
Abstract
Over the past decade, evolutionary psychologists have proposed that many moral stances function to promote self-interests. At the same time, behavioral geneticists have demonstrated that many moral stances have genetic bases. We integrated these perspectives by examining how moral condemnation of recreational drug use relates to sexual strategy (i.e., being more vs. less open to sex outside of a committed relationship) in a sample of Finnish twins and siblings (N = 8,118). Twin modeling suggested that genetic factors accounted for 53%, 46%, and 41% of the variance in drug condemnation, sociosexuality, and sexual-disgust sensitivity, respectively. Further, approximately 75% of the phenotypic covariance between drug condemnation and sexual strategy was accounted for by genes, and there was substantial overlap in the genetic effects underlying both drug condemnation and sexual strategy (rg = .41). Results are consistent with the proposal that some moral sentiments are calibrated to promote strategic sexual interests, which arise partially via genetic factors.
Some moral sentiments (e.g., stealing and murder are wrong) are almost universally endorsed, whereas others underlie substantial disagreement within societies. One example of the latter has contributed to fortunes spent on legal enforcement and to the incarceration of tens of thousands of individuals: that recreational drug use is immoral (Piaggio & Vidwans, 2019). At first blush, condemnation of other individuals’ drug use can seem puzzling because one individual’s use of drugs has little to no impact on an observer’s outcomes. Recent work aimed at solving this puzzle suggests that much of the variability in drug condemnation overlaps with orientations toward a more versus less committed sexual strategy (Kurzban et al., 2010; Quintelier et al., 2013), presumably because of associations between drug use and casual sex. For example, adolescent recreational drug use covaries with earlier sexual debut, more sexual partners, and engaging in unprotected sex (e.g., Lowry et al., 1994), and recreational drugs are often used at events where people seek out uncommitted sex (Garcia et al., 2012; Ross et al., 2003).
People who invest heavily in long-term, committed relationships have more to lose in social ecologies that afford opportunities for mate switching or so-called extrapair copulations, and people who pursue short-term mates benefit from such ecologies. Hence, individuals who are more commitment oriented should support rules that shift the social ecology toward high commitment in relationships. Individuals who are less commitment oriented should resist such rules because they would be the targets of condemnation and punishment simply for engaging in their preferred sexual behaviors (Weeden & Kurzban, 2017). Accordingly, findings indicate that sexual strategy (i.e., being more vs. less open to sex outside of a committed relationship) relates to moral views toward contraception and abortion (Weeden & Kurzban, 2014), pornography (Simpson & Gangestad, 1991), and same-sex marriage (Pinsof & Haselton, 2016). A first test of this hypothesis applied to moral condemnation of drug use found that, in U.S. college student and Amazon Mechanical Turk samples, sociosexuality, sexual-disgust sensitivity, and opinions on sexual policies were uniquely and more strongly related to condemnation of recreational drug use than Big Five personality traits, pathogen- and moral-disgust sensitivity, broader political orientation, and opinions on policies not related to sexuality (Kurzban et al., 2010). These findings have been replicated in The Netherlands, Belgium, and Japan (Quintelier et al., 2013).
The Strategic-Interest Hypothesis
Work suggesting that condemnation of drug use is contingent on one’s orientation toward a more versus less commitment-oriented sexual strategy is in line with recently developed evolutionary perspectives on moral condemnation (Petersen, 2015; Petersen & Laustsen, 2019; Pinsof & Haselton, 2016; Weeden & Kurzban, 2014, 2017), which suggest that one’s support for and opposition to moral rules serve fitness-promoting functions. For instance, recent findings suggest that physical formidability influences views on resource redistribution, showing that physically stronger men tend to approve of inequitable divisions of resources (Petersen & Laustsen, 2019)—a rule that tends to favor more physically imposing individuals across several species. Other evidence suggests that transient states such as hunger can nudge moral views in self-serving directions. For example, one study found that experimentally induced low blood glucose levels (indicating hunger) increased people’s support for social welfare (Aarøe & Petersen, 2013). But where do these strategic interests—such as those affecting drug condemnation—emerge from?
Genetic and Environmental Underpinnings of Moral Judgments
Much existing work is based on the assumption that moral sentiments toward recreational drugs are guided by broader political ideologies, which are socially transmitted from parents to offspring (Sears & Brown, 2013). Seemingly consistent with this assumption, findings have shown that people resemble their parents in terms of political orientation and religiosity (Jennings et al., 2009; Leonard et al., 2013) and attitudes toward drugs (Kandel, 2001). However, decades of behavioral-genetics research—using both twin studies and adoption studies—show that within-family similarities are largely undergirded by shared genes rather than shared environments, which presumably would include social transmission from parents (Plomin et al., 2016). And while meta-analyses suggest that some of the strongest shared environmental effects found in twin studies involve social values, such as those relating to societal attitudes, religion, and spirituality (Polderman et al., 2015), genetic effects tend to be substantially larger than shared environmental effects on both broader ideological orientation and stances on specific sociopolitical issues. For instance, in twin samples from Australia and the United States, genes account for 36% to 70% of the variance in attitudes toward abortion, gay rights, and homosexuality, whereas shared environmental effects account for 5% to 18% of the variance (Eaves & Hatemi, 2008; Verweij et al., 2008). In a similar vein, much of the within-family similarities in views toward recreational drug use might be underpinned by genetic effects.
Statement of Relevance
In the past five decades, the war on drugs has borne immense financial, social, and political costs and consequences. Therefore, understanding the factors that shape moral views toward recreational drug use is crucial. Recent research indicates that sexual strategy (i.e., being more vs. less open to sex outside of a committed relationship) underlies much of the variability in moral condemnation of recreational drugs, presumably because of the association between drug use and casual sex. We sought to estimate the extent to which genetic, shared environmental, and nonshared environmental effects underlie the covariance between condemnation of recreational drugs and sexual strategy. Results suggest that this covariance is largely accounted for by genetic effects and that much of the same genes influence both sexual strategy and condemnation of recreational drugs. These findings suggest that variation in sexual strategy underlies heritability in a key aspect of modern culture wars: condemnation of recreational drugs.
The Reactive Heritability of Moral Views
In aiming to integrate abundant (and seemingly fitness-relevant) variation in personality with a putatively universal evolved psychology, Tooby and Cosmides (1990) proposed that much of this variation reflects reactive heritability. This perspective suggests that clusters of behavioral traits are attuned to heritable aspects of an individual’s phenotype, resulting in (reactive) heritability of those behavioral trait clusters. For instance, physically stronger individuals are better able to impose costs on others and, thus, benefit more from aggressiveness than smaller individuals. As a result, a heritable physical feature (e.g., size) would underlie heritability in a behavioral feature (e.g., aggressiveness; Sell et al., 2009; Tooby & Cosmides, 1990). In a similar vein, such universal psychological mechanisms could drive people to adopt the political, coalitional, and moral positions that would bring advantages to themselves, producing divergent stances contingent on the demographic and lifestyle features of the individual—for instance, driving physically stronger men to adopt more accepting views of economic inequality (Petersen & Laustsen, 2019) and more favorable views on aggressive military action (Sell et al., 2009). Similarly, heritability in drug condemnation might stem from moral views being calibrated to one’s sexual strategy, given that the latter has consequences for how those moral rules affect the individual in question. Existing research suggests that sexual strategies are indeed heritable (Bailey et al., 2000; Sherlock et al., 2016). If moral condemnation of drug use reflects reactive heritability, the heritable components of drug condemnation are likely to overlap with the heritable components of sexual strategy.
The Current Study
Past work on drug condemnation and sexual strategy has relied exclusively on phenotypic correlations—that is, correlations between instruments measuring condemnation of recreational drugs and sexual strategy. Such work has not informed the degree to which the relation between the two reflects genetic or environmental covariance. Using a genetically informed sample of more than 5,000 twins and their siblings, we sought to assess the extent to which genetic, shared environmental, and nonshared environmental effects underlie condemnation of recreational drugs and sexual strategies. We further aimed to test the extent to which the relation between the two variables reflects similar genetic, shared environmental, or nonshared environmental effects.
Method
The instruments used in the study (both those administered to participants and English translations), data, and analysis scripts are accessible on OSF (https://osf.io/kmau4).
Participants
An invitation letter for a Web survey was sent to 33,211 twins and the siblings of twins identified in the Central Population Registry in Finland. A maximum of two reminder letters spaced 2 to 3 weeks apart were sent to participants who did not respond to the initial letter. Data collection began in November 2018 and concluded in January 2019. In total, 9,319 people completed at least part of the survey and gave consent for the use of their data for scientific purposes. We included only participants who completed all three measures of interest for this study (condemnation of recreational drug use, sociosexuality, and sexual-disgust sensitivity). The resulting sample included 8,118 individuals, of which 5,993 were twins and 2,125 were siblings of twins. This sample consisted of 1,296 complete twin pairs: 126 monozygotic men, 384 monozygotic women, 115 dizygotic men, 337 dizygotic women, and 334 dizygotic opposite-sex twins. Twins were between the ages of 18 and 45 years (M = 29.28 years, SD = 7.75), and the siblings were between 18 and 58 years (M = 32.0 years, SD = 8.36). Further details regarding data collection, zygosity estimation, and translation procedures are provided by Tybur et al. (2020).
Measures
Participants completed measures of drug condemnation and sexual strategy (sociosexuality and sexual-disgust sensitivity) used by Kurzban et al. (2010) and Quintelier et al. (2013). All instruments were presented in Finnish.
Drug condemnation
Participants read scenarios describing situations in which people used recreational drugs at parties or with friends (MDMA, marijuana, cocaine, or speed; scenarios were taken from the study by Quintelier et al., 2013, who had adapted them from the study by Kurzban et al., 2010). They then indicated their agreement with statements such as “Using MDMA in this way is morally wrong” and “Using MDMA in this way should be legally permitted” (reverse coded) for each of the scenarios. They also indicated their agreement with two additional sentiments: “People who use recreational drugs are dirty,” and “It is ok to obtain drugs for the purpose of making you feel good” (reverse coded). As in the study by Quintelier et al. (2013), all of the items were rated on a scale from 1 (strongly disagree) to 7 (strongly agree). A mean score of the items was computed (α = .93).
Sexual strategy
Following Kurzban et al. (2010) and Quintelier et al. (2013), we used measures of sociosexuality (Simpson & Gangestad, 1991) and sexual-disgust sensitivity (Tybur et al., 2009) to assess sexual strategy. Participants completed the revised Sociosexual Orientation Inventory (SOI-R; Penke & Asendorpf, 2008), which includes subscales for sociosexual attitudes (e.g., “Sex without love is OK”), behavior (e.g., “With how many different partners have you had sexual intercourse without having an interest in a long-term committed relationship with this person?” from 1 = 0 to 9 = 20 or more), and desire (e.g., “In everyday life, how often do you have spontaneous fantasies about having sex with someone you have just met?” from 1 = never to 9 = at least once a day). The SOI-R contains nine questions in total. We used the mean score of the full scale in the analyses (α = .81). Participants also completed the Three-Domain Disgust Scale (Tybur et al., 2009), which includes pathogen-, moral-, and sexual-disgust sensitivity subscales, the latter of which corresponds with sexual strategy (Tybur et al., 2015). This subscale consists of items such as “Watching a pornographic video” and “Performing oral sex,” which participants rated on a scale from 0 (not at all disgusting) to 6 (extremely disgusting; α = .82).
Covariates
We also collected measures of personality, religiosity, social dominance orientation, and right-wing authoritarianism. Personality was measured using the HEXACO-100 (Lee & Ashton, 2018), which assesses honesty-humility (α = .82), emotionality (α = .82), extraversion (α = .90), agreeableness (α = .82), conscientiousness (α = .79), and openness to experience (α = .82). The items were rated on a scale from 1 (strongly disagree) to 5 (strongly agree). Religiosity was measured with the Intrinsic Religiosity subscale of the Duke University Religion Index (Koenig & Büssing, 2010), which contains items such as “I try hard to carry my religion over into all other dealings in life,” rated from 1 (completely disagree) to 7 (completely agree; α = .93). Social dominance orientation was measured with the Social Dominance Orientation scale (SDO7; Ho et al., 2015), which contains items such as “An ideal society requires some groups to be on top and others to be on the bottom,” rated on a scale from 1 (strongly oppose) to 7 (strongly favor). We used the mean score of the full scale in the analyses (α = .92). Right-wing authoritarianism was measured with the Aggression-Submission-Conventionalism scale (Dunwoody & Funke, 2016), which contains items such as “People emphasize tradition too much” (conventionalism, reverse coded; α = .80), “People in positions of authority generally tell the truth” (authoritarian submission; α = .79), and “Strong punishments are necessary in order to send a message” (authoritarian aggression; α = .76). Items were rated on a scale from 1 (strongly oppose) to 7 (strongly favor).
Analytic approach
We employed the classical twin study design, which assesses monozygotic and dizygotic twins (and, here, siblings) to estimate the extent to which genetic effects, shared environmental effects, or unique (nonshared) environmental effects underlie trait variance. Because monozygotic twins share approximately 100% of their genes, whereas dizygotic twins and siblings share, on average, 50% of their segregating genes, higher correlations between monozygotic twins than between dizygotic twins and between twins and siblings indicate greater genetic influences on that trait. The classical twin design can also be used to partition the covariance between two variables into genetic and environmental components and to estimate the degree to which genetic and environmental influences overlap in explaining a trait—that is, the extent to which the same genetic or environmental factors affect both drug condemnation and sexual strategy.
Phenotypic analyses
First, we calculated phenotypic correlations between drug condemnation, sociosexuality, and sexual-disgust sensitivity. Second, we tested whether drug condemnation uniquely related to sociosexuality and sexual-disgust sensitivity by partialing out the effect of personality, religiosity, social dominance orientation, and the three Aggression-Submission-Conventionalism subscales (conventionalism, authoritarian aggression, and authoritarian submission). Correlations between all variables are provided in Table S1 in the Supplemental Material available online.
Genetic analyses
Genetic analyses were conducted with the OpenMx package (Version 2.19.5; Boker et al., 2011) in the R programming environment (Version 4.0.3; R Core Team, 2020) using the full-information maximum likelihood option, which allows for analyses of all available data and responses from twin singletons contributing to only the estimation of means and variances. We first fitted a saturated model, in which variable means were free to vary across men and women, and twin correlations were free to vary (within zygosity) across monozygotic twin pairs, dizygotic twin pairs, twin and sibling pairs, and opposite-sex twin pairs. We then constrained parameters and tested changes in model fit using likelihood-ratio tests, where the negative log-likelihood (−2LL) of the more constrained nested model is subtracted from the −2LL of the saturated model, and the difference is tested with a χ² test (α < .01). If model fit did not differ across nested models, we retained the more parsimonious model (i.e., the model with fewer degrees of freedom; for the model comparisons, see Table S2 in the Supplemental Material). Using structural equation modeling, we can partition observed phenotypic variance into additive genetic (A), nonadditive genetic (D), shared environmental (C), and nonshared environmental (E) components. Similarly, observed phenotypic covariance between variables can be partitioned into these components.
Results
Phenotypic analyses
Correlations were consistent with those found in past studies (Kurzban et al., 2010; Quintelier et al., 2013). Specifically, condemnation of recreational drugs was related to both sociosexuality and sexual-disgust sensitivity (rs = −.32 and .34, respectively), which were themselves strongly related (r = −.54, p < .001). Fisher r-to-z transformations indicated that the effect sizes for the relation between drug condemnation and sexual strategy were no different from those observed in Belgian, Dutch, and Japanese samples (Quintelier et al., 2013), but they were smaller than those observed in the United States (Kurzban et al., 2010; for comparison of the effect sizes, see Figs. S1 and S2 in the Supplemental Material). These relations remained when analyses controlled for the six HEXACO dimensions, religiosity, social dominance orientation, and the three facets of right-wing authoritarianism (sociosexuality: partial r = −.22, p < .001; sexual-disgust sensitivity: partial r = .23, p < .001; see Tables S3 and S4 in the Supplemental Material).
Preliminary genetic analyses
Correlations between dizygotic twins did not differ from correlations between twins and siblings—drug condemnation: χ2(1) = 2.93, p = .09; sociosexuality: χ2(1) = 5.15, p = .02; sexual-disgust sensitivity: χ2(1) = 0.89, p = .34. We therefore modeled genetic effects of .5 for both dizygotic twins and siblings. Next, we tested for sex differences in the means and genetic and environmental influences on drug condemnation, sociosexuality, and sexual-disgust sensitivity. Relative to men, women were more condemning of recreational drug use (d = 0.50). They were also more sociosexually restricted (d = 0.52) and more sensitive to sexual disgust (d = 1.31; for means and sex-specific twin correlations, see Tables 1 and 2). Twin correlations did not differ between same-sex male and female twin pairs (quantitative sex differences), suggesting no sex differences in the contribution of genetic and environmental influences on any of the variables. Further, twin correlations did not differ between the same-sex dizygotic and opposite-sex twins (qualitative sex differences), suggesting no sex differences in the genetic or shared environmental effects influencing drug condemnation, sociosexuality, and sexual-disgust sensitivity. On the basis of these results and following the aforementioned criteria, we fitted a model in which the genetic (A), shared environmental (C), and nonshared environmental (E) influences were constrained to be equal across the sexes, but mean scores were allowed to vary between the sexes.
Mean Scores for Men and Women and Correlations Within Monozygotic (MZ), Dizygotic (DZ), and Dizygotic Opposite-Sex (DOS) Twins and Siblings, Separately for Each Dependent Variable
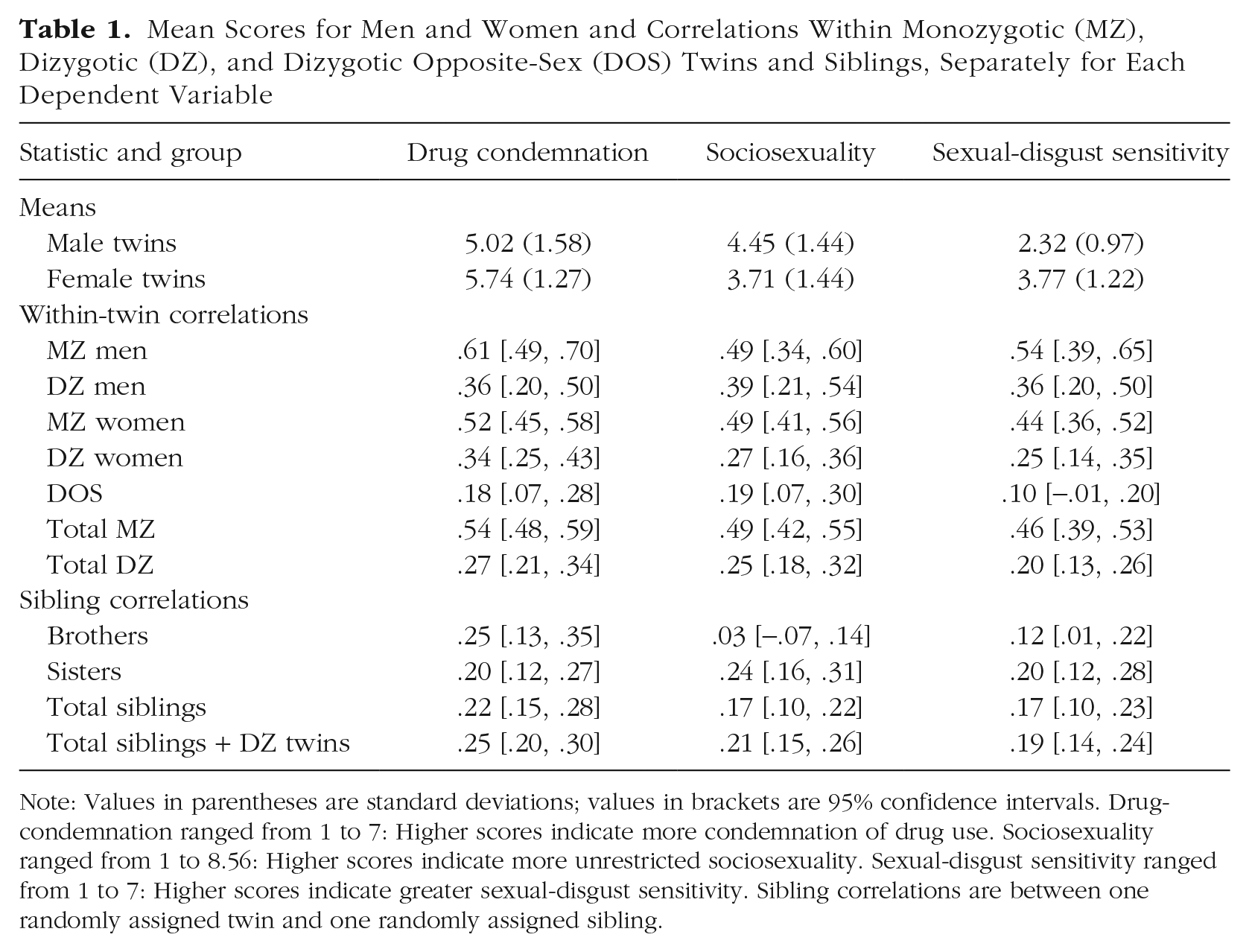
Note: Values in parentheses are standard deviations; values in brackets are 95% confidence intervals. Drug-condemnation ranged from 1 to 7: Higher scores indicate more condemnation of drug use. Sociosexuality ranged from 1 to 8.56: Higher scores indicate more unrestricted sociosexuality. Sexual-disgust sensitivity ranged from 1 to 7: Higher scores indicate greater sexual-disgust sensitivity. Sibling correlations are between one randomly assigned twin and one randomly assigned sibling.
Cross-Twin/Cross-Trait Correlations for Drug Condemnation, Sociosexuality, and Sexual-Disgust Sensitivity
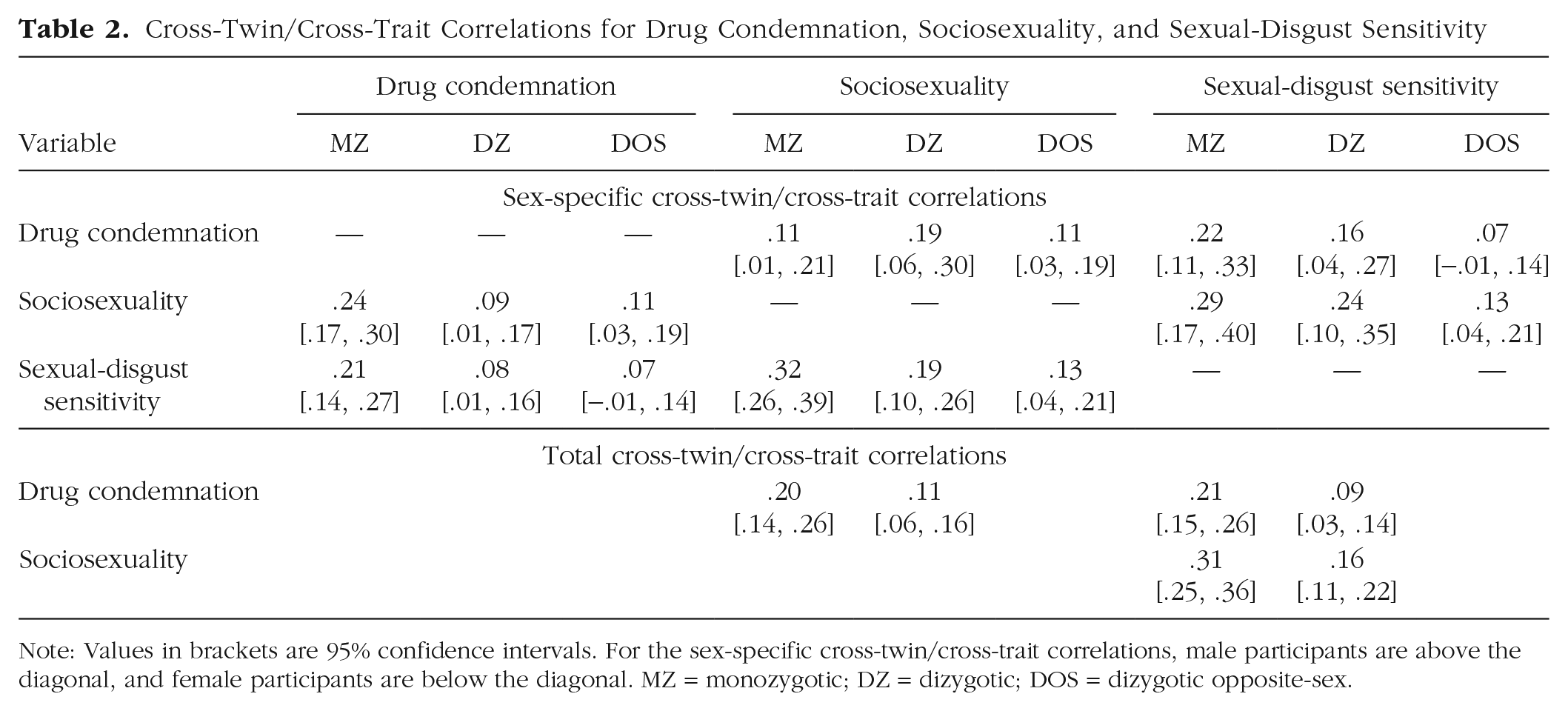
Note: Values in brackets are 95% confidence intervals. For the sex-specific cross-twin/cross-trait correlations, male participants are above the diagonal, and female participants are below the diagonal. MZ = monozygotic; DZ = dizygotic; DOS = dizygotic opposite-sex.
Genetic and environmental effects on drug condemnation, sociosexuality, sexual-disgust sensitivity, and their covariance
Correlations in monozygotic twins for drug condemnation (r = .54), sociosexuality (r = .49), and sexual-disgust sensitivity (r = .46) were approximately double their respective correlations in dizygotic twins (rs = .27, .25, and .20), suggesting no effect of the shared environment (C) on these variables. Cross-twin/cross-trait correlations for monozygotic twins were also approximately double those for dizygotic twins (Table 2), indicating a similar pattern for the genetic and environmental influences on the covariance between drug condemnation, sociosexuality, and sexual-disgust sensitivity. Further model comparison (using likelihood-ratio tests as described above) indicated no difference in model fit when the C component was dropped from the model, χ2(6) = 0.00, p = 1. Hence, we fitted an AE trivariate model (with sex-specific means estimated and shared environmental influence constrained to zero) in which the additive genetic effects and the nonshared environmental effects were estimated for each of the variables individually as well as for their covariance (Fig. 1).
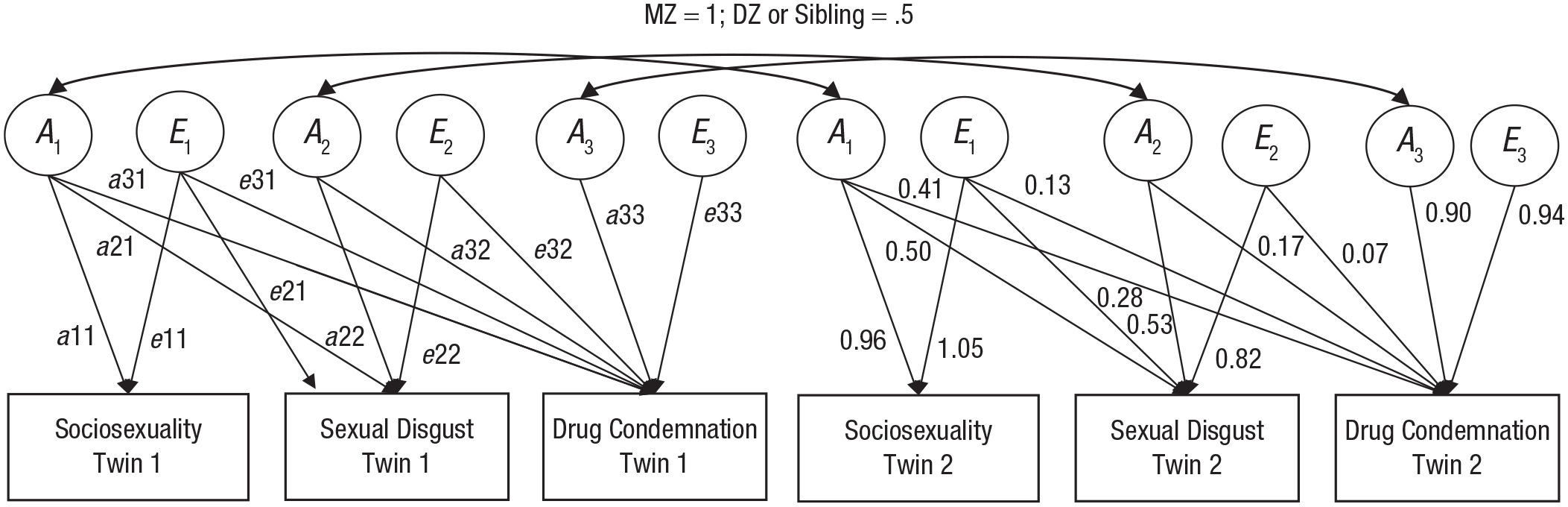
The trivariate model showing additive genetic (A) and nonshared environmental (E) effects on the three measures of interest. A1, A2, and A3 are the additive genetic effects, and E1, E2, and E3 are the unique environmental effects. Values along the single-headed arrows show factor loadings (on the left) and unstandardized path estimates from the best-fitting model (on the right). The double-headed arrows at the top of the figure indicate the constraint used in twin modeling. For monozygotic (MZ) twin pairs, correlations between A in Twins 1 and 2 are 1; for dizygotic (DZ) and sibling pairs, these correlations are .5. Twin 1 E is uncorrelated with Twin 2 E.
First, we estimated the extent to which drug condemnation, sociosexuality, and sexual-disgust sensitivity, as well as their phenotypic correlations, were accounted for by genetic, shared environmental, and nonshared environmental influences. From the trivariate AE model, we derived the following estimates: Additive genetic effects accounted for 53% of the variance in drug condemnation, 46% of the variance in sociosexuality, and 41% of the variance in sexual-disgust sensitivity, and unique environmental effects accounted for the remaining variance (Table 3). The majority (62%–76%) of the phenotypic covariance between drug condemnation, sociosexuality, and sexual-disgust sensitivity was accounted for by genetic effects (Table 3), and the remaining percentage reflected nonshared environmental effects, which includes measurement error.
Standardized Estimates of Additive Genetic (A), Shared Environmental (C), and Nonshared Environmental (E) Influences on Sociosexuality, Sexual-Disgust Sensitivity, Drug Condemnation (Univariate), and Their Covariance (Trivariate)

Note: Values in brackets are 95% confidence intervals.
Second, we estimated genetic correlations, which represent the overlap in genetic factors underlying both drug condemnation and sexual strategy—that is, the correlation between the univariate heritability estimates (A) of drug condemnation and the sexual-strategy variables (see Table 4). The genetic correlation (r) between drug condemnation and sociosexuality was .41, as was the genetic correlation between drug condemnation and sexual-disgust sensitivity, indicating that approximately 40% of the genetic underpinnings of drug condemnation overlap with the genetic underpinnings of sexual strategy. The nonshared environmental correlations were .13 and .12, respectively, indicating that slightly more than 10% of the factors that cause twins to be different from each other (including measurement error and idiosyncratic environmental effects) in drug condemnation overlap with factors that cause them to be different in sexual strategy. To account for the influence of the covariates, we also ran the trivariate model controlling for the six HEXACO dimensions, religiosity, social dominance orientation, and the three facets of right-wing authoritarianism. These analyses provided results similar as those reported here (see Table S5 in the Supplemental Material).
Genetic and Environmental Correlations for the Most Parsimonious (AE) Model

Note: A = additive genetic; E = nonshared environmental.
Discussion
Consistent with past work (Kurzban et al., 2010; Quintelier et al., 2013) and in line with evolutionary perspectives on moral condemnation (Petersen, 2015; Petersen & Laustsen, 2019; Pinsof & Haselton, 2016; Weeden & Kurzban, 2014, 2017), results indicated that high-commitment-oriented individuals are more condemning of recreational drug use, even after analyses controlled for personality, religiosity, and broader dimensions of political ideology. Further, twin modeling indicated that within-family similarities in both drug condemnation and sexual strategy reflect shared genetic rather than shared environmental effects. And, critically, there was substantial overlap in the genetic effects underlying both drug condemnation and sexual strategy (i.e., sociosexuality and sexual-disgust sensitivity). This finding is consistent with a reactive-heritability account of drug condemnation (Tooby & Cosmides, 1990).
Twin modeling detected no influence of shared environmental effects—which presumably would include parental rearing, influence of siblings, socioeconomic status, or childhood neighborhood—on either condemnation of recreational drugs or sexual strategy. These findings run counter to the idea that within-family similarities in views toward drugs and sex reflect social transmission from parents to offspring (Kandel, 2001); instead, such similarities appear to reflect shared genes. These findings add to the line of studies suggesting that both specific attitudes and broader ideology have a substantial heritable component and that the remaining variance in these variables tends to be shaped by nonshared (and, perhaps, nonsystematic; Turkheimer & Waldron, 2000) rather than shared (e.g., Eaves & Hatemi, 2008; Verweij et al., 2008) environmental factors. The absence of shared environmental effects on sexual strategy is also in line with findings of past studies, which have shown similar estimates of genetic and nonshared environmental effects on sociosexuality and sexual-disgust sensitivity (Bailey et al., 2000; Sherlock et al., 2016).
Limitations and future directions
We note a few limitations. First, the classical twin design cannot disentangle gene–environment interactions or gene–environment correlations. A gene–environment interaction, in which certain genes are activated only in certain (shared) environments, would result in an overestimate of A influences and an underestimate of C influences. Gene–environment correlations, in which people choose environments in accordance with their genetically predisposed views on sex (Westerlund et al., 2012), would result in an overestimate of the A component and an underestimate of the E component. Future studies are needed to disentangle any gene–environment interactions or gene–environment correlations that underlie sexual strategy or condemnation of recreational drugs.
Following the practice of past studies (Kurzban et al., 2010; Quintelier et al., 2013), we used measures of sociosexuality and sexual-disgust sensitivity to measure sexual strategy. Yet the sexual-disgust scale includes items that do not pertain to high- versus low-commitment sexual strategy per se (e.g., watching a pornographic video, performing oral sex). Nonetheless, sociosexuality and sexual-disgust sensitivity are highly correlated (r = −.54) and relate similarly to drug condemnation as well as to other political sentiments (Tybur et al., 2015). The present results show that 62% of the phenotypic covariance between sociosexuality and sexual-disgust sensitivity is explained by genetic sources, and the strong genetic correlation (r) of .69 suggests that both variables are shaped by the same genetic sources. Because a less commitment-oriented sexual strategy is characterized by sexual variety with multiple partners after short intervals of time, sexual disgust must presumably be downregulated to successfully pursue such a strategy (Al-Shawaf et al., 2015; O’Shea et al., 2019). However, sexual experiences may influence both sexual-disgust sensitivity and sociosexuality. Longitudinal studies that follow the same individuals from sexual debut to later adulthood can further shed light on the causality between sociosexuality and sexual-disgust sensitivity.
Concluding remarks
Over the past decade, political psychology and moral psychology have become increasingly influenced by biological perspectives. Some work in this area has described the genetic components that underlie variance in political views; other work inspired by evolutionary psychology theory has proposed that political views partially flow from psychological mechanisms for promoting rules advantageous to one’s fitness interests. Borrowing from Tooby and Cosmides’s (1990) conception of reactive heritability, we integrated these two perspectives to better understand condemnation of other individuals’ use of recreational drugs. The overlap in genetic factors underlying condemnation of recreational drug use and sexual strategy suggests that genetic variation in sexual strategy—putatively maintained via frequency-dependent selection (Bailey et al., 2000)—partially underlies a key aspect of modern culture wars: condemnation of recreational drug use. Future work can use genetically informed approaches similar to that employed here to further test the extent to which the heritable variation in our moral stances corresponds with heritable variation in other aspects of the phenotype. Such findings, combined with those reported here, can contribute to elucidating the biological factors underlying our moral and political psychology.
Supplemental Material
sj-docx-1-pss-10.1177_0956797621997350 – Supplemental material for Sex, Drugs, and Genes: Illuminating the Moral Condemnation of Recreational Drugs
Supplemental material, sj-docx-1-pss-10.1177_0956797621997350 for Sex, Drugs, and Genes: Illuminating the Moral Condemnation of Recreational Drugs by Annika K. Karinen, Laura W. Wesseldijk, Patrick Jern and Joshua M. Tybur in Psychological Science
Footnotes
Acknowledgements
We thank Kimmo J. Karinen for back-translating the questionnaire items from Finnish to English.
Transparency
Action Editor: Steven W. Gangestad
Editor: Patricia J. Bauer
Author Contributions
A. K. Karinen and J. M. Tybur designed the study. P. Jern collected the data, and L. W. Wesseldijk and A. K. Karinen analyzed the data. A. K. Karinen wrote the manuscript with input from L. W. Wesseldijk, P. Jern, and J. M. Tybur. All the authors approved the final manuscript for submission.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.