
Abstract
Statistical learning is a powerful mechanism that enables the rapid extraction of regularities from sensory inputs. Although numerous studies have established that statistical learning serves a wide range of cognitive functions, it remains unknown whether statistical learning impacts conscious access. To address this question, we applied multiple paradigms in a series of experiments (N = 153 adults): Two reaction-time-based breaking continuous flash suppression (b-CFS) experiments showed that probable objects break through suppression faster than improbable objects. A preregistered accuracy-based b-CFS experiment showed higher localization accuracy for suppressed probable (versus improbable) objects under identical presentation durations, thereby excluding the possibility of processing differences emerging after conscious access (e.g., criterion shifts). Consistent with these findings, a supplemental visual-masking experiment reaffirmed higher localization sensitivity to probable objects over improbable objects. Together, these findings demonstrate that statistical learning alters the competition for scarce conscious resources, thereby potentially contributing to established effects of statistical learning on higher-level cognitive processes that require consciousness.
Keywords
Since the inception of psychology as a scientific discipline, the question of how conscious experiences arise has been central. Our visual environment is characterized by structures and regularities. Is our conscious experience of the visual environment shaped by our past experiences of these regularities? This question is important, because, as human beings, we naturally seek to recognize patterns in features (e.g., the red and circular properties of an apple) and understand temporal and spatial properties of events (e.g., the arrangement and sequence of traffic lights). This inherent ability to extract distributional regularities from the environment, serving as a foundation for the behavioral functioning and adaptability of organisms in their surroundings, is commonly referred to as statistical learning. Over the past two decades, statistical learning has become a major area in cognitive research, as is indicated by its pervasive influence on a wide range of basic and higher-level cognitive processes (for reviews, see Bogaerts et al., 2020; Frost et al., 2019; Sherman et al., 2020). These processes include language acquisition (e.g., Saffran et al., 1996), perception of high-level perceptual units (e.g., events; Brady & Oliva, 2008), recognition and association of meaningful chunks for learning, memory (e.g., Brady et al., 2009), and social inference (e.g., Dotsch et al., 2017). Among these cognitive functions, many are predicated on the acquisition of conscious resources (e.g., Sabary et al., 2020). Surprisingly, despite extensive exploration on the functions of statistical learning, it is unknown whether statistical learning regulates the conscious access of sensory inputs before the level at which conscious functions (e.g., memory, language, inference) are regulated. To understand how statistical learning influences such higher-tier cognitive functions, one must first establish whether statistical learning affects conscious access of sensory inputs.
The hypothesis that statistical learning affects conscious access is supported by the overlap between the behavioral correlates of statistical learning and conscious access. Previous studies suggested that statistical learning generates memory chunks (Orbán et al., 2008), induces implicit anticipations (Turk-Browne et al., 2010), and regulates the allocation of attention (e.g., Wang & Theeuwes, 2018) and working memory resources (e.g., Brady et al., 2009; Umemoto et al., 2010). Given that these factors have been shown to influence the detection of interocularly suppressed stimuli (working memory, Gayet et al., 2013; anticipation, Denison et al., 2011, 2016; Pinto et al., 2015; and perhaps attention, Thibault et al., 2016—but see Gayet et al., 2020), one might hypothesize that statistical learning directly affects conscious access. Alternatively, it is conceivable that statistical learning does not affect conscious access, because similar learning processes (e.g., perceptual learning) do not affect detection of stimuli under interocular suppression (Mastropasqua et al., 2015; Paffen et al., 2018).
To investigate whether statistical learning affects conscious access, we first used a reaction-time-based breaking continuous flash suppression paradigm (b-CFS; Jiang et al., 2007) in Experiments 1 and 2 (Fig. 1). Here, the time it takes for observers to report some aspect of interocularly suppressed targets is related to the competitive strength of stimuli for entering visual awareness (e.g., higher-contrast targets will break suppression faster than low-contrast ones). To exclude the possibility that differential reaction times in b-CFS can reflect differences emerging after conscious access (e.g., decisional and postperceptual processes; Stein & Peelen, 2021), we further conducted two follow-up experiments. In Experiment 3, we used an accuracy-based b-CFS paradigm (Litwin et al., 2023) in which the localization accuracy for interocularly suppressed high- and low-probability feature stimuli were compared. In addition, a supplemental experiment (see Supplemental Material 2 in the Supplemental Material available online) used a visual-masking technique (Stein & Peelen, 2021) to compare the localization sensitivity to masked high- and low-probability feature stimuli.
Statement of Relevance
Our visual environment provides us with a continuous stream of complex information, most of which is highly structured. Cars, for example, have prototypical locations (e.g., on the road rather than in the sky) and share prototypical visual characteristics (e.g., a horizontally elongated shape). It is known that human beings are extremely proficient at extracting such structural regularities to facilitate a wide range of cognitive functions. However, it is unknown whether this prior learning of environmental structures shapes our consciousness—a core aspect of human cognition. The present study revealed for the first time that statistical learning can influence the selection of sensory input for conscious perception, prioritizing probable events over improbable events in conscious access. The direct effect of statistical learning on consciousness may explain how statistical learning serves a wide range of cognitive functions that benefit from or depend on consciousness, such as memory, learning, and inference.
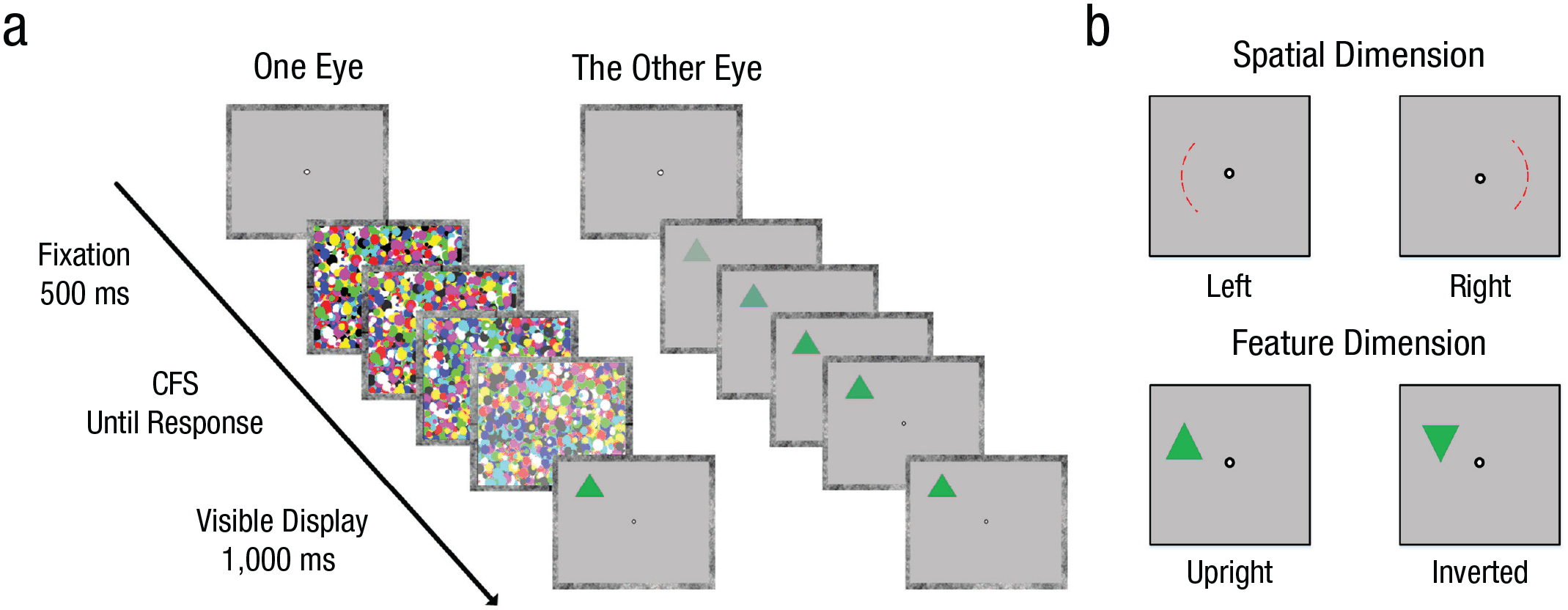
Trial overview of the breaking continuous flash suppression (b-CFS) paradigm used in Experiments 1 and 2 (a). Targets could appear on the left or right of fixation and could consist of upright or inverted triangles (b).
Open Practices
All materials, data, and analysis scripts for all experiments have been made publicly available via the Open Science Framework (OSF) and can be accessed at https://osf.io/2v4dh. Experiment 3 (https://osf.io/w7g25) and one supplemental experiment (Supplemental Experiment B, https://osf.io/y96ba) were preregistered at OSF.
Method (Experiment 1)
In Experiment 1, we examined whether the presence of statistical regularities of target locations influences conscious access to targets. Specifically, if statistical learning of spatial locations affects conscious access, we should observe shorter reaction times (in an orthogonal discrimination task) for targets that appear at high- versus low-probability locations.
Participants
The predetermined sample size for this first experiment was loosely based on previous statistical-learning studies (Wang & Theeuwes, 2018). Twenty-five healthy participants, naive to the purpose of the experiment, participated in Experiment 1 after signing an informed consent form. One observer who did not follow instructions (i.e., the participant did not look through the binocular stereoscope) was excluded from all data analyses and was replaced. The eventual sample contained 24 participants (19 women and 5 men, mean age = 24.13 years, SD = 2.49 years) for data analysis. All participants had normal or corrected-to-normal vision, and none had color blindness. The experiment was approved by the Ethics Committee of the Faculty of Social and Behavioral Sciences of Utrecht University. Participants received monetary compensation for their participation.
Apparatus and stimuli
The experiment was conducted on a 27-in. LCD monitor (2,560 × 1,440 pixels, 100-Hz refresh rate). The experiment took place in a darkened laboratory with all light sources turned off except for the computer screen, which was positioned at an effective viewing distance (the distance the light travels from monitor to eye) of 57 cm. The presentation area on the screen was composed of two parts (half-images presented on the left and right half of the monitor) which were viewed dichoptically through a stereoscope mounted on a chin rest. The stereoscope made it possible to independently stimulate the left and right eyes of participants, thus triggering interocular competition. To promote binocular fusion of the two competing images, we ensured that each display that contained a competing image had a gray background (16.2 cd/m², x = 0.283, y = 0.298, (9.5° × 7.2°) surrounded by a Brownian noise square frame with a thickness of 0.5°. The remaining part of the screen was set as a uniform black background with luminance of 0.05 cd/m².
The target stimulus consisted of an upright or inverted triangle (1.4° in height and length) that was presented to a single eye. This target was distanced 2.4° from a central fixation dot (a 0.3° white dot with a black edge). The other eye was presented with high-contrast colored masks that were changed at a rate of 10 Hz. These masks were made up of randomly arranged circles (diameter 0.3° to 1.5°) of different colors (they differed both in hue and luminance). One hundred and twenty different CFS masks were generated before the experiment and appeared in a random order (without replacement) across different trials. The dynamic high-contrast CFS masks were presented to one eye to perceptually suppress the static stimulus shown to the other eye, thereby rendering the static stimulus initially unconscious. The experiment was programmed using the Psychophysics Toolbox (Brainard, 1997) in MATLAB (2021a; The MathWorks, Natick, MA). Data analysis was performed in JASP (2022; Version 0.16.1; Love et al., 2019).
Procedure
Each trial started with a fixation dot that appeared at the center of the screen for 500 ms. After this, a dynamic CFS mask was presented to one eye of the participant. Meanwhile, the target was presented to the other eye of the participant with its intensity (i.e., opacity) ramping up from zero to full opacity within 1,000 ms (Fig. 1a). At the start of the trial, participants were unable to consciously perceive the triangle because it was interocularly suppressed by the CFS masks. Over time, the visibility of the dynamic CFS mask presented to one eye was gradually reduced, with its transparency increasing gradually. The increasing intensity of the target and the decreasing intensity of the mask jointly caused the target to be eventually released from interocular suppression, thus allowing participants to report upon the target. The presentation of the CFS masks and the target ended when a participant responded or after 10 s had passed. Participants were required to press one of two buttons to indicate the orientation of the triangle as quickly and accurately as possible (↑ for upright triangles, ↓ for inverted triangles). We manipulated how often a triangle was presented at different locations (left vs. right) but required participants to report triangle orientation (upright vs. inverted); see Figure 1b. Consequently, the response mapping was orthogonal to the experimental manipulation, minimizing the influence of response bias.
Targets (triangles) appeared on a specific side, either the left or right side of the screen, on 75% of all trials (the high-probability condition), and appeared on the other side of the screen on the remaining 25% of all trials (the low-probability condition). The location (left vs. right sides) being set as the high-probability location was counterbalanced between participants. In contrast, the orientation of triangles was counterbalanced (with random presentation orders) within participants across different trials. The distance from the target to the fixation dot was the same in all trials; that is, the target was positioned on the outline of an imaginary circle with a radius of 2.4°. To preserve a clear distinction between the left and right of fixation, we made sure that targets could appear only in the left and right quadrants of the imaginary circle on one of 20 equally interspersed locations. The exact position within a quadrant was drawn at random (and thus not counterbalanced), so that targets had an equal probability of appearing either (slightly) above or below the horizontal midline.
After the response, participants kept viewing the target display with both eyes for 1,000 ms. Meanwhile, participants also received sound feedback (500 ms): a high-pitched beep sound (2000 Hz) or a low-pitched beep sound (1500 Hz) for correct or incorrect responses, respectively. This phase was not only a feedback phase but also served as an important learning phase: Given that it is unclear whether statistical regularities can be extracted from interocularly suppressed stimuli, we showed the targets without suppression at the end of each trial to ensure that participants could learn the statistical regularities from stimuli that were not interocularly suppressed.
The formal experiment was composed of eight practice trials and six blocks of 32 formal trials. Here, we manipulated the prevalence of the target locations, so that it was more likely to appear to the left of fixation (75% of trials) than to the right of fixation for half of the participants, and vice versa for the other half of the participants. We refer to these as regularity blocks. The color of the target was green (7.70 cd/m², x = 0.288, y = 0.444 at full opacity) in the regularity blocks. To enhance the motivation of participants, we provided them with feedback about their average performance at the end of each block, showing their mean reaction times and the mean accuracy of that block. After checking the performance, participants had the opportunity to take a self-initiated rest. We set up a mandatory rest every three blocks for all participants. In addition, before the formal experiments, we also asked participants to finish three blocks of 32 trials in which the statistical regularity was absent (nonregularity blocks); see Supplemental Material 3.
To probe participants’ explicit knowledge about the existence of the manipulated regularities, we asked participants to estimate the probability of targets appearing on the left or right of fixation (the regularity dimension), as in previous studies (e.g., Wang & Theeuwes, 2018). Notably, this coarse post hoc measurement alone is insufficient for drawing conclusions about the implicit nature of statistical learning (see Giménez-Fernández et al., 2020; Vadillo et al., 2022). The modest sample size in our experiments might not guarantee adequate power for relating the magnitude of the effect of statistical learning on conscious access to individual participants’ awareness of the regularities (Vadillo et al., 2020). However, the debated (un)consciousness of statistical learning (e.g., Conway, 2020; Giménez-Fernández et al., 2020; Turk-Browne et al., 2005) is not the focus of our study. For the transparency of the results, we present descriptive statistics and correlation analysis, with a cautionary note that these statistics cannot be taken as definitive evidence for or against participants’ awareness of the regularities.
Results (Experiment 1)
The b-CFS task
Incorrect responses were excluded from all data analyses (2.41% of all trials). The accuracy of participants ranged from 89.58% to 100%, with an average accuracy of 97.59% (SD = 0.02). To test whether statistical learning develops over time, we further compared the reaction times (RTs) to targets at high- and low-probability locations across time. To this end, we divided the experiment into epochs of 64 trials (the conditions, high versus low probability, were fully counterbalanced within each of these individual epochs). We conducted a repeated-measures ANOVA with the factors of probability (high vs. low) and epoch (1 to 3) to investigate how the influence of statistical regularities on RTs would evolve over time.
Results showed that the main effect of probability was significant, F(1, 23) = 6.05, p = .022, η p 2 = .21. Specifically, RTs for the high-probability locations (3.66 s, SD = 1.81) were 0.47 s shorter than for the low-probability locations (4.13 s, SD = 2.00); see Figure 2a. The shorter RTs for targets presented at high-probability locations indicate that visual input gains faster access to consciousness when appearing at a probable (rather than improbable) location. The main effect of epoch was also significant, F(2, 46) = 8.84, p < .001, η p 2 = .28, reflecting a general decrease in RTs over the course of the experiment. Importantly, the absence of an interaction between probability and epoch, F(2, 46) = 0.30, p = .742, η p 2 = .01, suggests that the difference in RTs to targets appearing on high- compared to low-probability locations did not change over the course of the experiment (Fig. 2b). To test whether the statistical-learning effect appeared at early stages of the experiment, we further conducted t tests to compare RTs to targets on high- and low-probability locations in the first epoch. Results showed that RTs for the high-probability locations were already shorter than for the low-probability locations in Epoch 1, t(23) = 3.26, p = .003, Cohen’s d = 0.67, 95% confidence interval (CI) = [0.20, 0.90]. These data show that participants rapidly extracted the statistical regularities of the target location and that the effect of statistical regularities on conscious access did not change over time.
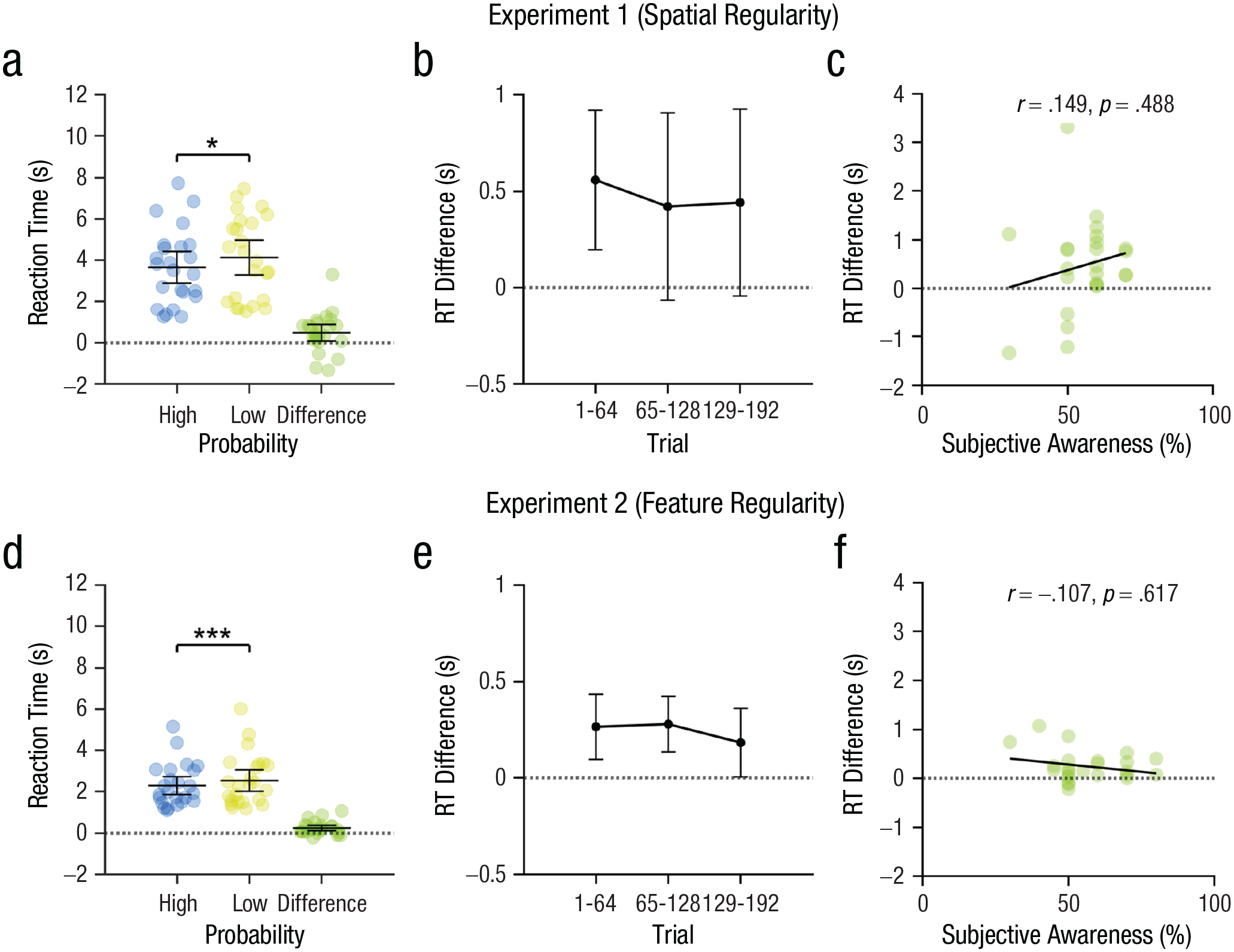
The results of Experiments 1 and 2. In (a) and (d), mean reaction times (RTs) in the high-probability and low-probability conditions are displayed, with individual dots representing individual participants; in (b) and (e), RT differences between the high- and low-probability conditions over time are shown. The correlation between participants’ RT difference and their subjective awareness of the statistical regularities (i.e., their estimates of the probability of a high-probability location or feature occurring) are illustrated in (c) and (f). Error bars represent 95% confidence intervals of the mean. Asterisks indicate significance. *p < .05. ***p < .001.
Intertrial priming effects
An alternative account for the RT difference between the high- and low-probability trials could be an intertrial priming effect. For example, a previous study suggested that targets repeated on successive trials gain preferential access to consciousness (Peremen et al., 2013). There were more consecutive trials for the high-probability location trials than for the low-probability location trials in our study, so it is possible that the faster target detection in high-probability trials was due to the advantage caused by such intertrial priming instead of the learned regularities. To exclude such a possibility, we compared RTs for high-probability trials preceded by a high-probability trial with those preceded by a low-probability trial, and we compared RTs for low-probability trials preceded by a low-probability trial with those preceded by a high-probability trial (Golan & Lamy, 2023). Results showed that for high-probability trials, there was no difference in RTs between trials that were preceded by a high-probability trial (3.66 s, SD = 1.81) and those that were preceded by a low-probability trial (3.63 s, SD = 1.84), t(23) = 0.43, p = .673, Cohen’s d = 0.09, 95% CI = [–0.13, 0.19]. For low-probability trials, we also found no difference in RTs between trials preceded by a low-probability trial (4.09 s, SD = 2.21) and those preceded by a high-probability trial (4.12 s, SD = 1.97), t(23) = 0.21, p = .838, Cohen’s d = 0.04, 95% CI = [–0.34, 0.41]. These results show that the observed difference between RTs for high- and low-probability locations is unlikely to have been caused by intertrial priming.
The subjective-awareness ratings
Out of the 24 participants, there were 14 participants who correctly guessed the regularity (no preferences: 8; incorrect guesses: 2). A rank-based Spearman’s correlation test (p < .05 in the Shapiro-Wilk test of normality) indicates that in our sample there was no significant correlation between the calculated subjective-awareness score and the difference in RTs between high- and low-probability locations in the b-CFS task (r = .15, p = .488; see Fig. 2c), suggesting that the effect on conscious access did not depend on the level of awareness of the regularity. As mentioned above, the results here provide only weak evidence for the level of awareness of the regularity of the participants and need to be interpreted with caution because of the limitations of the methods (e.g., lack of statistical power).
Method (Experiment 2)
Experiment 2 was aimed at extending the findings of Experiment 1 by testing whether the presence of statistical regularities of target features (instead of target locations) also accelerates conscious access of targets comprising high-probability features. If statistical learning affects conscious access at the level of features, we should observe shorter reaction times to targets comprising high-probability (vs. low-probability) features.
The methods were generally identical to those of Experiment 1 except for the following changes. First, a new group of 24 healthy participants (20 women and 4 men, mean age = 25.08 years, SD = 3.17) were recruited for the experiment. The sample size was set to match that of Experiment 1. Second, in the regularity blocks, we manipulated the probability of targets being upright or inverted, instead of manipulating the probability of target locations. Participants were asked to determine target locations (left or right) as quickly as possible, instead of reporting the target orientation (see Fig. 1).
Results (Experiment 2)
The b-CFS task
Incorrect responses were excluded from all data analyses (1.54% of all trials). The accuracy of participants ranged from 94.79% to 100%, with an average accuracy of 98.46% (SD = 0.01).
As in Experiment 1, we also conducted a repeated-measures ANOVA with the factors of probability (high vs. low) and epoch (1 to 3) in Experiment 2. Results showed that the main effect of probability was, again, significant, F(1, 23) = 15.19, p < .001, η p 2 = .40. Specifically, RTs for the high-probability features (2.30 s, SD = 1.03) were 0.24 seconds shorter than for the low-probability features (2.54 s, SD = 1.23); see Figure 2d. The shorter RTs for targets presented with high-probability features indicated that visual input gains faster access to consciousness when appearing with a probable (rather than improbable) feature. Apart from this, the main effect of epoch was also significant, F(2, 46) = 14.34, p < .001, η p 2 = .38, which reflects a general decrease in RTs over the course of the experiment.
The interaction between probability and epoch was not significant, F(2, 46) = 1.22, p = .305, η p 2 = .05, suggesting that the difference in RTs to targets with high- compared to low-probability features did not change over the course of the experiment (Fig. 2e). As in Experiment 1, we further conducted t tests to compare RTs to targets on high- and low-probability features in the first epoch; this showed that RTs for the high-probability features were already shorter than for the low-probability features in Epoch 1, t(23) = 3.39, p = .003, Cohen’s d = 0.69, 95% CI = [0.11, 0.44]. These data show that the statistical regularities of the target feature were extracted rapidly and affected conscious access accordingly.
Intertrial priming effects
As in Experiment 1, we conducted analyses to exclude possible intertrial priming effects. Results showed that for high-probability trials, there was no difference in RTs between trials preceded by a high-probability trial (2.30 s, SD = 1.04) and those preceded by a low-probability trial (2.27 s, SD = 1.04), t(23) = 0.41, p = .688, Cohen’s d = 0.08, 95% CI = [–0.11, 0.17]. For low-probability trials, we also found no difference in RTs between trials that were preceded by a low-probability trial (2.46 s, SD = 1.40) and those preceded by a high-probability trial (2.56 s, SD = 1.22), t(23) = 0.76, p = .452, Cohen’s d = 0.16, 95% CI = [–0.16, 0.35]. Therefore, the differential RTs between high- and low-probability features is unlikely to have been caused by intertrial priming.
The subjective-awareness ratings
Out of the 24 participants, 11 participants correctly guessed the regularity (no preferences: 9; incorrect guesses: 4). A rank-based Spearman’s correlation test (p < .05 in the Shapiro-Wilk test of normality) showed that there was no significant correlation between the calculated subjective-awareness score and the difference in RTs between high- and low-probability features in the b-CFS task, r = –.11, p = .617 (see Fig. 2f). Again, these findings provide only weak evidence for the absence of a relation between awareness of the regularity and conscious access, and they should be approached with caution because of the methodological constraints (e.g., lack of statistical power).
Method (Experiment 3)
The goal of Experiment 3 was to test whether statistical learning affects conscious access by excluding processing differences emerging after conscious access (e.g., shifts in response criteria, which affect RTs). To this end, we used an accuracy-based variant of the b-CFS paradigm (Litwin et al., 2023) in which we compared the localization accuracy for high-probability versus low-probability features, given the same stimulus-presentation time. The methods and hypothesis of this experiment were preregistered before data collection (https://osf.io/w7g25).
In this paradigm, participants performed a nonspeeded two-alternative forced-choice (2AFC) localization task during viewing of a b-CFS presentation (i.e., CFS masks to one eye and a target to the other eye). The duration of the CFS presentation was predetermined before every trial and kept identical between conditions of interest (high- and low-probability conditions, in our case). Because responses in this paradigm are nonspeeded, participants’ responses on the forced-choice task reflect how much information they obtained about a stimulus within a given presentation duration. If for a specific presentation duration (e.g., yielding ~80% localization accuracy on average across high- and low-probability conditions), participants have more information about the identity or location of a stimulus in condition A compared to condition B, we can establish that conscious access of the stimulus (the identity or location) was faster in condition A than in condition B. This precludes any effect of decisional biases and postdetection effects, because it exhaustively measures the amount of information available to the participant within a specific time frame (Litwin et al., 2023).
Participants
After reviewing the effect size (Cohen’s d = 0.34) of a previous study that used the bias-free b-CFS paradigm to measure conscious access (Litwin et al., 2023), we concluded that a sample of 55 participants was needed for an experimental power of 80% with an alpha level of 0.05 for a planned one-tailed paired-samples t test (power calculation performed in G*Power). We opted to preregister a one-tailed test because we had clear predictions on the directionality of the effect following Experiments 1 and 2 and the supplemental experiment (see Supplemental Material 2). To counterbalance the between-subjects condition (i.e., upright or inverted triangles as the high- or low-probability feature), we recruited one more participant than was specified in the preregistration (i.e., 56 participants). A new group of 61 participants was recruited. For each participant, we simultaneously ran two independent staircases (for stimuli appearing left and right of fixation). According to the preregistered analysis plan, we excluded 5 participants whose average accuracy in both staircases was lower than 65% or higher than 95% from data analysis. This resulted in the planned sample size of 56 participants (49 women and 7 men, mean age = 24.59 years, SD = 2.77).
Apparatus and stimuli
The apparatus and stimuli used in Experiment 3 were the same as in Experiments 1 and 2.
Procedure
For any given participant, either the upward pointing triangle or the downward pointing triangle was selected as the high-probability feature (e.g., the triangle pointed upward in 75% of all trials), whereas the other one was the low-probability feature (e.g., the triangle pointed downward in the remaining 25% of all trials). Which one was selected as the high-probability feature was counterbalanced across participants. Within both the low- and high-probability conditions, different target location conditions (left or right of fixation) occurred equally often.
At the beginning of each trial, a central white fixation dot (with a black edge) appeared at the center of the presentation area for 500 ms (Fig. 3a). After this, a dynamic CFS mask (consisting of a number of Mondrian images, randomly chosen from 120 generated images, and replaced at 10 Hz without repetition) randomly appeared to the dominant eye of participants. Between 300 and 600 ms after the onset of the dynamic CFS mask (to the dominant eye), the target (a triangle) was presented to the nondominant eye of the participant (either to the left or right side of fixation, with equal probability), and remained on the screen for the duration that was predetermined by the staircase procedure (ranging between 1 and 6 s). During the presentation of the target, the intensity (i.e., opacity) of the target linearly ramped up within 2 s from zero to the eventual opacity (30%, 50%, or 60% of the original stimulus opacity of Experiments 1 and 2, depending on performance in the practice session; see Supplemental Material 4), regardless of the determined target presentation duration.
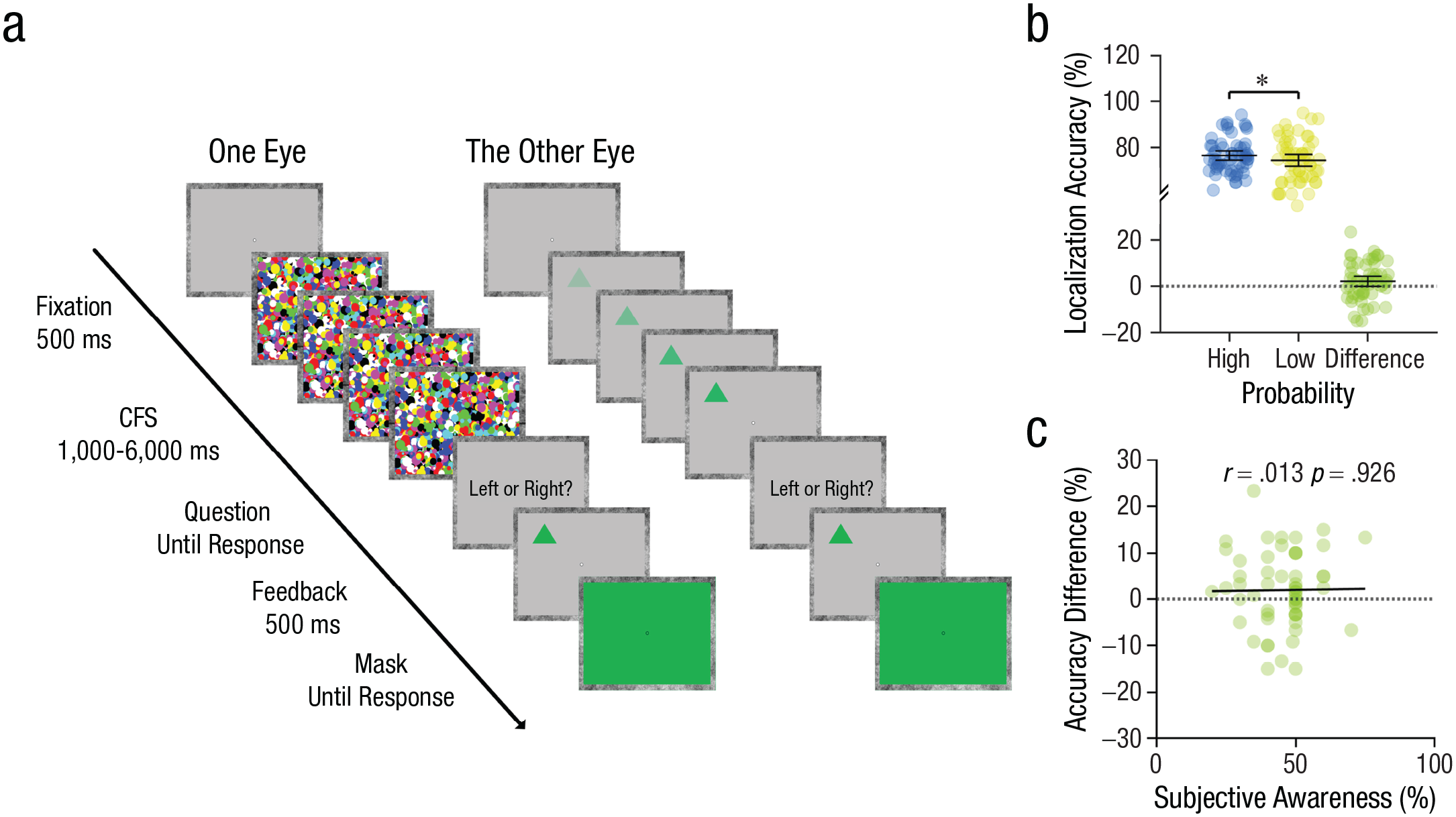
Trial outline (a) of the accuracy-based breaking continuous flash suppression (b-CFS) paradigm used in Experiment 3. The localization accuracy in the high-probability and low-probability conditions is shown in (b); the correlation between accuracy difference and subjective awareness is shown in (c). Individual dots represent individual participants, and error bars represent 95% confidence intervals of the mean. Asterisks indicate significance (*p < .05).
After the presentation of target and mask stimuli, a message—“presented left or right?”—appeared on the screen, requiring participants to press one of two arrow keys (← for left, → for right) to indicate on which side of fixation the target was presented (i.e., a 2AFC localization task). Participants were instructed to respond as accurately as possible, without any time pressure. After the response, the target stimulus remained present for 500 ms for both eyes, which ensured that participants got the opportunity to learn that one triangle orientation was more prevalent than the other. At the same time, they received auditory feedback—a high-pitched beep sound (2000 Hz) or a low-pitched beep sound (1500 Hz)—indicating a correct or incorrect response, respectively. At the end of the trial, the presentation area was filled with the same (green) color that was used for the target triangle to minimize afterimages at the target location before onset of the next trial. After pressing the space bar, the next trial began.
To avoid ceiling or floor effects (or to have more trials available for data analysis), we had to keep the overall localization performance at a consistent level across different visual fields and different phases of the experiment. To this end, we used a two-down/one-up adaptive staircase method that allowed the algorithm to reliably converge on the individual presentation-time thresholds to yield a localization accuracy of 80.35% (García-Pérez, 2001). We ran two interleaved staircases for different presentation positions (left vs. right side of the central fixation, respectively), considering the differences in nasal and temporal visual hemifields (Sahakian et al., 2022). The localization performance of the high- and low-probability trials were thus compared within staircases first and averaged afterward. More details for controlling the overall localization performance can be found in Supplemental Material 4.
Participants completed 24 trials for determining eye dominance and 32 practice trials in the pre-experiment, and then completed five blocks of 32 trials in the formal experiment. At the end of the experiment, we measured participants’ awareness of statistical regularities in a questionnaire, as in Experiments 1 and 2. The results of the awareness test, although not preregistered, are included here to ensure completeness.
Data analysis
According to the preregistered analysis plan, we used a one-tailed paired-samples t test to compare the localization accuracy of the high-probability condition to that of the low-probability condition. To avoid ceiling or floor effects, we excluded the data from an entire staircase (i.e., left or right target location) if the average accuracy of that staircase exceeded the predefined ceiling or floor (i.e., accuracy < 65% or > 95%). According to this preregistered data-exclusion principle, 5 participants were completely excluded from data analysis, and for 20 participants, the data from one staircase were excluded. For each of these 20 participants, the exclusion of one staircase may have resulted in decreased precision of estimation for the average localization accuracy. However, a considerable number of trials (80 trials) were left after data exclusion. In addition, similar to Experiments 1 and 2, we included an unregistered exploratory test to examine intertrial priming effects. Because of the insufficient number of trials, no additional data filtering was applied to the trials of included participants. The descriptive statistics and the correlation analysis for the relationship between the statistical-learning effect on conscious access and participants’ awareness of the regularities are also provided.
Results (Experiment 3)
The b-CFS task
Results showed that localization accuracy was higher for high-probability targets (0.77, SD = 0.08) compared to low-probability targets (fraction correct = 0.75, SD = 0.10), t(55) = 1.88, p = .033, Cohen’s d = 0.25, 95% CI = [0.00, ∞]; see Figure 3b. The higher localization accuracy for high-probability (versus low-probability) features was in line with our hypothesis in the preregistration, providing evidence that conscious access is enhanced for targets with high-probability (versus low-probability) features. We found no systematic difference between the staircased presentation durations of high-probability feature trials (2.04, SD = 0.90) and low-probability feature trials (2.03, SD = 0.87), t(55) = 1.03, p = .308, Cohen’s d = 0.14, 95% CI = [–0.01, 0.03], thus excluding the possibility that the observed accuracy difference was due to unbalanced presentation durations between the two conditions. Importantly, we measured localization accuracy given a fixed stimulus-presentation time, so the differential localization performances cannot be accounted for by decisional biases and postdetection effects but must reflect that—within the given timeframe—more information about stimulus location was extracted in one condition than in the other.
The higher localization sensitivity for probable versus improbable features was replicated in a supplemental experiment (see Supplemental Material 2) using a different blinding technique (the detection-discrimination dissociation paradigm, or DDD paradigm; Stein & Peelen, 2021) that uses backward masking instead of interocular suppression. Moreover, this experiment further showed that the ability to discriminate between high- and low-probability feature stimuli did not influence the magnitude of the statistical-learning effects. Therefore, the advantage of probable features in localization sensitivity did not depend on the awareness of the dimension that carried the regularities (i.e., the feature). These findings further confirm that the statistical-learning effects we observed reflect differences in conscious access, rather than differences in postperceptual or decisional processes.
For further evidence of the robustness of the results, see Supplemental Material 1, which depicts the effect sizes as a function of included participants for Experiments 1 through 3 and the supplemental experiment.
Intertrial priming effects
As in Experiments 1 and 2, an exploratory test was conducted to examine potential intertrial priming effects. Results showed that for high-probability trials, there was no difference in accuracy between trials preceded by a high-probability trial (fraction correct = 0.75, SD = 0.09) and those preceded by a low-probability trial (fraction correct = 0.74, SD = 0.12), t(55) = 0.86, p = .394, Cohen’s d = 0.12, 95% CI = [–0.01, 0.03]. For low-probability trials, there was no difference between trials preceded by a low-probability trial (fraction correct = 0.75, SD = 0.17) and those preceded by a high-probability trial (fraction correct = 0.72, SD = 0.12), t(55) = 1.44, p = .156, Cohen’s d = 0.19, 95% CI = [–0.01, 0.07].
The subjective-awareness ratings
As in Experiments 1 and 2, we asked participants to guess the proportion of upward and downward triangles. Results showed that out of 56 participants, there were 7 participants who correctly guessed the regularity (no preferences: 22; incorrect guesses: 27). The Pearson correlation test (p > .05 in the Shapiro-Wilk test of normality) showed that there was no significant correlation between the calculated values of subjective awareness and the accuracy difference (between high- and low-probability features) in the b-CFS task, r = .01, p = .926 (Fig. 3c).
General Discussion
In this study, three different paradigms provided converging evidence for the influence of statistical learning on conscious access. In reaction-time-based b-CFS experiments, targets broke through interocular suppression faster when they appeared at probable locations or contained probable features. In a preregistered accuracy-based b-CFS experiment, we successfully replicated the advantage of probable features in breaking through suppression, while excluding potential contributions of decisional and postperceptual factors (i.e., factors influencing the behavioral responses that arise after conscious detection of the stimulus). Using a different masking technique (i.e., backward masking) and experimental task (the DDD paradigm), the supplemental experiment provided the additional evidence that the perceptual advantage for probable objects was not correlated to the conscious access of the dimension that carried the regularities; this further demonstrates the generalizability of our findings across perceptual blinding techniques (i.e., backward masking). Taken together, our study goes beyond existing work in showing that statistical learning alters the priority of sensory inputs for conscious access, facilitating the entry of probable targets into consciousness relative to improbable targets.
This finding provides a new perspective on how statistical learning underlies many basic and higher-order cognitive functions. In numerous studies, statistical learning was suggested as a powerful mechanism for determining what information is prioritized in perception (e.g., the effect of statistical learning on visual attention; Chun & Jiang, 1998; Wang & Theeuwes, 2018). We show that this selectivity may begin at the early stages of visual processing, even before sensory inputs enter conscious experience. Because consciousness is of limited capacity, a large amount of information in the environment competes for conscious access; the regulation of conscious access by statistical learning may enable the processing of potentially relevant objects and events for higher-order cognitive functions (e.g., conscious attention, Wang & Theeuwes, 2018; language, Saffran et al., 1996; learning and memory, Brady et al., 2009 and Umemoto et al., 2010; inference, Dotsch et al., 2017). For instance, the superior recognition of regular stimuli (e.g., regular shapes and meaningful words) by humans from early life onward (e.g., Saffran et al., 1996) may be (partially) attributed to their faster entry into consciousness. For cognitive mechanisms that highly rely on conscious resources (such as working memory; Brady et al., 2009; Umemoto et al., 2010), statistical learning may increase the amount of conscious resources allocated, thus allowing for deeper processing and information integration.
We propose that there are several possible (and not necessarily mutually exclusive) mechanisms by which statistical learning regulates the priority of stimuli before they are consciously perceived. First, the influence of statistical learning on conscious access might be mediated by spatial or featural attention toward stimuli before they are consciously perceived. Although it could be reasoned that top-down attention is involved in this process, earlier work has shown that top-down attention is not sufficient for enhancing conscious access (Gayet et al., 2020; Stein & Peelen, 2021). For example, Gayet et al. (2020) showed that, even though observers were explicitly incited to attend a feature—that is, given a strong attentional template—for a concurrent search task, stimuli containing the attended feature were not released from suppression faster. A more likely hypothesis, then, is that the invisible stimuli triggered bottom-up attention and directed the allocation of attention toward certain locations (Hsieh et al., 2011; Jiang et al., 2007) and features (e.g., Sun et al., 2016). Second, statistical learning may evoke implicit anticipation of upcoming visual events (Turk-Browne et al., 2010), which has been shown to modulate the detection of interocularly suppressed stimuli (Denison et al., 2011, 2016; Pinto et al., 2015). Predicting the upcoming image from a sequence of images, for instance, can facilitate detection of expected images during interocular suppression (Denison et al., 2011), and the anticipation of visual stimuli can evoke feature-specific activity patterns in early visual cortex (e.g., Kok et al., 2014). In this way, statistical learning may preactivate stimulus-specific representations in primary visual cortex, thereby lowering the effective threshold for probable stimuli to breach conscious access. Another possibility is that, through neuroplasticity (Conway, 2020), statistical learning increases the sensitivity or selectivity of V1 neurons to specific (e.g., probable) stimuli or patterns. Future studies should arbitrate between these possible ways in which statistical learning modulates conscious access.
The fields of statistical learning and perceptual learning have exhibited a trend toward convergence in methods and results in recent years (Fiser & Lengyel, 2019). However, previous studies found no significant impact of perceptual learning on conscious access (Mastropasqua et al., 2015; Paffen et al., 2018). For example, perceptual training on gratings with specific orientations (Mastropasqua et al., 2015) or specific motion directions (Paffen et al., 2018) did not facilitate the breakthrough of trained stimuli relative to untrained stimuli during interocular suppression. Furthermore, it has been shown that perceptual learning must be associated with threat to elicit feature-specific effects on conscious access (Stein, 2019), whereas we show that statistical learning, without being associated with threat, can also exert an impact on conscious access. The comparison between our findings and the existing literature therefore suggests that statistical learning and perceptual learning may exert distinct influences on conscious access. Relatedly, behavioral studies (e.g., Trevethan et al., 2012) and neurological studies (e.g., He & Gan, 2022) reliably showed evidence that perceptual learning can improve cortical blindness to a certain extent, increasing detectability of stimuli that were previously not consciously perceived. Considering that statistical learning appears to have a more pronounced impact than perceptual learning in modulating conscious access of masked stimuli, this opens up the question of whether statistical learning would also be more effective than perceptual learning in the treatment of patients with cortical blindness.
The statistical-learning effects shown here appear to be reminiscent of the preferential conscious access of familiar stimuli (human faces, language, etc.) over unfamiliar stimuli (Gobbini et al., 2013; Jiang et al., 2007; Ramon & Gobbini, 2018). In these studies, however, familiarity typically relates to explicit and lifelong learning processes, which have been shown to influence perception differently from the short-term extraction of statistical regularities, even within the same context (e.g., Dogge et al., 2019). At the same time, the advantage of probable stimuli over improbable stimuli may, at first glance, seem to contradict the advantages of surprising stimuli, such as faster breakthrough of images of minority members over majority members (Kardosh et al., 2022) or the faster breakthrough of scenes containing unexpected objects (Mudrik et al., 2011). However, the preferential conscious access of surprising stimuli is still up for debate because of the mixed results in the literature (e.g., Biderman & Mudrik, 2018). We suggest that contextual factors might influence the directionality of these effects (i.e., the advantage for probable over improbable objects, or vice versa). For example, it is possible that statistically unlikely objects are prioritized over statistically likely objects when the statistically unlikely objects are more informative (Denison et al., 2016). Statistically likely objects may also be deprioritized because they are distracting (e.g., Wang & Theeuwes, 2018). Thus, although we have shown that statistical learning increases conscious access to high-probability stimuli, it is possible that, under certain circumstances or task requirements, statistical learning might differently modulate conscious access (e.g., facilitating conscious access to improbable stimuli). This possibility should be further investigated.
In conclusion, we have consistently shown in four experiments that the visual system rapidly extracts statistical regularities from streams of sensory input to promote the selection of information for conscious processing. Given that conscious resources are scarce, and that access to consciousness is a prerequisite for a myriad of cognitive functions, our findings provide a mechanism for how statistical learning subserves these cognitive functions.
Supplemental Material
sj-docx-1-pss-10.1177_09567976241263344 – Supplemental material for Statistical Learning Facilitates Access to Awareness
Supplemental material, sj-docx-1-pss-10.1177_09567976241263344 for Statistical Learning Facilitates Access to Awareness by Luzi Xu, Chris L. E. Paffen, Stefan Van der Stigchel and Surya Gayet in Psychological Science
Footnotes
Transparency
Action Editor: Krishnankutty Sathian
Editor: Patricia J. Bauer
Author Contributions
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.