
Abstract
Forest clearance is one of the main drivers of soil erosion and hydrological changes in mires, although climate may also play a significant role. Because of the wide range of factors involved, understanding these complex links requires long-term multi-proxy approaches and research on the best proxies to focus. A peat core from NW Spain (Cruz do Bocelo mire), spanning the last ~3000 years, has been studied at high resolution by physical (density and loss on ignition (LOI)), geochemical (elemental composition) and palynological (pollen and non-pollen palynomorphs) analyses. Proxies related to mineral matter fluxes from the catchment (lithogenic tracers, Glomus and Entorrhiza), rainfall (Bromine), mire hydrology (HdV-18), human pressure (Cerealia-type, nitrophilous taxa and coprophilous fungi) and forest cover (mesophilous tree taxa) were the most useful to reconstruct the evolution of the mire and its catchment. Forest clearance for farming was one of the main drivers of environmental change from at least the local Iron Age (~2685 cal. yr BP) onwards. The most intense phase of deforestation occurred during Roman and Germanic times and the late Middle Ages. During these phases, the entire catchment was affected, resulting in enhanced soil erosion and severe hydrological modifications of the mire. Climate, especially rainfall, may have also accelerated these processes during wetter periods. However, it is noteworthy that the hydrology of the mire seems to have been insensitive to rainfall variations when mesophilous forest dominated. Abrupt changes were only detected once intense forest clearance commenced during the Iron Age/Roman transition (~2190 cal. yr BP) phase, which represented a tipping point in catchment’s ability to buffer impacts. Overall, our findings highlight the importance of studying ecosystems’ long-term trajectories and catchment-wide processes when implementing mire habitat protection measures.
Keywords
Introduction
During the Holocene, and more specifically in the late Holocene, human activities have played, directly or indirectly, an increasing role in the evolution of terrestrial ecosystems. One of the key processes is deforestation (Williams, 2000). Evidence of small-scale human disturbance to woodland cover by hunter–gatherers has been already detected during the Mesolithic (Brown, 1997; Innes and Blackford, 2003; Innes and Simmons, 1988; Schuldenrein, 1986; Siiriäinen, 1980; Smith, 1970; Williams, 2000). More widespread forest clearance for cultivation and grazing has resulted in land degradation from at least the Neolithic onwards (Carrión et al., 2010a; Mazoyer and Roudart, 2006; Starkel, 2005). Late-Holocene climate change, although weaker in amplitude than the dramatic shifts that occurred in the last glacial cycle, has been shown to be larger and more frequent than commonly recognized (Mayewski et al., 2004), and has also influenced environmental change.
Soil erosion is becoming one of the most significant geomorphic processes acting at the Earth’s surface (Pimentel, 2006; Wilkinson and McElroy, 2007). Soil erosion is largely caused by human activity and climate. Rain and wind determine climatic erosivity, whereas air temperature controls the occurrence of frost, snowfall, snowmelt and soil moisture, the latter affecting the susceptibility to soil erosion (Boardman and Poesen, 2006). However, today in Europe, the principal causes of soil erosion are agricultural practices, deforestation, overgrazing and construction, all of which are strongly influenced by land use and policy (Boardman and Poesen, 2006; Grimm et al., 2002). Globally, moderate to severe soil degradation affects almost 2000 million hectares of arable and grazing land, an area larger than that of the United States and Mexico combined (FAO, 1995), and has become a serious public health issue (Pimentel, 2006), resulting in increased awareness among scientists and policy-makers. To fully evaluate the importance of soil erosion, a long-term perspective is needed, as it could provide insights on how ecosystems shift in response to soil erosion and the relative contributions of climate and human transformations. Significant soil erosion as a consequence of human activities and climate has been detected in many parts of the world. Examples include the early Neolithic in the Peloponnesus Peninsula, where high sedimentation rates suggest that Holocene soil erosion was triggered by human activity and amplified by precipitation (Fuchs, 2007; Fuchs et al., 2004) and from the late Bronze Age in the Drama Basin of Macedonia, where deforestation and agricultural activities made the river system less resilient to natural soil erosion and more sensitive to small changes in climate (Lespez, 2003).
Most of the long-term studies on soil erosion have been undertaken on polycyclic, colluvial or alluvial soils (Benito et al., 1991; Costa Casais et al., 2009; Dreibrodt et al., 2009; Fuchs, 2007; Fuchs et al., 2004; Kaiser et al., 2007; Kirch, 1996; Lespez, 2003; Martínez Cortizas et al., 2000, 2009b; Rochette Cordeiro, 1992; Van Andel et al., 2013; Zádorová et al., 2013). However, peatlands and lakes can also be used to infer erosion as they are good archives of dust/sediment fluxes (Foster et al., 2000; Giguet-Covex et al., 2011; Hölzer, 1998; Le Roux et al., 2012; Lomas-Clarke and Barber, 2004; López-Merino et al., 2010; Martínez Cortizas et al., 2005; Schofield et al., 2010; Shotyk et al., 2001; Simonneau et al., 2013). Moreover, they offer ideal conditions for the preservation of geochemical and palynological proxies to infer environmental changes.
Most palaeoenvironmental reconstructions, especially geochemical studies, use ombrotrophic mires, in which inorganic inputs are derived exclusively from atmospheric deposition (Clymo, 1987). Although less studied, minerotrophic mires can also be used to reconstruct soil erosion at catchment scale since their inorganic inputs are commonly derived from rock–water interactions in the surrounding soils and underlying sediments (Shotyk, 2002). Moreover, they also offer suitable conditions to evaluate variations in water run-off because of disturbances in their catchment. Understanding how disturbances can affect mires, which maintain high levels of biodiversity and are important carbon sinks, could provide additional insights in order to manage such sensitive ecosystems and to prevent further damage.
In this study, we analyse the palaeoenvironmental evolution of a minerotrophic mire located in NW Spain, focusing on the link between climate, human activities, forest evolution, soil erosion and changes in local hydrology. Previous palaeoenvironmental studies in the area performed on peat records mainly focused on vegetation change and peatland inception (Aira Rodríguez et al., 1994; Taboada Castro et al., 1993, 1996). Here, we present a higher resolution multi-proxy study combining physical, geochemical and palynological analyses, as well as a multivariate statistical approach aiming to (1) decipher how late-Holocene anthropogenic transformations and climate change affected both the physical and ecological evolution of the mire’s catchment, at both regional and local scales, and (2) get insights into the behaviour of specific palynological and geochemical proxies for soil erosion and peatland moisture.
Material and methods
Study area and sampling
Cruz do Bocelo (42°59′N; 8°01′W; 730 m a.s.l.) is a minerotrophic mire located in the western side of the O Bocelo range (Melide, NW Spain; Figure 1), overlying a geological substratum of cataclastic deformed orthogneiss with granitic composition (García Salinas, 1978). The climate is temperate humid, with average annual temperatures of 12–13°C, and annual precipitation of 1600–1800 mm (Martínez Cortizas and Pérez Alberti, 1999). Present land use consists in an association of pasture, horticultural crops, scrubland (Ulex, Erica and Cytisus species) and afforestations of Pinus pinaster, Eucalyptus globulus and, to a lesser extent, Betula, with scattered patches of Quercus robur and Castanea sativa. The mire is used for grazing cattle and disturbed on one side by the recent construction of a road. Surface vegetation is mainly Sphagnum, Carex, Drosera and Juncaceae species, with Calluna vulgaris, Erica tetralix and Ulex spp. in drier and more degraded areas.
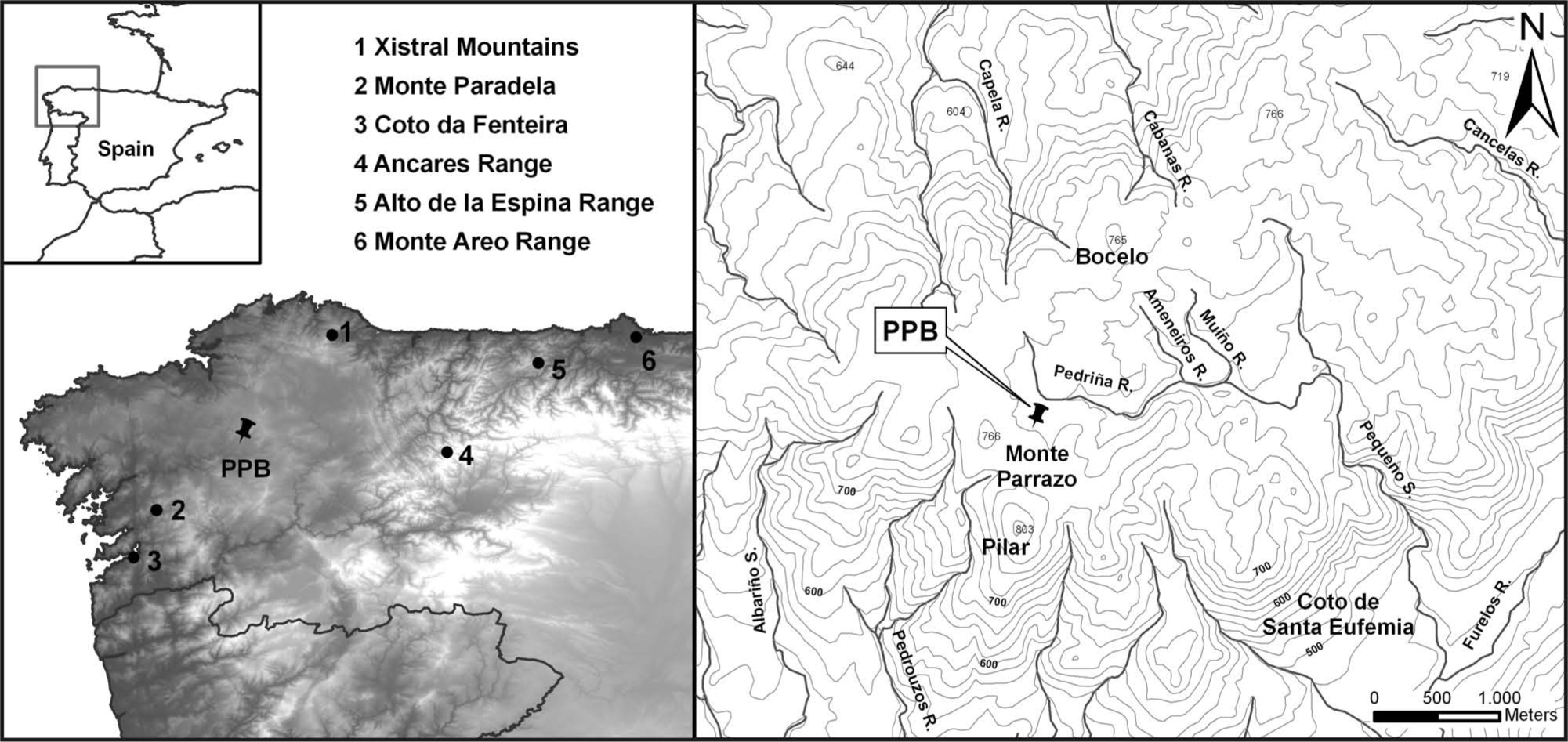
Location of PPB core in Cruz do Bocelo mire (NW Spain) and other places discussed in the text.
A 140-cm-long core (PPB) was collected in 2007 using Waardenar and Russian corers. Below 130 cm, the sediment was a mixture of gravel and sand. The extracted peat sections were protected in plastic guttering and stored under cold conditions (4°C) prior to laboratory analyses. The core was sub-sampled into 2-cm-thick slices, obtaining 70 samples. Sub-samples were taken to measure peat bulk density (BD) and loss on ignition (LOI) and for palynological analysis. The remaining material was dried at 105°C and milled to very fine powder prior to elemental composition analyses.
Physical, geochemical and palynological analyses
BD was determined after drying (105°C) peat plugs to constant weight. The same plugs were then heated at 550°C for 5 h for LOI. Additionally, the concentrations of major and minor (Si, Al, Ti, Ca, K and S), trace lithogenic (Rb, Sr, Zr and Th), redox-sensitive elements (Fe and Mn) and halogens (Cl and Br) were determined using x-ray fluorescence dispersive EMMA-XRF analysers (Cheburkin and Shotyk, 1996). The instruments are hosted at the RIAIDT facility of the University of Santiago de Compostela. The calibration was performed using 26 certified reference materials for organic matrices consisting of tree and vegetable leaves (SRM1515, SRM1547, SRM1575, SRM1570a, SRM1573a, SRM1575a, BCR62), coals (SRM1635, SRM1632b, LECO501020, LECO502433, LECO502435), coke (SRM2718, SRM2719), wheat flour (SRM8436, SRM8437, SRM8438), other plant derivates (SRM8412, SRM8432, SRM8433, BCR129, BCR60), animal derivates (SRM8414, BCR150) and peat (NJV942, NIMT/UOE/FM/001). For the inorganic samples of the base of the core, the calibration included 36 certified reference materials, consisting of rocks and minerals (GSR6, SG1a, SRM1d, SRM278, SRM2780, SRM688, 5365, AGV1, DTS1, SRM607, SRM70a), sands and clays (SRM1413, SRM81a, BCSCRM348, SRM679, SRM97b, SRM98b), ashes (SRM1633a, SRM1633b, SRM2690, SRM2691), soils and sediments (SO2, SO3, SRM2586, BCRCRM277b, LKSD1, LKSD2, MAG1, PACS1, RM8704, SRM1646, SRM1646a, SRM1944, SRM2702, SRM2703) and industrial sludge (SRM2782). Quantification limits were as follows: Si (0.05%), Ti and Fe (0.002%), Al (0.002% for organic; 0.2% for inorganic matrices), Ca (0.002%; 0.01%), K (0.002%; 0.05%), S (0.009%; 0.03%), Zr (0.5 µg/g), Th (2.5 µg/g), Rb (0.5 µg/g; 5 µg/g), Sr (0.5 µg/g; 5 µg/g), Mn (5 µg/g; 30 µg/g), Br (0.5 µg/g; 2 µg/g) and Cl (40 µg/g; 350 µg/g).
The classic methodology (Fægri and Iversen, 1989) with concentration in heavy liquid (Goeury and Beaulieu, 1979) was applied to obtain pollen, spores and non-pollen palynomorphs (NPP). Laboratory work was performed at the Archaeobiology Laboratory of the CCHS (CSIC, Madrid). Pollen counting was conducted at 400× magnification, and at least 500 terrestrial pollen grains (trees, shrubs and herbs) were counted and used for the total land pollen (TLP) sum. Hydro-hygrophytes, fern spores and other NPP counts were excluded from the TLP, although their values are also expressed as percentages of TLP. Identification of pollen types and fern spores was achieved with the aid of keys and atlases (Fægri and Iversen, 1989; Moore et al., 1991; Reille, 1999), while NPP classification follows the nomenclature proposed by the Hugo de Vries (HdV) Laboratory (University of Amsterdam). Microfossil diagrams were drawn using Tilia (Grimm, 1992, 2004). The two basal samples (140–136 cm) were palynologically sterile because of the abundance of mineral matter.
Numerical methods
When dealing with a large set of variables, the use of multivariate statistical approaches help summarize common patterns of variation beyond the raw data and to get insights into the underlying environmental factors.
For geochemical data, principal component analysis (PCA; Hotelling, 1933; Pearson, 1901) was applied using SPSS 15.0, in correlation mode and by applying a varimax rotation. Prior to analysis, the data were standardized (Z-scores) to avoid scaling effects and obtain average-centred distributions (Eriksson et al., 1999). The square of the factor loadings, multiplied by 100, was used as a measure of the explained variance of each variable by each principal component.
For palynological data, constrained incremental sum-of-squares (CONISS) cluster analysis (Grimm, 1987) was performed after applying a square-root transformation and Edwards and Cavalli Sforza’s chord-distance dissimilarity measure to all taxa counts in order to delineate pollen assemblage zones.
Radiocarbon dating and chronology
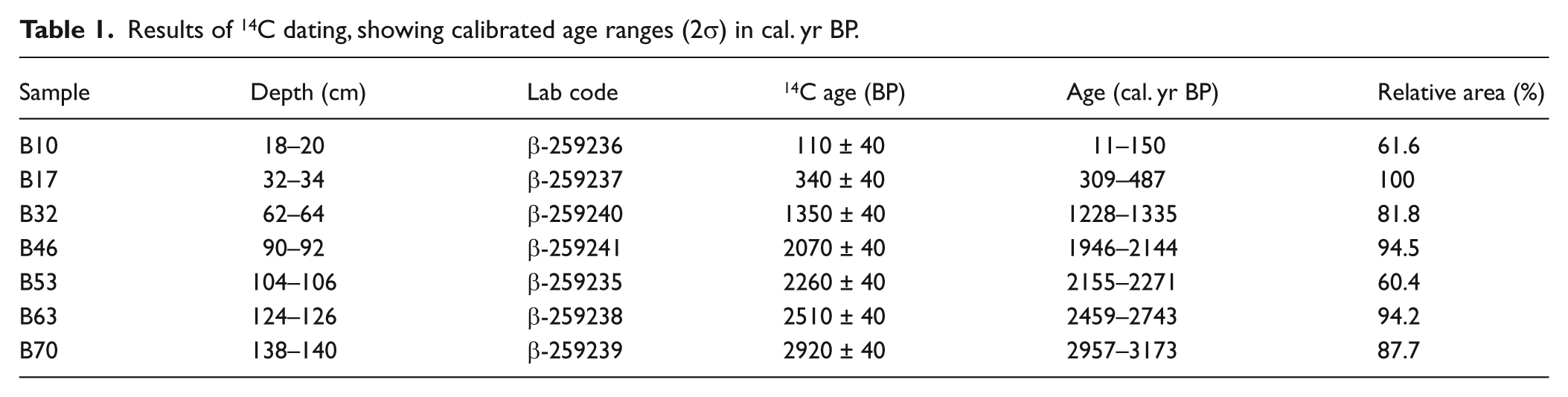
Seven peat samples were sent to Beta Analytic Inc. (Miami, USA) where they were dated by AMS after an acid-wash pretreatment. The obtained 14C dates (Table 1) were calibrated using the IntCaL09.14C curve (Reimer et al., 2009), and an age–depth model was built using Clam.R 1.0 (Blaauw, 2010). The best fit was provided by a smooth-spline solution with a smooth factor of 0.2 (Figure 2). Ages are expressed as calibrated years before present (cal. yr BP) at 2σ level and are set to the year of sampling (2007) by adding the difference from 1950 to all estimated ages. According to this model, the 140-cm sequence represents the last ~3000 cal. yr.
Results of 14C dating, showing calibrated age ranges (2σ) in cal. yr BP.
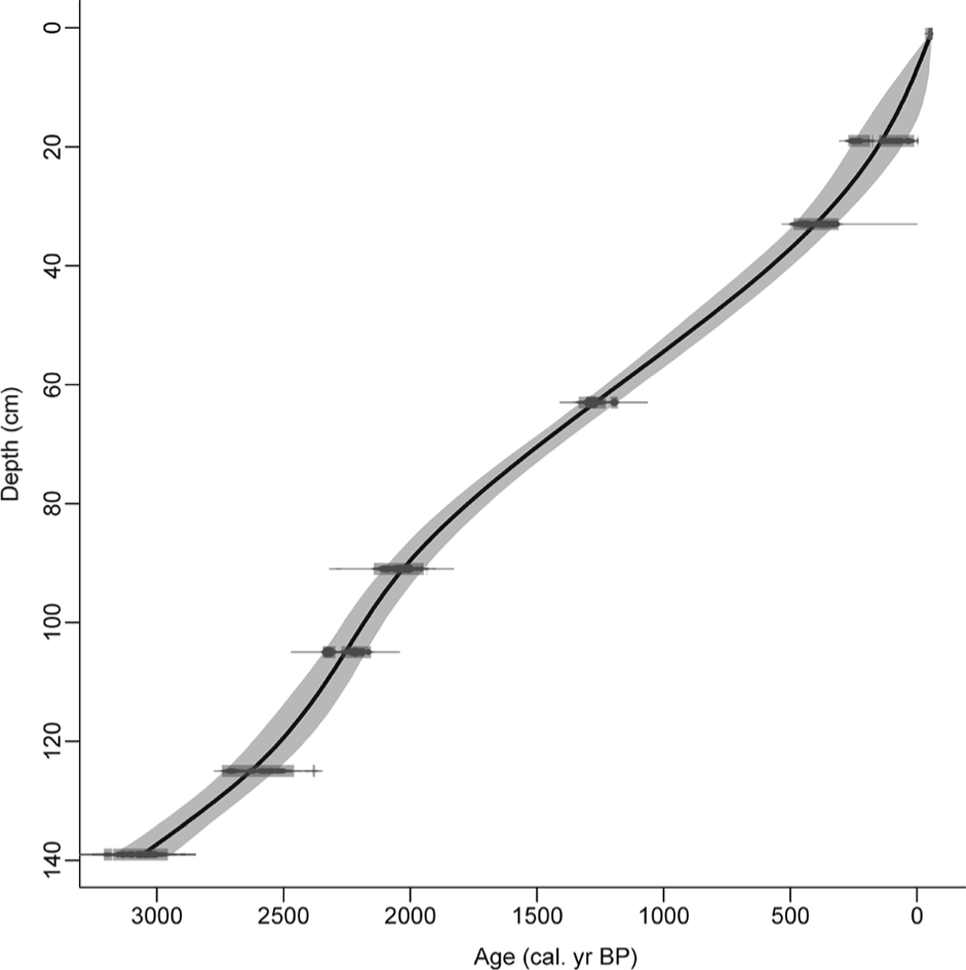
Age–depth model of Cruz do Bocelo mire, fitted with a smooth-spline (smooth = 0.2) using Clam.R (Blaauw, 2010). Blocks in the radiocarbon ages represent the 95% confidence level in radiocarbon dates calibration, and the grey-shaded area the highest density ranges.
Results
Geochemical record
LOI, BD and elemental composition (Figure 3) variations are summarized in three principal components (Figure 4), which explain 84.3% of the total variance. PC1 explains 48.8% of the variance. Aluminium, Rb, Ti, Sr, K, Si, Zr and Th show high positive loadings, BD has a moderately positive loading, and LOI and S have a high negative and a moderate negative factor loading, respectively. The high positive loadings of the lithogenic elements and LOI are indicative of the amount of mineral matter in the peat. Even variables with moderate loadings support this interpretation. One of the main properties controlling peat BD (positive loading) is mineral matter content, and S (negative loading) is a biophilic and organically bound element. The record of factor scores (Figure 4) indicates that the amount of mineral matter is highest at the bottom part of the core (>130 cm) probably because of its proximity to the inorganic basal sediment. Seven sections with higher inputs of inorganic material (E1–E7; Figure 4) occur in the remainder of the core, and they may reflect soil erosion episodes.
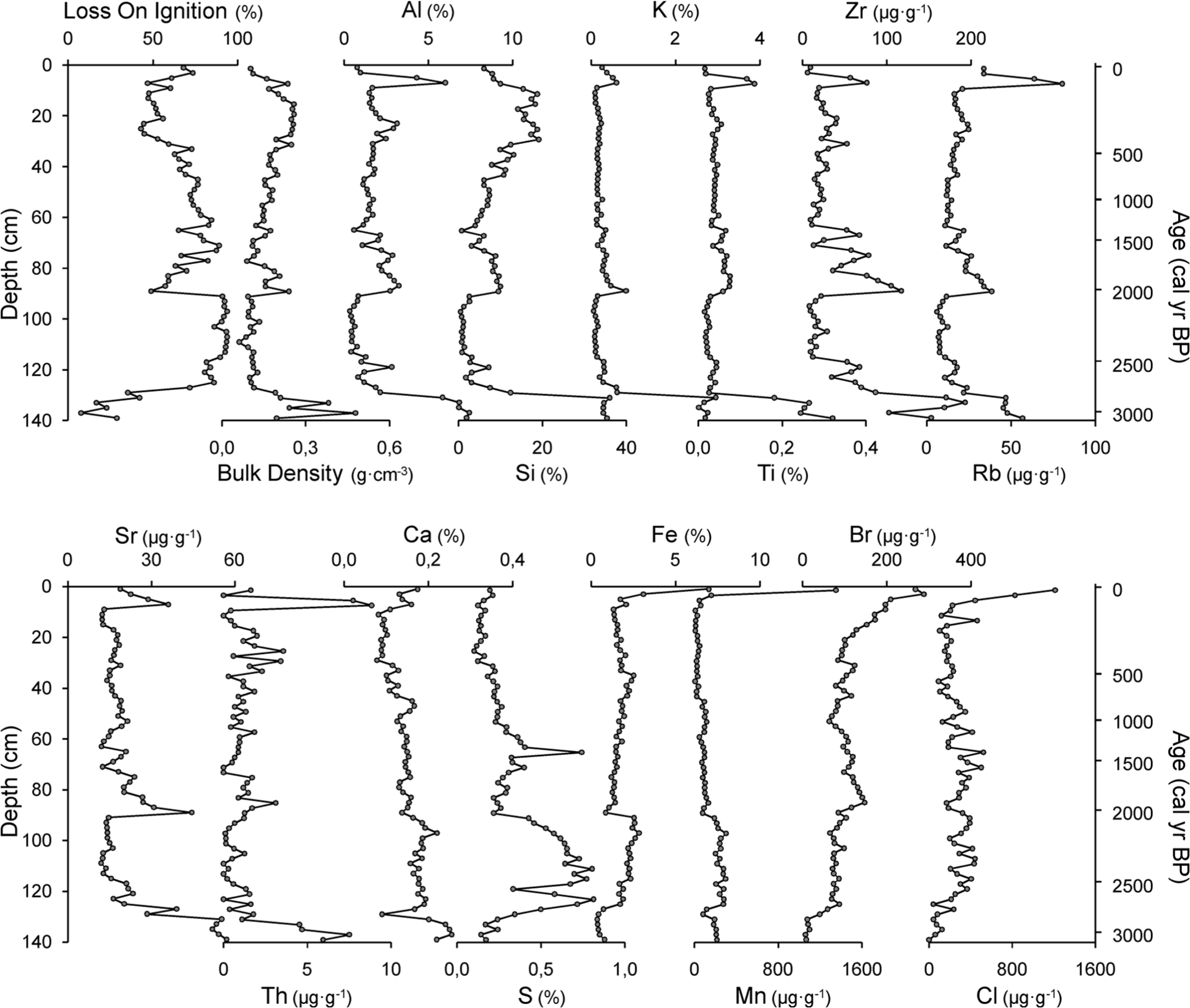
Geochemical results (physical properties and elemental composition) of the PPB core sampled at Cruz do Bocelo mire.
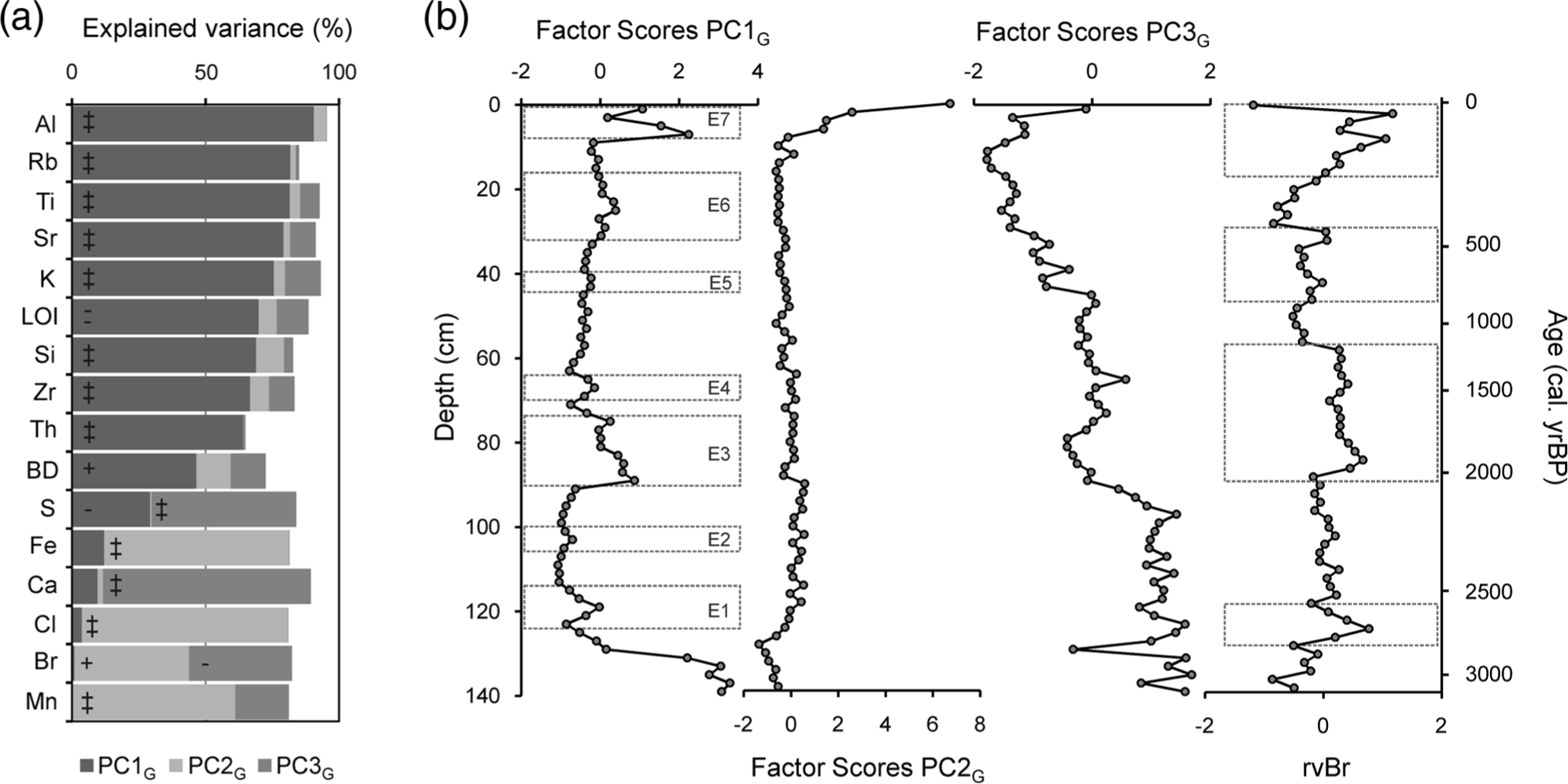
PCA results of geochemical data (physical properties and elemental composition). (a) Percentage of explained variance (square of factor loadings × 100) of the principal components extracted. (b) Records of factor scores of the extracted principal components. ‘‡’: high positive loadings (>0.7); ‘+’: moderate positive loadings (0.5–7); ‘=’: high negative loadings (<−0.7) and ‘-’: moderate negative loadings (>−0.7 and <−0.5).
PC2 explains 19.2% of the variance. Cl, Fe and Mn show high positive loadings, and Br shows moderate positive loading. Factor scores are negative from 140 to 118 cm, and then remain around zero up to 8 cm when their values shift sharply to become positive. Fe and Mn are redox-sensitive elements, while Cl and Br are sourced from the oceans (Kabata-Pendias and Pendias, 2001). Although they share some variance, this is unlikely to be related to a unique source/environmental factor, and it seems to be strongly influenced by their high values at the top of the core. The surface enrichment in Fe and Mn is probably related to oxic conditions in the upper sections of the mire which are more favourable conditions for oxidized, less mobile forms of these elements to form (Chesworth et al., 2006). Halogen accumulation in peat and soils is dependent on atmospheric wet deposition (Johanson et al., 2003), enzymatic halogenation of the organic matter (Biester et al., 2006) and dehalogenation under reducing conditions (Van Pée and Unversuch, 2003).
PC3 explains 16.3% of the variance, and it reflects the inverse relationship between biophilic elements such as Ca and S (high positive factor loadings) and Br (moderate negative loading). The record of the scores can be divided in three sections: >90 cm, with positive scores (except from one sample at 129 cm); 90–42 cm, with scores around zero; and <42 cm, with negative scores.
Most of the variation of Br is explained by PC2 and PC3, but a significant part (18%) remains unexplained (Figure 4). We interpret that PC2 and PC3 are related to the processes controlling halogenation and dehalogenation within the peat. The remaining, unexplained, variation is most likely related to atmospheric wet deposition because there is a good chronological match between the residual Br variation (after eliminating the effect of PC2 and PC3) in the PPB core and previous reconstructions of humidity changes in NW Spain (Mighall et al., 2006 and references therein; see Figure 7).
Palynological record
The regional palynological signal (Figure 5, the complete diagram in supplementary information, Supplementary Figure SI1a, available online) is characterized by the transition from the dominance of arboreal pollen (AP), particularly mesophilous tree taxa, to herbs, indicative of a more open landscape. Shrub percentages remain low throughout the sequence and other trees, like Pinus, only become relevant because of recent afforestation. The local palynological signal (Figure 6; Supplementary Figure SI1b, available online) has relatively stable contributors like Cyperaceae and Filicales monolete and trilete. The record is also characterized by frequent and intense compositional fluctuations affecting Pteridium aquilinum, HdV-18, coprophilous fungi – such as Sordaria-type, Sporormiella-type and Cercophora-type and Entorrhiza (HdV-527).
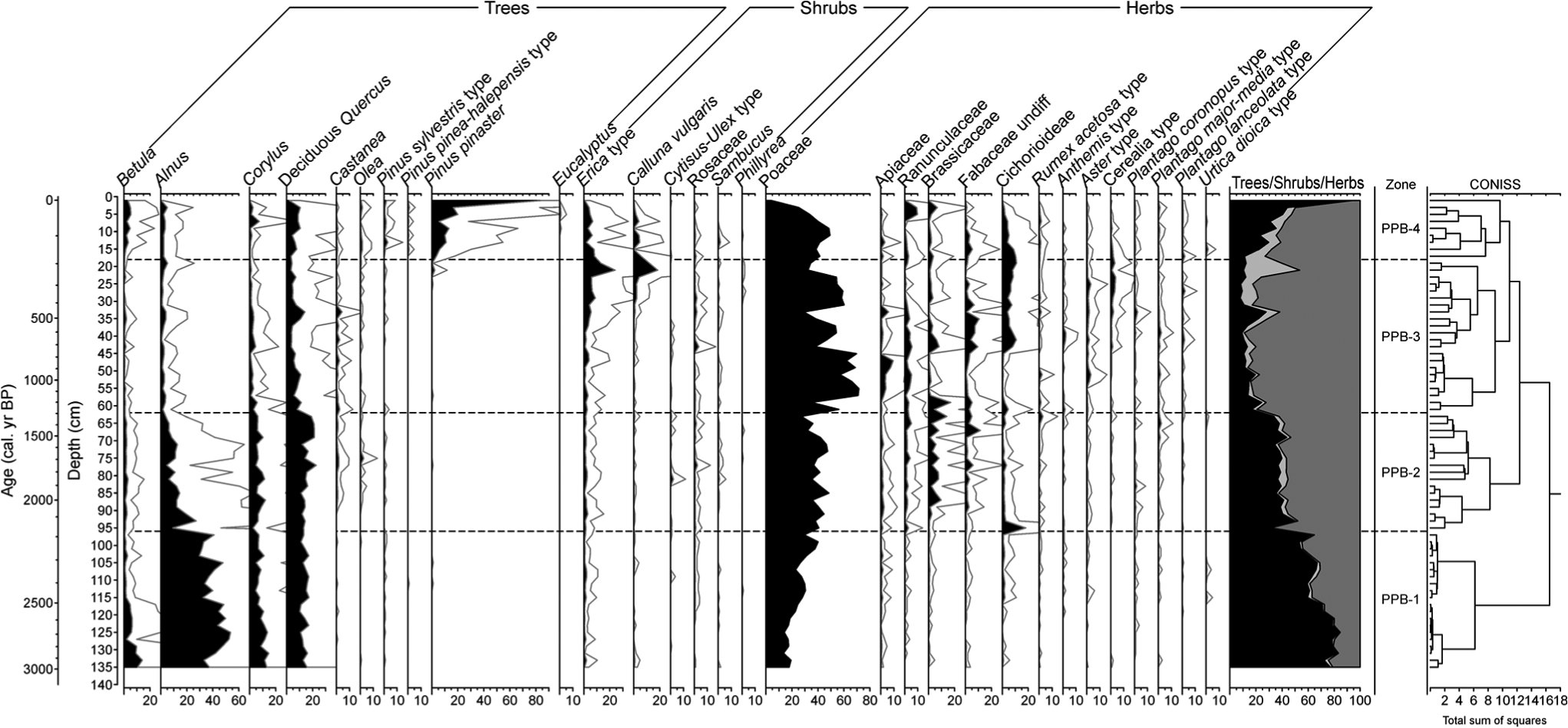
Synthetic palynological diagram of the Cruz do Bocelo mire showing the results for regional types (trees, shrub and herbs). Exaggeration curves have a factor of 5.
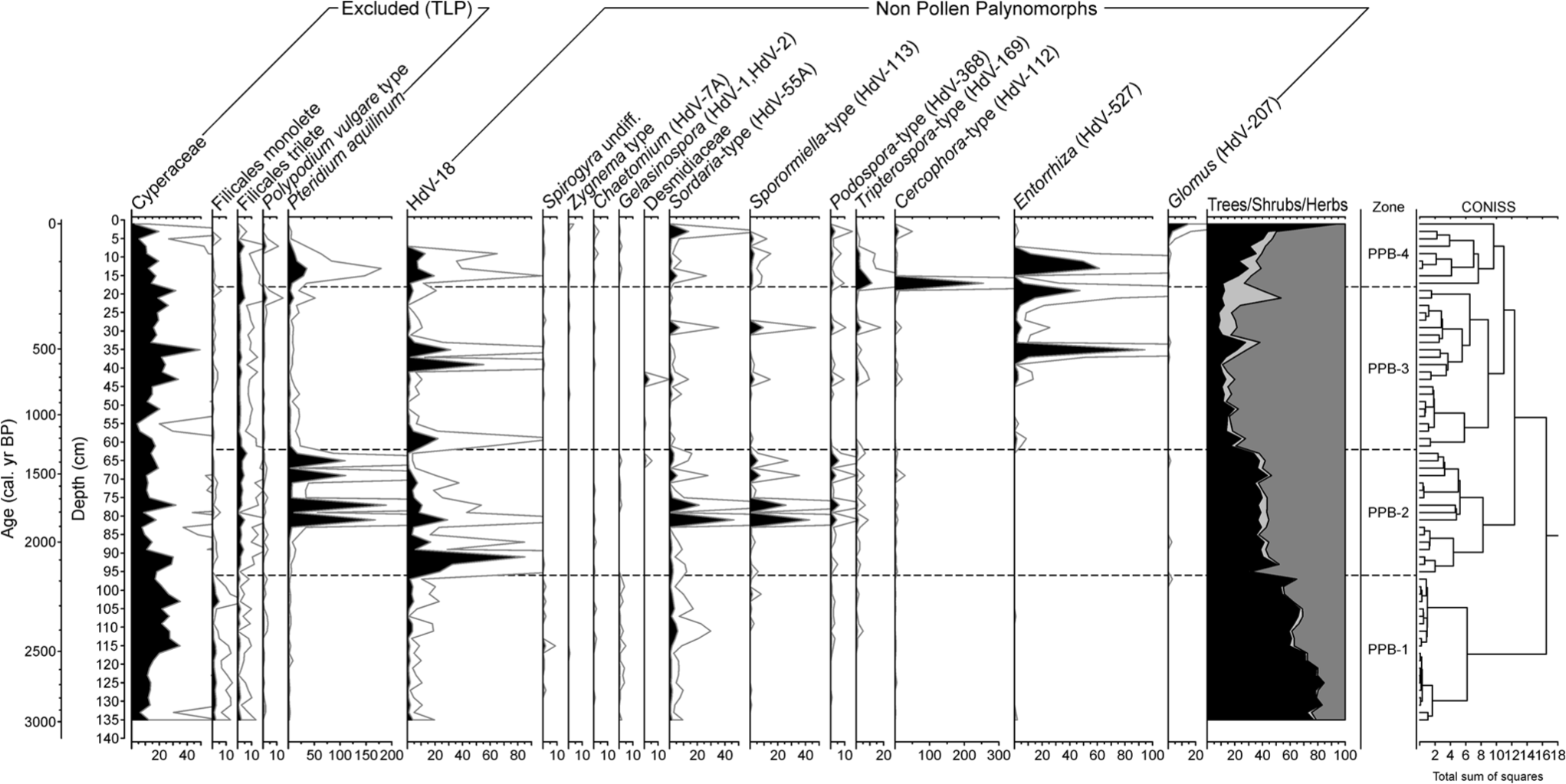
Synthetic palynological diagram of the Cruz do Bocelo mire showing the results for local types (excluded from the total land pollen sum and non-pollen palynomorphs). Exaggeration curves have a factor of 5.
Four palynological zones were identified according to CONISS. PPB-1 (136–96 cm; 3120–2175 cal. yr BP) is characterized by high percentages of AP (68.9 ± 9.5%), especially Alnus, Quercus, Corylus and Betula. Two phases of forest disturbance were recorded: at 126–110 and 106–98 cm depth. The first one mainly affected Alnus and Betula, while the second one just Alnus. Poaceae is well represented. Trace amounts of Cerealia-type is recorded at the bottom of the zone. Cyperaceae is the most abundant hydro-hygrophyte taxa, indicating a well-developed mire vegetation. The coprophilous fungi Sordaria-type occurs throughout the zone, increasing its percentages from 114 cm depth upwards. The mycorrhizal fungus Glomus is present at 94–96 cm depth. It has been often associated to erosion events (Anderson et al., 1984; Argant et al., 2006; Van Geel, 2001; Van Geel et al., 1989, 2003), since their spores may come from the soils of the catchment. However, peatlands are suitable environments for plants mychorrhized by Glomus (Kołaczek et al., 2013), so its presence may also reflect that type of association.
PPB-2 (96–62 cm; 2175–1295 cal. yr BP) is characterized by a major shift from a well-forested environment to a more open one. Mesophilous tree taxa, although still maintaining considerable values (38.1 ± 4.7%), decrease as Poaceae, and other herbaceous taxa such as Apiaceae, Ranunculaceae, Brassicaceae, Fabaceae and Cichorioideae all increase in representation. Percentages of cultivated trees such as Castanea and Olea increase in this zone. Cerealia-type increases its presence within this zone, but it never exceeds 1% TLP, while P. aquilinum and Sordaria-type, Sporormiella-type and Podospora-type show several peaks at 81, 77, 69 and 65 cm depth. HdV-18 also increases representation. Mighall et al. (2006) used HdV-18 as a proxy of past rainfall changes in an ombrotrophic peatland in the Xistral Mountains. However, Cruz do Bocelo is minerotrophic, so water supply not only depends on rainfall but also on catchment run-off. Glomus is present at 86–88 and at 64–66 cm depth.
In PPB-3 (62–18 cm; 1295–185 cal. yr BP), Poaceae reaches its maximum percentage for the PPB record. Grassland is the dominant type of vegetation as herbs represent a 77.5 ± 10%, while mesophilous pollen taxa are reduced to 13.6 ± 5.3%. Cerealia-type, Plantago spp. and nitrophilous taxa such as Cichorioideae, Anthemis-type and Aster-type increase in values. By the end of the zone, Erica-type and C. vulgaris increase in representation. Among NPP, coprophilous fungi such as Sordaria-type, Sporormiella-type, Podospora-type and Tripterospora-type, as well as Entorrhiza, show several peaks. Entorrhiza species are parasites on a variety of plants (Vánky, 1994), and their basidiospores have been related to clay sedimentation environments (Van Geel et al., 1983). Glomus is present at 36–38 and at 32–34 cm depth. HdV-18 shows an increase at the beginning of the zone, which occurs simultaneously with a decrease in mesophilous pollen taxa, and two peaks centred at 39 and 35 cm depth.
PPB-4 (<16 cm; <185 cal. yr BP) is characterized by a rapid increase in P. pinaster, a species that, like Eucalyptus, has been regularly planted since the second half of the 19th century in NW Iberia. Eucalyptus is entomophilous, and therefore only present in small percentages. P. aquilinum and Sordaria-type, Sporormiella-type, Podospora-type, Tripterospora-type and Cercophora-type abundance are indicative of continued use of grasslands for grazing. Moreover, Entorrhiza and Glomus show also increase during this zone.
Discussion
Chronology of environmental changes
Bronze and Iron Ages: moderate human pressure
Mesophilous forest was most widespread during this phase (PPB-1). On a local scale, any of the changes in rainfall (humidity indices, Figure 7) has had little observable impact on mire hydrology (HdV-18), probably because the forest acted as a buffer. Nevertheless, the presence of Cerealia-type (Figure 7) at the bottom of the sequence reveals that cultivation took place from at least the late Bronze Ages, although late Neolithic and early Bronze Age ceramics, lithic industries and megalithic burials (tumuli and dolmens) from the area attest to an even earlier phase of human occupation (Acuña Castroviejo and Mejide Cameselle, 1991; Criado Boado, 1991; Prieto Martínez, 1995). First evidence of deforestation in the PPB core occurs during the Iron Age (at ~2685–2400 cal. yr BP and ~2310–2220 cal. yr BP), when the increase in coprophilous fungi and/or nitrophilous taxa, indicative of animal husbandry, coincides with a slight decline in mesophilous forest. Contemporary increases in peat mineral matter content (PC1; E1: ~2640–2470 cal. yr BP and E2: ~2310–2250 cal. yr BP) suggest the inception of soil erosion by human activities during this early disturbance phase. While both forest disturbances have a similar magnitude, it is noteworthy that E2, which occurred during drier conditions (humidity indices; Figure 7), is less pronounced than E1, which happened during a wetter phase. The transition between the local Bronze and Iron Ages is considered to be a critical phase of changes in the exploitation of resources in NW Iberia (Martínez Cortizas et al., 2009a). During this period, evidence of soil erosion and acidification is found in many soil sequences in NW Spain (e.g. Benito et al., 1991; Costa Casais et al., 2009; Martínez Cortizas et al., 2000, 2009b). In some cases, these changes are accompanied by prominent charcoal layers indicating that fire was used to manage the forest. Soil erosion was already of concern for humans from at least the mid-Bronze Age. Although there are issues with regard to the dating of agrarian terraces, the first generation of man-made terraces date to ~3300 cal. yr BP in the Saa Valley (Martínez Cortizas et al., 2009a), and an analogous Iron Age structure was found in Castro de Follente (López-Sáez et al., 2009).
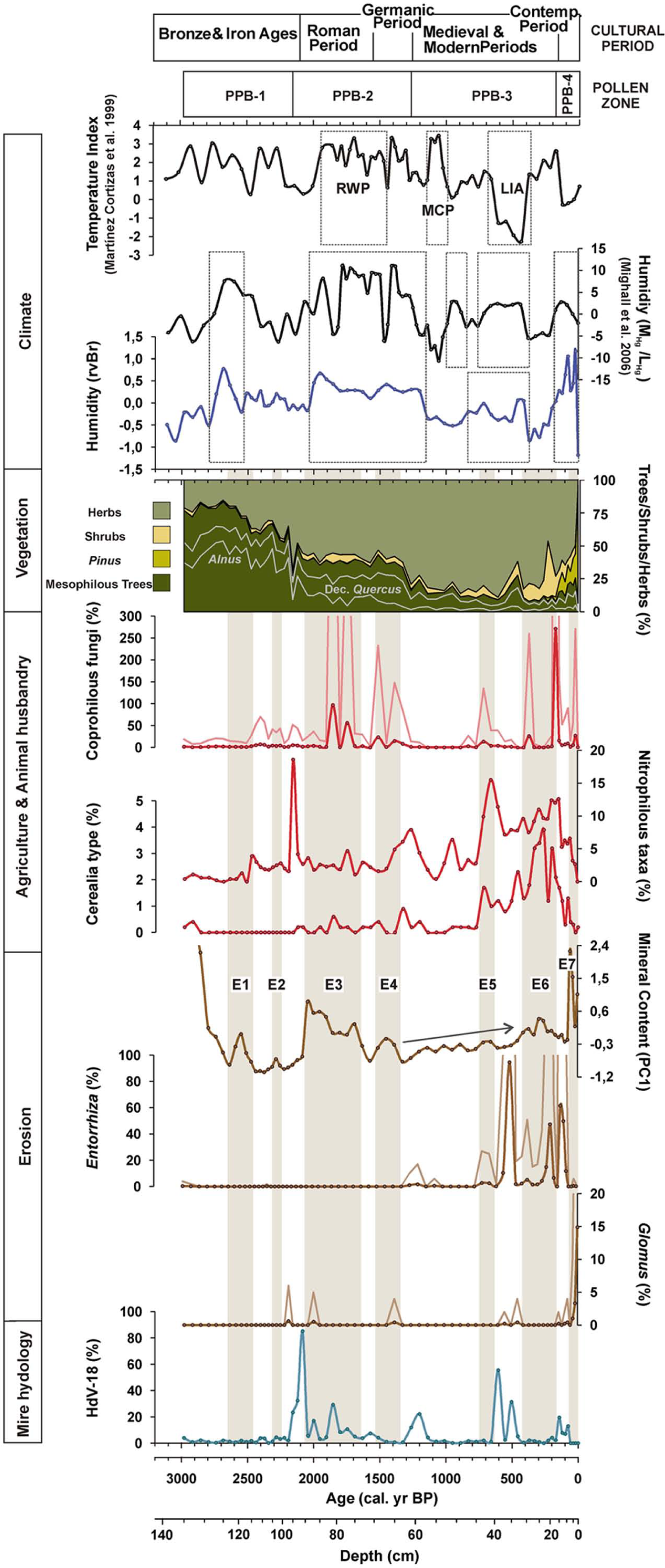
Chronology of the main environmental changes reflected by the Cruz do Bocelo records. Nitrophilous taxa = Anthemis-type + Asphodelus albus-type + Aster-type + Cardueae + Cichorioideae + Rumex acetosa-type + Urtica dioica-type; Coprophilous fungi = Cercophora-type + Podospora-type + Sordaria-type + Tripterospora-type + Sporormiella-type; PC1 (mineral content): we have avoided the representation of the most mineral samples near the substrate in order to amplify changes in the peat section of the core.
From Roman to late Middle Ages: mesophilous forest falls down
The most dramatic environmental change occurred at the transition from the Iron Age to the Roman Period (PPB-1/PPB-2 boundary). It involved changes at both a regional and local scale. Mesophilous forest suffers another large decline (mesophilous trees sum down to 43%) in a short period of time (~2190–2160 cal. yr BP) in favour of grassland. Alnus is the most affected tree. Simultaneous increases in coprophilous fungi and nitrophilous taxa link this decline to grazing. A dramatic increase in HdV-18 points to a higher mire water level. The presence of Glomus suggests soil erosion, and a slight increase in mineral matter (PC1) points to a disturbance phase of fairly modest intensity.
A short-lived period of forest recovery is detected (~2120 cal. yr BP). Thereafter, deforestation was more or less continuous from Roman times until the late Middle Ages (~660 cal. yr BP), characterized by two major phases of forest clearance: the first during Roman times (~2120–1750 cal. yr BP) and the second during the Germanic Period (~1510–1260 cal. yr BP). These phases are synchronous with the expansion of grassland and both grazing and cultivation. Despite the overall decrease in mesophilous forest, the percentages of Olea and Castanea increased. This could be related to warmer temperatures (Figure 7), especially in the case of Olea, although the increased presence of the abovementioned anthropogenic indicators does not rule out the possibility that Olea and Castanea were deliberately managed.
Deforestation during Germanic times represented a tipping point in the history of the mesophilous forest, as its previous importance would never be retained. Analogous responses found in other pollen records of NW Spain indicate that the irreversible decline in mesophilous forest was part of a wider process. Forest declines were found at ~1500 cal. yr BP in a soil sequence in Monte Paradela (Carrión et al., 2010b; Kaal et al., 2011; López-Merino et al., 2012); at ~1400 cal. yr BP in Pena da Cadela and Borralleiras da Cal Grande, two ombrotrophic mires located in the Xistral Mountains (Martínez Cortizas et al., 2005; Mighall et al., 2006); at ~1300 cal. yr BP in Monte Areo, a mire located in the Monte Areo Range (López-Merino et al., 2010); and at ~1200 cal. yr BP in Suárbol bog (Muñoz Sobrino et al., 1997) and at La Molina mire (López-Merino et al., 2011), located in the Ancares and in the Alto de la Espina Ranges, respectively (Figure 1). Thereafter, from ~1260 to ~660 cal. yr BP, pollen of taxa associated with mesophilous forest was at their lowest level (14.6 ± 4.1%). In contrast to other NW Iberian sequences (Allen et al., 1996; López-Merino et al., 2010; Martínez Cortizas et al., 2005; Mighall et al., 2006; Muñoz Sobrino et al., 1997), in the O Bocelo Range, no abrupt reductions in forest cover seem to have occurred. However, a further, slight, deforestation was recorded for the end of the Middle Ages (~710–600 cal. yr BP) at Cruz do Bocelo.
Large increases in the mineral content of the peat (PC1), as well as occasional rises in Glomus abundance, indicate severe soil erosion events in the catchment related to forest clearance during both the Roman Period (E3: ~2040–1690 cal. yr BP) and Germanic Period (E4: 1510–1390 cal. yr BP). During Medieval times, mineral inputs (PC1) to the mire gradually increased, culminating in E5 (~710–660 cal. yr BP).
Previous high-resolution studies of soil erosion and its link to forest evolution in NW Iberia (Martínez Cortizas et al., 2005; Xistral Mountains) found that forest clearances during Neolithic, Metal Ages, Roman, Germanic and Medieval times resulted in enhanced fluxes of mineral matter to mountain bogs. These chronologies are quite similar to those found at Cruz do Bocelo mire. However, regional differences are suggested when comparing the relative magnitude of each erosive phase. Erosion during Roman and Germanic times seems to have been more severe in the Bocelo range than in the Xistral Mountains. While medieval soil erosion seems to have been most relevant over the last ~5000 years in the Xistral Mountains, no similar record was observed at O Bocelo, suggesting that human transformation of the landscape varies by region and with altitude across NW Iberia.
Changes in regional vegetation and farming activities between ~2120 and ~600 cal. yr BP also affected the ecological evolution of the Cruz do Bocelo mire. During Roman and German phases of forest decline, high and abrupt increases in HdV-18 and P. aquilinum occurred. P. aquilinum has been associated with temporary woodland recession as it holds a pivotal role in succession (Marrs et al., 2000) and quickly invades perturbed sites (Ouden, 2000). However, its presence during this period and the good agreement with increases in coprophilous fungi may indicate successional changes on the mire surface because of cattle trampling. Moreover, increases in HdV-18 indicate that the tree cover loss would have decreased water retention in the catchment leading to higher superficial run-off and wetter conditions in the mire; this was probably intensified by the higher rainfall detected during this period (Figure 7).
‘Little Ice Age’ and contemporary human impact
A change in the trend between accelerated forest reduction and soil erosion occurred between ~605 and ~460 cal. yr BP. Evidence for enhanced soil erosion during this time includes increases in Glomus and Entorrhiza, the gradual increase in the mineral content of the peat (PC1) and the rise in silicon and LOI (Figure 3; 32–40 cm), which may reflect enhanced inputs to the mire. This appears to be the only soil erosion event which is not associated with a decrease in the mesophilous forest. It is chronologically framed within the cooler and more humid conditions of the ‘Little Ice Age’ (LIA), and it is probable that climate had a major influence on this episode of erosion. Moreover, increases in HdV-18 indicate wetter conditions in the mire. In NW Iberia, evidence of increased soil erosion and enhanced mineral particle inputs during the LIA have already been documented in Coto da Fenteira Atlantic ranker (Martínez Cortizas et al., 2000) and in Pena da Cadela bog (Martínez Cortizas et al., 2005; Xistral mountains), respectively (Figure 1). Elsewhere in Europe, evidence of higher mineral matter fluxes during the LIA has also been demonstrated by Meurisse et al. (2005) in peat-dune complexes from Northern France, by De Jong et al. (2007) in a raised bog from South Sweden and by De Vleeschouwer et al. (2009) in a bog from Northern Poland.
Although the idea of enhanced erosion during the LIA is commonly discussed, natural forest cover over Europe could have prevented dust deposition in peatlands (De Vleeschouwer et al., 2009). Thus, environments like Cruz do Bocelo where forest clearance occurred over several millennia are key areas for the detection of LIA-induced erosion and provide evidence that this climatic deterioration would have affected the ability of buffering anthropogenic changes. After the cold conditions of the LIA (~410–160 cal. yr BP), anthropogenic indicators (Cerealia-type, nitrophilous taxa and coprophilous fungi) show elevated values that, together with the increase of ericaceous shrub, show land degradation. However, as the humidity indices point to drier conditions (Figure 7), climate could have also influenced this shift. The low percentage of mesophilous trees and increases in peat mineral content (PC1) indicate the occurrence of another erosion phase (E6).
The recent afforestation with Pine and Eucalyptus commenced approximately two centuries ago, although it has intensified in the last decades, having a profound impact in mire catchment. Afforestation is responsible for the highest percentage of total AP in the whole record as the representation of the native mesophilous trees remains very low. Synchronous increases in the mineral content of the peat and Glomus indicate that despite increased arboreal cover, erosion was still very intense (E7: last ~80 years). This suggests that Pine and Eucalyptus afforestation is not as effective as the mesophilous forest in preventing soil erosion. Studies in this field indicate that harvesting practices can increase soil degradation and forest floor disturbance, which may result in soil loss (Beasley and Granillo, 1988; Blackburn et al., 1986; Castillo et al., 1997; Edeso et al., 1999; Rab, 1996). However, wetter climatic conditions (Figure 7) may have also enhanced soil loss and fluxes of mineral matter to the mire. Moreover, during the last decade, the construction of a road on one side of the mire possibly affected the fluxes of mineral matter.
Soil erosion and local moisture proxies responses
The combination of geochemical and palynological approaches allowed us to detect synergic effects and to interpret environmental changes at both regional and local scales at PPB mire. The geochemical principal component reflects the mineral content of the peat (PC1) and has proven to be a highly sensitive erosion index as it was linked to forest reduction during the last ~3000 years. But soil erosion also seems to have been affected by climate, particularly by rainfall. Glomus, however, was a less sensitive proxy for soil erosion, as it only increased at specific samples and often only during the episodes of higher erosion intensity, while Entorrhiza, which has been related to clay sedimentation (Van Geel et al., 1983), only increased during the LIA and in recent times.
The variable response of each of these erosion indicators is a very good example of how a multi-proxy approach strengthens the interpretation of environmental changes and helps in understanding the response of different proxies to the same environmental stressors. It is also important to know how a given proxy should be interpreted in different environments. In this regard, the fact that HdV-18 may respond to water-table fluctuations in minerotrophic mires (driven by climate or increases in water run-off because of deforestation in the catchment) may be of importance when comparing such patterns reconstructed from ombrotrophic bogs, in which it has been related to changes in rainfall (Mighall et al., 2006). This could be of interest for future reconstructions using peat cores from minerotrophic mires.
Conclusion
The integration of physical, geochemical and palynological data suggests that climate and human activities have been involved in intense landscape changes in the Cruz do Bocelo mire’s catchment during the last three millennia.
PCA on geochemical data identified a proxy for the mineral content of the peat (increases in the content of lithogenic elements), which was valuable to infer soil erosion events on a catchment scale, as well as a proxy for changes in humidity (Bromine residual variance), which agrees reasonably well with previous palaeoclimatic records, and was key to climatically contextualize environmental changes and to obtain more accurate interpretations.
The approaches presented in this paper show how human activities and climate have affected both soil erosion and the hydrology of the Cruz de Bocelo’s catchment. Although a forested landscape dominated ~3000 years ago, mesophilous forest decline and the development of an open landscape are a more or less continuous feature of the record since ~2685 cal. yr BP (local Iron Age). A link between forest clearance episodes and indicators of farming (Cerealia-type, coprophilous fungi, nitrophilous taxa) was clearly established. Deforestation would have triggered a decrease in water retention capacity of the catchment soils that resulted in increased run-off inputs into, and wetter hydrological conditions in, the mire. In this sense, the most prominent feature of the Cruz do Bocelo record is the dramatic change at the Iron Age/Roman period transition. Until then, the mire buffered climate and human impacts, while from this point onwards its hydrology responded abruptly to landscape changes in the catchment. Forest reduction enhanced the physical instability of the soils leading to soil erosion and increased fluxes of mineral matter to the mire. However, the interplay with climatic conditions accelerated these processes.
Differential responses of soil erosion (mineral content vs Glomus and Entorrhiza) and local moisture (HdV-18) proxies according to their sensitivity or to the type of archive (ombrotrophic vs minerotrophic mire) must be taken into account when reconstructing environmental change. In this sense, the finding that HdV-18 may be a useful proxy of water-table fluctuations in minerotrophic mires is of importance for future research.
Environmental management requires a thorough understanding of long-term environmental processes. In addition, peatlands are key natural ecosystems because of their biodiversity and environmental functions, and although some of them are being protected by different legislation, many are still suffering from degradation, especially minerotrophic mires. Cruz do Bocelo is a good example since the most significant amount of erosion over the last ~3000 years is the one taking place at present. The data presented here indicate that when implementing protection measures in mires, a whole catchment approach should be considered.
Footnotes
Acknowledgements
We are grateful to Xavier Pontevedra Pombal (USC) and Cruz Ferro Vázquez (Incipit-CSIC) for their assistance during the fieldwork; to José Antonio López-Sáez (CCHS-CSIC) for his support during different stages of the research, especially assistance with palynological identification; to Professor Suzanne Leroy (Brunel University) for her comments on early versions of the manuscript and to Tim Mighall for his comments and his corrections in the final English version of the manuscript
Funding
This work was funded by the projects CGL2010-20672 (Plan Nacional I+D+i, Spanish Ministry of Science and Innovation) and 10PXIB200182PR (General Directorate of I+D, Xunta de Galicia). N Silva-Sánchez and L López-Merino are currently supported by a FPU predoctoral scholarship (AP2010-3264) funded by the Spanish Government and a MINT postdoctoral fellowship funded by the Brunel Institute for the Environment, respectively.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.