
Abstract
Although police officers are carefully selected for their high emotion-regulation abilities, excessive aggression in police officers has been reported, particularly in socially challenging situations known to elicit high state testosterone levels. Adequate regulation of emotional actions depends on the prefrontal cortex’s control over the amygdala. We investigated the effects of trait aggression and endogenous testosterone on this emotional-control neurocircuitry in 275 healthy, high-functioning police recruits using a functional MRI social-emotional task eliciting impulsive and controlled approach-and-avoidance actions. Higher levels of aggression were counteracted by increased anterior prefrontal cortex (aPFC) control over the amygdala when control over automatic emotional actions was required. Crucially, testosterone had a detrimental effect on this aggression-dependent aPFC recruitment: Police recruits with relatively high trait aggression and high state testosterone showed reduced aPFC control over the amygdala during emotion regulation. This provides a mechanistic explanation for inadequate behavioral control during socially challenging situations in otherwise well-functioning individuals.
Keywords
The ability to regulate emotional reactions is crucial for adequate, adaptive emotional behavior and largely depends on control by the prefrontal cortex over the amygdala (Etkin, Büchel, & Gross, 2015). This is particularly relevant for first responders, such as police officers, who often need to exert aggression in a controlled manner and are typically selected on the basis of strong emotion control (van der Linden, Born, Phielix, & Touw, 2013). However, excessive aggression in police officers has been reported, particularly in socially challenging situations (Atherley & Hickman, 2014), and this has exposed a need for improved insight into the relationship between aggression and emotion control mechanisms.
Studies on human aggression have, however, mainly focused on various psychiatric conditions, such as psychopathy, antisocial personality disorders, and borderline personality disorders. In these patient studies, aggression has been associated with poor prefrontal control over regions facilitating emotional responses, especially the amygdala (Coccaro, Sripada, Yanowitch, & Phan, 2011). For instance, a recent study on aggressive delinquents showed reduced activity in the anterior prefrontal cortex (aPFC), as well as reduced aPFC–amygdala functional connectivity during control over emotional actions (Volman et al., 2016). However, levels of trait aggression vary widely in the healthy population, and surprisingly little is known about effects of interindividual differences in trait aggression on emotional control in healthy, high-functioning individuals, who do not show the deficits in other cognitive or behavioral domains that most psychiatric samples do. Given that police recruits need to pass a stringent selection procedure and demonstrate well-controlled behavior overall (including functional use of anger), it is unlikely that trait aggression in this group is related to reduced prefrontal control in the same way as in the psychiatric samples. Instead, we hypothesized that the aggressive tendencies in this group are driven by increased reactivity to emotional cues within limbic structures such as the amygdala but that this increased reactivity can be offset by enhanced aPFC control in police recruits with high trait aggression.
Nevertheless, such a compensatory mechanism can be impeded by situational factors, such as high state testosterone levels. Testosterone plays an important role in social behavior and has been linked to frontal–amygdala decoupling (Mehta & Beer, 2010; Van Wingen, Ossewaarde, Bäckström, Hermans, & Fernández, 2011). For example, salivary testosterone was associated with reduced negative coupling of aPFC and amygdala in both healthy males and male delinquents (Volman et al., 2016; Volman, Toni, Verhagen, & Roelofs, 2011). Importantly, socially challenging situations are known to increase testosterone levels (Archer, 2006). It has been shown that dominant and impulsive individuals are more prone to be affected by situation-dependent testosterone levels in guiding motivational behavior (Carré et al., 2017), raising the question of whether the same goes for trait aggression. Along these lines, we also hypothesized that high testosterone may have a detrimental effect on frontal control over the amygdala in police officers with relatively high trait aggression who are otherwise characterized as having adequate emotion control.
To test these hypotheses, we measured neural control over emotional actions in a large sample of police recruits (N = 275). To do so, we used functional MRI (fMRI) while participants completed a well-established social-emotional task eliciting impulsive and controlled actions—an approach-avoidance task (Fig. 1a). Controlled actions, requiring control over automatic action tendencies, are associated with longer reaction times (RTs), stronger aPFC recruitment, and more negative aPFC–amygdala coupling (Volman et al., 2011; Volman et al., 2016). Here, we tested whether trait aggression is associated with increased recruitment of this aPFC–amygdala network during emotional control in well-functioning police recruits. Moreover, we investigated whether relatively high state levels of testosterone are associated with reduced neural emotional-control abilities in police recruits with relatively high levels of aggression.
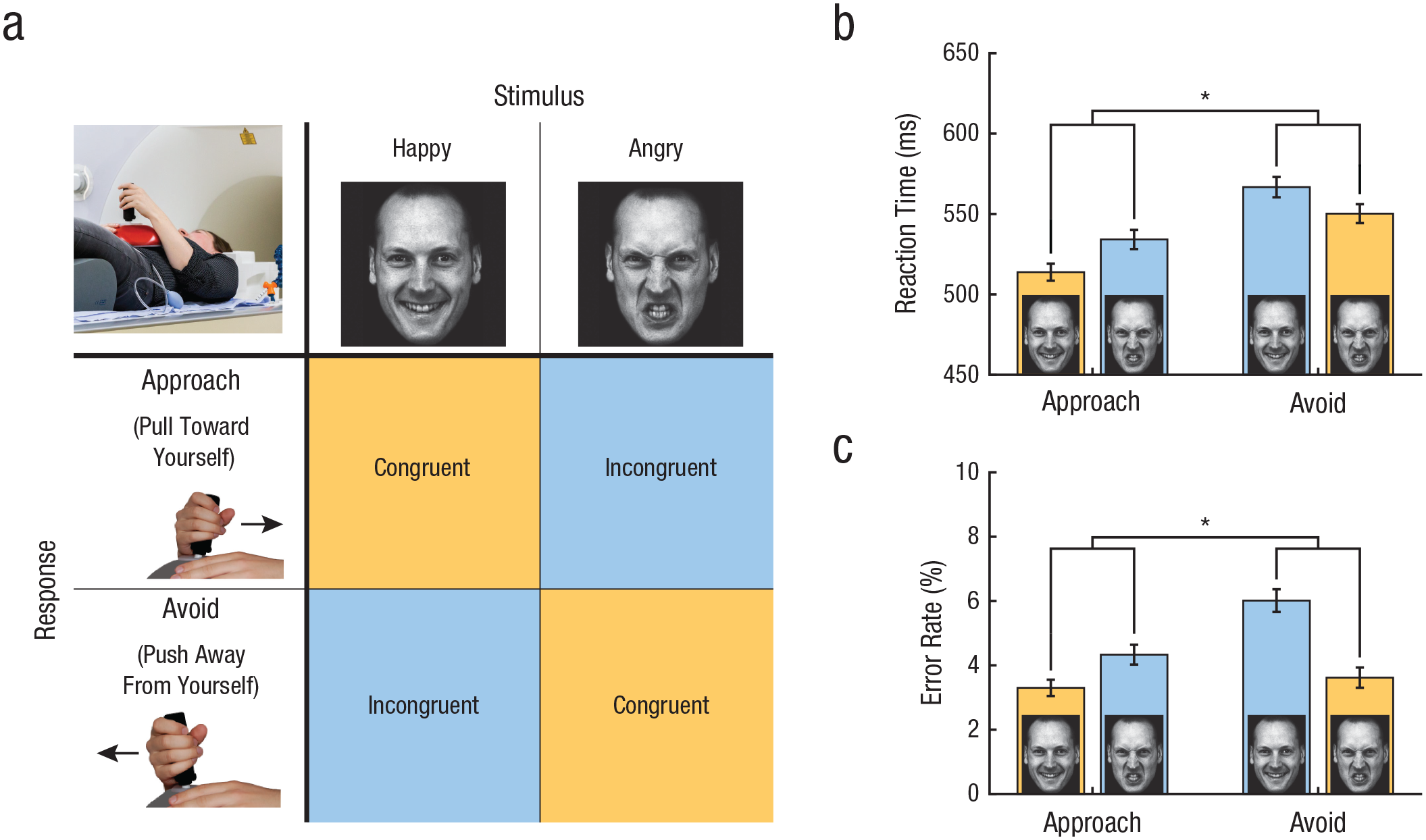
The approach-avoidance task: paradigm and results. In congruent blocks of the approach-avoidance task (a), participants approach happy faces and avoid angry faces by pulling or pushing a joystick, respectively. In incongruent blocks, participants avoid happy faces and approach angry faces (panel adapted from Volman et al., 2016; images are AM10HAS and AM10ANS from the Karolinska Directed Emotional Faces database of Lundqvist, Flykt, & Öhman, 1998). The graphs show (b) mean reaction time and (c) mean error rate on congruent and incongruent trials for each movement type. Asterisks indicate significant differences (p < .001). Error bars represent standard errors of the mean.
Method
Participants
A total of 320 police recruits from the Dutch Police Academy participated in the study. Exclusion criteria consisted of current psychiatric and neurological disorders, current or past endocrine or neurological treatment, current drug or alcohol abuse, and current use of psychotropic medication. Complete data sets (MRI data, hormone assessments, and questionnaires) were available for 285 participants. Two additional participants were excluded because of anatomical abnormalities or poor MRI quality. Three participants were excluded because of poor task performance (i.e., fewer than 10 correct trials in one or more of the four task conditions), and 5 were excluded because of extreme scores on the variables of interest. Outliers more than 3.29 SD from the mean were excluded, as recommended by Tabachnik and Fidell (2001). Consequently, 275 participants between 18 and 45 years of age (Mdn = 23 years; 209 males, 66 females) were included in the final analyses. Note that a sample size above 250 is regarded as sufficient for stable estimates of correlations with a small expected effect size typical for effects of individual differences in traits such as aggression (Schönbrodt & Perugini, 2013). The study was conducted in accordance with the principles of the Declaration of Helsinki and approved by the Independent Review Board Nijmegen, The Netherlands.
Procedure
The approach-avoidance task was part of the baseline measurement of a large prospective study assessing the role of automatic defensive responses in the development of trauma-related psychopathology in police recruits (Netherlands Trial Registry No. NTR6355). For details regarding the design of the prospective study, see Koch et al. (2017). On arrival in the lab, participants completed several questionnaires, including the Reactive Proactive Questionnaire (RPQ; Cima, Raine, Meesters, & Popma, 2013). They underwent a 7-min structural T1-weighted scan. Approximately 1 hr before the approach-avoidance task, a saliva sample was collected for hormonal assessments. In the MRI scanner, participants completed a 3-min training session to familiarize themselves with the task; they then completed the approach-avoidance task (12 min). Both the approach-avoidance task and the saliva collection were conducted in the afternoon.
Experimental task
The approach-avoidance task consisted of eight blocks of 12 trials each, with an interblock interval of 21 s to 24 s and an intertrial interval of 2 s to 4 s (Tyborowska, Volman, Smeekens, Toni, & Roelofs, 2016). At the start of each block, participants were instructed to respond to each facial expression as quickly as possible by pushing the joystick away from themselves (i.e., an avoiding movement) or pulling the joystick toward themselves (i.e., an approaching movement), depending on the valence of the facial expression. In congruent blocks, participants were required to make approaching movements in response to happy faces and avoiding movements in response to angry faces. For incongruent blocks, the instructed response mapping was reversed: Avoiding movements were used for happy faces, and approaching movements for angry faces (Fig. 1a). The four congruent and four incongruent blocks were presented alternately, and the first block type was counterbalanced across participants. The stimuli were facial expressions from 36 models (18 male), each showing two emotions (happy and angry). Stimuli presentation and acquisition of joystick positions were controlled by a PC running Presentation software (Version 16; Neurobehavioral Systems, http://www.neurobs.com). For further details about the task, see the article by Tyborowska et al. (2016).
Self-reported aggression questionnaire
Reactive and proactive aggressive symptoms were assessed with a Dutch version of the Reactive Proactive Questionnaire (RPQ; Cima et al., 2013; Raine et al., 2006), which is frequently used to assess aggression (e.g., Bobes et al., 2013). The RPQ is a 23-item questionnaire on a 3-point scale (0 = never, 1 = sometimes, 2 = always), with good test-retest reliability (all intraclass coefficients > .41) and adequate convergent validity (all rs > .16; Cima et al., 2013). RPQ scores showed high internal consistency (Cronbach’s α = .8).
Salivary measurements
Saliva was collected approximately 1 hr prior to task execution, following previous work in which we applied a comparable task design (participants completed a functional MRI approach-avoidance task) and found that testosterone levels remain stable within a 1-hr time frame (Tyborowska et al., 2016). Participants minimized physical exercise and did not smoke more than one cigarette on the day of the experiment. They also refrained from cigarettes, food, and drink (except water) for at least 1 hr before their saliva was collected. The samples were collected by passive drool of 2 ml into Salicap containers (IBL International, Hamburg, Germany), which were stored at −20 °C on the same day. Sample quality was assessed in the lab, and levels below the threshold to reliably detect testosterone values were considered undetectable. This resulted in 0% missing values in men and 2% in women. Testosterone concentration was measured using a chemiluminescence immunoassay (IBL International) with a sensitivity of 0.0025 ng/mL. Cortisol concentration was measured using a commercially available chemiluminescence immunoassay (IBL International) with a high sensitivity of 0.16 ng/mL. Additional checks showed that the testosterone levels demonstrated the expected pattern: higher in males than in females (see Table S1 in the Supplemental Material available online) and a negative correlation with age in males that was trend significant (r = −.12, p = .087) despite the limited age range of the majority of men.
Materials and apparatus
MRI data were acquired with a Siemens 3T Magnetom Prisma scanner (Siemens, Munich, Germany), using a 32-channel head coil. An ascending dual-echo echo-planar imaging sequence—37 transversal slices, repetition time (TR): 1,740 ms, echo time (TE): 11 and 25 ms, field of view (FOV): 212 mm, flip angle 90°, voxel size = 3.3 × 3.3 × 3.0 mm—was used for the acquisition of the functional scans (Poser, Versluis, Hoogduin, & Norris, 2006). Structural images were obtained using a combined magnetization-prepared rapid acquisition gradient echo and generalized autocalibrating partially parallel acquisitions sequence—192 slices, TR: 2,300 ms, TE: 3.03 ms, FOV: 256 mm, flip angle 8°, voxel size = 1.0 × 1.0 × 1.0 mm.
Behavioral analysis
Behavioral data were analyzed using MATLAB 2015 (The MathWorks, Natick, MA) and SPSS Statistics (Version 21). For each trial, a reliable measure of joystick movement onset was reconstructed from the joystick displacement measures (Tyborowska et al., 2016). Trials in which a movement in the wrong direction, or no movement at all, was made, were considered to be errors. Trial blocks with an error rate at or above chance level were excluded from both error-rate analyses and RT analyses. RT was defined as the time between the onset of the presentation of the facial expression and the onset of the movement. Error trials, trials with an extreme RT (< 100 ms or > 1,500 ms), and trials with RTs more than 3 standard deviations from the individual mean were excluded from the RT analyses. A log transformation on individual RTs was applied to obtain a normal distribution. Mean RTs were calculated for each level of the experimental factors (valence and movement), resulting in four variables (approach–happy, approach–angry, avoid–happy, avoid–angry). These mean RT scores and error rates were entered in two separate repeated measures analyses of variance (ANOVAs), with the within-subjects factors movement (approach, avoid) and valence (happy, angry). Gender, RPQ score, testosterone level, and cortisol level were included as covariates, as were all two-way and three-way interactions between hormones and aggression. The α level was set at .05.
Analysis of questionnaire and hormone levels
A log transformation was applied to the testosterone and cortisol levels to correct for a skewed distribution. Outliers more than 3.29 standard deviations from the mean for each variable within each gender group were excluded (2 participants for RPQ score, 1 participant for testosterone level, and 2 participants for cortisol level). Replicating previous studies (Dabbs, 1990; Miller & Lynam, 2006), results of independent-samples t tests indicated that men, compared with women, scored higher on aggression as measured by the RPQ, t(273) = 2.99, p = .003, and had higher salivary testosterone levels, Welch’s t(76.88) = 17.88, p < .001. Therefore, scores for these variables were standardized within each gender for further analyses. For consistency, salivary cortisol values were standardized within each gender. Trait aggression was measured using the RPQ sum scores, which is a robust measure of aggression (Brugman et al., 2017).
fMRI preprocessing
Imaging data were preprocessed and analyzed using the MATLAB toolbox for Statistical Parametric Mapping (SPM; Version 12; www.fil.ion.ucl.uk/spm). The first six volumes of each participant’s data set were discarded to allow for T1 equilibration. The two echoes were combined to form a single time series, using echo-time weighted combination. Subsequently, principal component analyses were used to filter out motion-related slice-specific noise components (Nieuwhof et al., 2017). The image time series were spatially realigned using a least-squares approach with the six rigid-body transformation parameters; they were then slice-time corrected. The T1-weighted image was spatially coregistered to the mean of the functional images. Subsequently, the T1-weighted image was segmented into gray-matter, white-matter, and cerebrospinal-fluid compartments. After normalization to standard Montreal Neurological Institute (MNI) space, the functional images were spatially smoothed using an isotropic 8-mm full-width at half-maximum Gaussian kernel. Finally, an automated-removal-of-motion-artifacts (AROMA) procedure was conducted to filter out motion-related artifacts (Pruim et al., 2015).
Single-participant analysis
Following Volman et al. (2011) and Tyborowska et al. (2016), the fMRI time series were analyzed using an event-related approach within the framework of the general linear model. The following effects were considered separately: approach–happy, approach–angry, avoid–happy, and avoid–angry. The time of stimulus presentation (onset) and the time between stimulus presentation and response (duration) were convolved with the canonical hemodynamic response function. Misses and on-screen information (i.e., instructions preceding each block and feedback messages) were modeled as separate regressors. Potentially confounding effects of residual head movement were modeled using original, squared, cubic, first-order, and second-order derivatives of the movement parameters. Three further regressors described the time course of signal intensities of white matter, cerebrospinal fluid, and the portion of the magnetic resonance image outside the skull. The fMRI time series were high-pass filtered (cutoff = 128 s). To correct for temporal autocorrelation, we used a first-order autoregressive model.
Group analyses
Contrast images of the effects of interest (approach–happy, approach–angry, avoid–happy, avoid–angry) were entered into a random-effects multiple regression analysis, separately for each condition and each gender. RPQ sum scores, log-transformed testosterone levels, and log-transformed cortisol levels were standardized within each gender group and added as gender-specific and condition-specific covariates, yielding 24 extra regressors. Participant-specific regressors were added to control for overall between-subjects effects. Following previous work (Tyborowska et al., 2016; Volman et al., 2011), we tested for a congruency effect by contrasting the activation differences during both affect-incongruent conditions (approach–angry, avoid–happy) with those during both affect-congruent conditions (approach–happy, avoid–angry). Subsequently, we tested whether aggression, as measured by the RPQ sum scores, was associated with differences in the congruency effect. Finally, we tested whether this potential relationship between aggression and congruency effect was modulated by testosterone levels by adding a testosterone-by-aggression interaction term to the model, separately for gender and task conditions. These effects were assessed at the whole-brain level, as well as within a priori defined regions of interest—namely the aPFC and bilateral amygdala, regions that have been shown to be sensitive to approach-avoidance-task-related congruency effects before (Volman et al., 2011; Volman et al., 2016). Following Tyborowska et al. (2016), we used the lateral frontal pole region defined in the frontal cortex parcellation atlas of Neubert, Mars, Thomas, Sallet, and Rushworth (2014) as a volume of interest (VOI) for the aPFC, whereas the amygdala VOI was anatomically defined on the basis of the Automated Anatomical Labeling Atlas (Tzourio-Mazoyer et al., 2002). The reported activation clusters were corrected for multiple comparisons using family-wise error (FWE) correction. Inferences for whole-brain analyses were made at the peak level (pFWE < .05). Small volume corrections (SVCs; p < .05) were performed for the VOIs on the basis of an initial cluster-forming threshold of p < .005. Anatomical inference was drawn by superimposing the thresholded SPM T maps on the canonical SPM single-participant T1 image.
To explore the effect of aggression on the activation change between congruent and incongruent trials, we extracted the mean beta values for the significant clusters in the amygdala and aPFC, as found in the aggression-by-congruency interaction effects. Subsequently, we tested the correlation (Pearson’s r) between the gender-normalized RPQ sum score and beta values for both conditions and both regions.
To test the generalizability and specificity of the effects of trait aggression, we selected a group of 72 (54 males) age- and gender-matched civilians. Data acquisition, analysis of behavioral data, preprocessing of imaging data, and first-level modeling of imaging data were performed using the same procedure as described in the Method section. Contrast images of all participants (police and civilian) were then entered into a group analysis following the procedure described in the Method section, with one adaptation: Instead of two groups (male, female), four groups were defined (male police, female police, male civilian, female civilian), and each task condition and covariate was modeled separately for each group.
Functional-connectivity analyses
Interregional connectivity between the bilateral amygdala and bilateral aPFC during emotional control was explored using a psychophysiological-interaction analysis (Friston et al., 1997), with the bilateral amygdala VOI as the seed region. Participant-specific contrast images were generated describing the psychophysiological interaction between the time course of the amygdala VOI and the time course of the affect-incongruent versus affect-congruent conditions within the approach-avoidance task. These images were entered in a multiple-regression analysis, with gender-normalized RPQ scores, log-transformed testosterone levels, log-transformed cortisol levels, and the RPQ-sum-score-by-testosterone interaction term as regressors. In addition to the whole-brain analysis, we performed a VOI analysis with the same bilateral anatomical aPFC mask used for the group-task activation analyses (Neubert et al., 2014).
Results
We included 275 police recruits (209 males, 66 females; age: M = 24.70 years, SD = 5.32 years) from the Dutch Police Academy (see Table S1 for specifics on trait aggression, endogenous testosterone, and cortisol levels for each gender). Average aggression levels of the police recruits were within the range of normal nonoffender adults (Cima et al., 2013), and average testosterone levels were within the normal range for each gender (Dabbs, 1990). At the behavioral level, the typical approach-avoidance congruency effects were observed, reflecting emotional action control during incongruent responses: a significant Movement (approach, avoid) × Valence (happy, angry) interaction for RTs indicated that participants were slower to respond during incongruent trials compared with congruent trials, F(1, 269) = 57.189, p < .001, η p 2 = .175 (Fig. 1b). Moreover, participants made more errors during incongruent trials compared with congruent trials, indicated by a significant movement-by-valence interaction for error rates, F(1, 269) = 22.365, p < .001, η p 2 = .077 (Fig. 1c; see also Supplementary Behavioral Results in the Supplemental Material). Gender, trait aggression, and endogenous testosterone did not affect these behavioral congruency effects on RTs and accuracy, and aggression and endogenous testosterone were not significantly correlated (r = .046, p = .45). Thus, our task successfully evoked the emotional bias in approach-avoidance behavior, and performance at the behavioral level was unrelated to individual differences in aggression or testosterone. This latter finding is consistent with previous results using this mild fMRI-adapted version of the approach-avoidance task (Tyborowska et al., 2016; Volman et al., 2011).
Trait aggression is reflected in the neural circuitry of emotional control
First, we tested the hypothesis that trait aggression in police recruits is associated with relatively stronger recruitment of neural control circuits during control over emotional actions. Replicating previous studies (Roelofs, Minelli, Mars, van Peer, & Toni, 2009; Volman et al., 2011), our results showed that a congruency effect was observed in the left aPFC: Activation was increased during incongruent trials (avoid–happy and approach–angry) compared with congruent trials (avoid–angry and approach–happy; p = .003, whole-brain FWE corrected MNI values for local maxima: x = −22, y = 50, z = 8; see Fig. 2). This effect was also visible in the right aPFC, although it did not reach statistical significance (threshold p < .005, uncorrected, MNI values for local maxima: x = 24, y = 60, z = 16).
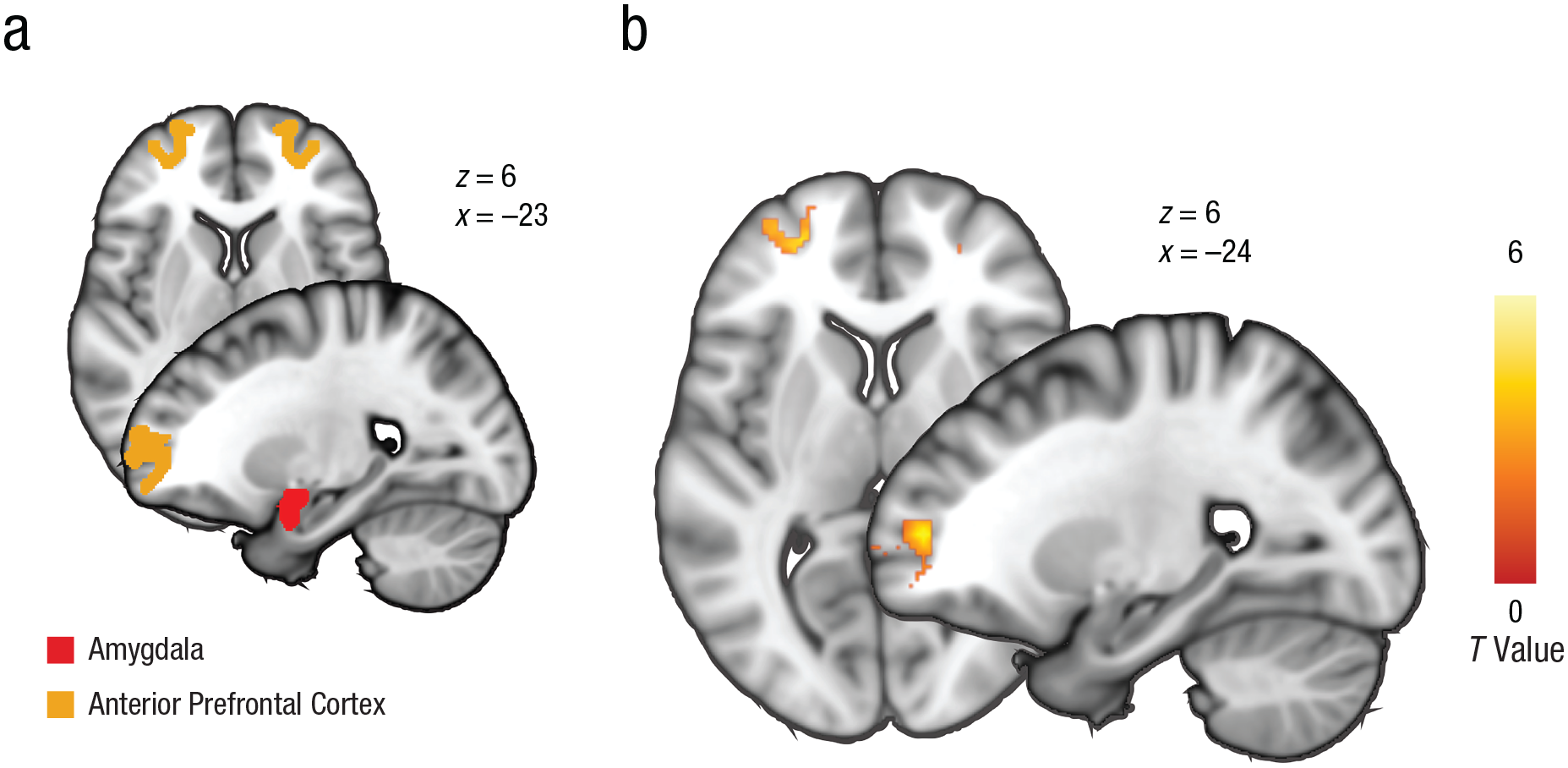
Brain masks and task effects. The brain images in (a) show anatomical masks of the bilateral amygdala and the anterior prefrontal cortex (aPFC) based on parcellation (defined by structural connectivity; Neubert, Mars, Thomas, Sallet, & Rushworth, 2014) selected as volumes of interest. The brain images in (b) show areas of activation in the left aPFC during emotional control (p < .05, family-wise error corrected, whole-brain corrected; incongruent > congruent trials). The cluster is masked by the bilateral aPFC mask at an uncorrected threshold of p < .005 for display purposes. (See Table S4 in the Supplemental Material available online for full cluster statistics.) Values shown next to brain images are Montreal Neurological Institute coordinates.
We further investigated whether activation differences within the aPFC and amygdala were related to behavioral performance. Consistent with previous work using the same task (Bramson, Jensen, Toni, & Roelofs, 2018; Roelofs et al., 2009), analyses showed that aPFC congruency effects were linked to behavioral congruency effects (increased error rates for incongruent trials vs. congruent trials), r = .17, p = .005. Note that all subsequent reported effects were assessed within the a priori defined regions of interest: the bilateral aPFC and the bilateral amygdala (see Fig. 2a and the Method section; whole-brain-level effects are listed in Table S2 in the Supplemental Material). In support of our first hypothesis, results showed that higher levels of aggression were coupled with a stronger increase in bilateral aPFC activation and a stronger decrease in bilateral amygdala activation for incongruent compared with congruent trials; this is compatible with increased recruitment of emotional-control neurocircuitry (left aPFC: pSVC = .021, right aPFC: pSVC = .001; local maxima: left aPFC: x = −18, y = 58, z = −10, right aPFC: x = 18, y = 58, z = −10; left amygdala: pSVC < .001, right amygdala: pSVC = .019; local maxima: left amygdala: x = −18, y = 2, z = −18, right amygdala: x = 24, y = 2, z = −16 (see Fig. 3). See Table S2 for significant clusters outside the regions of interest. Post hoc tests of the effect of trait aggression on bilateral amygdala activation for incongruent versus congruent trials revealed that this effect was primarily driven by increased amygdala activation during the congruent condition in individuals with relatively high aggression (r = .14, p = .02), whereas the aggression-related reduction during incongruent trials was not significant (r = −.10, p = .10; Fig. S1A).
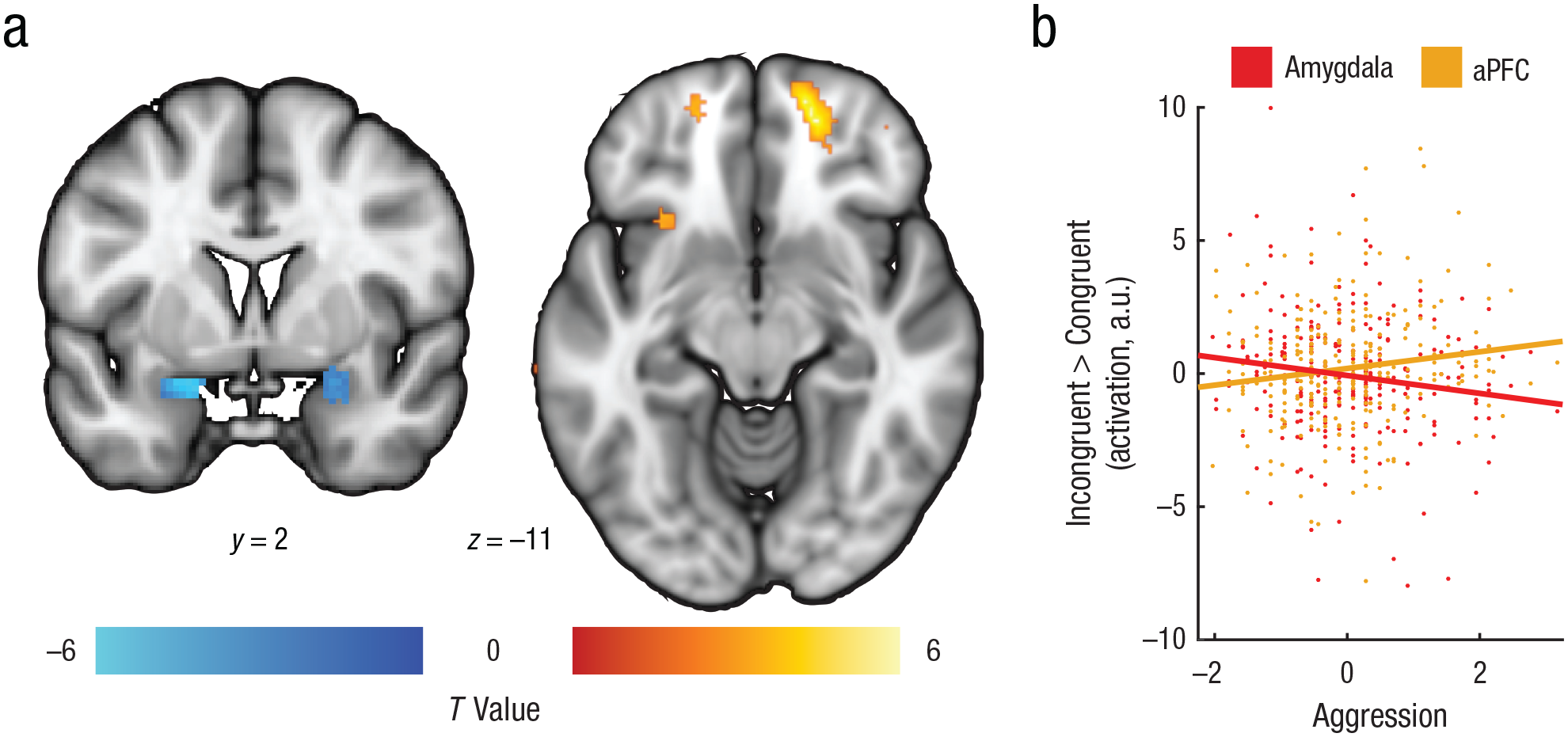
Aggression effects on brain activation during emotion control. The effects of aggression on bilateral amygdala and bilateral anterior prefrontal cortex (aPFC) activation during emotional control (incongruent > congruent trials) are shown in (a). The left image is masked with a bilateral amygdala mask, and both clusters are shown at an uncorrected threshold of p < .005 for display purposes (see Table S4 in the Supplemental Material available online for full cluster statistics). Values shown next to brain images are Montreal Neurological Institute coordinates. The scatterplot (b) illustrates the relationship between self-reported aggression (Reactive Proactive Questionnaire sum z scores) and differential activation in the amygdala and aPFC for incongruent > congruent trials; contrast estimates are given in arbitrary units (a.u.s) extracted from the bilateral activation clusters. Best-fitting regression lines are shown for results from each region.
For the aPFC, the effects of trait aggression were driven by stronger recruitment during the incongruent condition than during the congruent condition (congruent: r = −.01, p = .86; incongruent: r = .14, p = .02; Fig. S1B). Thus, high trait aggression was associated with increased emotional reactivity (reflected by increased amygdala activation during congruent trials), possibly requiring compensation during emotional control (reflected by increased aPFC activation during incongruent trials). Follow-up analyses exploring the potential modulating effects of valence (angry faces vs. happy faces) and gender within the amygdala and aPFC on the aggression-related effects revealed no significant clusters. In sum, trait aggression is related to increased aPFC activation during successful control over automatic emotional action tendencies, as well as with increased amygdala activation during automatic emotional actions.
Endogenous testosterone moderates the impact of trait aggression on emotional control
In line with our second hypothesis of the detrimental effects of testosterone on frontal emotion control, results showed that the aggression-related recruitment of the left aPFC during emotional action control decreased as a function of state testosterone levels. A congruency-by-aggression-by-testosterone interaction effect indicated that the effects of aggression on aPFC recruitment during emotional control (incongruent > congruent trials) depended on testosterone: Aggressive police recruits with relatively high state testosterone levels showed less left aPFC recruitment compared with aggressive police recruits with relatively low testosterone levels (pSVC = .001, local maxima: x = −24, y = 54, z = −6; Fig. 4). For the bilateral amygdala, we did not find such effects. Follow-up analyses exploring potential modulating effects of valence (angry faces vs. happy faces) and gender on the testosterone-by-aggression interaction effect within the aPFC yielded no significant results.
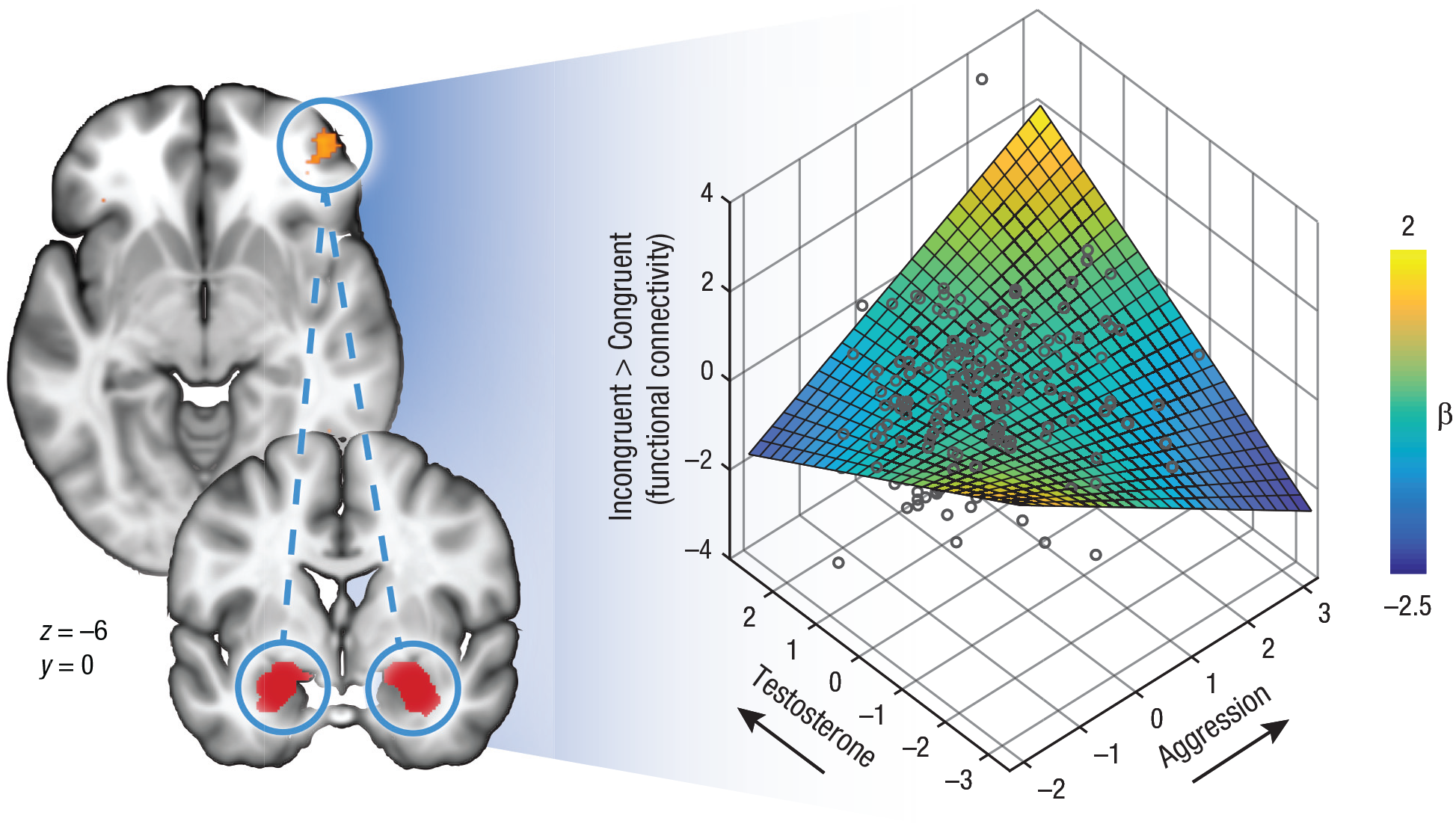
Relationship of testosterone and aggression with functional connectivity between the bilateral amygdala (seed region) and the anterior prefrontal cortex (aPFC). In the brain images on the left, the bilateral amygdala in highlighted in red, and the right aPFC is highlighted in yellow. Values shown are Montreal Neurological Institute coordinates. The 3-D scatterplot shows the individual data points. The 3-D surface was constructed from the best-fitting regression lines.
Next, we assessed the effects of testosterone and aggression on interregional connectivity within the frontoamygdalar network during emotional action control. A psychophysiological-interaction analysis was conducted using the bilateral amygdala as the seed region, because activation in this region was specifically linked to the aggression-by-congruency interaction as described above. This analysis provided further support for our second hypothesis on the negative association between endogenous testosterone and frontal-emotional control. A congruency-by-aggression-by-testosterone interaction effect in the right aPFC (pSVC = .035, local maxima: x = 42, y = 52, z = −6) indicated that individuals with relatively high aggression and low state testosterone showed the expected emotion-control-related pattern of negative aPFC–amygdala connectivity. In contrast, aggressive individuals with high state testosterone did not show this pattern of negative connectivity, β = 0.19, p = .007 (Fig. 4; see Supplementary fMRI Results in the Supplemental Material for all simple-slopes effects). There were no significant main or interaction effects of congruency or aggression on either a whole-brain level or within the bilateral aPFC. Thus, our data indicate that high trait aggression in combination with high testosterone levels is associated with reduced recruitment of the left aPFC and less negative coupling between the right aPFC and the bilateral amygdala.
To test whether the results held for each gender group separately, we correlated the extracted mean beta values of the incongruent > congruent contrast for all significant aPFC and amygdala clusters (including the psychophysiological-interaction effect) to aggression and the testosterone-by-aggression interaction, separately for males and females. These post hoc analyses demonstrated that all results held in both gender groups (all ps < .05). Moreover, to control for effects of age and interactions between cortisol and testosterone (Mehta & Josephs, 2010), we performed post hoc regression analyses incorporating these variables on all significant clusters. All effects remained significant (p < .002).
Discussion
Our results indicate that increased trait aggression is associated with increased aPFC activation during successful control over automatic emotional action tendencies, as well as with increased amygdala activation during automatic emotional actions. Interestingly, aPFC control over the amygdala in police recruits with high trait aggression decreased as a function of high state testosterone. These results suggest that aggressive individuals who are mentally healthy and high functioning are able to compensate for their (amygdala-driven) emotional reactivity using a frontal-control network, involving the aPFC, but that this system fails under conditions of high testosterone levels. These findings are relevant for populations that need to handle socially challenging situations, which are known to increase testosterone levels (Archer, 2006). Moreover, they provide a neuroendocrine explanation for the paradox that police officers, who are selected for their high cognitive control, may be impeded under emotionally challenging situations associated with testosterone elevations.
Altered functioning of the prefrontal–amygdala circuitry in relation to aggressive traits is consistent with previous findings in aggressive human samples (Coccaro et al., 2011), as well as in lesion studies in animals (De Bruin, Van Oyen, & Van De Poll, 1983). However, most human studies have focused on individuals with psychiatric conditions, such as borderline personality disorder (Bertsch et al., 2018), intermittent explosive disorder (Silverman & Siever, 2002), and psychopathy (Volman et al., 2016). Within patient populations, the findings point to reduced functioning of the prefrontal cortex, including damage to the frontal cortex, reduced baseline activity, and reduced glucose metabolism (Coccaro et al., 2011). We expand on this knowledge by showing that in a nonclinical, well-functioning sample of police recruits, high trait aggression is associated with increased recruitment of similar frontal-control structures, thereby possibly counteracting increased aggression-related emotional reactivity of the amygdala during congruent responses. These findings may be explained by the fact that police recruits are selected on the basis of high control in emotion-evoking situations (van der Linden et al., 2013), indicating adequate compensatory mechanisms for dealing with aggressive impulses. In line with this interpretation, additional analyses indicated that our police recruits had larger aPFC involvement during emotion control than an independent healthy control sample of 72 age- and gender-matched civilians (see the Supplemental Material). Moreover, high levels of aggression do not necessarily lead to unsuccessful behavior when adequately compensated for. For example, in contrast to psychopathic delinquents, successful people who score high on psychopathy traits do not show structural and functional prefrontal impairments, and they even display enhanced executive functioning (Gao & Raine, 2010).
Emotional-control-related aPFC activity and connectivity with the amygdala in police recruits with relatively high aggression were reduced as a function of high state testosterone levels. By demonstrating that testosterone influences the prefrontal cortex and its connectivity during social behavior, these findings extend previous research. For example, Mehta and Beer (2010) showed a relationship between high endogenous testosterone, reduced activation in the orbitofrontal cortex, and aggressive behavior in the ultimatum game. Volman et al. (2011) used the same approach-avoidance task as in the current study and found that in healthy individuals, endogenous testosterone is associated with reduced aPFC activity and reduced negative connectivity between the aPFC and the amygdala, indicating reduced control over emotional behavior.
Our finding that trait aggression moderates the effect of testosterone on prefrontal activity and connectivity expands on these observations, and it fits well with other recent studies indicating that the control-reducing effects of testosterone depend on individual characteristics: Testosterone levels are associated with reduced aPFC activation during emotion control in psychopathic individuals specifically (Volman et al., 2016), and only impulsive and dominant individuals are sensitive to the effects of exogenous testosterone on aggressive decisions (Carré et al., 2017).
Together these studies suggest that during emotional action control, highly aggressive individuals are more sensitive to the effects of high testosterone because their emotional control system is already challenged to a larger extent than that of less aggressive individuals. Such increased sensitivity has previously been associated with poor serotonin function, which codes for reactive aggressive outbursts in animals, in particular when coupled with high testosterone (Higley et al., 1996). Future genetic-hormone interaction studies should be employed to investigate whether genetic variations in the serotonin system underlie the effects of aggression on neural markers of emotional action control, as shown here.
It is noteworthy that studies on emotion regulation and aggression report the involvement of various subregions of the prefrontal cortex, including the orbitofrontal cortex and the medial prefrontal cortex (Coccaro et al., 2011; Mehta & Beer, 2010). Although we observed aggression-related activation extending into the orbitofrontal cortex, our results point to a crucial role for the adjacent lateral aPFC (or lateral frontal pole). This structure developed late in human lineage and has no obvious homologue in macaques (Neubert et al., 2014). It has been associated with task switching and cognitive branching—that is, maintaining representation of alternative courses of action (Mansouri, Koechlin, Rosa, & Buckley, 2017). This function seems crucial to optimal performance during the approach-avoidance task, which requires arbitration between the instructed response rules and the automatic action tendencies triggered by the emotional stimulus (Roelofs et al., 2009; Tyborowska et al., 2016; Volman et al., 2011). By enabling consideration of alternative long-term outcomes over immediate action outcomes, the aPFC may also support the ability to systematically engage in prosocial activities. This fits with the ability to override automatic impulses in favor of a more abstract goal, even in healthy individuals with relatively high aggression, and hence, heightened emotional reactivity, as shown in this study. Consistent with knowledge about the aPFC (Mansouri et al., 2017) as well as a recent meta-analysis of fMRI studies using the approach-avoidance task (Koch, Mars, Toni, & Roelofs, 2018), our findings revealed that bilateral task-related aPFC activation was dependent on aggression levels. The fact that the testosterone-by-aggression interaction modulated activity in the left aPFC may be a function of statistical thresholding; a few previous studies have reported unilateral effects either in the left or right aPFC (Bertsch et al., 2018; Roelofs et al., 2009; Volman et al., 2016).
A strength of this study is the large, homogenous sample of 275 police recruits and the combination of measurements taken at the neural, endocrine, behavioral, and symptom (trait aggression) levels. Note that for stable estimates of correlations, only sample sizes above 250 are regarded as sufficient (Schönbrodt & Perugini, 2013). However, trait aggression was assessed using a self-report questionnaire—the RPQ. Although this questionnaire is well validated and appears to differentiate well between aggressive delinquents and healthy controls (Cima et al., 2013; Raine et al., 2006), it would be helpful in future research to include real-life observations of aggression during police work. From this perspective, it is interesting that within this sample of police officers, accuracy of rapid shooting decisions in a separate behavioral task (Hashemi et al., 2019) was negatively correlated with endogenous testosterone levels, underlining the relevance of testosterone in impulsive or aggressive decision making (see Fig. S2 in the Supplemental Material). Because we did not manipulate testosterone levels directly (i.e., by testosterone administration), we cannot draw any conclusions about the causal role of testosterone in the relationship between aggression and emotional control. However, in line with the suggested disinhibiting effects of testosterone, recent studies on testosterone administration in women during similar approach-avoidance tasks showed faster RTs (Enter, Spinhoven, & Roelofs, 2016) and increased amygdala activation (Radke et al., 2015) during incongruent threat-approach responses compared with a placebo control group. Like previous researchers, we used an fMRI-adapted approach-avoidance task with a relatively mild social challenge optimized to detect individual differences in neural effects (Tyborowska et al., 2016; Volman et al., 2016). Accordingly, the behavioral effects were not related to variations in aggression and testosterone. However, behavioral work with more provocative (zooming) versions of the approach-avoidance task have shown that approach-avoidance behavior is influenced by individual characteristics, such as social anxiety or psychopathy (Heuer, Rinck, & Becker, 2007; von Borries et al., 2012). This zooming version of the task, in which the face grows larger when the joystick is pulled and decreases in size when the joystick is pushed away, is not compatible with fMRI, as the congruency effects would be confounded by the perceptual consequences of the zooming. However, future studies using an additional behavioral zooming task would enable further investigation of the mediating role of the brain in the relationship between hormones and behavior.
Finally, although a control sample was included to verify whether police officers overall differed from other healthy individuals in task-related activations, this sample (54 males, 18 females) was too small to reliably assess individual differences in the modulating effects of aggression and testosterone (e.g., Schönbrodt & Perugini, 2013). Future research is therefore needed to elucidate whether our findings generalize to an unselected general population.
To conclude, whereas trait aggression appears to be counterbalanced by increased frontal activation during emotional action control, this compensatory mechanism seems to go awry when testosterone levels are high. This potential neuroendocrine mechanism for the occasional outbursts of poor behavioral control, such as excessive violence, may have implications for selection processes and individualized training programs for first responders.
Supplemental Material
Kaldewaij_SupplementalMaterial_rev – Supplemental material for High Endogenous Testosterone Levels Are Associated With Diminished Neural Emotional Control in Aggressive Police Recruits
Supplemental material, Kaldewaij_SupplementalMaterial_rev for High Endogenous Testosterone Levels Are Associated With Diminished Neural Emotional Control in Aggressive Police Recruits by Reinoud Kaldewaij, Saskia B. J. Koch, Wei Zhang, Mahur M. Hashemi, Floris Klumpers and Karin Roelofs in Psychological Science
Footnotes
Acknowledgements
We thank all our participants; Annika Smit and other colleagues at the Dutch Police Academy for their help with recruiting participants; Ingrid Kerstens, Tiele Döpp, Naomi de Valk, Leonore Bovy, and Lisanne Nuijen for their help with recruiting participants and conducting the experiment; and Vanessa van Ast for her help with setting up the study.
Action Editor
Eddie Harmon-Jones served as action editor for this article.
Author Contributions
All authors contributed to the study design. R. Kaldewaij, W. Zhang, and M. M. Hashemi conducted the experiment. R. Kaldewaij analyzed the data. All authors interpreted the results, wrote the manuscript, and approved the final version of the manuscript for submission.
Declaration of Conflicting Interests
The author(s) declared that there were no conflicts of interest with respect to the authorship or the publication of this article.
Funding
This study was funded by the Netherlands Organisation for Scientific Research (NWO Vici Grant 453-12-0010) and by a starting grant from the European Research Council (StG2012 313749), both awarded to Karin Roelofs.
Open Practices
Data for this study are available from the corresponding author on request. Materials have not been made publicly available, and the design and analysis plans were not preregistered.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.