
Abstract
According to accounts of neural reuse and embodied cognition, higher-level cognitive abilities recycle evolutionarily ancient mechanisms for perception and action. Here, building on these accounts, we investigate whether creativity builds on our capacity to forage in space (“creativity as strategic foraging”). We report systematic connections between specific forms of creative thinking—divergent and convergent—and corresponding strategies for searching in space. U.S. American adults completed two tasks designed to measure creativity. Before each creativity trial, participants completed an unrelated search of a city map. Between subjects, we manipulated the search pattern, with some participants seeking multiple, dispersed spatial locations and others repeatedly converging on the same location. Participants who searched divergently in space were better at divergent thinking but worse at convergent thinking; this pattern reversed for participants who had converged repeatedly on a single location. These results demonstrate a targeted link between foraging and creativity, thus advancing our understanding of the origins and mechanisms of high-level cognition.
Humans have an exceptional capacity for creative thought. Consider the engineer who invents a novel device by combining older parts or the poet who describes a mundane experience in multiple unexpected ways. Whether creating new inventions or writing poetry, humans appear singular among animals in their ability to think flexibly and innovatively. But this presents a mystery: How did we evolve such extraordinary creativity?
The evolutionary origins of human creativity have implications for its proximal mechanisms—that is, for how creativity is accomplished, today, in our minds. One proposal offers an especially parsimonious account: Our creative prowess might be rooted in the evolutionarily ancient machinery of spatial search. On this account, creativity is an “exaptation” (Gould & Vrba, 1982) that recycles older abilities to think about, explore, and interact with space (Hills, 2006; Hills et al., 2008, 2010; Rhodes & Turvey, 2007; Todd & Hills, 2020; cf. James, 1890, on mental life as a series of “flights and perches”). This account suggests that our ancestors’ mundane creative activities, such as searching for food, facilitated adaptation to changing environments. Over time, our capacity to navigate space in flexible ways may have been exapted for more abstract forms of movement, such as foraging in semantic memory or moving metaphorically through a space of abstract ideas (Hills, 2006; Todd & Hills, 2020).
The human capacity for high-level creativity, therefore, may involve “recycling” (Dehaene & Cohen, 2007) or “reusing” (Anderson, 2010) more ancient neural circuitry that evolved for navigating space. We shall refer to this account as creativity as strategic foraging. On this account, mechanisms for creativity evolved from our capacity for complex, continuous, embodied interactions with the environment—searching for food, water, shelter—but now have been exapted for more abstract forms of mental foraging. Creative searches for words, ideas, or solutions engage processes akin to physically searching the external world (Hills, 2019; Hills et al., 2008, 2010; Malaie et al., 2023; Montez et al., 2015; Rhodes & Turvey, 2007; Todd & Hills, 2020). For example, Hills et al. (2008) investigated one prediction of this account: that strategies used to search through actual space might carry over to more abstract searches. They had participants search through a virtual environment in which resources were either spatially clustered or spatially diffuse. They then completed an abstract search task where they had to form meaningful words from sets of scrambled letters. After searching in spatial clusters, participants spent more time with each cluster of letters before moving on to the next set of letters, as if they were adopting the same strategy in the abstract space of words that they had recently used successfully to search literally through space.
But creativity is not a monolith. Some creative tasks involve generating many viable ideas (e.g., Beaty et al., 2015; Campbell, 1960; Kenett et al., 2018; Mednick, 1962). Others require finding a single response that fulfills a set of constraints (e.g., Duncker, 1945; Gick & Holyoak, 1980). Divergent and convergent thinking, two facets of creativity, may involve different approaches to spatial foraging, with divergent thinking relying on wide exploration and convergent thinking on finding a single “location” in a space of ideas. If true, then divergent thinking might benefit from priming a diverse exploration of space, whereas convergent thinking might benefit from priming a targeted search for a single destination. Support for this proposal comes from studies of verbal fluency and spatial search, which have found evidence for shared mechanisms involved in searching both for words in the mental lexicon and for goals in physical space (Hills, 2006; Hills et al., 2008, 2010, 2012, 2015; Troyer et al., 1997). Similarly, a recent study found that drawing divergent lines to multiple targets (compared with repeating a line to the same target) improved divergent thinking, as measured by a word stem completion task (Malaie et al., 2023).
Various lines of research, therefore, support a link between spatial search and creativity, and there are reasons to suspect that the relationship is quite targeted, with specific forms of creativity (divergent vs. convergent) recycling specific strategies for foraging through space. Here, we systematically test this proposal. To investigate a targeted link between spatial search and creative thought, we had participants search through space in one of two ways, followed by a test of either divergent or convergent thinking (Fig. 1). In the spatial search task, participants sought targets on a map of Venice; between subjects, we manipulated whether the targets were dispersed throughout the map (Fig. 1, top row) or clustered together at the same location (Fig. 1, bottom row). Participants then completed a task that required either convergent thinking (compound remote associate test; Fig. 1, right column) or divergent thinking (word stem completion; Fig. 1, left column). Creativity as strategic foraging predicts a systematic and targeted relation between spatial search and subsequent creativity: Participants who searched through space in a dispersed way should perform relatively better on subsequent tests of divergent thinking, whereas those who searched for a single target should perform better on subsequent tests of convergent thinking.
Statement of Relevance
Humans are incredibly creative. Where did this ability come from? One possibility is that creative thinking is not an entirely new mental ability but builds on older abilities, like the ability to search through space (e.g., for food). Generating many different ideas (divergent creativity) might recycle mental processes involved in searching through space for multiple locations; finding one perfect idea (convergent creativity) might be like finding a single, targeted location in space. Here we report that people who searched a city map for different locations were then better at generating multiple different solutions on an unrelated test of creativity; people who searched the city for a single, targeted location were then better at finding the single best solution to a puzzle. This connection between spatial search and mental creativity is consistent with accounts of human creativity that treat it as an extension of abilities that we share with other species. These findings suggest new ways to unlock creative potential by harnessing connections among movement, search, and insight.
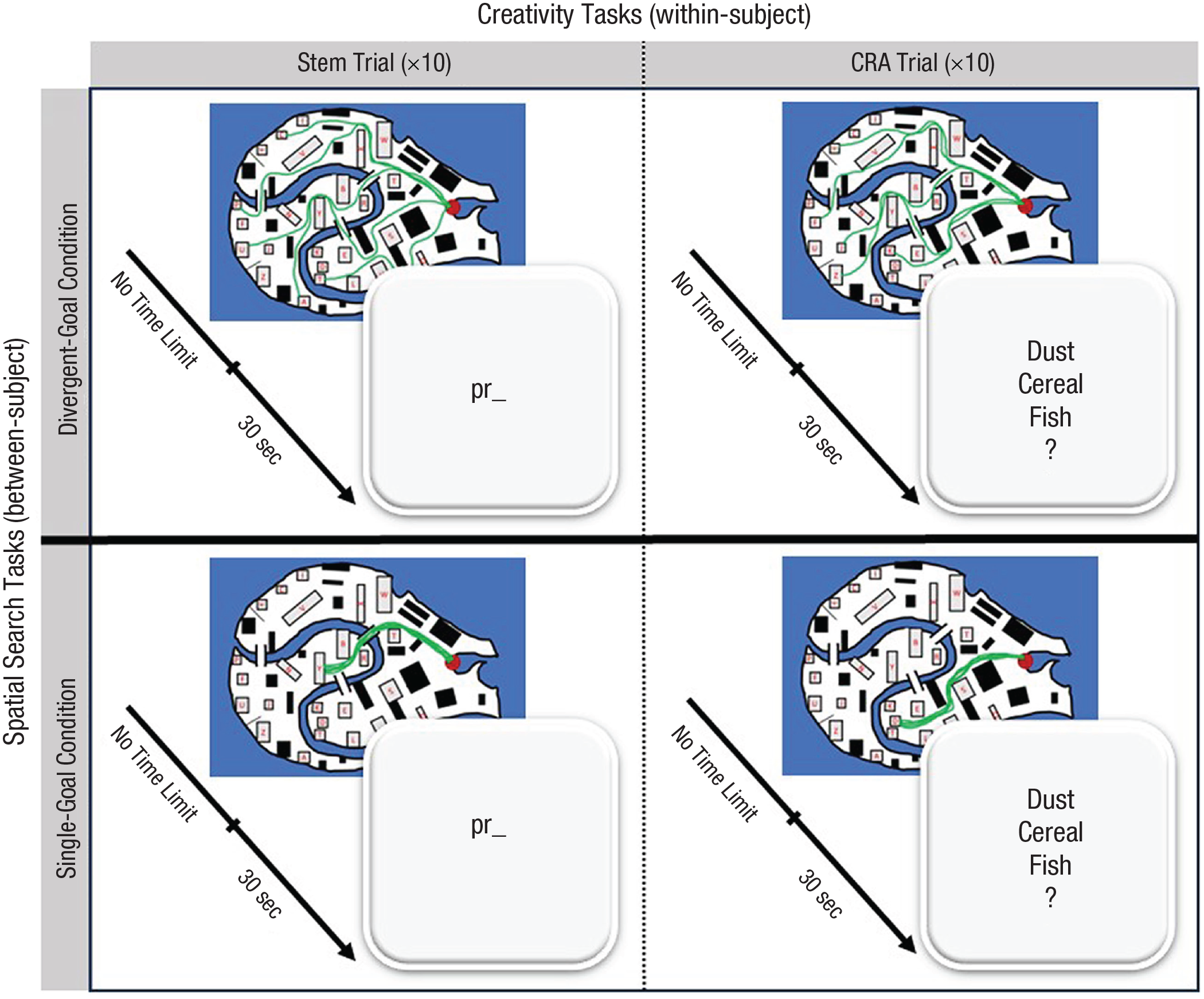
Design and procedure. On each trial (illustrated in each of the four panels), participants first completed a spatial search task, followed by a test of creativity. Between subjects, we manipulated whether participants searched for multiple divergent targets (top row) or a single repeated target (bottom row). On the spatial search task, participants had unlimited time to draw lines (green) five times from the initial location (red dot) to the target location(s). Target locations were cued by letters displayed at the top of the screen. Each spatial search was followed immediately by either a stem completion item (left column) or a compound remote associate item (right column), which lasted up to 30 s. Participants completed a block of 10 trials for each task, and task order was randomized.
Method
We report how we determined our sample size, all data exclusions, all manipulations, and all measures in the study (Simmons et al., 2012). As we noted earlier, this experiment, including all the tasks and procedures participants underwent, are publicly available for demonstration purposes at https://soranmalaie.github.io/FYP_Venice_map.
Participants
We recruited participants from Prolific, an online platform for recruiting high-quality research participants (N = 143; 72 women, 68 men, three other gender; age M = 42.37 years, SD = 12.78, range = 20–69; education ranging from less than a high school diploma to a doctoral degree). Sample size was determined ahead of data collection and was based on the sample size used in a similar study, Malaie et al. (2023), which used a sample of 63 participants. Here, to assure a well-powered design, we took advantage of our online design to double the target sample size. A post hoc power analysis confirmed that we had a power of 0.84 to detect a medium effect. To maintain homogeneity in our sample and control for potential language and cultural variations that could impact the study’s outcomes, we limited participation to native English speakers living in the United States. Participants were compensated $4.67.
Design
Participants first gave informed consent. In a mixed design, participants were assigned to one of two spatial search conditions (divergent goals vs. single goal) and completed two tests of creative thinking (for details of the procedures, see the Task Procedures section). In the divergent-goals condition, participants had to find targets distributed throughout a series of maps. Each map had five target locations, and participants had to draw five routes from a fixed origin to each of the locations (Fig. 1, top row). In the single-goal condition, participants had to draw five routes that converged on the same target location (Fig. 1. bottom row). The start location was constant across conditions, but target location(s) varied in every trial. Spatial search condition assignment was counterbalanced between participants; the two spatial search conditions did not differ significantly in age, gender, or education (all ps > .14). All participants, regardless of their spatial search condition, completed the same two tests of creativity: one requiring divergent thinking, the other requiring convergent thinking, with creativity task order randomized (Fig. 1, left vs. right column). For each creativity task, participants completed 10 trials (20 creativity items overall), with trial order randomized within each creativity task. On each trial, participants first completed the spatial search immediately followed by one creativity item.
The study was conducted online, programmed in jsPsych (de Leeuw, 2015), and hosted on GitHub (https://github.com/). Condition assignment was handled by DataPipe (pipe.jspsych.org), which also sends participant data directly to the Open Science Framework (OSF; osf.io) for archiving. After completing the tasks, participants completed an author recognition task and provided demographic information. The author recognition task is a measure of reading experience and exposure to print, which we included for a future investigation of individual differences in creative thinking. (See Supplemental Material for the full list of demographic questions.) No other measures were collected or analyzed. The study took approximately half an hour to complete (M = 36 min).
Task procedures
Spatial search task
In the spatial search task, repeated before each creativity item, participants searched a simplified map of Venice, Italy (Fig. 1, inset). Using the mouse cursor, participants had to draw five routes along roads from a fixed start location (red dot) to the target location(s) (indicated by letters). A cover story explained that, on each trial, participants were delivering five items from the start to the target location(s). We manipulated, between subjects, whether the search involved divergent routes or routes that all converged on the same location. In the divergent-goals condition (Fig. 1, top row), participants had to draw routes to five distinct target locations, thus searching widely. In the single-goal condition (Fig. 1, bottom row), participants had to draw five routes that converged repeatedly on the same target location. The target locations(s), which differed on each trial, were indicated by letters that appeared at the top of the screen. After drawing all five routes on a map, participants then completed one item of either the divergent thinking or convergent thinking tasks.
Divergent thinking task
To measure divergent thinking, we used a stem completion task (Graf & Mandler, 1984; Malaie et al., 2023; Warrington & Weiskrantz, 1970, 1974). On each trial (N = 10), participants were presented with a two-letter stem (e.g., “PR”) and had 30 s to type as many words that started with that stem (e.g., “proud,” “present,” “principle”). Participants were informed that they were prohibited from using proper nouns (e.g., “Prague”) and from using simple suffixes to generate multiple derivates of the same word (e.g., “principles,” “principled”). Participants were informed that incorrect words would not result in negative points, so if they were uncertain about whether a word was acceptable, they should submit it and continue searching for other words. Because this task requires participants to explore their mental lexicon to generate multiple, different solutions to a single prompt, we adopted it as a measure of participants’ divergent thinking. The same items were used by Malaie et al. (2023) to investigate divergent thinking.
Convergent thinking task
To measure convergent thinking, we used a compound remote associate task (CRA; Bowden & Jung-Beeman, 2003). On each CRA trial (N = 10), participants were presented with three words and had to identify a fourth word that formed a common two-word phrase with each of the given words. For instance, if presented with the words “dust,” “cereal,” and “fish,” the correct solution would be “bowl” because it forms the phrases “dust bowl,” “cereal bowl,” and “fishbowl.” Participants had up to 30 s to type their solution. We selected items of moderate difficulty from Bowden and Jung-Beeman’s (2003) norming study. Because this task requires participants to converge on a single solution that fulfills multiple constraints, we adopted it as a measure of participants’ convergent thinking.
Analysis
Visual inspection of the routes drawn in the spatial search task confirmed that participants had followed the instructions. On the stem completion task, some participants were unable to enter responses because of browser incompatibility and were removed prior to analysis (23 out of 143 total participants). To calculate the number of valid responses on each stem completion trial, we removed responses that did not start with the designated two-letter stem. We then removed responses that were not English words, in two stages: First, we used the qdap package in R to automatically identify nonwords; second, responses flagged as nonwords were inspected visually for infrequent words that were not in the default dictionary or responses that were obvious misspellings of common words. (For instance, the default dictionary in qdap does not include “zoopathology” or “abhorrent.”) To remove word repetitions and simple suffix-based variations, we used the snowball package in R to extract the root of each word and kept only unique responses. 1
On the CRA, no responses or participants were removed. Responses were considered correct if they matched the standard response supplied by Bowden and Jung-Beeman (2003).
All analyses used Bayesian mixed-effects models, implemented using the brms package in R. Word stem completion performance was analyzed using a Bayesian linear mixed-effects model of the number of words generated on each trial. CRA performance was analyzed using a Bayesian logistic mixed-effects model of trial-level accuracy. Both of these models consisted of a fixed intercept, a fixed effect of spatial search condition (single goal vs. divergent goals), and random intercepts for participants and items. Combined performance on both tasks was analyzed using a Bayesian linear regression of task-level performance (z-scored). This model consisted of a fixed intercept, fixed effects of spatial search condition (single goal vs. divergent goals), task (stem completion vs. CRA), and their interaction and random intercepts for participants. All models used noninformative priors. Model convergence was confirmed by potential scale reduction factor (R-hat) values of 1 for all parameters. Model specifications using R syntax are available in the Supplemental Material. And R code is on OSF: https://osf.io/bc8yg/?view_only=ee89f79308a944c090725742172abdcf.
Results
Stem completion task
The results are summarized in Table 1. As predicted, engaging in divergent spatial search caused a significant improvement in subsequent word generation. Compared with participants in the single-goal condition (Msingle = 5.272 words/trial), participants in the divergent-goals condition generated nearly one additional word on every trial (Mdivergent = 6.056 words/trial). A Bayesian linear mixed-effects model confirmed that divergent-goal search improved performance on the stem completion task (effect of divergent goals vs. single goal, b = 0.81, 95% Bayesian credible interval [CI] = [0.19, 1.45]; Fig. 2a). This pattern of results was found consistently across items (Fig. 2b), where performance on each item was better for participants in the divergent-goals than in the single-goal condition. Thus, engaging in diverse spatial search (divergent-goals search) improved performance on a test of divergent thinking.
Bayesian Linear Mixed-Effects Model of Stem Completion Task
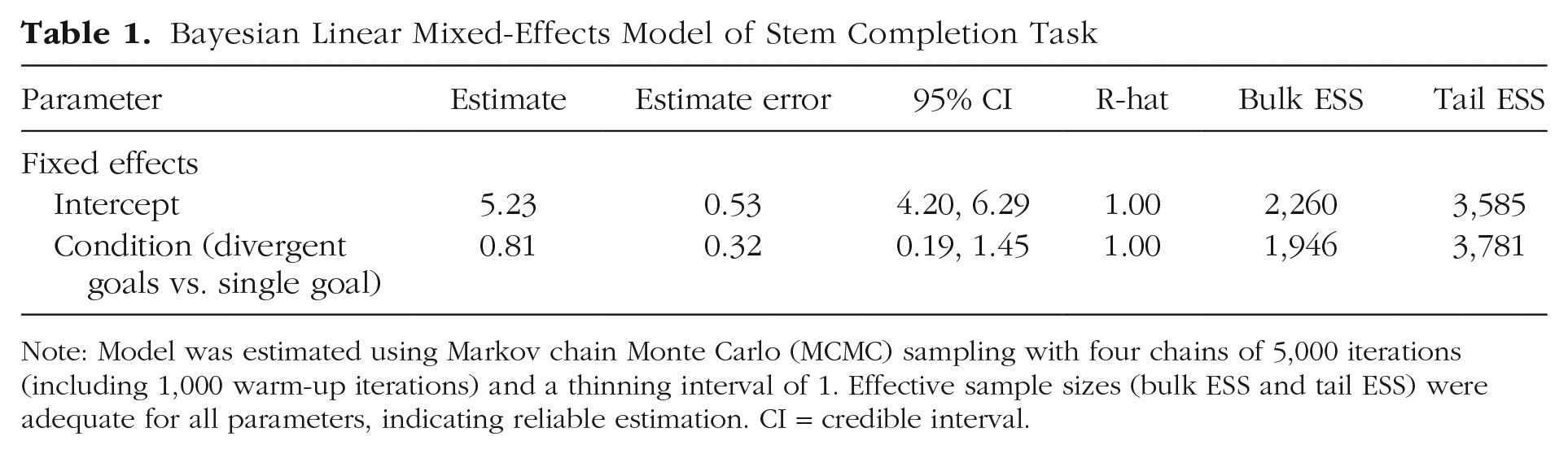
Note: Model was estimated using Markov chain Monte Carlo (MCMC) sampling with four chains of 5,000 iterations (including 1,000 warm-up iterations) and a thinning interval of 1. Effective sample sizes (bulk ESS and tail ESS) were adequate for all parameters, indicating reliable estimation. CI = credible interval.
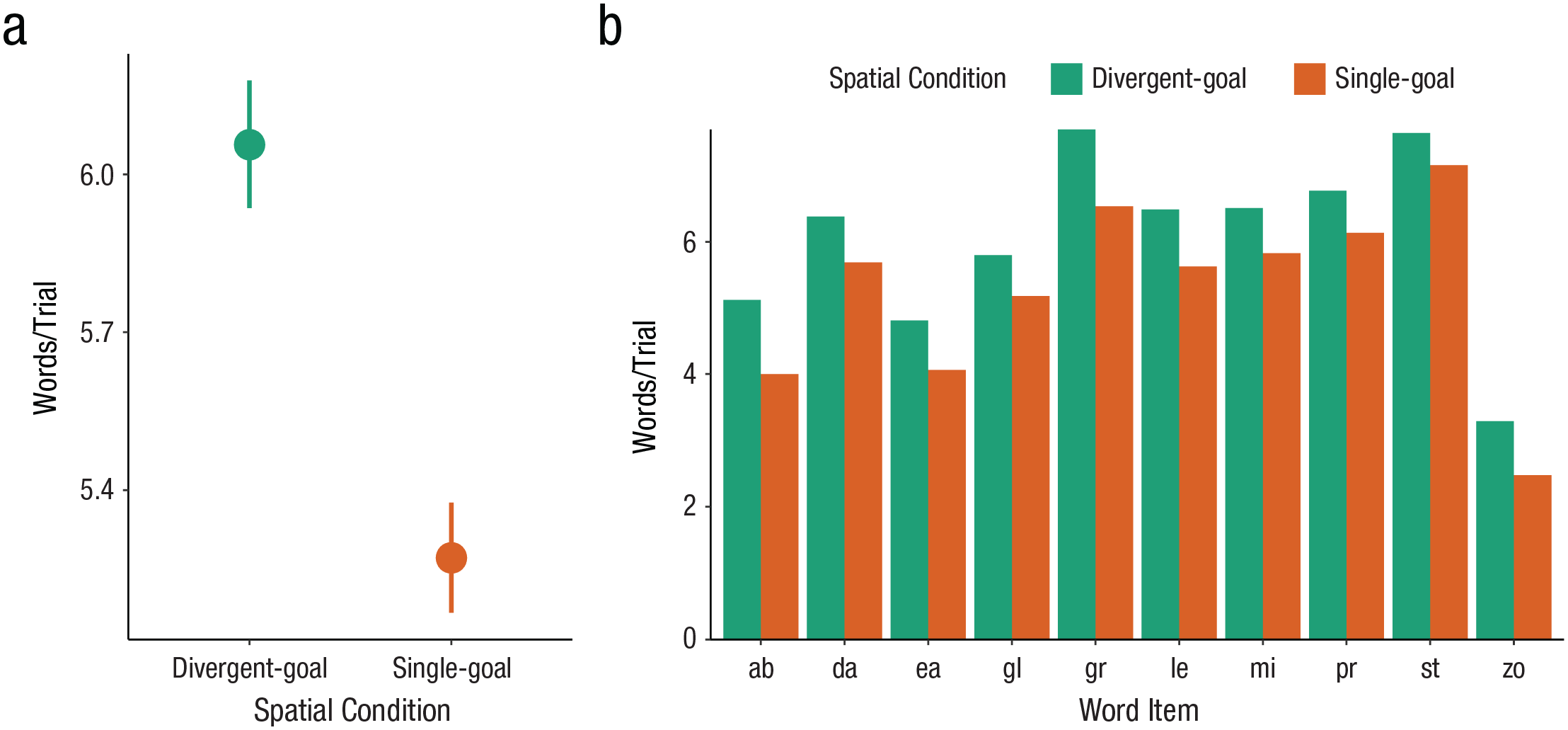
Performance on the stem completion task. (a) Participants who had completed a diverse spatial search generated significantly more words. (Points show means; error bars show standard errors.) (b) The effect of spatial search was consistent across items. Bars show the mean number of words generated for each item, split by spatial search condition. After completing a diverse spatial search (green), participants generated significantly more words for all times.
Compound remote associate task
The results are summarized in Table 2. Unlike for the stem completion task, but as predicted, performance on the CRA was significantly better for participants who had engaged in spatial searches that converged on a single location (Msingle = 65% correct) than those who had engaged in divergent search (Mdivergent = 55% correct). A Bayesian logistic mixed-effects model confirmed that single-goal search improved performance on the CRA (effect of single goal vs. divergent goals, b = 0.58, 95% Bayesian CI = [0.08, 1.10]; Fig. 3a). This pattern of results was found consistently across items (Fig. 3b), whereby performance on each item was better for participants in the single-goal than in the divergent-goals condition. Thus, engaging in spatial search that converged on a single location (single-goal search) improved performance on a test of convergent thinking.
Bayesian Logistic Mixed-Effects Model of Compound Remote Associate Task
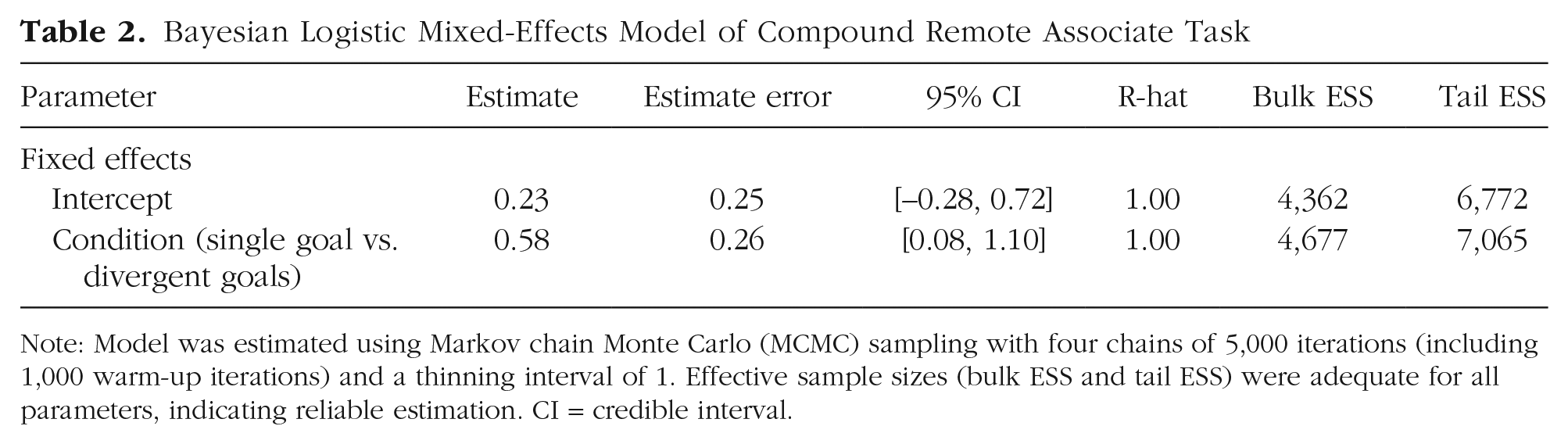
Note: Model was estimated using Markov chain Monte Carlo (MCMC) sampling with four chains of 5,000 iterations (including 1,000 warm-up iterations) and a thinning interval of 1. Effective sample sizes (bulk ESS and tail ESS) were adequate for all parameters, indicating reliable estimation. CI = credible interval.
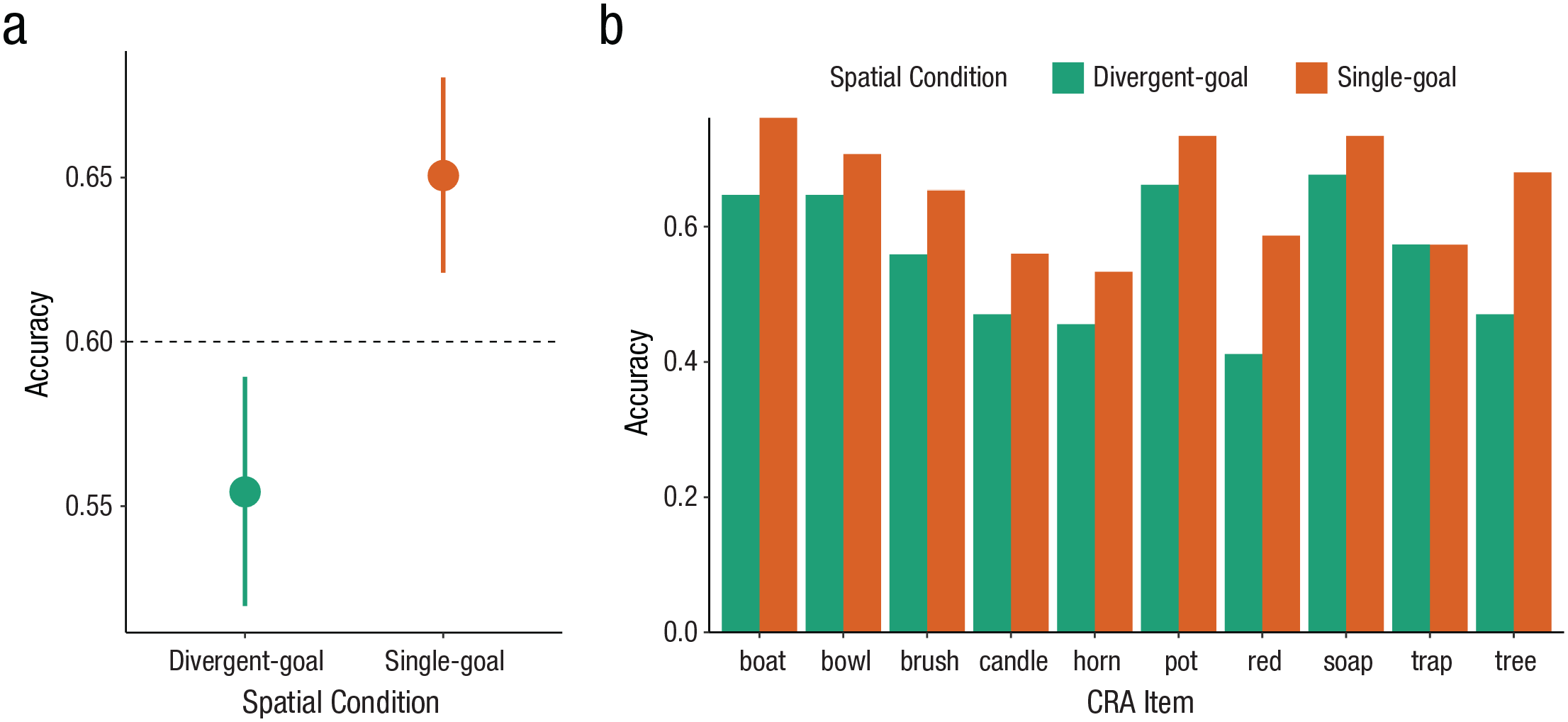
Performance on the compound remote associates task. (a) Participants who had completed a single-goal spatial search were significantly better at converging on the correct response. The dotted line shows “baseline” performance in the norming study by Bowden and Jung-Beeman (2003). (Points show mean accuracy; error bars show standard errors.) (b) The effect of spatial search was consistent across items. Bars show the mean accuracy for each item, split by spatial search condition. After completing a single-goal spatial search (orange), participants were significantly better at finding the correct response.
Combined data
To confirm the targeted benefits of spatial search for different kinds of creative thinking, we analyzed performance on both tasks in a single model. To compare across tasks, we z-scored participants’ aggregate performance on each task; positive (negative) values thus indicate that a participant performed better (worse) than average on that task. Task performance was analyzed with a Bayesian linear mixed-effects model with fixed effects of spatial search condition (divergent goals vs. single goal), task (stem completion vs. CRA), and their interaction. Results are summarized in Table 3. As predicted, the two different types of spatial search had selective and targeted impacts on creative thinking, as captured by the interaction between spatial search condition and task (b = 0.71, 95% Bayesian CI = [0.37, 1.05]); Fig. 4). After searching for multiple spatial locations and drawing divergent spatial routes, participants were systematically better at divergent thinking but worse at convergent thinking. Conversely, after repeatedly drawing spatial routes that converged on a single spatial location, participants were systematically better at convergent thinking but worse at divergent thinking.
Bayesian Linear Mixed-Effects Model of Performance on Both Tasks
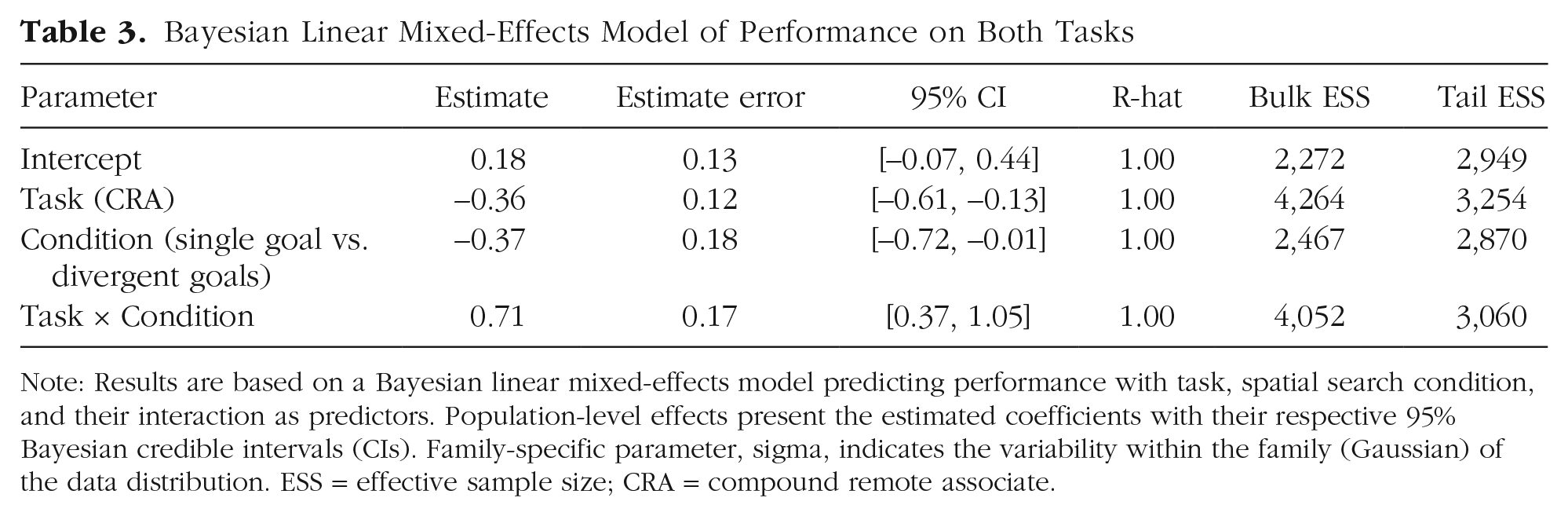
Note: Results are based on a Bayesian linear mixed-effects model predicting performance with task, spatial search condition, and their interaction as predictors. Population-level effects present the estimated coefficients with their respective 95% Bayesian credible intervals (CIs). Family-specific parameter, sigma, indicates the variability within the family (Gaussian) of the data distribution. ESS = effective sample size; CRA = compound remote associate.
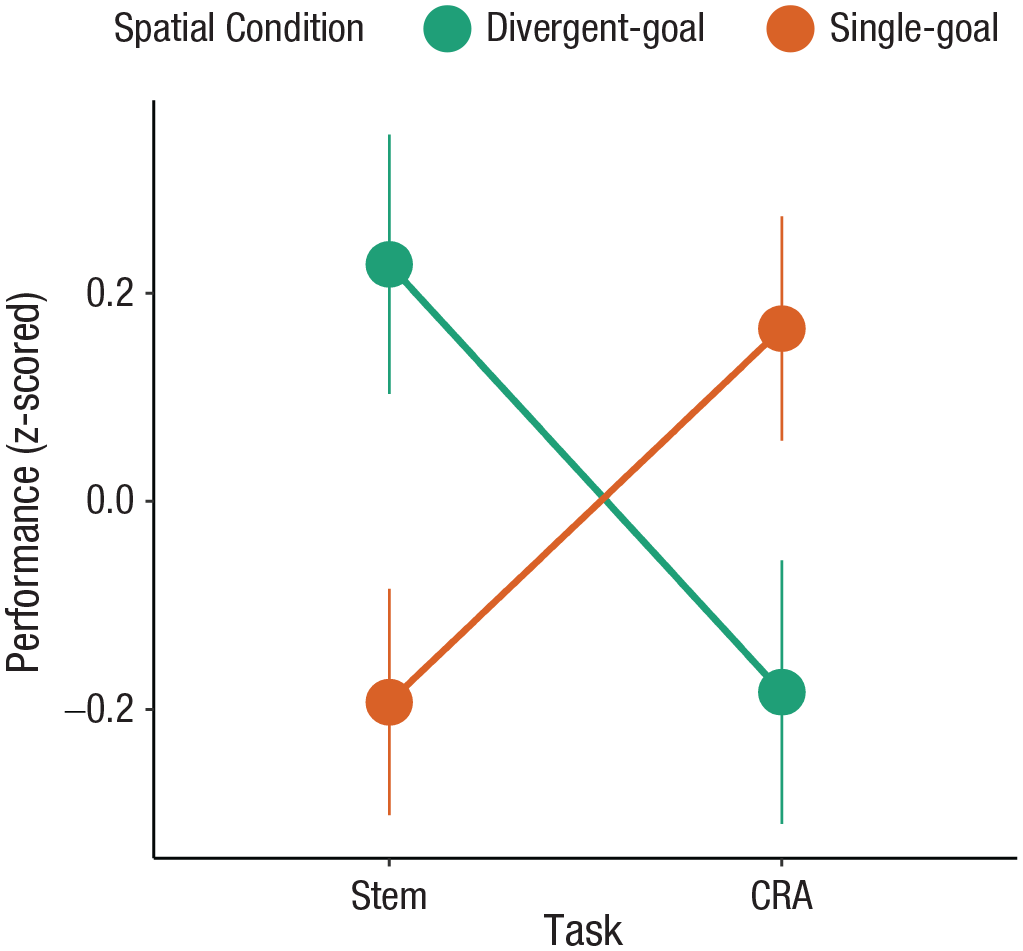
Combined performance across both the stem completion and compound remote associates (CRA) tasks. Performance was z-scored within each task. Participants who had searched for multiple, dispersed spatial targets were better at divergent thinking (stem completion) but worse at convergent thinking (CRA), compared with participants who had searched for a single spatial target.
Discussion
We explored the relationship between creative thinking and evolutionarily ancient spatial search abilities. Specifically, we investigated whether performance on divergent and convergent creative thinking tasks was systematically linked to associated forms of spatial search, as proposed by the creativity-as-strategic-foraging account. Consistent with our predictions, participants who had searched divergently in space performed better on a test of divergent creativity but worse on a test of convergent creativity, whereas participants who had searched repeatedly for the same spatial location showed the opposite pattern. In short, divergent and convergent spatial search had targeted impacts on divergent and convergent creativity. These findings are consistent with the idea that high-level cognitive processes, including creativity, draw on evolutionarily ancient spatial foraging abilities, repurposing them for abstract tasks (Hills, 2006, 2019; Hills et al., 2008, 2010; Rhodes & Turvey, 2007; Todd & Hills, 2020). Notably, these effects were elicited through interactions with a city map, a common daily-life activity, highlighting the ecologically valid connection between concrete spatial search and abstract creative thinking.
Our findings align with existing proposals that the cognitive mechanisms underlying foraging in physical space are recycled when we “forage in mind” for more words, ideas, or memories (Hills, 2006; Todd & Hills, 2020). If ancient mental procedures involved in physical foraging are repurposed and exapted for cognitive processes, then the creative search for novel solutions in seemingly abstract tasks may rely on the same fundamental mechanisms used for searching in physical space. The exact nature of the shared mechanisms, however, remains open to future work. One possibility is that neural circuits that evolved to support spatial memory and search are conserved across tasks that involve some kind of foraging and thus reused to support computationally similar processes during nonspatial search. Indeed, such regions have recently been implicated in semantic and phonetic foraging (Lundin et al., 2023; Ovando-Tellez et al., 2022) and creative thinking (Bottini & Doeller, 2020). Indeed, the existence of common neural circuits that are deployed both for spatial foraging and for creativity offers a parsimonious explanation of our finding that spatial search primed creative thinking, because the activation of a particular strategy for spatial foraging would carry over to subsequent creative thinking. This mechanistic account, grounded in recent empirical findings, echoes William James’s (1890) much older speculation that foraging for food and exploring the mind rely on shared mechanisms.
More generally, these results support a view of cognition as embodied, grounded in mechanisms for perception and action (e.g., Barsalou, 1999; Lakoff & Johnson, 1980). Traditional amodal approaches fail to capture the intricate interplay between cognitive functions and spatiomotor activities (e.g., Fodor, 1983; Mahon & Caramazza, 2008). In contrast, our findings suggest a tight link between spatial search strategies and high-level cognitive processes. This interaction emphasizes the connection between external physical experiences and internal mental processes.
Similarly, these results resonate with a treatment of cognition as a continuous trajectory through a high-dimensional state space of ideas (Spivey, 2008). In this view, the stream of thought is a journey through this state space, with some locations and concepts easily accessible and others more difficult. Succeeding at a particular task—such as word stem completion or CRA—will require a particular kind of trajectory through state space. When the task requires a cognitive trajectory that visits many disparate locations, then priming widespread exploration may help; when the task requires a trajectory that finds a specific, hard-to-reach region of state space, then encouraging a strategy of targeted exploitation may help. The priming results reported here may reflect the influence of spatial search on the continuous trajectory of thought.
Creativity as strategic foraging shares features with conceptual metaphor theory (Lakoff & Johnson, 1980). Conceptual metaphor theory argues that our conceptualization of abstract domains, such as time and number, is built on a foundation of spatial metaphors (Casasanto & Bottini, 2014; Lakoff & Johnson, 1980; Marghetis & Núñez, 2013; Winter et al., 2015). In English and other languages, creative thought is described in spatial terms—“thinking outside the box,” “thinking fluidly,” “moving forward” with an idea—and some studies have tested whether these figures of speech capture how people think about creativity (Kuo & Yeh, 2016; Leung et al., 2012; Oppezzo & Schwartz, 2014; Slepian & Ambady, 2012). Note, however, that conceptual metaphor theory and creativity as strategic foraging have different explanatory targets. Conceptual metaphor theory is an account of how we conceptualize creativity—how we think about creativity, how we make sense of it as an attribute or activity. Creativity as strategic foraging, by contrast, is an account of how we do creativity. For example, English speakers conceive of loving relationships, metaphorically, as journeys (e.g., “they are at a crossroads in their relationship”; Lakoff & Johnson, 1980). But the act of actually loving somebody is something else entirely; it involves processes of desire, attachment, physiological attunement, and more. Thus, our conception of love is distinct from the actual process of loving. Conceiving is not the same as doing. Seen in this light, evidence that we conceptualize creativity in spatial terms is orthogonal to how creativity is actually done. Creativity as strategic foraging is an account of the doing of creative thought.
The current results help explain inconsistencies in the literature on creativity and walking. Several studies have found that walking enhances creativity, especially when walking freely along an unconstrained route (e.g., Kuo & Yeh, 2016; Leung et al., 2012; Murali & Händel, 2022), but others have found that walking can inhibit creativity (Malinin, 2019; Oppezzo & Schwartz, 2014). Creativity as strategic foraging offers an explanation: Different kinds of creativity recycle different foraging strategies. Free walking, therefore, should help divergent thinking while potentially hindering convergent creativity, which is indeed the pattern that has been reported in the literature (Malinin, 2019).
The design of the current study invites future work to clarify the connection between spatial foraging and creative thinking. For one, the study reported here was conducted online, and online studies have known limitations, such as limited control of the testing environment, a higher risk of distractions, and the absence of physical intervention by researchers during the experiment (for a recent review, see Gagné & Franzen, 2023); on the other hand, our online design allowed us to reach a more diverse population than is common in lab-based studies, as evidenced by the wide range of ages (20 to 69 years old) and educational attainments (from participants with no high school diploma to those with a doctoral degree) in our sample. In addition, the priming design used here cannot distinguish between two versions of the creativity-as-strategic-foraging account. In one account, discussed earlier, creativity redeploys the same neural machinery as spatial foraging, similar to accounts of embodied or grounded cognition (e.g., Barsalou, 1999; Dehaene & Cohen, 2007; Pezzulo et al., 2011). Alternatively, the neural systems that support spatial foraging and creativity may be distinct but tightly coupled. Future investigations are required to clarify the exact implementational details of the connection between spatial foraging and creative thought. For instance, bidirectional interactions between spatial search and creative thought would suggest the operation of an integrated neural system for foraging in space and mind. Recent neuroimaging studies lend credence to such an integrated system. For instance, the brain’s spatial circuits have been implicated in creative thinking (Bottini & Doeller, 2020), and shifts in semantic foraging during tests of verbal fluency are associated with activity in hippocampal and cerebellar regions otherwise known to support spatial memory and search (Lundin et al., 2023). Future work should clarify the exact processing pathways that are responsible for divergent and convergent search both in literal space and in creative worlds of ideas and imagination.
Conclusion
In an online study of convergent and divergent creativity, we demonstrated that different strategies for searching in space had significant and targeted impacts on different kinds of creativity. The findings support the notion that evolutionarily ancient sensorimotor processes are repurposed and redeployed for the abstract tasks faced by modern humans. By understanding and harnessing the influence of spatial characteristics, we can potentially enhance cognitive processes related to creativity and problem-solving. Future research in this area could further explore the underlying mechanisms and potential applications of spatial search strategies in various domains.
Supplemental Material
sj-docx-1-pss-10.1177_09567976241245695 – Supplemental material for Divergent and Convergent Creativity Are Different Kinds of Foraging
Supplemental material, sj-docx-1-pss-10.1177_09567976241245695 for Divergent and Convergent Creativity Are Different Kinds of Foraging by Soran Malaie, Michael J. Spivey and Tyler Marghetis in Psychological Science
Footnotes
Acknowledgements
We would like to express our gratitude to Dr. John A. Bargh for his invaluable suggestion to use the map of Venice as a testing ground for the ideas presented in this article. We also extend our heartfelt appreciation to the members of Marghetis Lab and Spivey Lab for their valuable feedback and insights.
Transparency
Action Editor: Vladimir Sloutsky
Editor: Patricia J. Bauer
Author Contributions
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.