
Abstract
Salient stimuli have the potential to distract us from our immediate goals. Much research has therefore aimed to understand how we learn to use attention to resist distraction by salient stimuli. We propose a new hypothesis whereby an initial instance of distraction can improve future suppression of salient stimuli. Across three experiments (N = 120 college students, aged 18–35 years), we provide evidence for this hypothesis using a new eye-tracking approach. The results demonstrated that an initial instance of distraction occurred before salient distractors were suppressed. Notably, if this initial instance of distraction was eliminated or weakened via experimental manipulations, learned suppression of the distracting stimuli was greatly reduced. Together, these findings suggest that attentional capture can serve as a learning signal that improves future attentional control. They also indicate that learned suppression emerges rapidly, which has strong implications for models of attention and cognitive control.
In everyday life, we frequently encounter salient stimuli, such as pop-up notifications or brightly colored billboards, that attempt to distract us from our goals. Much research has investigated whether salient stimuli have the power to involuntarily attract attention (attentional capture). Here, salience is defined by whether an object differs from neighboring objects in terms of low-level perceptual features (Nothdurft, 1993). For instance, a lone red object among several blue objects is considered highly salient (the color singleton in Fig. 1). Proponents of bottom-up theories, a classic approach, have proposed that salient stimuli have an automatic power to capture attention (Theeuwes, 1992), and this proposal has led to significant debate about top-down control over attention (Folk et al., 1992).
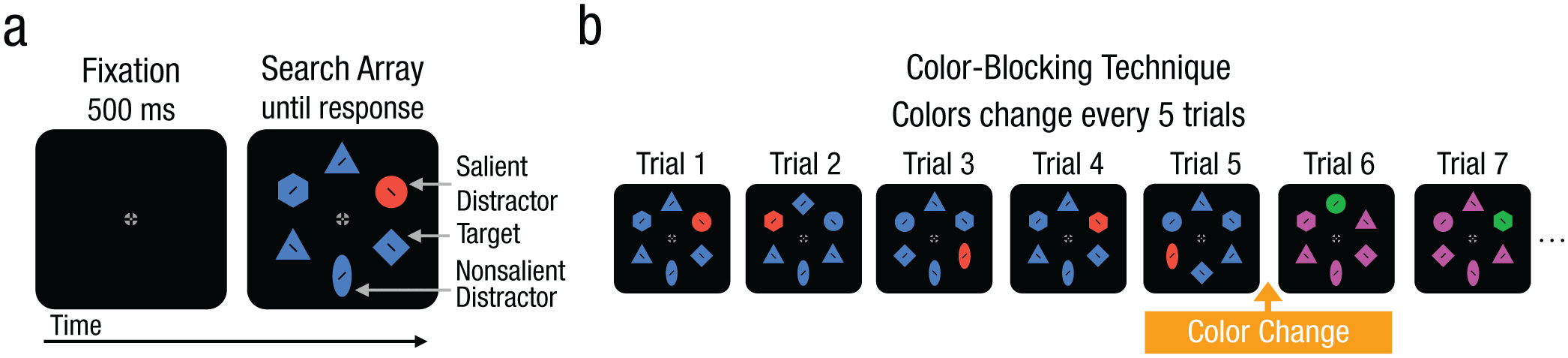
Stimuli and task for Experiment 1. Trial progression for the oculomotor suppression task is shown in (a): Participants searched for a target shape (i.e., a diamond) and attempted to ignore a salient distractor that was a color singleton. The color of the salient distractors (b) was blocked for short runs of five trials, which allowed evaluation of rapidly learned suppression with high statistical power.
Much evidence now suggests a hybrid account whereby attentional capture can be mitigated by suppressive processes. The signal-suppression account proposes that salient stimuli do have an automatic power to capture attention but that capture can be prevented via inhibition of salient stimuli (Sawaki & Luck, 2010). This account has been supported by converging evidence from psychophysics, eye tracking, event-related potentials (ERPs), and single-unit recordings (e.g., Chang & Egeth, 2019; Cosman et al., 2018; Drisdelle & Eimer, 2023; Gaspar & McDonald, 2014; Gaspelin & Luck, 2018; Gaspelin et al., 2015, 2017; Ma & Abrams, 2023; Sawaki & Luck, 2010; Stilwell & Anderson, 2025; Stilwell & Gaspelin, 2021; Won et al., 2019; H. Zhang et al., 2025). Much of this research has shown that salient stimuli can be suppressed below baseline levels to prevent attentional capture from occurring. For instance, studies of shifts of gaze have shown that eye movements are preferentially biased away from the salient distractors during visual search, which suggests that they are being suppressed to prevent distraction (e.g., Gaspelin et al., 2017; Y. Zhang & Gaspelin, 2024).
There is also growing evidence that the ability to suppress salient stimuli is learned. For example, attentional capture is greatly reduced when salient stimuli repeatedly appear in the same color or location (e.g., Gaspelin & Luck, 2018; Savelson et al., 2025; Vatterott & Vecera, 2012; Wang & Theeuwes, 2018). These results imply that prior experience leads to learned biases that powerfully influence attentional allocation (Anderson et al., 2011, 2021; Awh et al., 2012). As a result, recent formulations of the signal-suppression account suggest that attentional suppression is largely the result of implicit learning (Gaspelin et al., 2025).
Here, we propose a new hypothesis about how attention learns to suppress salient stimuli, called the capture-dependent suppression hypothesis. We propose that an initial instance of attention capture will improve later suppression of salient stimuli. This hypothesis was explained in detail by Gaspelin et al. (2025). The crux is that, after each attentional shift, the currently attended object is compared with an internal representation of the target (Yu et al., 2023). If the attended object does not match the target, the features and locations associated with the object are suppressed in attentional gain controls to reduce subsequent distraction. This leads to a straightforward prediction: Any instance of capture will help strengthen future suppression of the features and locations associated with a salient stimulus. Notably, this learning could occur relatively quickly (e.g., within a few exposures to the salient distractor) because the comparison process occurs after each attentional shift.
The present study will examine the capture-dependent suppression hypothesis. Three experiments will test the prediction that initial capture facilitates a learning process that improves suppression. As will be seen, the findings support a new model of attentional control in which an initial instance of capture improves suppression in future encounters with distracting stimuli.
Research Transparency Statement
General disclosures
Study 1 through 3 disclosures
Experiment 1
Participants searched for a target shape and attempted to ignore a salient distractor while shifts of gaze were recorded (Fig. 1). A new color-blocking technique was introduced in which the display colors changed every five trials. This approach allowed us to measure the oculomotor processes associated with rapidly learned suppression with great statistical power. We will specifically analyze whether shifts of gaze to the salient distractor are captured or suppressed as a function of the sequence in the color run.
There are competing predictions about potential outcomes. According to the capture-dependent suppression hypothesis, there should be an initial instance of capture on the first trial of a color run when the color has just changed (Trial 1), followed by learned suppression of the distractor color on later trials in the run (Trials 2–5). In contrast, a purely bottom-up account predicts that top-down control cannot eliminate capture. As a result, the salient distractor should capture attention, regardless of position within a color run.
Method
Participants
Forty participants from the University of Missouri were recruited for payment of $20 per hour (20 men and 20 women, Mage = 23.1 years). The sample size was determined a priori based upon a previous study (Y. Zhang & Gaspelin, 2024). Assuming an oculomotor suppression effect is the same magnitude as this study (dz = 0.94), a minimum of 17 participants would be needed to obtain 95% power. Because this is a new task and the effect size of the learning effect was unknown, we erred on the side of caution and recruited a larger sample than needed. As will be seen, we also replicated the key findings in Experiments 2 and 3. All participants had normal color vision, as indicated by an Ishihara test, and normal or corrected-to-normal visual acuity, as evaluated by a Snellen chart. The research protocol was approved for ethical considerations by the internal review board at the University of Missouri (ID No. 2097167).
Apparatus
Stimuli were presented using PsychToolbox for Linux. An Asus VG248QG monitor presented stimuli on a black background at a viewing distance of 100 cm in a dimly lit room. A photosensor was used to measure the timing delay of the video system (7 ms), and this value was subtracted from all latencies. An EyeLink 1000 Plus desk-mounted eye tracker measured gaze position monocularly from the right eye at 500 Hz.
Stimuli and procedure
Participants performed an adapted version of an oculomotor suppression task (Gaspelin et al., 2017). As shown in Figure 1a, search displays are composed of six shapes arranged in a notional circle, with an eccentricity of 4.3° from the screen center. Each search display contained one diamond (1.7° × 1.7°) and one circle (1.7° in diameter). The remaining items could be hexagons (1.7° × 1.7°), triangles (1.7° height and 1.7° width), or ovals (2.0° × 1.4°), and these shapes were randomly selected with the exception that no more than two instances of a given shape could be present in a search display. Each shape featured a small black line (0.2° × 0.1°) that was tilted at a 45° angle to the left or right. These lines were small, making them difficult to see parafoveally and therefore necessitating eye movements to perform the task. Each search display included a fixation cross positioned at the screen center, which consisted of a gray circle (30.2 cd/m², x = .290, y = .294) with a diameter of 0.5°, containing a black crosshair with a central gray dot (0.1° in diameter). Prior research has shown that this type of fixation cross improves fixation stability compared with other types of fixation crosses (Thaler et al., 2013).
Each participant searched the displays for a specific target shape (e.g., a diamond). The target shape remained constant for the entire experiment and was counterbalanced across participants: a diamond for half of the participants and a circle for the other half. Heterogeneous distractor shapes were used to make the target nonsalient and to prevent participants from searching broadly for any popout stimulus (Bacon & Egeth, 1994); this has been shown to be important for oculomotor suppression to develop (Gaspelin et al., 2017). On every trial, a salient distractor appeared as a uniquely colored object that was a different color from the other search objects. This salient distractor could never be the target. Participants were informed of this and were instructed to ignore the salient distractor to the best of their ability.
As shown in Figure 1b, the display colors changed every five trials (see related studies by Reinhart & Woodman, 2015; Vatterott & Vecera, 2012). To accomplish this, we generated a color wheel of 360 isoluminant color pairs using the International Commission on Illumination (CIE) color space (L = 40, R = 90). For each color run, the target color was randomly selected from this color wheel, and the salient-distractor color was selected to be 180° from the target color on the color wheel (i.e., the opposing color). This was meant to produce a highly salient color singleton. The colors of the nonsalient distractors always matched the target color. When the display colors changed, a new color pair was selected at random, with the exception that the new color pair must be at least 60° from the previous color pair. This ensured the new color set was sufficiently different from the previous color set.
Each trial began with the fixation cross. To initiate a trial, participants had to maintain central fixation for 500 ms within a circular region of 1.5° around the fixation cross. Once this requirement was met, the search display appeared. Participants searched for the target shape (e.g., a diamond) and reported the tilt of the line inside the target (left or right). Manual responses were made via a speeded button press on a game pad with the left- and right-shoulder buttons, respectively. The search array appeared until a response was made or until a 2,000-ms time-out. If the participants responded inaccurately or a time-out occurred, a 200-Hz tone and a prompt message saying either “Wrong!” or “Too Slow!” (respectively) were displayed for 300 ms before participants moved on to the subsequent trial.
Performance feedback on mean accuracy and response time (RT) was provided at the end of each block. These block breaks also warned participants if their accuracy fell below 90% in a given block. Before the start of a new block, the eye-tracking system was recalibrated using a 9-point calibration. During the main task, the eye-tracking system was also recalibrated if a participant failed to fixate the fixation cross for more than 8 s.
The experiment consisted of 60 practice trials and 480 experimental trials, organized into nine blocks. Given that each color run consisted of five trials, this meant that there were 96 trials for each trial bin in the color run (i.e., 480 total trials divided by 5 trials in each color run). This allowed us to have robust statistical power to study learned oculomotor suppression.
Data analysis
The onset of an eye movement was determined by a minimum acceleration threshold of 9500°/sec2 and a minimal eye velocity threshold of 30° per second. Our analyses focused on the speed and destination of the first eye movement during visual search. To categorize the destination of the first eye movement, we used an approach from previous studies using a similar paradigm (e.g., Gaspelin et al., 2017; Gaspelin & Luck, 2018; Stilwell et al., 2023; Y. Zhang & Gaspelin, 2024). An annulus was defined and centered on the fixation cross with an inner radius of 1.5° from fixation and an outer radius of 7.5° from fixation. The first eye movement landing within the annulus was identified as the first eye movement, and the nearest search item was then selected as the destination of the eye movement. This approach effectively creates wedge-shaped interest areas around each display object. Saccadic latency was measured as the start time of the first eye movement that landed within the annulus.
An exclusion criterion was established a priori. Participants were replaced if their manual RT, saccade latency, and manual error rates were more than 3.5 SDs from the mean. We also replaced participants who made eye movements on less than 75% of trials. No participants met the exclusion criteria. For these 40 participants, the practice block was excluded from analysis. Trials with RTs below 200 ms or above 2,000 ms (1.6%) were removed, along with those in which participants did not move their eyes from central fixation (0.1%) or had abnormal saccade latencies (less than 50 ms or greater than 1,000 ms; 5.6%). Additionally, trials with incorrect manual responses (2.0%) were omitted from all analyses but the manual error-rate analysis. In total, 8.3% of trials were excluded.
Because the display colors changed every five trials, we structured the analyses by aggregating data within these five-trial color runs, forming standardized Trials 1 through 5 across the experiment for subsequent analysis. Thus, Trial 1 is the initial trial in which the color changed. As previously mentioned, this meant there were 96 trials for each trial bin, which granted us strong statistical power to observe learned suppression of salient distractors.
All analyses of variance (ANOVAs) reported in this article include Greenhouse–Geisser corrected p values, and Cohen’s d for within-subject t tests is reported as dz. For one-sample t tests in the color-run analyses, the false discovery rate (FDR) correction was applied to control for multiple comparisons (Benjamini & Hochberg, 1995).
Bayesian analyses were conducted using JASP (Version 0.95.1; JASP Team, 2025). For Bayesian t tests, Bayes factors (BF10) quantify the relative evidence for the alternative versus the null model based upon the ratio of their marginal likelihoods. When BF10 was less than 1, we report the reciprocal BF01 = 1 / BF10. All Bayesian t tests used the default Jeffreys-Zellner-Siow (JZS) prior on the standardized effect size implemented in JASP with a zero-centered Cauchy prior with a scale parameter (r) of .707 (Rouder et al., 2009). For Bayesian ANOVAs, we report inclusion Bayes factors (BFincl) from JASP, which quantify the change from prior-inclusion odds to posterior-inclusion odds for each effect under Bayesian model averaging. When BFincl < 1, we report the reciprocal exclusion Bayes factor BFexcl = 1/BFincl. Bayesian ANOVAs used JASP’s default coefficient priors (r scale fixed effects = .5; r scale random effects = 1) with a uniform model prior (see Rouder et al., 2012); marginality was enforced for fixed effects, and no covariates were included.
Results
Manual responses
Table 1 depicts manual RTs and error rates as a function of the number of trials with the same color. As can be seen, manual RTs were slowed on Trial 1 compared with Trials 2 through 5, which is not unexpected (attentional capture occurs on Trial 1 when the color changes, slowing the detection of the target). A one-way repeated-measures ANOVA was conducted on manual RT with the factor of trial bin. There was a main effect of trial bin, F(4, 156) = 33.84, p < .001, η p 2 = .46, BFincl > 1,000. In addition, a paired-samples t test confirmed that responses were slower on Trial 1 than Trial 5, t(39) = 10.37, p < .001, dz = 1.60, BF10 > 1,000.
Manual Response Times (ms) and Error Rates across Trial Bins in Experiments 1 and 2
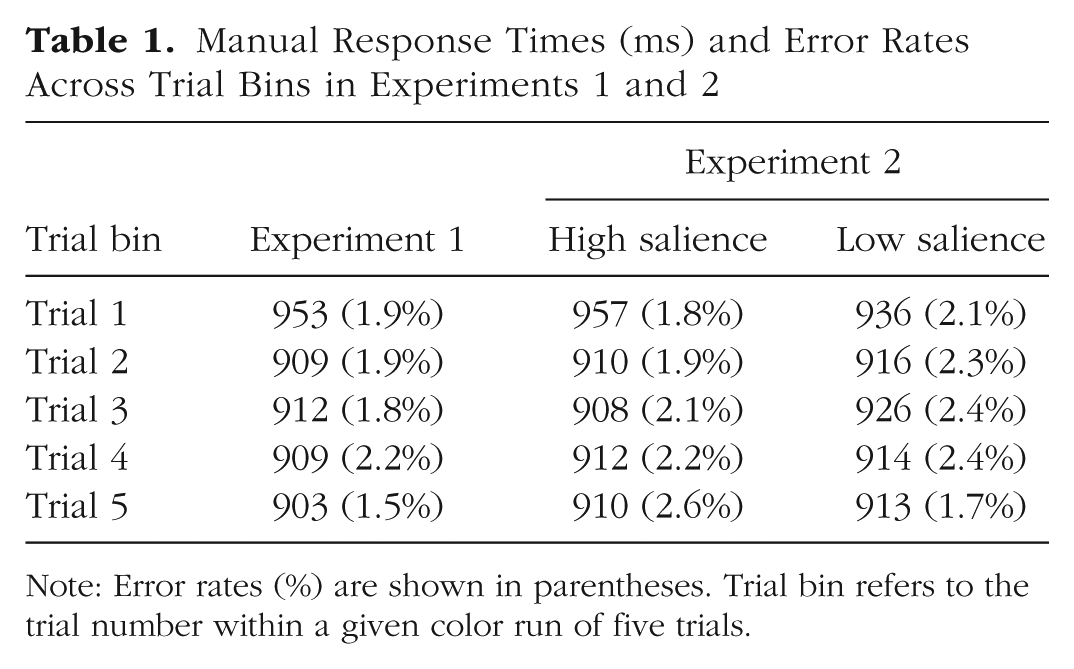
Note: Error rates (%) are shown in parentheses. Trial bin refers to the trial number within a given color run of five trials.
The same one-way repeated-measures ANOVA was conducted on manual error rates with the factor of trial bin. The analysis did not reveal a significant main effect of trial bin, F(4, 156) = 0.95, p = .440, η p 2 = .02. A complementary Bayesian analysis provided strong evidence for the null over the alternative (BFexcl = 15.43).
Overall, the results are consistent with an account in which capture occurred on the initial color-switch trial (Trial 1), which slowed detection of the target. However, such a slowing effect could also occur because the target color changed, slowing visual search without any attentional capture by the salient distractor. The next section will therefore use the destination of first eye movements to directly assess overt attentional allocation to the salient distractor.
First eye movements
According to the capture-dependent suppression hypothesis, first eye movements should be biased toward the salient distractor on Trial 1 when the color changes (initial capture). On Trials 2 through 5, suppression should develop as participants gain experience with the color configuration. As shown in Figure 2, the observed results are directly consistent with this prediction. Figure 2a depicts the percentage of first eye movement to each search item (target, nonsalient distractor, and salient distractor) as a function of trial bin. The percentage of eye movements to the nonsalient distractors has been divided by four to give a per-item estimate. As can be seen, there is evidence of oculomotor capture on Trial 1, meaning that the salient distractor is more likely to be fixated than the baseline level of the average nonsalient distractor. On Trials 2 through 5, however, there is evidence of oculomotor suppression (i.e., the salient distractor is fixated below the baseline level of the nonsalient distractors).
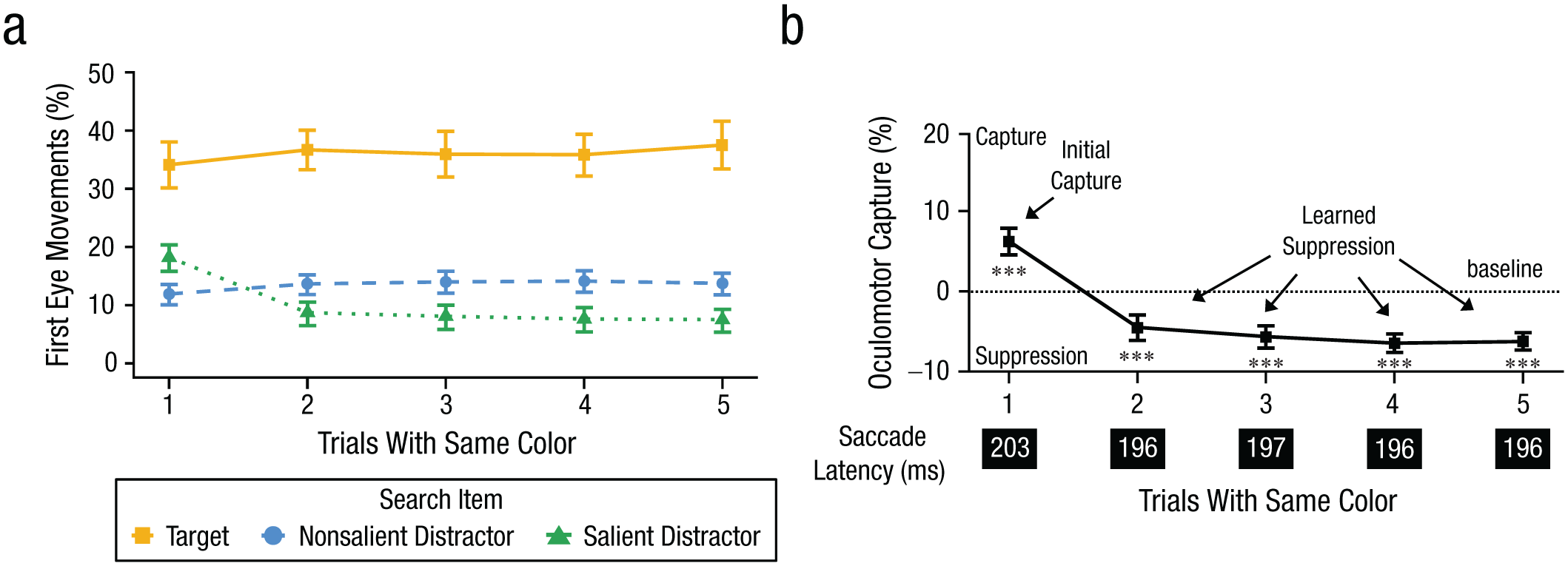
Results for Experiment 1. Shown in (a) are the first eye movements to each search item as a function of trial bin with the same color. Shown in (b) are oculomotor capture effects by salient distractors as a function of trial bin with the same color. Positive values indicate initial capture (Trial 1), whereas negative values reflect learned suppression (Trials 2–5). The downward trend illustrates the rapid shift from capture to suppression. Mean saccadic latency for each trial bin is provided below the x-axis. Error bars represent within-subject 95% confidence intervals (Morey, 2008). ***p < .001.
The pattern of initial capture and learned suppression can be more easily seen in Figure 2b. Here, we calculated an oculomotor capture effect, which is a difference score between the percentage of first eye movements to the salient distractor minus the average nonsalient distractor. This is a common metric used to evaluate whether initial eye movements were captured by the salient distractor (Gaspelin et al., 2017). A positive value indicates that the salient distractor captured attention above baseline levels and a negative value indicates that the salient distractor was suppressed below baseline levels. As shown, there was an initial capture effect on Trial 1, and this turned into an oculomotor suppression effect on Trials 2 through 5.
A one-way repeated measures ANOVA was conducted on oculomotor capture effects with the factor of trial bin. There was a main effect of trial bin, F(4, 156) = 60.72, p < .001, η p 2 = .61, BFincl > 1,000. A series of planned one-sample t tests was conducted to examine whether the oculomotor capture effect significantly deviated from zero in each trial bin. On Trial 1, the initial capture effect was significantly greater than zero (+5.9%), t(39) = 6.50, p < .001, dz = 1.03, BF10 > 1,000. This initial capture was rapidly followed by learned suppression, with significant oculomotor suppression observed as early as Trial 2 (–4.8%), t(39) = −5.98, p < .001, dz = −0.95, BF10 > 1,000. This pattern of suppression continued across Trial 3 (−5.6%), t(39) = −6.86, p < .001, dz = −1.09, BF10 > 1,000; Trial 4 (−6.8%), t(39) = −9.27, p < .001, dz = −1.47, BF10 > 1,000; and Trial 5 (−6.1%), t(39) = −10.24, p < .001, dz = −1.62, BF10 > 1,000.
Taken together, the results indicate a highly efficient learning mechanism that rapidly reduces the interference from a salient distractor after only a single instance of capture (see also Savelson et al., 2025). The result is consistent with the capture-dependent suppression hypothesis: Initial capture preceded learned suppression.
Learned suppression as a function of initial capture
We next conducted an analysis aimed at linking capture on Trial 1 with suppression on Trials 2 through 5. According to the capture-dependent suppression hypothesis, initial capture should increase future suppression and reduce the impact of the salient distractor on subsequent trials. If this is true, the rate of learned suppression should be improved following an initial instance of capture. In other words, the number of eye movements to salient distractors should be greatly reduced if Trial 1 resulted in capture.
For each color run, we classified whether the first eye movement was directed to the salient distractor on Trial 1 (i.e., capture vs. no capture). We then computed a learning slope on Trials 2 through 5. Learning slopes were then computed by plotting the first eye movements to the salient distractor on Trials 2 through 5 for each participant and fitting a linear slope. Here, a negative slope indicates a reduction in capture by the salient distractor, whereas a slope of zero indicates no reduction in capture. Participants were excluded from the analysis if capture occurred on less than 10 trials on Trial 1, because there would be inadequate power to compute a learning curve on Trials 2 through 5. Three participants were excluded for this reason.
The reduction in capture was significantly greater following capture (−1.4% per trial) than following no capture (−0.1% per trial), t(37) = 2.23, p = .032, dz = 0.37, BF10 = 1.58. In other words, after capture on Trial 1, the rate of attending to the singleton was reduced by an average of 1.4% for each subsequent trial on Trials 2 through 5. When capture did not occur on Trial 1, orienting to the salient distractor was only reduced by an average of 0.1% on each subsequent trial. This pattern is consistent with the capture-dependent suppression hypothesis in that it shows that the salient distractor was more strongly downweighted following an initial instance of capture.
Learning effects across the experiment
We were interested in how stable the learning effects were across the experiment. For instance, it is possible that changing the color of the salient distractor would become less effective as participants habituate to the repeated color changes. We therefore compared initial capture effects on Trial 1 in the first half of the experiment (+6.0%) and the second half of the experiment (+5.8%). The analysis did not reveal a significant difference, t(39) = −0.21, p = .831, dz = −0.03. Complementary Bayesian analysis provided moderate evidence for the null over the alternative (BF01 = 5.74). In addition, we calculated learned suppression effects by using oculomotor capture effects on Trial 1 minus Trial 5. The learned effects did not differ significantly in the first half of the experiment (12.0%) and second half of the experiment (12.1%), t(39) = −0.06, p = .949, dz = −0.01, with Bayesian analysis indicating moderate evidence for the null hypothesis (BF01 = 5.85), showing that the data are 5.8 times more likely under the null hypothesis than under the alternative. Altogether, these results show that the learned suppression effect was fairly consistent across the experiment.
Saccadic latency
According to a saccadic slowing account, participants might learn to suppress salient distractors by slowing eye movements to prevent attentional capture from occurring. Indeed, such saccadic slowing effects have been observed in previous studies to improve top-down control (e.g., Anderson & Mrkonja, 2021; Donk & van Zoest, 2008; van Zoest et al., 2004). Figure 2b therefore depicts saccadic latency for each trial bin. The saccadic latency was calculated as the time to initiate the first eye movement after the onset of the search stimulus. As can be seen, we observe the opposite pattern predicted by a saccadic slowing account of the results. If anything, saccadic latencies were significantly faster on Trial 5 (196 ms) than on Trial 1 (203 ms), t(39) = −5.53, p < .001, dz = −0.87, BF10 > 1,000. Thus, the learned suppression effect cannot be explained by mere slowing of eye movements to prevent capture.
Dwell time
Learned suppression might also improve the ability to disengage from salient distractors after they capture attention. We therefore computed dwell times on salient distractors as a function of the number of trials with the same color (Table 2). This analysis was restricted to trials in which the first eye movement was directed to the salient distractor, and we then computed the duration of the initial fixation on the salient distractor. As can be seen, dwell time generally decreased from Trial 1 to Trials 2 through 5. A one-way repeated-measures ANOVA was conducted on dwell time to the salient distractor with the factor of trial bin. There was a main effect of trial bin, F(4, 156) = 4.87, p < .001, η p 2 = .11, BFincl = 24.64. In addition, a paired-samples t test confirmed that dwell time was longer on Trial 1 than Trial 5 (140 vs. 118 ms), t(39) = 4.46, p < .001, dz = 0.64, BF10 = 342.47. This pattern indicates that participants also learned to disengage from salient distractors more quickly when capture did occur as they gained experience within a trial run.
Dwell Times (in ms) on Salient Distractors across Trial Bins in Experiments 1 and 2
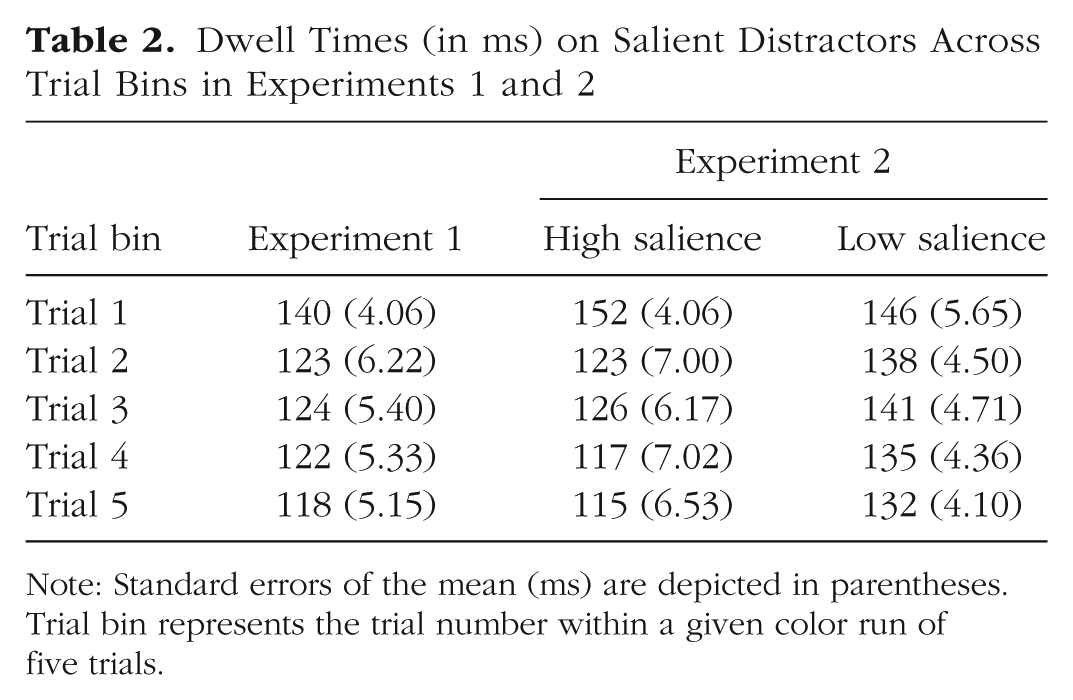
Note: Standard errors of the mean (ms) are depicted in parentheses. Trial bin represents the trial number within a given color run of five trials.
Discussion
The results demonstrated a clear pattern of initial capture (on Trial 1 when the color changed) followed by a learned suppression (Trials 2–5). We also observed evidence that initial capture improved later learned suppression. These findings uniquely suggest that learned suppression can develop rapidly and also align with the prediction of the capture-dependent suppression hypothesis that an initial event of capture will increase suppression of features associated with the salient distractor.
Experiment 2
Experiment 2 was designed to further examine the linkage between initial capture and learned suppression. Prior studies have shown that high-salience distractors are more strongly suppressed than low-salience distractors (e.g., Stilwell & Anderson, 2025; Stilwell et al., 2023; Y. Zhang & Gaspelin, 2024). This result might seem counterintuitive, but the capture-dependent suppression hypothesis offers a straightforward explanation. High-salience distractors initially capture attention, which then improves learned suppression. Low-salience distractors, however, will not initially capture attention and therefore will not trigger a suppressive process. Experiment 2 directly tests this prediction by comparing shifts of gaze to high- and low-salience distractors with our new color-blocking approach (Fig. 3).
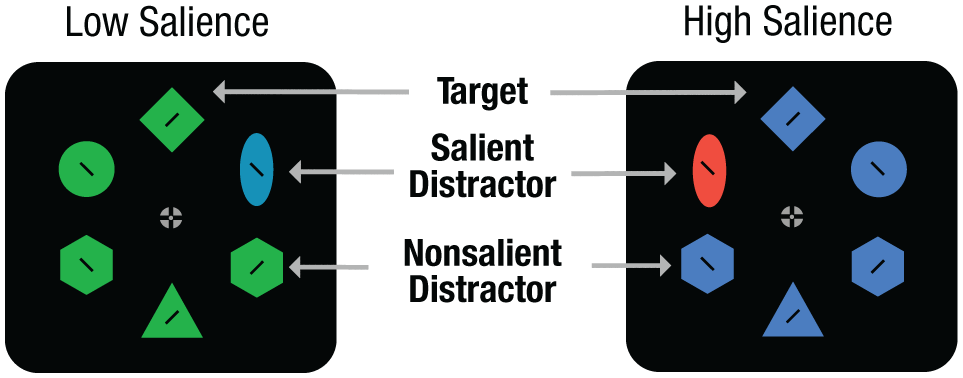
Stimuli and task for Experiment 2. Display colors were changed every five trials. Low-salience and high-salience distractors were compared.
The capture-dependent suppression hypothesis predicts that low-salience distractors will not initially capture attention and that this lack of capture will weaken subsequent suppression of the salient stimuli. A purely bottom-up account, however, predicts that capture should persistently occur in the high-salience condition and should be larger than that of the low-salience condition, regardless of prior experience with color.
Method
All methods were identical to Experiment 1’s, except for the following changes.
A new sample of 40 participants was recruited from the University of Missouri for payment of $20 per hour (16 men and 24 women, Mage = 18.5 years). No participants were removed as outliers.
The same color-blocking technique was used as in Experiment 1 (i.e., the display colors changed every five trials). The key modification was that there were low-salience and high-salience distractor conditions (Fig. 3). The high-salience condition was identical to Experiment 1’s, in which the salient distractor color was 180° from the target color on the color wheel. In the low-salience condition, the salient distractor color that was 30° from the target color on the color wheel was selected, resulting in a low-contrast singleton. High- and low-salience conditions were randomly interspersed throughout the experiment. In other words, every five trials new display colors were randomly selected. This salient distractor could have either high or low salience.
The eye-tracking recording and analysis were the same as in Experiment 1. The first block was excluded as practice. Trials with RTs less than 200 ms or greater than 2,000 ms were removed from analysis (2.2% of trials). Trials in which participants did not move their eyes from central fixation (0.3%) and trials with abnormal saccade latencies (< 50 ms or > 1,000 ms, comprising 5.4% of trials) were also removed. Trials with incorrect manual responses (2.2%) were omitted from all analyses except for the manual error-rate analysis. In all, 8.9% of trials were excluded.
Results
Manual responses
We had no direct predictions about manual responses but analyzed them for the sake of completeness. Manual RTs were slower on Trial 1 compared with later trials (Trials 2–5) in both high- and low-salience conditions (see Table 1). As suggested in Experiment 1, the manual RTs could be slowed on Trial 1 because of capture, but some slowing might also occur because of changes in display colors, which disrupt attentional guidance. A two-way repeated-measures ANOVA was conducted on manual RT with two within-subject factors—trial bin (Trials 1–5) and distractor salience (low-salience and high-salience). There was a significant main effect of trial bin, F(4, 156) = 16.57, p < .001, η p 2 = .30, BFincl > 1,000, indicating that manual RTs varied systematically across trials, with faster responses in later trials. The main effect of distractor salience was not significant, F(1, 39) = 0.10, p = .750, η p 2 = .00, with Bayesian analysis indicating inconclusive evidence (BFexcl = 0.88). Crucially, there was a significant interaction between trial bin and distractor salience, F(4, 156) = 3.50, p = .009, η p 2 = .08, BFincl = 5.29. This indicates that the reduction in RT was greater for the high-salience condition (47-ms reduction) than the low-salience condition (24-ms reduction).
In addition, the same two-way repeated-measures ANOVA was conducted on manual error rates with the factors of trial bin and distractor salience. The analysis did not reveal a significant main effect of trial bin, F(4, 156) = 0.34, p = .854, η p 2 = .01, with Bayesian analysis providing strong evidence for the null hypothesis (BFexcl = 120.44). Likewise, there was no significant main effect of distractor salience, F(1, 39) = 0.15, p = .703, η p 2 = .00, and a complementary Bayesian analysis revealed strong evidence for the null hypothesis (BFexcl = 10.54). Moreover, there was no significant interaction between trial bin and distractor salience, F(4, 156) = 1.14, p = .341, η p 2 = .03, BFexcl > 1,000. Overall, these results indicate that participants consistently maintained a high level of accuracy throughout the experiment.
First eye movements
The key prediction in this experiment was that low-salience distractors should not capture attention on Trial 1 and therefore would not be suppressed on Trials 2 through 5. Figure 4a depicts the percentage of first eye movement to each item type (target, nonsalient distractor, and salient distractor) as a function of trial bin with the same color for both low- and high-salience conditions. Figure 4b shows oculomotor capture effects for both distractor types. As can be seen, the high-salience distractor exhibited initial oculomotor capture on Trial 1, followed by suppression on Trials 2 through 5. Low-salience distractors, however, showed no initial capture and weakened suppression in later trials. This pattern is broadly consistent with the capture-dependent suppression account, which suggests that initial capture is necessary for learned suppression to develop.
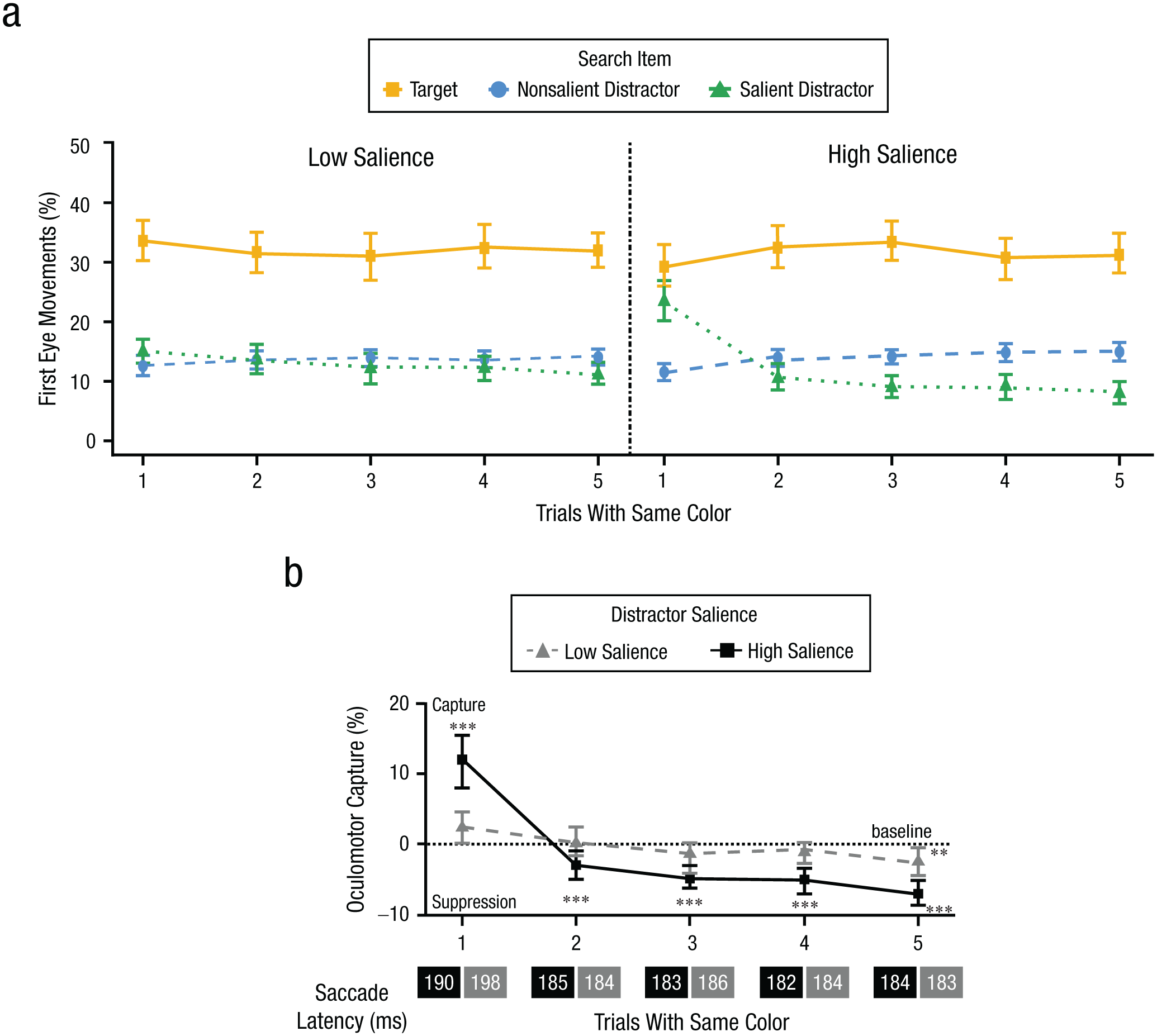
Results for Experiment 2. First eye movements to each search item are shown in (a) as a function of trial bin, with the same color for both low- and high-salience conditions. In (b) we show oculomotor capture effects for Trials 1 through 5 for each salience condition. The high-salience distractor exhibits initial capture followed by suppression. The low-salience distractor, however, shows no initial capture, and the resulting learned suppression is weakened. The saccadic latency for each trial number is provided below the x-axis labels. Error bars represent within-subject 95% confidence intervals (Morey, 2008). ***p < .001.
A two-way repeated measures ANOVA was conducted on oculomotor capture effects with the factors between trial bin and distractor salience. The analysis revealed a significant main effect of trial bin, F(4, 156) = 40.17, p < .001, η p 2 = .51, BFincl > 1,000, indicating an overall decrease in capture from Trial 1 to Trial 5. The main effect of distractor salience was not significant, F(1, 39) = 1.54, p = .221, η p 2 = .04, BFexcl = 3.60. Crucially, a significant interaction between trial bin and salience type was observed, F(4, 156) = 17.10, p < .001, η p 2 = .30, BFincl > 1,000. This indicates that the reduction in oculomotor capture was greater in the high-salience condition (18.9%) than in the low-salience condition (5.0%).
Next, a series of planned one-sample t tests was conducted to assess whether the oculomotor capture significantly deviated from zero across trial bins in each salience condition. In the high-salience condition, a significant initial capture effect was observed in Trial 1 (+12%), t(39) = 6.26, p < .001, dz = 0.99, BF10 > 1,000, followed by rapidly learned suppression on Trials 2 through 5, [Trial 2 (−3.3%): t(39) = −2.78, p = .008, dz = −0.44, BF10 = 4.81; Trial 3 (−4.7%): t(39) = −5.12, p < .001, dz = −0.81, BF10 > 1,000; Trial 4 (−5.7%): t(39) = −5.35, p < .001, dz = −0.85, BF10 > 1,000; Trial 5 (−6.9%): t(39) = −8.51, p < .001, dz = −1.35, BF10 > 1,000]. In the low-salience condition, there was a nonsignificant trend of a small capture effect on Trial 1 (+2.3%), t(39) = 2.11, p = .041, dz = 0.33, BF10 = 1.25, but this did not pass the significance threshold of the FDR adjustment for multiple comparisons (p < .02). No significant deviation from zero was observed on Trials 2 through 4 (ps > .05), with all BF01 values between 1.73 and 5.75. A small suppression effect emerged in Trial 5 (−2.7%), t(39) = −3.17, p = .003, dz = −0.50, BF10 = 11.68, after prolonged exposure.
Pairwise comparisons further revealed that initial capture in high-salience conditions was significantly stronger than in the low-salience conditions on Trial 1 (+12% vs. +2.3%), t(39) = 4.57, p < .001, dz = 1.00, BF10 = 477.60, whereas suppression in Trials 2 through 5 was significantly greater in the high-salience condition (ps < .05). Specifically, on Trial 5, oculomotor suppression was stronger for the high-salience distractor than for the low-salience distractors (−6.9% vs. −2.7%), t(39) = 3.83, p < .001, dz = 0.43, BF10 = 61.27.
In sum, the current results demonstrate that the strength of initial capture (on Trial 1) corresponds to the strength of learned suppression (on Trials 2–5). When initial capture did not occur in the low-salience condition, learned suppression was weakened in later trials.
Learned suppression as a function of initial capture
As an exploratory analysis, we evaluated whether capture on Trial 1 improved the development of learned suppression on Trials 2 through 5 by computing learning slopes. We classified whether the first eye movements were directed to the salient distractor on Trial 1 (capture vs. no capture). To improve the power to conduct this analysis, we collapsed this analysis across both salience conditions. Participants were excluded from the analysis because of inadequate power to compute a learning curve (i.e., resulting in less than 10 mean observations in Trials 2–5). Four participants were excluded for this reason.
We next examined how capture influenced the rate of learned suppression by fitting linear slopes to the reduction in first eye movements to salient distractors on Trials 2 through 5. The same pattern was observed as in Experiment 1: The reduction in capture was significantly greater following capture (slope = −1.8% per trial) than when capture did not occur (slope = −0.5% per trial), t(35) = 2.09, p = .044, dz = 0.35, BF10 = 1.24. This analysis provides additional evidence that initial capture improved learned suppression, as predicted by the capture-dependent suppression account.
Learning effects across the experiment
We compared initial capture effects for the high-salience condition on Trial 1 between the first and second halves to examine whether participants would learn to suppress distractors across the experimental session. The results did not reveal a significant difference between the first and the second half (+13% vs. +11%), t(39) = −0.67, p = .508, dz = −0.11. A complementary Bayesian analysis provided moderate evidence for the null hypothesis (BF01 = 4.75), indicating that the initial capture effect was relatively stable throughout the entire experimental session. We also calculated learned effects by using the oculomotor capture effects on Trial 1 minus Trial 5. These effects did not differ significantly in the first half (19.4%) and the second half of the experiment (18.3%), t(39) = −0.40, p = .693, dz = −0.06, and a Bayesian analysis again provided moderate support for the null hypothesis (BF01 = 5.44). Altogether, this suggests that the pattern of learned suppression over the course of a color run was relatively stable across the experimental session.
Saccadic latency
Figure 4b depicts saccadic latency for each trial bin with the same color (underneath the trial bin), with data points for the high-salience condition enclosed in black rectangles and those for the low-salience condition enclosed in gray rectangles. As a reminder, a saccade-slowing account would predict that learned suppression is accomplished merely by slowing saccades to improve top-down control. However, the opposite pattern was also observed here: Saccadic latencies were significantly faster on Trial 5 than Trial 1 for both the high-salience condition (184 vs. 190 ms), t(39) = −3.88, p < .001, dz = −0.61, BF10 = 70.42, and the low-salience condition (183 vs. 198 ms), t(39) = −7.76, p < .001, dz = −1.23, BF10 > 1,000. Thus, the observed learned suppression effects cannot be attributed to a slowing of eye movements to prevent capture.
Dwell time
We also evaluated whether reactive learned suppression was improved with experience by evaluating dwell time on singletons as a function of trial bin (Table 2). As in Experiment 1, dwell times were slower on Trial 1 compared with later trials (Trials 2–5) for both high- and low-salience conditions. A two-way repeated-measures ANOVA was conducted on dwell time with two within-subject factors: trial bin (Trials 1–5) and distractor salience (low-salience and high-salience). There was a significant main effect of trial bin, F(4, 156) = 8.27, p < .001, η p 2 = .17, BFincl > 1,000, indicating that dwell time to the salient distractor systematically decreased across trials. The main effect of distractor salience was also significant, F(1, 39) = 10.16, p = .003, η p 2 = .21, BFincl = 6.86, with dwell time being longer in the low-salience condition than in the high-salience condition. In addition, there was a marginally significant interaction between trial bin and distractor salience, F(4, 156) = 2.13, p = .08, η p 2 = .05, BFincl = 2.25. Taken together, these results demonstrate that learned suppression improved with experience across the color run and provide some evidence that this process is modulated by distractor salience.
Discussion
Experiment 2 compared high- and low-salience distractors using the same task in which display colors changed every five trials. The results showed that high-salience distractors initially captured attention and then were suppressed on subsequent trials. Low-salience distractors did not initially capture attention, and learned suppression was greatly weakened on future trials. These results confirm the predictions of the capture-dependent suppression hypothesis and help to strengthen the proposed relationship between initial capture and learned suppression.
Experiment 3
Experiment 3 tested whether reducing initial capture would weaken learned suppression. Participants completed a task similar to Experiments 1 and 2, but the display colors changed every three trials (Fig. 5). Notably, there were two conditions meant to manipulate the strength of attentional capture in initial trials (Trials 1 and 2). In the central-task condition, participants identified a small letter at fixation, and eye movements were prohibited during this central task. We expected that this would reduce attentional capture by focusing overt and covert attention at the center of the display. In the search-task condition, participants searched for a target shape, and capture was permitted to occur.
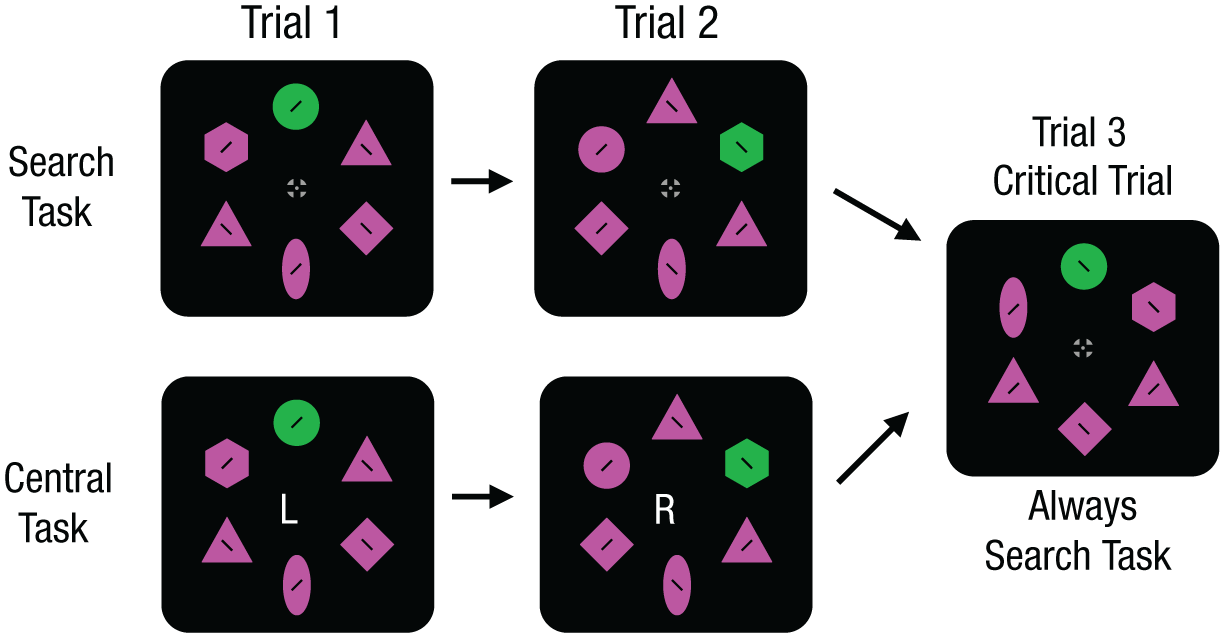
Stimuli task for Experiment 3. Experiment 3 used the same color-blocking technique, but the colors changed every three trials. Separate tasks manipulated the strength of capture on initial trials (Trials 1 and 2). For the central task, participants responded to a letter presented at fixation, which was meant to reduce capture on initial trials. For the search task, participants searched for a target shape as in prior experiments, and capture was permitted to occur as usual. The key question was whether suppression would occur on the critical trial, which was always a search task (Trial 3).
The key prediction relates to performance on the critical trial (Trial 3), which was always a search task. The capture-dependent suppression account predicts that suppression should be weakened on the critical trial if preceded by the central task (which reduced initial capture) than if preceded by the search task (which allowed initial capture).
Method
All methods were identical to Experiment 1, except as follows. Forty-two participants were recruited from the University of Missouri for payment of $20 per hour (21 men and 21 women, Mage = 21.8 years). As explained below, 2 participants were removed as outliers, resulting in a final sample of 40.
The eye-tracking task in Experiment 3 was similar to that used in Experiments 1 and 2 (Fig. 5). In this experiment, the display colors changed every three trials, and the distractors were always highly salient, which was 180° from the target color on the color wheel. Notably, there were two different tasks. Before each trial began, a pre-cue was presented for 1,000 ms that indicated the upcoming task to perform (“search” or “central”). In the search-task condition, participants performed a search task in which they looked for a target shape and attempted to ignore a salient distractor (similar to previous experiments). This should allow the salient distractor to capture attention on the initial trial and kick-start suppressive processes. In the central-task condition, participants held fixation at the center of a screen and classified a letter (L or R) using a game pad. This central task was meant to reduce attentional capture by the salient distractor by holding attention at the center of the screen. On these trials, any trial sequences where fixation was broken (1.4° from the center of the screen) were removed from the analysis. In addition, a feedback tone was played to indicate an erroneous eye movement to the participants (i.e., so they knew to try to avoid this).
Trials 1 and 2 were either the search task or the central task. Trial 3 (the critical trial) was always the search-task trial. The key question was whether reducing capture via a central task would impair suppression on the critical trial. According to our account, initial capture should increase future suppression. This account therefore predicts that suppression should be weakened more on critical trials after the central task than after the search task.
The experiment comprised 120 practice trials and 480 experimental trials, organized into 10 blocks. Participants were removed if the final proportion of included trials across all blocks fell below 75%. Two participants were removed because of this criterion. Trials with RTs less than 200 ms or greater than 2,000 ms were removed from analysis (2.9%). Trials with incorrect responses (2.4%) were omitted from all analyses but the manual error-rate analysis. In all, 5.1% of trials were excluded.
Results
Oculomotor capture and suppression on search trials
We first analyzed runs that consisted of search trials only (i.e., the top portion of Fig. 5). We expected to observe a similar trend as in Experiments 1 and 2: Trial 1 (color switch) should exhibit initial capture, followed by learned suppression on Trials 2 and 3. As shown in Figures 6a and 6b, we observed results consistent with this prediction: Capture preceded later suppression.
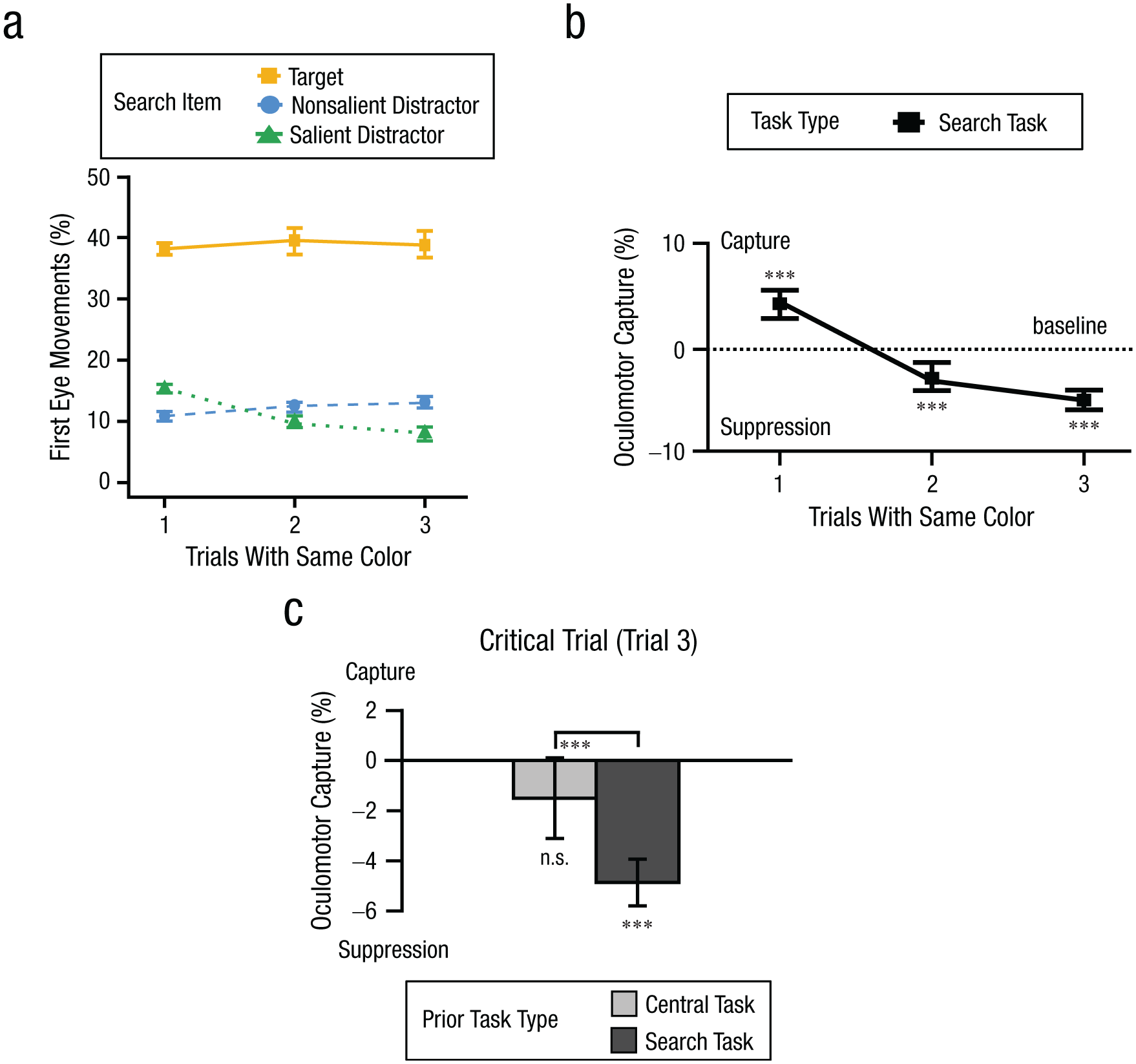
Results for Experiment 3. In (a) are shown first eye movements to each search item as a function of trial bin with the same color on search-task trials. In (b) we illustrate oculomotor capture effects across Trials 1 to 3 in runs of the search-task trials. A positive value indicates initial capture (Trial 1), whereas negative values indicate learned suppression (Trials 2 and 3). In (c), oculomotor capture effects are shown on the critical Trial 3 as a function of prior task type. Error bars represent within-subject 95% confidence intervals (Morey, 2008). ***p < .001.
A one-way repeated-measures ANOVA was conducted on oculomotor capture effects with the factor of trial bin (Trials 1–3) on runs of search trials. The analysis revealed a significant main effect of trial bin, F(2, 78) = 61.80, p < .001, η p 2 = .61, BFincl > 1,000, indicating a systematic shift in oculomotor capture across trials. To further explore this pattern, we conducted a series of planned one-sample t tests to assess whether oculomotor capture significantly deviated from zero across trials in the search-task condition. As shown in Figure 6b, the initial capture effect was significantly greater than zero (+4.3%) in Trial 1, t(39) = 5.84, p < .001, dz = 0.92, BF10 > 1,000, indicating a robust attentional bias toward salient distractors. This initial capture was rapidly followed by learned suppression, with significant suppression observed as early as Trial 2 (−2.7%), t(39) = −3.80, p < .001, dz = −0.60, BF10 = 56.57. This pattern of suppression strengthened in Trials 3 (−4.9%), t(39) = −8.68, p < .001, dz = −1.37, BF10 > 1,000, reflecting robust learned suppression triggered by initial capture, with the magnitude of suppression increasing over time.
Suppression on the critical trial
The key prediction is related to performance on the critical trial. Attentional capture should be reduced in the central task condition, where attention needs to be focused on the fixation cross. Thus, oculomotor suppression should be weaker on the critical trial (Trial 3) when it was preceded by a central task than when it was preceded by a search task. Figure 6c depicts oculomotor capture effects on the critical trial as a function of the task on the preceding two trials. As shown, the results conform to the prediction that the central task reduced suppression.
A planned one-sample t test was conducted to assess whether oculomotor capture significantly deviated from zero on Trial 3. In the central-task condition, the results did not reveal significant deviation from zero (−1.6%), t(39) = −1.67, p = .102, dz = −0.27, suggesting that capture was not significantly below baseline. A corresponding Bayesian analysis provided anecdotal evidence in favor of the null hypothesis (BF01 = 1.64). In the search-task condition, the results revealed that suppression was significantly below the baseline of zero, t(39) = −8.68, p < .001, dz = −1.37. A corresponding Bayesian analysis provided extreme evidence in favor of the alternative hypothesis (BF10 > 1,000). Critically, a paired t test on critical Trial 3 demonstrated that suppression was significantly larger when preceded by the search task than when preceded by the central task (−5.0% vs. −1.6%), t(39) = −3.80, p < .001, dz = −0.60, and a Bayesian analysis provided very strong evidence in favor of an effect, BF10 = 59.68. This provides evidence that the central task on Trials 1 and 2 reduced the strength of suppression on the critical trial.
We also compared the capture effect on Trial 3 (the critical trial) in the central-task condition with Trial 1 in the search-task condition. Oculomotor capture was significantly weaker on Trial 3 of the central-task condition than on Trial 1 of the search-task condition (−1.5% vs. +4.3%), t(39) = 5.12, p < .001, dz = 0.81, BF10 > 1,000. Thus, Trial 3 in the central-task condition was not equivalent to Trial 1 in the search-task condition. As suggested above, a potential explanation is that the central task reduced capture by the salient distractor but may not have fully prevented attentional capture: Covert attention could have shifted to display items on some trials and spread in parallel between the central letter and display items. Thus, some degree of covert capture may have occurred despite our best efforts to minimize it.
An additional possibility is that a form of latent learning to ignore the salient distractor occurred, in addition to proactive suppression. For example, the central task might have encouraged covert suppression of all peripheral items, or the central white target might itself function as a salient singleton that changes the learning context. In short, it is clear that some level of ignoring occurred despite our efforts to prevent capture. However, it is also clear (from the previous paragraph) that the central task did reduce learned suppression, consistent with the predictions of the capture-dependent suppression hypothesis. We will return to this issue in the General Discussion.
Discussion
The findings from Experiment 3 further strengthen the evidence of a relationship between initial capture and learned suppression. By using a central task to weaken initial capture, suppression was weakened on the critical search trial and did not occur below baseline levels when preceded by search trials. This suggests that capture is an important precursor that can increase learned suppression.
General Discussion
The present study tested a new hypothesis about how we learn to ignore salient stimuli. This capture-dependent suppression hypothesis proposes that an initial instance of capture acts as a learning signal that increases subsequent suppression of features and locations associated with a salient distractor. This new hypothesis therefore proposes a direct relationship between the initial attention-capturing power of a stimulus and the later ability to ignore it.
Three experiments examined this new hypothesis. In Experiment 1, we used a new task in which the display colors changed every five trials. Shifts of gaze were captured on the initial trial when the colors switched, followed by learned suppression, demonstrating that successful suppression begins with an initial instance of capture. Experiment 2 provided further evidence for a relationship between initial capture and learned suppression by manipulating distractor salience. High-salience distractors initially captured attention, followed by learned suppression. Low-salience distractors, however, did not initially capture attention and were also not as effectively suppressed. In Experiment 3, we further examined the role of initial capture in improving suppression by employing an experimental manipulation that prevented attentional capture from occurring during initial trials (i.e., with a central task that occupied attention). When initial capture was prevented, learned suppression was eliminated.
The current findings advance theories of attention in two important ways. First, the results highlight the need for a hybrid model of attentional control in which both bottom-up salience and top-down goals can influence attentional allocation, because evidence of both capture and control was found within each color run. Second, the results also highlight that learned suppression can develop rapidly, after only one or two exposures to a salient stimulus (see also Adam et al., 2024; Savelson et al., 2025), which is much faster than previously assumed (Gaspelin & Luck, 2018; Vatterott & Vecera, 2012). This indicates that attentional control is highly adaptive in applying feature-based suppression to salient distractors.
The current results also bridge an important gap between the signal-suppression account (Gaspelin et al., 2025) and theories of selection history (Anderson et al., 2021). For instance, the current hypothesis is similar to that of Anderson et al. (2021), who suggested that selection-history effects might require attention to the critical stimulus for learning to emerge. The current account also has similarities to the search-and-destroy account of Moher and Egeth (2012), which proposed that distractors must be attended to before they are ignored. The present findings could be viewed as an intertrial extension of this principle: Initial capture in a previous trial triggers suppression on a subsequent trial. Unlike search-and-destroy accounts, however, the signal-suppression account proposes that salient distractors are proactively suppressed without first being attended to after learning has occurred (Gaspelin et al., 2025).
In Experiment 3, capture was reduced in the central task condition on the critical trial. One possibility is that our central task did not entirely prevent capture, especially covert capture, which allowed some capture-dependent suppression to occur. However, some prior studies have also suggested that passive viewing can allow participants to latently habituate to salient stimuli and mitigate future distraction (Turatto, 2023; Won & Geng, 2020). Therefore, one possibility is that some level of latent learning occurred. It seems plausible that latent learning and capture-dependent suppression represent distinct mechanisms used to ignore distracting information but may also occur concurrently to guide attention. This will be an important question for future studies.
There are some limitations to the current findings. First, learning was manipulated by blocking the colors of salient distractors. Future research might evaluate whether similar results are observed for other forms of distractor suppression, such as those involving spatial biases (Wang & Theeuwes, 2018). Second, the current study used oculomotor measures of attention. Future research might therefore use measures of covert attention, such as ERP components, to further establish whether suppression depends on initial distraction (e.g., N2pc and PD component; Gaspelin et al., 2023; Luck, 2011). Third, the present findings may have limited generalizability because participants were college students recruited from a U.S. university; replication with a more diverse sample (e.g., broader age ranges and nonstudent community samples) may be useful in future research. Finally, future studies with larger samples and designs explicitly tailored to individual-differences approaches could further test whether capture variability reliably predicts suppression learning.
In conclusion, the present findings highlight a new principle about how distracting information is ignored. Rather than viewing distraction as a problem, we suggest that distraction may serve a functional purpose as a training signal to initiate future attentional control.
Footnotes
Transparency
Action Editor: Zhicheng Lin
Editor: Simine Vazire
Author Contributions