in WTD reconstructed in AND12 appear reliable, as they do not exceed the associated error range, with the exception of the near-surface drop in water tables. KAR13 shows somewhat drier conditions between 1540 and 1370 cal. BP, followed by a slight increase in water tables culminating at 1010 cal. BP. Much more pronounced is a sharp recent dry shift that was dated around the first half of the 19th century in KAR13, although the age–depth model of this core shows a wide age uncertainty for this depth (Figure 5).
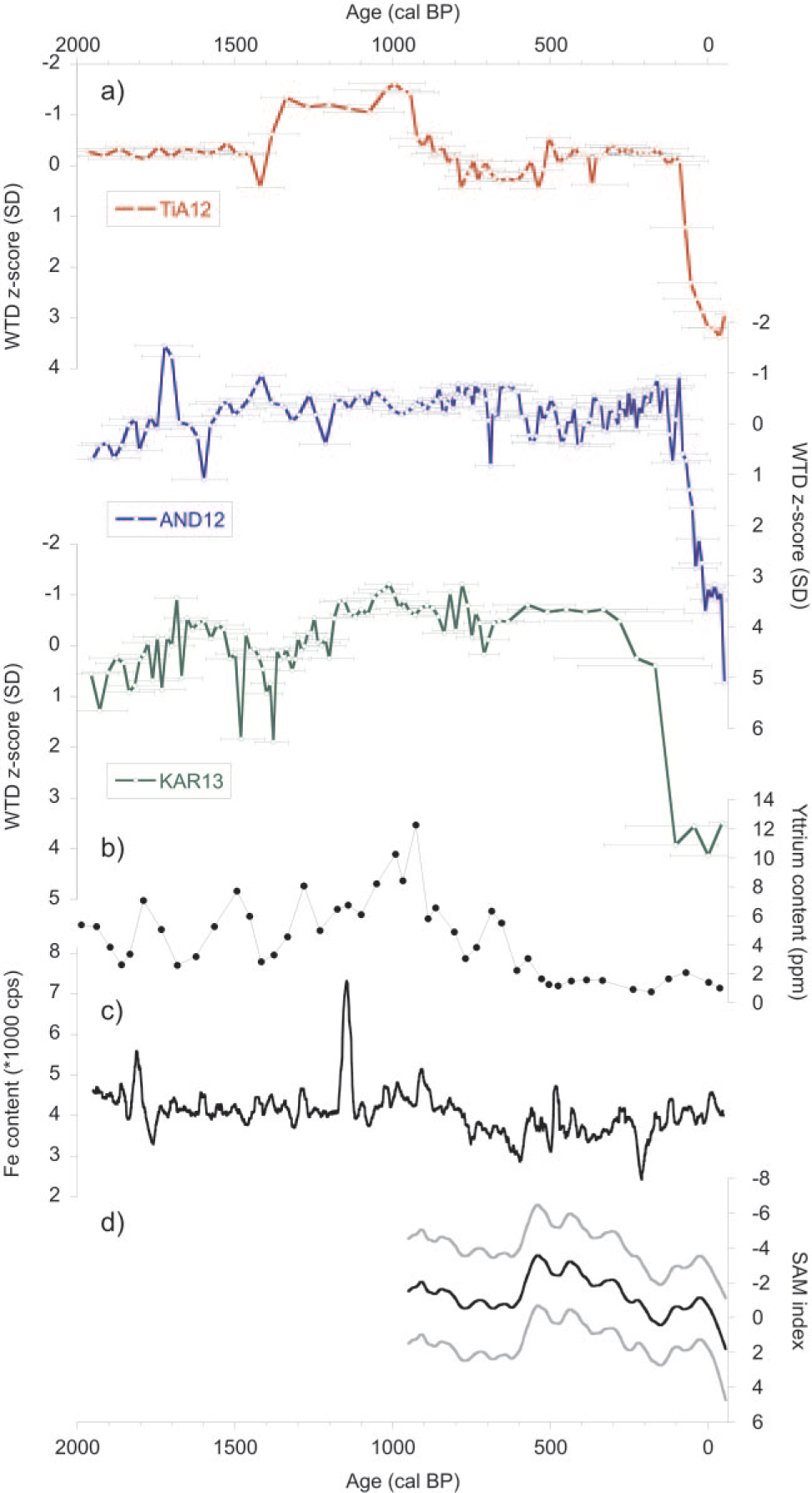
(a) Standardised WTD reconstructions for the last 2000 years for each core. Horizontal bars represent the chronological error for each sample; (b) Yttrium content from stalagmites on the western side of the Andes (Schimpf et al., 2011); (c) Fe content from lake records from Lago Fagnano (Waldmann et al., 2010); (d) SAM reconstruction (70-year LOESS; Abram et al., 2014). Graphs (b) and (c) represent wetter conditions along the vertical axis.
Discussion
Linking testate amoeba assemblages and climate change
Reconstructing past climate variables using testate amoeba records from peat bogs implies assumptions about relationships between (1) surface wetness and testate amoeba assemblages and (2) climate and peat bog hydrology. The former assumption is justified here by the study of near-surface assemblages and the associations with field-measured environmental conditions which showed a significant relationship expressed in the transfer function (van Bellen et al., 2014). Although multiple environmental variables affect testate amoeba assemblages, variables quantifying some measure of surface humidity, for example, WTD, generally explain the most variability. In many studies of modern testate amoeba ecology, WTD alone explains a minor part of the variation in assemblages (Amesbury et al., 2013; Charman et al., 2007; Lamarre et al., 2013; Payne, 2013; van Bellen et al., 2014), although WTD still represents the most important ‘measurable’ environmental variable. This phenomenon is not uncommon in ecology and should normally still allow a quantification of past water tables from fossil assemblages.
Assumption 2, regarding the nature and stability of the relationship between climate and hydrological conditions, was not studied specifically here. A direct water input from the watersheds surrounding the bogs may be excluded, because this would have been reflected along the cores by shifts in trophic status and, as a result, vegetation. Previous studies generally identify summer water deficit, expressed as precipitation minus evapotranspiration as the main driver for peatland water-table dynamics in temperate, northwestern Europe (Charman, 2007; Charman et al., 2009) and continental northern United States (Booth, 2010). In this balance, precipitation plays a more dominant role than temperature. Since the Fuegian bogs seem to develop currently under relatively warm-dry conditions (Loisel and Yu, 2013), they may have been more sensitive to any warm-dry shifts than to cold-wet shifts during the late-Holocene. For these reasons, the WTD records presented here should not be interpreted as simple linear representations of past water deficit. Further studies are required to assess the drivers of hydrological change in South American bogs as these may not be exactly equivalent to Northern Hemisphere examples.
Testate amoeba assemblages and WTD reconstructions
The WTD reconstructions presented here have relatively high uncertainty with poor abundance-weighted correlation between RDA Axis 1 scores and weighted-averaging optima for AND12 and KAR13 (Figure 4) and a weak transfer function performance. The transfer function was created from samples originating from the same sites and testate amoeba analyses were performed by SvB in both studies; therefore, taxonomic inconsistency is unlikely to be significant (Payne et al., 2011). Payne (2011) recommended limiting the application of transfer functions to fossil samples accumulated under the same trophic conditions as those covered by the training set. Preliminary plant macrofossil analyses for each of the cores showed that the sampled sections accumulated under ombrotrophic conditions, with a strong dominance of S. magellanicum during the entire 2000-year length of the records. One section in KAR13 suggested an episode of S. fimbriatum dominance between 154- and 122-cm depth (~1620–1370 cal. BP). S. fimbriatum has a tendency to build hummocks under dry and slightly minerotrophic conditions with relatively high levels of disturbance (Laine et al., 2011). This section coincides with occasional presence of A. muscorum and Alocodera cockayni (Figure 2). Ordinations and correlation with palaeoSig showed that fossil assemblages might not have been mainly driven by a hydrology-related variable, or at least not during the entire late-Holocene period considered here (Figure 4). An alternative explanation for the non-significant randomTF result may be the very low long-term species turnover, resulting from low species diversity and low long-term variability in assemblages, observed in each of the records and in AND12 in particular. However, this does not exclude the possibility that some important changes in testate amoeba assemblages were indeed driven by shifts in WTD.
The major decrease in reconstructed water tables at the top of the records is mainly because of the dominance of A. muscorum, and to a lesser extent Corythion-Trinema type and E. tuberculata type at these levels. Based on direct comparisons between measured WTD and contemporary testate amoeba assemblages, Swindles et al. (2015a) concluded that reconstructed magnitudes in WTD should be interpreted with caution, especially for reconstructions at the dry end of the gradient. This corroborates the independent validation results of the transfer function which showed a higher root mean square error of prediction at the dry end (van Bellen et al., 2014).
A. muscorum is considered resistant to decay (Mitchell et al., 2008; Swindles and Roe, 2007) and therefore it is unlikely that its rarity or absence in fossil samples is the result of a preservation bias. Each of the records shows evidence of a minor A. muscorum presence in Tierra del Fuego before the 20th century, excluding the possibility of a recent introduction of this species into the region (cf. Wilkinson, 2010). One possibility is that the relative abundance of A. muscorum in the near-surface samples may have been enhanced by increased intensity of UV-B radiation because of stratospheric ozone depletion. Field experiments have shown that assemblages of living testate amoeba in Fuegian peatlands are sensitive to UV-B radiation, although 99% of light may be attenuated at 6-mm depth in the Sphagnum carpet (Searles et al., 1999, 2001). In one experiment, Robson et al. (2005) stretched two types of plastic film over a total of 20 sample plots which created either ‘near-ambient’ or ‘reduced’ UV-B conditions. This setup was maintained for 6 years and testate amoeba assemblages were subsampled several times per growing season. Near-ambient surface UV-B was estimated to be relatively beneficial to A. muscorum at the 0- to 0.5-cm depth interval, with an ~50% decrease in absolute numbers of A. muscorum under reduced UV-B conditions (Robson et al., 2005). Thus, the high dominance of A. muscorum in the near-surface samples may be partly explained by a relatively enhanced UV-B and therefore the reconstructed water tables for these levels may be too low. Preliminary results from independent carbon isotope analysis of Sphagnum cellulose, which allows for a reconstruction of changes in bog hydrology (WTD) and relative humidity, support this hypothesis.
In order to test this effect on the reconstructions, we reduced by 50% the presence of A. muscorum in the transfer function. Both adjusted (50%-reduced A. muscorum) and original fossil records were used as input to infer WTD. None of the reconstructed WTD based on the artificially reduced A. muscorum presence showed marked differences compared with the original reconstructions, which suggests the potential influence of UV-B on A. muscorum presence since the 1970s may not have been important enough to affect the inferred WTD from subfossil samples (Figure 7). In studies from around the globe, A. muscorum has been widely recognised as a xerophilous taxon (Amesbury et al., 2013; Booth, 2008; Charman et al., 2007; Lamarre et al., 2013; Qin et al., 2013).
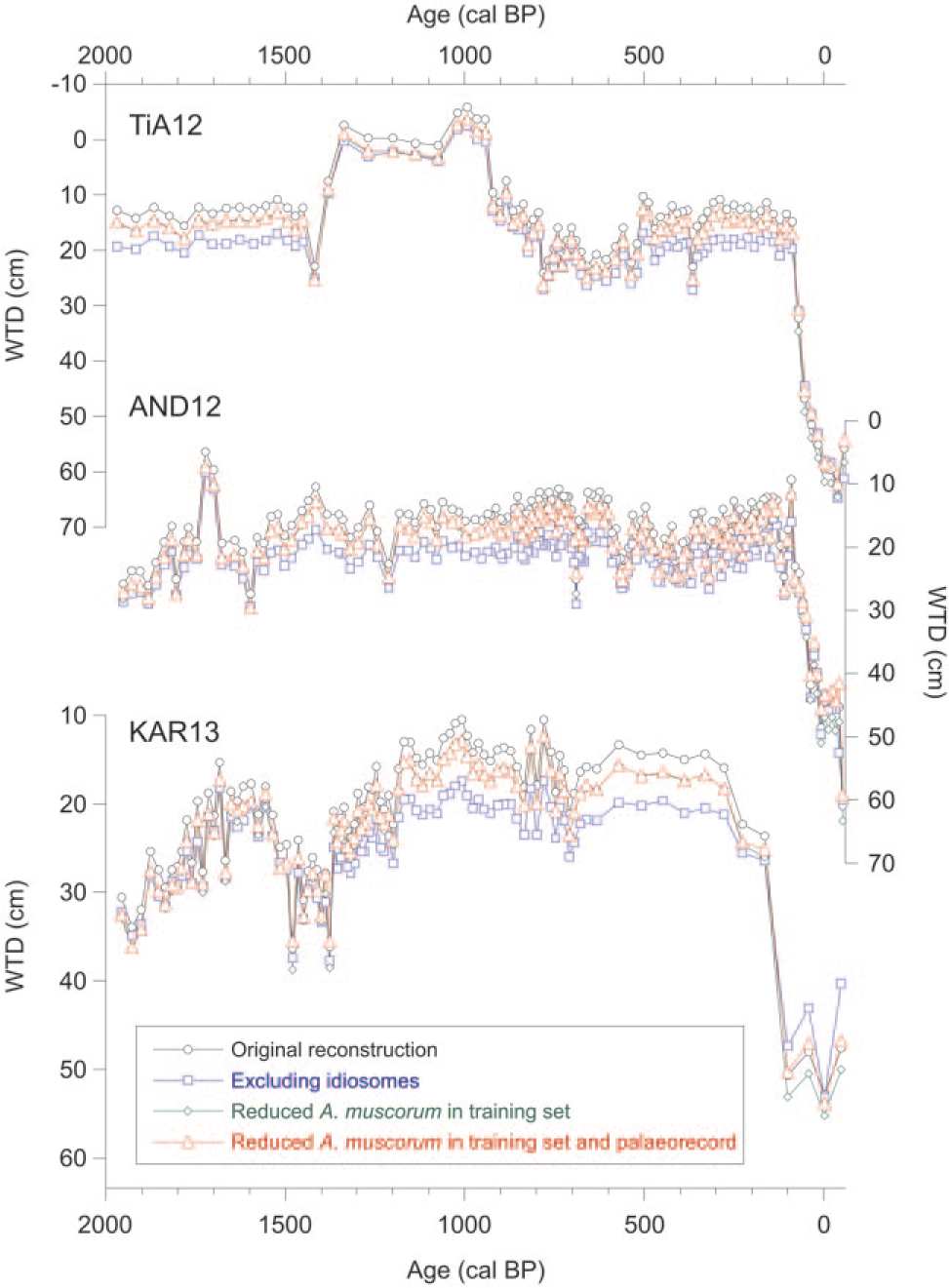
Comparisons of the original reconstruction and reconstructions based on various versions of the transfer function and palaeorecords.
Most idiosomic tests, including those of Corythion-Trinema type and E. tuberculata type, are composed of biosilica and may be poorly preserved in peat (Mitchell et al., 2008). To address possible issues with differential preservation, we reconstructed WTD after excluding from the transfer function the decay-sensitive idiosomes (most Euglyphida, including Corythion-Trinema type and Sphenoderia, but not Assulina). As expected, excluding idiosomes resulted in a slightly drier reconstruction for most of the last 2000 years, except for the top section (Figure 7). This may be explained by the fact that most idiosomes are rather xerophilous. Excluding these in the training set, dry microsites become represented by non-idiosomic taxa which were otherwise associated with wetter microsites. As a result, the modelled optimum WTD of these non-idiosomic taxa shifts towards the dry end of the gradient. Abundance of non-idiosomes in fossil samples then results in a drier reconstruction, but especially so for the levels where idiosomes are absent and non-idiosomes abundant. Most important, however, is that the reconstructed recent dry shift was reproduced excluding idiosomes.
Additional uncertainty relating to the inferred WTD changes may result from the important presence of D. pulex. This species has a poorly established ecology, because it is relatively rare in surface samples but often dominant in fossil assemblages. Its optimal WTD and tolerance were quantified by van Bellen et al. (2014) occupying an intermediate position along the water-table gradient, which agrees with its apparent relative position in Amazonian peatlands (Swindles et al., 2014). However, the rareness of D. pulex in southern Patagonian surface samples relative to its dominance in fossil samples (Figure 8) leads to a relatively large error in quantitative reconstructions.
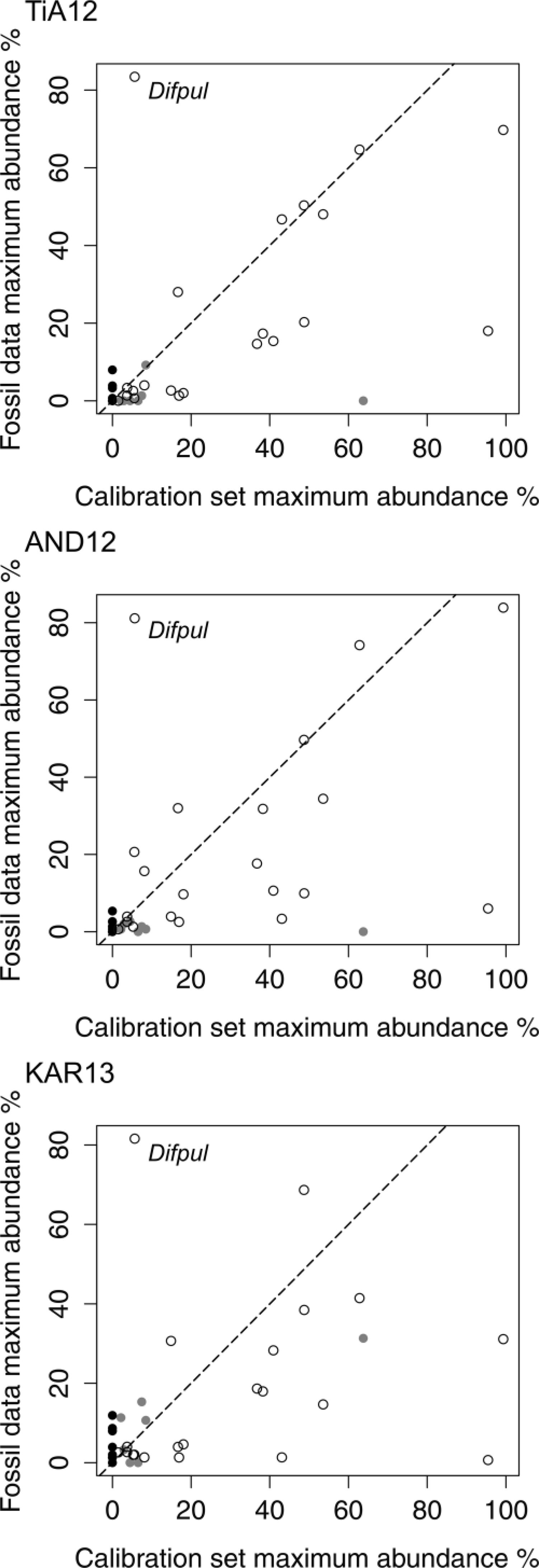
Correlation between the maximum abundance of taxa in the calibration set and in each core. Taxa absent from the calibration set are shown in black and taxa with a Hill’s N2 lower than 5 are shown in grey. Difpul = Difflugia pulex.
Quantification of testate amoeba influx (expressed as number of tests cm−2 yr−1) in future studies may provide an understanding of the importance of changes in UV-B radiation, because enhanced UV-B radiation is reported to positively affect testate amoebae, possibly because of leaching of nutrients from leaf cells and changes in Sphagnum leaflet morphology (Searles et al., 2001).
Matching modern and fossil assemblages
The microtopographical gradients currently observed in Fuegian peatlands are among the highest globally for temperate bogs as reflected by measured WTD ranges which exceeded 100 cm along a single transect during the austral summer of 2012 (van Bellen et al., 2014). These long gradients may be inconsistent with theory, because hummocks, lawns and drier hollows are all dominated by S. magellanicum, with S. falcatulum fringing the pools. The potential of S. magellanicum to sustain the entire microtopographical gradient challenges theories which support the influence of Sphagnum species-specific production and decay rates on peat bog microtopography (Nungesser, 2003). It should be noted that the dominance of S. magellanicum appears independent of any human disturbance, as it was equally observed in both palaeorecords and field observations over a wider region (Grootjans et al., 2010; Mauquoy et al., 2004). The wide range in WTD measured in the field explains both the low water-table optimum of A. muscorum exceeding 50-cm depth and the inferred deep water tables for the recent decades.
Fuegian testate amoeba assemblages are relatively species-poor in both the fossil dataset and surface samples (van Bellen et al., 2014) compared with datasets from Northern Hemisphere peat bogs (Booth, 2002; Lamentowicz and Mitchell, 2005), although comparisons between datasets are complicated by variations in the timespan represented by individual samples and records and variations in trophic status. Testate amoeba species richness has been shown to decrease with increasing latitude in the Southern Hemisphere (Smith, 1982, 1996), with very few taxa identified in Antarctica (Mieczan and Adamczuk, 2014; Royles et al., 2013; Smith, 1992). This may be the result of limited dispersal mechanisms, because the dominance of the SWWB, together with the sparseness of terrestrial habitats west of the South American continent may reduce the potential for (latitudinal) dispersal of testate amoebae towards southern Patagonia (Wilkinson et al., 2012). The relatively uniform species richness and similarity of fossil assemblages between sites in this study suggests the dominance of large-scale controls on testate amoeba dispersal.
Late-Holocene WTD reconstructions and climate
In line with relatively stable testate amoeba assemblages, reconstructed WTDs show relatively little variation for centuries, notably between 800 and 100 cal. BP (Figure 6). The absence of consistent changes in the peatland WTD records presented here highlights the potential importance of other, locally acting variables on WTD dynamics, or the influence of autogenic change and ecohydrological feedback mechanisms which may have affected the peatland sensitivity to external forcing (Swindles et al., 2012). The suggested higher sensitivity to warm-dry shifts, compared with cold-wet shifts, may be one example of non-linear response.
Nevertheless, we reconstructed a drier period between 1500 and 1400 cal. BP which coincides with the warm-dry conditions observed in a record from Lago Cipreses, ~400 km NW of our study region (Moreno et al., 2014). This period was followed by a wet shift centred ~1400 cal. BP which has been characterised globally by cooler climatic conditions, possibly linked with a meteorite flux or volcanic eruptions that affected climate (Baillie, 2007; Ferris et al., 2011; Larsen et al., 2008). These cooler conditions may have been the trigger for the wet shift observed in TiA12 and to a lesser extent in KAR13. Interestingly, the period often associated with the Medieval Climate Anomaly (MCA) was characterised in TiA12 and KAR13 by a maximum in observed water levels, culminating at ~1010 cal. BP. Increased precipitation was also reconstructed from different proxies on both sides of the Andes during this period. Speleothem records from the western flank of the Cordillera near 53°S show maximum precipitation between 1000 and 900 cal. BP (Schimpf et al., 2011), while increasing Fe content in lake records from Lago Fagnano suggests regional wetting east of the Andes (Waldmann et al., 2010; Figure 6).
Finally, we reconstructed a slight decrease in water-table levels towards the onset of the ‘Little Ice Age’ (LIA), which was most pronounced in TiA12. Using plant macrofossil and peat humification analyses, a decrease in local water table was also reconstructed around 490 cal. BP in Andorra bog (Chambers et al., 2014). Speleothem records suggest that the transition from MCA to LIA may have been characterised by an ~30% decrease in precipitation (Schimpf et al., 2011).
Recent change
Notwithstanding the aforementioned effect of enhanced UV-B radiation on testate amoebae, our data suggest that the nature of the response of testate amoeba communities to environmentally influenced changes in peat bog hydrology during recent decades is unlike any experienced during the last 2000 years. The reconstructed recent drop in water table is the only late-Holocene shift observed in all three of the records. SAM reconstructions show a clear increase starting in the first half of the 20th century (Figure 6) and a positive SAM is generally associated with an increased influence of SWWB over the study region (Archer and Caldeira, 2008). The near-surface drying trend coinciding with an increased SWWB activity suggests an ‘eastern response’, that is, a negative correlation between SWWB intensity and precipitation, of Tierra del Fuego peatlands to the increased westerlies. This interpretation corroborates previous findings from various types of proxy records from southern Patagonia. Villalba et al. (2012) reconstructed suppressed Nothofagus betuloides growth, forced by warm-dry conditions since the 1950s in Tierra del Fuego. This trend was associated with stratospheric ozone depletion and a persistent positive phase of the SAM. Lacustrine records of pollen and charcoal from southern Patagonia show changes in regional vegetation towards dominance of herbs and ferns to the detriment of Nothofagus, together with an increase in charcoal presence suggesting that fires became more important since 1890 CE (Moreno et al., 2014). Other proxy records suggest a more recent warming effect starting during the 1970s. Strelin and Iturraspe (2007) reconstructed glacier retreat of the Cordón Martial glacier, located 6 km southwest of Andorra bog, accelerating from the 1970s. A glacier record from Gran Campo Nevado, at the western side of the Andes, was interpreted as reflecting warmer and wetter conditions between 1984 and 2000 CE (Möller et al., 2007). It must be noted, however, that 20th-century glacier dynamics have been reported to be very sensitive to orographic effects on microclimate in southern Patagonia, with differing trends in dynamics between north- and south(west)-facing slopes due to strong gradients in precipitation (Holmlund and Fuenzalida, 1995).
Although the age–depth models presented here are generally well resolved, the near-surface shift shows a relatively wide age uncertainty, especially in KAR13. Age–depth models based only on 14C dates generally struggle to represent accurately the near-surface accumulation because of the presence of a plateau in the calibration curve during this period and a general increase in the ‘apparent’ peat accumulation rate near the surface associated with the presence of relatively undecomposed acrotelm peat. Current TiA12 and AND12 records and chronologies suggest the dry shift occurred sometime between 1850 and 1900 CE, yet the shift in KAR13 was very poorly chronologically resolved and therefore dated to one or several centuries earlier with an error range of up to 500 years (Figure 6). For now, we cannot exclude the possibility that the sharp shift in amoeba assemblages and associated drop in water table occurred anytime during the 20th century. More precise dating techniques may be applied in an upcoming paper to improve the chronological constraints on the reconstructed trend.
SAM and SWWB influence on Tierra del Fuego climate
The poor relationship between past SAM/SWWB and WTD records from peat bogs may be the result of the varying direction in the relationship between SAM and southern Patagonian climate. Within a timeframe covering the last 50 years, changes occurred in the typical hemispheric circulation pattern associated with austral spring SAM (Silvestri and Vera, 2009). In addition, SAM dynamics during the austral summer have been affected over the last several decades by the depletion of stratospheric ozone (Fogt et al., 2009; Thompson et al., 2011). We suggest examining sites at the northern boundary of the SWWB to increase the understanding of late-Holocene SWWB dynamics in terms of latitudinal shifts.
Conclusion
The three peatlands studied here record relatively minor fluctuations in WTD during the last 2000 years. It is uncertain whether this is due to minor precipitation, temperature or evapotranspiration changes in the core region of the SWWB during the last 2000 years or species-poor testate amoebae assemblages in Tierra del Fuego peat deposits. A wet shift around 1400 cal. BP was found in both TiA12 and KAR13 and coincided with a previously reported climatic event. These cores also record high water tables at ~1010 cal. BP. AND12, in contrast, showed few changes in WTD during the last 2000 years except for the near-surface ~40-cm drop in water table which remains poorly chronologically constrained. The generalised near-surface shift may partly result from an increase in UV-B radiation coinciding with the drought effect on testate amoeba assemblages, to the benefit of xerophilous A. muscorum. However, this recent increased dryness may well have been forced primarily by drier and warmer conditions, possibly driven by a more dominant positive SAM phase and an increase in the importance of the SWWB in this region, as it corresponds to trends reconstructed from other proxies in Tierra del Fuego.
Footnotes
Acknowledgements
We thank Ricardo Muza and the Wildlife Conservation Society (WCS) Karukinka Park rangers for facilitating access to Karukinka Park. We also thank François De Vleeschouwer, Gaël Le Roux, Heleen Vanneste, Sébastien Bertrand, Zakaria Ghazoui and Jean-Yves De Vleeschouwer for fieldwork assistance. Nelson Bahamonde (INIA, Punta Arenas, Chile) and Ernesto Teneb (UMag, Punta Arenas, Chile) provided logistical support for the fieldwork in Chile. Dr Andrea Coronato (CADIC, Ushuaia) kindly provided logistical support for the research in Argentina. We also thank Ramiro Lopez for fieldwork assistance, Claudio Roig for access and permission to sample Tierra Australis and Audrey Innes for laboratory assistance. We appreciate the critical reviews by Graeme Swindles and an anonymous reviewer which improved the manuscript.
Funding
This work was supported by the Natural Environment Research Council (grant numbers NE/I022809/1, NE/I022981/1, NE/I022833/1 and NE/I023104/1).