
Abstract
Migration to urban centres is among the most important forces in contemporary urban studies. In this paper, we study how the demography and epidemic profile of a community are altered when they transition from living in nomadic conditions in a forested environment to a peri-urban settlement in a city of the Amazon basin. We analyse demographic and epidemic data with a multilevel model to understand individual and community-level effects in terms of the risk of malarial infection. We show that malaria becomes endemic when the population settles in the peri-urban area of the city. We also show that the reproductive rate of women in the group increases as they become sedentary, and that while individual fertility rates have no effect on risk of contracting malaria, population-level fertility rates are associated with malaria endemicity.
I. Introduction
Since 1900, the total global malarious land area has almost halved, being reduced from 77.6 million square kilometres in 1900 to 39.8 million in 2010.(1) However, almost half of the world population lives at risk of contracting malaria, with a 2015 estimate of 212 million cases per year.(2) Today 3.41 billion people are exposed, more than three times the 892 million exposed in 1900.(3) The decline in malarial incidence and areas at risk, combined with general increases in at-risk populations, suggests the transformation of malaria’s epidemiological characteristics. That is, malarial infection risk has become more intense in circumscribed areas around the world.
Actual incidence of malarial infection has been declining in the world over the past few decades, but often re-emerging in locations where it was no longer a persistent problem, as shown by the data collected by Nájera et al.(4) about the most severe epidemics in the world during the 20th century. Five countries (India, the former USSR, Brazil, Ethiopia, Sri Lanka) have had epidemics with more than 1 million cases and more than 10,000 deaths. In those nations, people are vulnerable due to their socioeconomic status, cultural and environmental factors, and homes near sites of urban agriculture.(5)
Since malaria is generally associated with rural environments where Anopheline mosquitoes thrive, studies have found that transmission of malarial infection is reduced in urban areas such as in the Republic of the Congo,(6) Benin (except for coastal areas)(7) and Burkina Faso.(8) However, all of these studies were conducted in large cities (population greater than 500,000, most with more than 1 million), whereas most of the global increase of urban inhabitants is expected to occur in smaller cities.(9) Currently, more than half of the world population lives in urban areas, and the number is expected to increase, particularly in cities with fewer than 1 million inhabitants.(10)
Urbanization has been relegated to a secondary role in models to understand changes in the vulnerability of the world population over time, even though the effect of urban population growth on epidemiology is more predictable, better understood, and can be anticipated more accurately than before.(11)
As with the effect of urbanization on the epidemiology of malaria, the effect of migration on urbanization is understudied. Some researchers have addressed the degree to which the increase in urban dwellers that is expected to occur over the next few years will be influenced by changes in demographic profiles of current urban populations, migration from rural areas, or both (but the discussion remains open).(12)
Thus, the effect of demographic changes on the epidemic profile of human populations remains poorly understood. Even though it is a popular area of research in epidemiology, most of the work focuses on the effect of malarial infection upon human demographics and not the other way around.(13)
Population change over time has traditionally been explained by a model known as the demographic transition. Generally it presents a decline in birth rates and then a decline in fertility rates.(14) The model consists of five steps. First, high birth rates and high death rates keep in balance the size of a population. Second, death rates fall due to improvements in healthcare and sanitation while birth rates remain high, increasing the size of a population. Third, the gap between birth and death rates narrows due to the availability of contraception and fewer children being needed to work – due to the mechanization of farming, which slows but does not stop the increase of the population. Fourth, low birth rates and low death rates are stable, maintained by a large population with wide access to birth control and a desire for smaller families. And fifth, an ageing population and the continued desire for smaller families, with people opting to have children later in life, start to decrease the size of the total population.
Together, those dynamics account for a rapid population expansion that is often described as the end of a Malthusian stage.(15) Altogether, the population dynamics described as the demographic transition show remarkable consistency among countries; there is a high degree of heterogeneity as to the rate, timing, and factors associated with this transition in particular locales.(16) Most importantly, little is known about how the demographic transition affects sub-populations within a country.(17) Demographic transitions are normally described in terms of national averages, which are coarse and aggregated, and thus not capable of or designed to capture this process in subregions or specific populations.(18)
Also, migration and the pressure of urbanization play an integral role in the demographic transition.(19) Zelinsky was the first to suggest that the demographic transition is composed of a vital transition (changes in birth and death rates), as well as a mobility transition (which refers to resettlement of the rural population in urban settings).(20) De Vries further developed the idea, noting a decline in urban mortality by means of acute reductions in infectious diseases as one of the main forces of the demographic transition (including a mobility component).(21) Yet the role of migration (and disease) in the demographic transition remains greatly understudied, in spite of promising research connecting the pressure of urbanization, the demographic transition, and changes in epidemic profiles.(22)
In this paper, we address the effects that becoming urban dwellers has upon a population’s demographic and epidemic patterns. We study how the demography and epidemic profile of a community are altered when it moves from living in nomadic conditions in a forest environment to a peri-urban settlement in a city of the Amazon basin. While the effect of malaria infection on human demography has been documented in great detail, with multiple reports of malaria infection skewing genetic variability,(23) few, if any, studies exist that enquire about the effect of underlying demographic patterns on malaria epidemics (i.e., how change in population structure affects epidemic patterns).
Moreover, we address whether demographic parameters affect malaria epidemics. Does population structure affect the epidemiology of malaria at both individual and population levels? Inverting the traditional logic used for malaria research may reveal interesting and important insights with applications for reduction of malaria transmission and infection.
II. Background
a. Malaria and demographic studies
The Garki project,(24) the most comprehensive longitudinal study conducted on malaria to date, included an extensive chapter on demography. This project enquired exclusively about the effect of the disease on the host population and yielded interesting findings. No significant differences were detected in the population structures of individuals based on presence or absence of advantageous genetic mutations. This research also highlights one of the most challenging aspects of malaria: there was no detectable preventive effect on malaria mortality from different interventions and controls between locations.(25)
Similarly, Pringle found no effect of changing epidemic patterns on the fertility rates of human populations of the Mkomazi valley of South Pare in Eastern Africa, when malaria re-emerged due to changes in public health policies (suspension of dieldrin spraying to control vector populations).(26) However, the same study did find an association between malaria infection and child mortality.(27) These results are consistent across other countries. In Kenya, Spencer et al. found no effect of providing chloroquine phosphate on malaria-induced mortality or fertility.(28)
Substantial evidence indicates that demographic dynamics are closely related to malaria epidemiology. Socioeconomic research has shown that high infant and child mortality rates are positively associated with high fertility rates, while negatively associated with household income and education level.(29) Substantial evidence links malaria eradication programmes with an increase in fertility and decrease in low birth weight in different countries, such as Sri Lanka,(30) nations in sub-Saharan Africa(31) and Papua New Guinea.(32) However, some global analysis suggests that this relationship holds stronger in areas of unstable transmission, and is not characteristic of Latin America and the Caribbean.(33) As well, some authors suggest that the relationship between malaria and low birth weights can be overestimated when the latter is not adjusted by gestational age.(34)
Furthermore, recent evidence suggested that differences in malaria intensity were not only attributable to human genetic diversity.(35) Kwiatkowski argues that malaria has been the most powerful force for current selection on human genetic variants, producing differences in genetic resistance and susceptibility to human malaria around the world. Yet the haplotypic diversity is the result of gene–environment interactions too.(36) Thus, demographic dynamics, such as fertility, are part of the environmental dimension, which is needed in order to understand much better the intensity of malarial infection in human populations.(37) Here, we propose a novel model to evaluate the relationship between malaria and fertility: increased fertility in certain areas promotes malaria endemicity by increasing the susceptible proportion of the population. This is consistent with documented observations of southern Italy, where malaria endemicity occurs in Sardinia, precisely where fertility is higher.(38) Furthermore, that population – subject to intense malaria exposure – has developed higher frequencies of genetic traits that confer protection against malaria infection(39) as well as a delayed and more effective childbearing (lower mortality of offspring) than found in neighbouring communities.(40) The case of Sardinia is an example of how much the demographic transition can vary, and how much the influence of malaria can affect the population trajectory undergoing such a transition.(41)
b. Individual and population-level risks
While most models view demography as either a risk factor at an individual level for malaria or a consequence of malaria epidemics, here we propose a different approach: we evaluate the probability of malaria infection in terms of individual and population-level risk factors. This distinctive approach follows a line of work by contemporary epidemiologists focusing on the ecological fallacy, where aggregated data are mistakenly used to assess individual-level risk factors.(42) The advent of computers provided the opportunity to study disease at an individual level, which, in combination with the development of genetic methods, shifted the focus of epidemiology to detailed studies of host behaviours and risk factors.(43) This trend was a response to predominantly population-level studies that preceded it, which due to the lack of computational power to run multilevel statistical tests, presented analyses that involved group-level effects of risk factors for disease.(44) However, multiple epidemiologists have begun working over the past 20 years on the implementation of multilevel statistical models that explore both individual and population-level effects simultaneously.(45)
The notion that understanding individual and population-level dynamics is important to correctly portray epidemiological aspects of disease has been associated with malaria since the first mathematical formulations in the field.(46) In his theory of dependent happenings more than 100 years ago, Ross suggested that the number of infected individuals within a population is one of the main drivers of individual risk of infection by plasmodia.(47) Similarly, in vaccine-preventable diseases, individual risk may be characterized by whether the individual is vaccinated, along with the level of vaccination of the entire population, because the level of herd immunity is crucial to estimate the individual’s risk of infection (independent of the individual’s vaccination status).(48)
In vector-borne diseases, population and individual-level effects interact in complex ways. Without an appropriate understanding of multilevel effects upon an individual’s risk of becoming infected, erroneous interpretations can arise from multilevel phenomena, yielding ineffective public health interventions. Koopman and Longini Jr discuss the case of yellow fever, where at low infestation, individual risk factors prevail, such as the presence of mosquito breeding sites in the household, while at high pathogen loads among the population, a person can be inoculated by infected mosquitoes breeding in neighbouring houses, the market or school.(49)
c. Epidemic and demographic profile of Colombia
Colombia’s population grew by a factor of 11 in 110 years, from 4.7 million in 1905 to a projected 53 million in 2015.(50) By the second half of the 20th century, Colombia had changed from being a predominantly rural population (only 31 per cent of the population lived in an urban area in 1938) to predominantly urban (72 per cent urban inhabitants in 2000).
The most interesting demographic characteristic is that the country underwent the demographic transition during different, but consistent, periods of violence. Not only does death due to violence not seem to counter the trend of decreasing mortality and increasing life expectancy, it probably played a role by increasing the number of people living in urban areas. Although mortality rates began to drop in the 1940s, it was not until the 1950s that acute reductions were seen in mortality rates, which also coincided with the rapid urbanization of the population and the fastest growth rate, over a span of 40 years. The onset of Colombia’s 10 per cent fertility decline occurred in the second half of the 1970s. Thus, the gap between the two observed patterns for the demographic transition is approximately 20 years. While the demographic transition experienced by Northern Europe occurred over 75–100 years, this transition in Colombia is happening faster and faster, and the remarkable decrease in fertility is largely attributable to a nonprofit organization, Profamilia, led by progressive doctors.(51) This explains the apparent paradox that Colombia’s fertility decline was more pronounced than that of similar countries, like Mexico, with higher indexes of socioeconomic development.
Interestingly, the indigenous population has experienced demographic change in a completely different manner than the rest of the Colombian population. Its indicators vary considerably from the average rates documented across the country, depicting an alternative phenomenon to the demographic transition. In 1993–1994, 45 per cent of the indigenous population was under 15 years of age, the fecundity rate was 6.5 children per woman, infant mortality for 1990 was estimated at 63.3 per 1,000 live births, and life expectancy was 57.8 years for females and 55.4 years for males. This suggests that the indigenous population remains in pre-demographic transition rates.(52)
d. Malaria in the Amazon basin
Outbreaks of malaria in the Amazon have been associated primarily with two factors. First, migration processes alter the ratio of susceptible and infected individuals in such a way as to promote epidemics. Second, perturbations of primary or secondary forest alter the composition and behaviour of vector populations. The two factors often coincide with human occupation of the territory through diverse lifestyles, reflected in a wide range of intensity of malaria infection among different populations. As a result, malarial infection in the Amazon basin is considered to be associated with low incidence and prevalence in small and dispersed populations. In other words, it is hypoendemic (occasionally epidemic),(53) primarily associated with men of working age who travel into the forest (an occupational risk hazard).
Padilla et al. described similar results for urban and peri-urban malaria cases in the Pacific coast and Eastern regions from Colombia. There, malarial infection seems to be endemic, unstable and of low intensity;(54) thus it is increasingly difficult to detect foci of transmission.(55)
The principal vector responsible for malarial transmission in the Amazon basin is Anopheles darlingi, a species that breeds in forest fringes (between agricultural land and intact or secondary forest regrowth), with behavioural plasticity and genetic diversity that suggest it could be a species complex.(56) This mosquito exhibits two patterns of hours of peak activity in different locations, but is primarily associated with crepuscular blood meals of human origin.(57) The high vectorial capacity of Anopheles darlingi (the mosquito’s capacity to become infected with malarial gametocytes, live through the complete parasite cycle, which is the sexual stage of the parasite or sporogony, and finally infect another human with sporozoites) is one of the driving factors for malaria’s persistence in the Americas.(58) The characteristics of this vector enhance the persistence of malaria in the Amazon, which is being deforested due to factors such as logging, clearing for cattle ranchers and agriculture (mainly manioc, corn, rice and plantain), mining, road construction, and migration.(59)
The role of migrant populations in previously undisturbed areas has been particularly problematic in the Amazon. In Brazil, colonization programmes increased the population of the Amazon from 7.2 million in 1970 to 18.7 million in 1996. The number of malaria cases in the region, accounting for 99.7 per cent of the total number of cases of the country, also increased dramatically over this period, and by 1999 the number was estimated at 600,000.(60) A longitudinal study conducted in the region described malaria as unstable and hypoendemic, affecting mostly adult males.(61)
In Peru, malaria re-emerged between 1992 and 1997 in the Amazon region of Loreto, increasing 50-fold during that period.(62) The characteristics of the population affected are also consistent with the descriptions of the Brazilian Amazon, affecting mostly adult males who work in the forest.(63) One difference that stands out in the Peruvian case is that resistant strains of malarial parasites have been detected using both in-vivo and in-vitro testing.(64)
Similar conditions have been described for Colombia, where malaria occurs mostly among men of working age who labour in the forest.(65) While there is significant variation in malaria infection among ethnic minorities of the populations studied (even after controlling for family and household size),(66) further efforts have not aimed to account for the observed differences in disease prevalence between populations.(67) A notable exception suggests that warfare among headhunter societies of the Amazon not only reduces competition for resources, but also scares people away from their territory. This results in lower population densities, which reduce the presence of pathogens dispersed in the metapopulation (spatially separated populations belonging to the same species). When colonization forces increased the population densities of this region, the likelihood of malarial outbreaks was also observed to increase. Durham’s study(68) can be understood in terms of cooperation strategies where an almost unrecognized common good exists, and collective action leads to a context where the cost is experienced by everyone, there is no incentive for individuals to subvert, and the benefits are experienced by everyone in the group. The difference between individual and collective risk factors has been described as particularly important for vector-borne diseases in response to disease intensity.(69)
Other studies have reported isolated pockets of asymptomatic and endemic malaria among indigenous communities,(70) in what constitutes a confusing panorama of malaria in the Amazon. Studies suggest that indigenous practices are adaptive and reduce malarial transmission,(71) while conversely, indigenous communities are reported to experience asymptomatic malaria that is consistent with high parasite loads in the population.(72) However, no clarity exists as to the specific aspects that make some indigenous populations susceptible to malaria infection, while others are unaffected.
e. Urbanization in the Amazon basin
The seminal work addressing the advent of urbanization in the Amazon basin(73) describes the colonization of the Brazilian frontier as the effect of different forces (such as private capital, public policy, and economic forces) interacting with the process of urbanization. And these effects have become interconnected in an unpredictable and complex fashion. The urbanization of the Amazon is not necessarily associated with an increase of agricultural production(74) (and for different countries in the region, the urban population is the fastest-growing segment of the country).(75) Rural women inhabiting the Amazon have been documented to be more likely to migrate to urban areas than men in Ecuador,(76) where the demographic characteristics of the migrant population are associated with the type of occupation that promotes habitat degradation (more habitat fragmentation and dispersed land tenure than in the Brazilian Amazon).(77) Other studies suggest that men also migrate to “boom towns” of 10,000–20,000 people that form spontaneously in opportunistic migration, aiming to receive land or build a new business.(78) Thus, this regional process of urbanization matches a large body of literature calling for a redefinition of the dichotomous classification of urban and rural – one that can describe differences among the span of human settlements, and above all, the interaction and mobility between them.(79)
While most of the work studying the environmental effects of the urbanization of the Amazon has addressed habitat fragmentation,(80) some authors have also associated the process of urbanization in the Amazon with disease outbreaks (and in general with an unhealthier lifestyle).(81)
f. San José del Guaviare
During the 1990s and early 2000s, Guaviare was one of the most violent regions of Colombia, with an active war involving the army and illegal groups including paramilitaries and guerrillas. Illegal markets, including drugs, animal skins and, more recently, ornamental fish, have been the last of a series of booms. These booms perpetuate a series of occupation attempts in a region where cultural diversity, marginality and abandonment by the central government pose major barriers to livelihoods for the majority of the population.
Guaviare is one of Colombia’s largest departments by area (53,460 square kilometres), yet one of the least populated (107,934 inhabitants, 2.02 persons per square kilometre). Its GDP per capita is the equivalent of US$ 3,060 (contrasted with the national average of US$ 7,950).(82) According to an indicator developed by the UN Economic Commission for Latin America and the Caribbean (CEPAL), this department ranks 28th out of 29 in competitiveness. The CEPAL indicator takes account of human capital, science and technology, strength of the economy, infrastructure, and public management. At least 43 per cent of this department’s GDP comes from social or community services and personal businesses. Plantain (63.4 per cent) and manioc (30.7 per cent) are the main crops grown during the whole year, while rice (53.6 per cent), and corn (46.4 per cent) are subsistence crops. This region ranks 4th in molasses and 5th in rubber production among 29 departments studied.
Demographically, the Amazon region is one of the most fertile populations in Colombia, with 30.7 births per thousand per year from 2005 to 2010 (the national figure is 19.9). The mean death rate is slightly lower (5.7 per thousand) than the national average (5.8), while the negative migration rate (-8.4 per thousand) is greater than the national average (-2.3), and the growth rate (1.65) is higher than the national average (1.2).(83)
Guaviare was selected for a case study because this department presents several interesting aspects related to malaria and ethnicity. Most importantly, Guaviare harbours a multiplicity of ethnic communities living at risk of malaria infection, which have experienced forced displacement as well as living in distinctive conditions that would likely affect malaria transmission rates. Few malarial studies have been conducted in the area, and holoendemic (ubiquitous) malaria has not been reported to date there or throughout Colombia.(84)
The Nükak are a nomadic population organized in territorial groups, yet some groups have begun a sedentarization process in the past 30 years. Each group consists of a territorial or local band formed by an aggregation of agnates (people with the same male ancestor) and their allies.(85) Since the 1960s, the Nükak began having fortuitous encounters with settlers. In the late 1970s, intense colonization and the arming of coca agriculturalists reduced their territory. In 1988, a severe influenza epidemic affected the Nükak and forced several local groups to migrate to nearby villages to seek medical attention. In the first five years of permanent contact, epidemics killed nearly 40 per cent of the Nükak population.(86) As a result, healthcare became the linchpin of state programmes aimed at reducing mortality among the Nükak.
The armed conflict intensified in the early 2000s and resulted in the forced displacement of local groups to the region’s capital, San José del Guaviare, one of the cities with the fastest-growing urban populations in Colombia.(87) Since then, some local groups have lived in shelters in peri-urban areas of San José del Guaviare for significant periods to escape violence in their home regions. In these shelters, the Nükak live in precarious conditions and depend on humanitarian assistance of state agencies.
Here, we present data collected during two field seasons, for two separate Nükak populations, and data collected in a historical Nükak population.
Chécamo
Fieldwork was conducted from 31 December 2009 to 24 January 2010 by Carlos Del Cairo, in Chécamo, in the headwaters of the Inirida River close to the Las Chulas rapids, southeast of San José del Guaviare. In the early 2000s, this settlement was established by missionaries who were formerly part of the group New Tribes Mission, but are now affiliated to the indigenous religious network Misincol. With the missionaries’ assistance, among the few people fluent in both Spanish and Nükak, we reconstructed a genealogy for each domestic group using open-ended interviews, which allowed for the exploration of the memory of each woman’s births. Because it is taboo for the Nükak to talk about dead people, missionaries who were familiar with the practices and beliefs of the community handled this with care and respect.
Aguabonita
Field research for this study was conducted with two local assistants during the period October 2011 through May 2012 in Aguabonita, a peri-urban location of San José del Guaviare. The Nükak settlement consisted of 108 individuals during this study period. The Aguabonita settlement began as a temporary solution in 2006 for the Nükak population that had suffered forced displacement from their ancestral territory, the Nükak Indigenous Reserve, then under the control of illegal armed forces and with buried landmines. The Aguabonita settlement consists of various shelters, mostly built following techniques of neighbouring non-Nükak ethnic minorities, consisting of a basic structure with a palm leaf roof that provides support to hang hammocks. Sometimes the whole population decided to build a temporary camp elsewhere in the property, in both forested and cleared areas. At other times, all the house structures were occupied, while at times only one or two structures would be in use.
III. Research Study
A census of the community was conducted, asking all the individuals in the settlement their names (they change their Spanish names periodically) in the Nükak and Spanish languages, age, household, mother and father. Every person was assigned an individual ID code. Individual case reports of malaria morbidity for the population of Aguabonita were available through collaboration with the local Secretary of Health. Each case report is laboratory confirmed. Historical data on genealogy and medical records were compiled from archived field reports filed by Diehl.(88) Census data, including genealogies and morbidity reports, were gathered.(89) Figure 1 shows the study populations and their distance to nearby settlements.
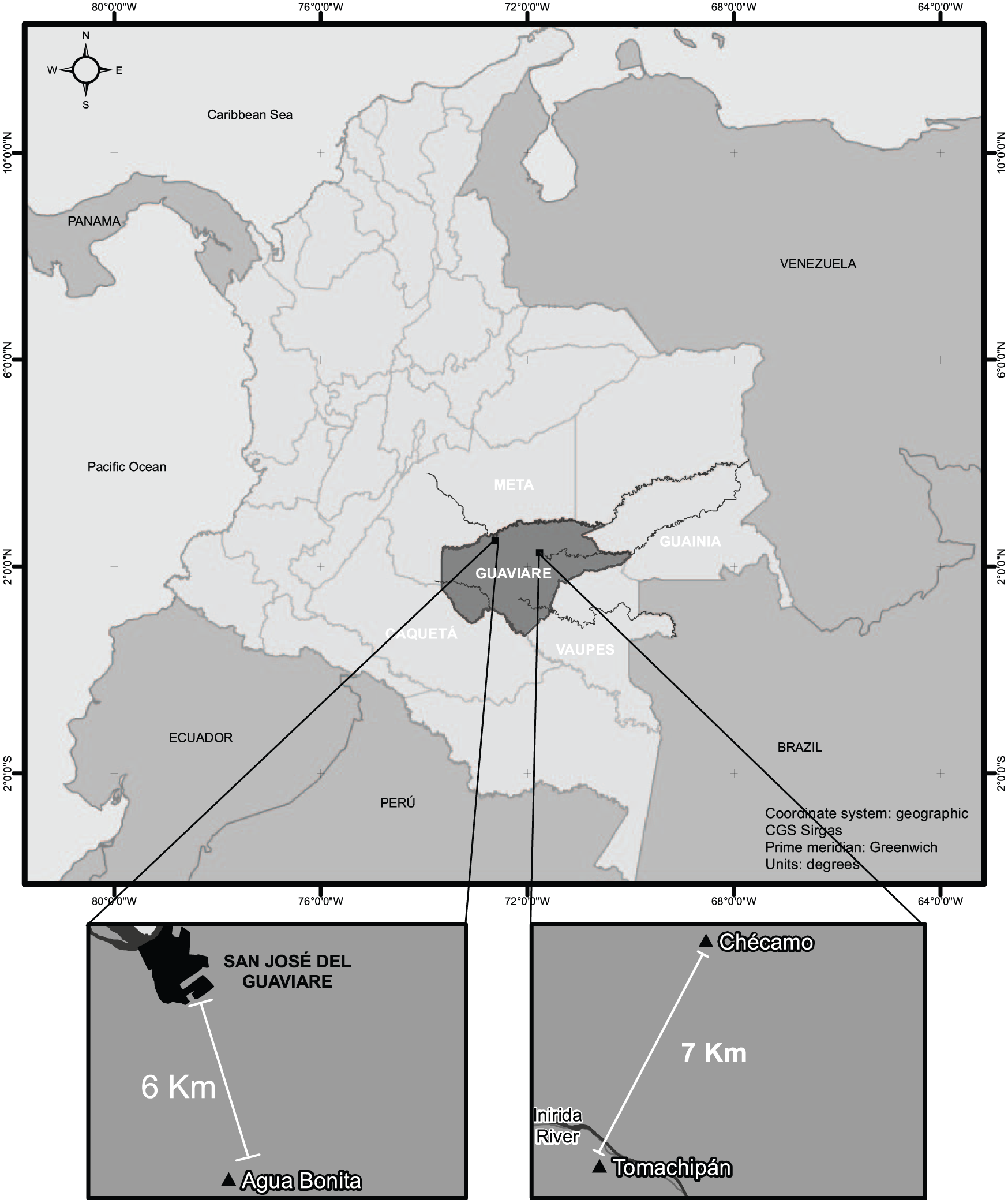
The studied populations and their distance to nearby settlements
The population sample comprises 108 individuals/foetuses in Aguabonita (50 females, 56 males, 2 in utero), 69 in Chécamo (33 females, 35 males, 1 in utero), and 54 in the sample documented by Diehl(90) (30 females, 24 males). Figure 2 shows the population pyramids for each population.
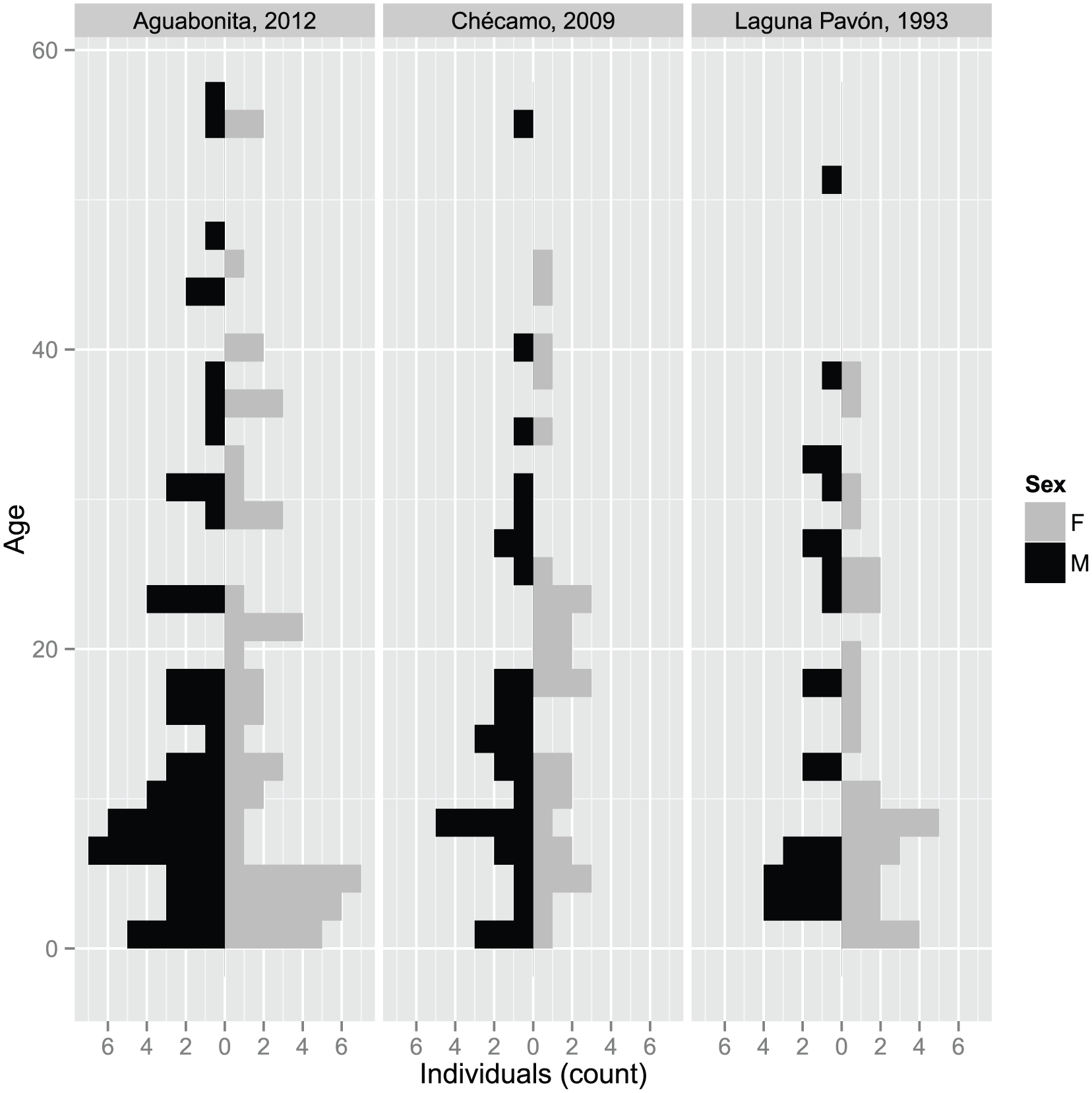
Population pyramids for the three Nükak populations
We used archival research performed by Diehl(91) to estimate malaria prevalence among the Nükak population before 1992; they reported 61 cases of malaria. The period during which these data were gathered was 1980–1988. Regional public health records used to estimate malarial prevalence among the Nükak population of Aguabonita reported 216 malarial cases from 2007 to 2011.
a. Models
We used a statistical model to perform a hypothesis test with the aim of determining whether the fertility patterns of women from the Aguabonita group were affected by the transition to becoming sedentary. We used data from other populations to explore whether the demographic patterns after settling in Aguabonita differed not only from their previous history, but also when compared with other Nükak groups living in various conditions of mobility.
The association of relevant variables and the birth hazard rate (the probability of a particular woman giving birth in a given year) was explored with a Cox proportional regression model. Model 1 explored associations with age, parity (number of viable pregnancies per woman) and settlement status, as well as the interaction between parity and settlement status. Model 2 excluded the interaction parameter. Model 3 excluded settlement status. Model 4 excluded parity. Age in all models was converted into three orthogonal polynomials to account for non-linear associations.
Descriptive statistics composed of histograms of malaria case reports were performed for the Nükak population of Aguabonita, as well as for the population studied by Diehl in 1992–1993. The purpose of this analysis was to identify patterns of endemic, asymptomatic malaria, as described for other indigenous populations of the Amazon basin.(92)
The association of malaria morbidity and demographic variables was explored by calculating the mean values for the Aguabonita population and the archival records collected by Diehl. A multilevel Poisson regression was used to explore the relationship between demographic variables and malaria outcome across populations (see the online supplement for a figure that explains this process).
IV. Findings
a. Demographics
After controlling for age and parity, being urban (sedentary) was found to have a significant positive effect on the birth hazard rate. Table 1 shows the Akaike’s information criterion (AIC) table for the models described, where Model 1 performed better than all the other models. Table 1 shows that the number of births increased as an effect of becoming sedentary and at a rate comparable to parity. Specifically, this model tested whether the population of Aguabonita experienced a demographic transition in 2005. The association suggested that as the population settled in the peri-urban area, birth intervals declined. Furthermore, the comparative sample with other Nükak populations suggested that by becoming sedentary the population of Aguabonita underwent a similar demographic transition as the population of Chécamo, a population that also presents a more sedentary existence, after controlling for the demographic characteristics of the population observed by Diehl in 1993.
Summary table for all models, including different parameters and interactions
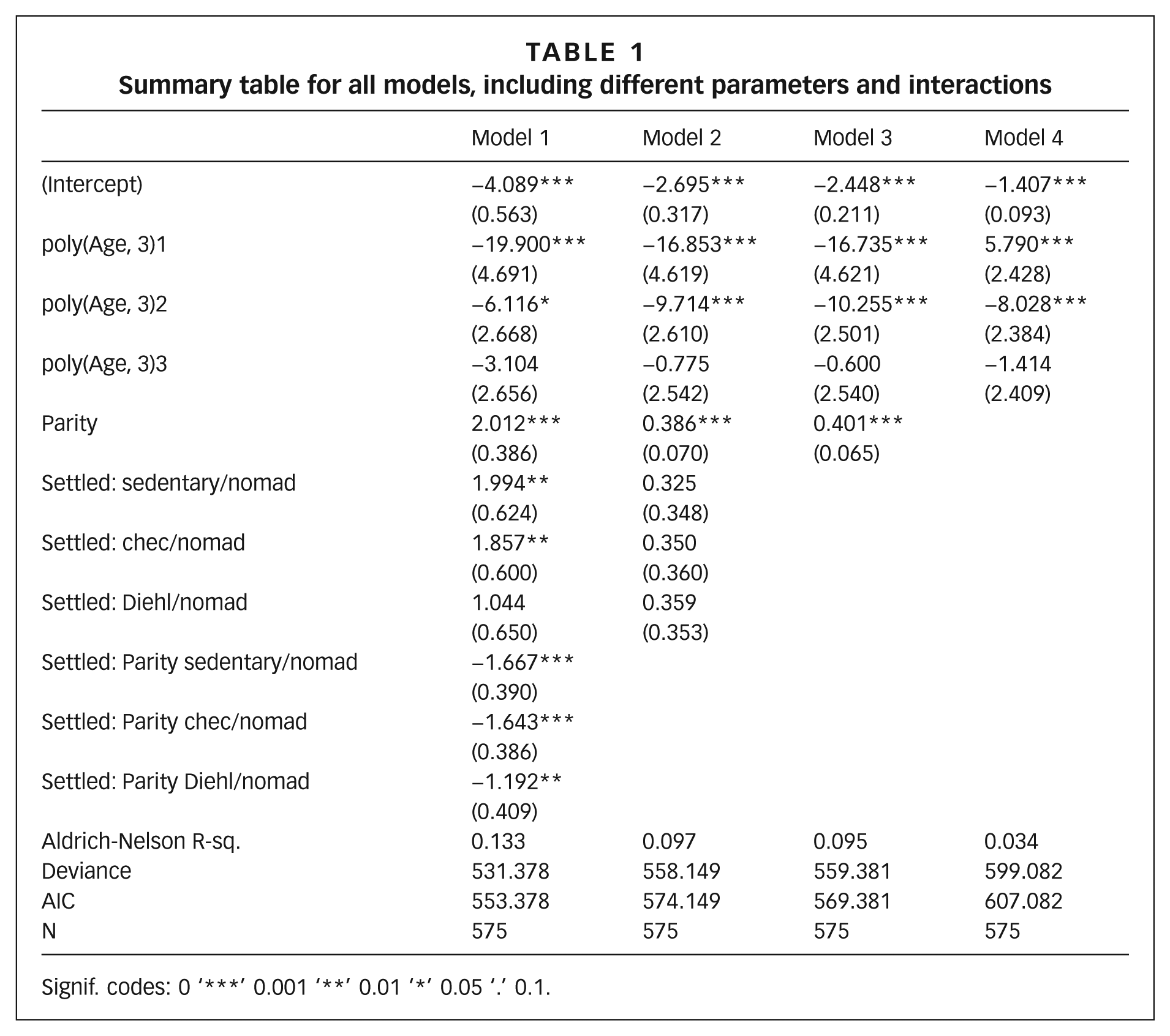
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1.
To examine recruitment (the number of children from each mother who make it to adulthood), we analysed the relationship between a woman’s fertility and the probability of survivorship of her progeny. Data were not available for time of death, and because death is a delicate subject among the Nükak, offspring survival probability was the best proxy to estimate this relationship. A linear model with child survivorship probability given a woman’s number of children found a significant negative effect between these two variables (p = 0.007, adjusted R2 = 0:8734; n = 11).
b. Epidemic
Figure 3 shows age-specific prevalence for the two focal populations. The epidemic pattern of malarial prevalence for the two populations showed two dramatic differences: the population of Aguabonita exhibits a pattern where prevalence was higher among children under 5 (holoendemic), consistent with reports for Amazonian populations living under endemic conditions, where asymptomatic malaria persists in adults.(93) The population studied by Diehl(94) indicated that the prevalence pattern before contact was not endemic, with a basic reproduction number (the number of infectious cases generated by a single case, on average – R0) for malaria lower than 1, below an epidemic threshold.
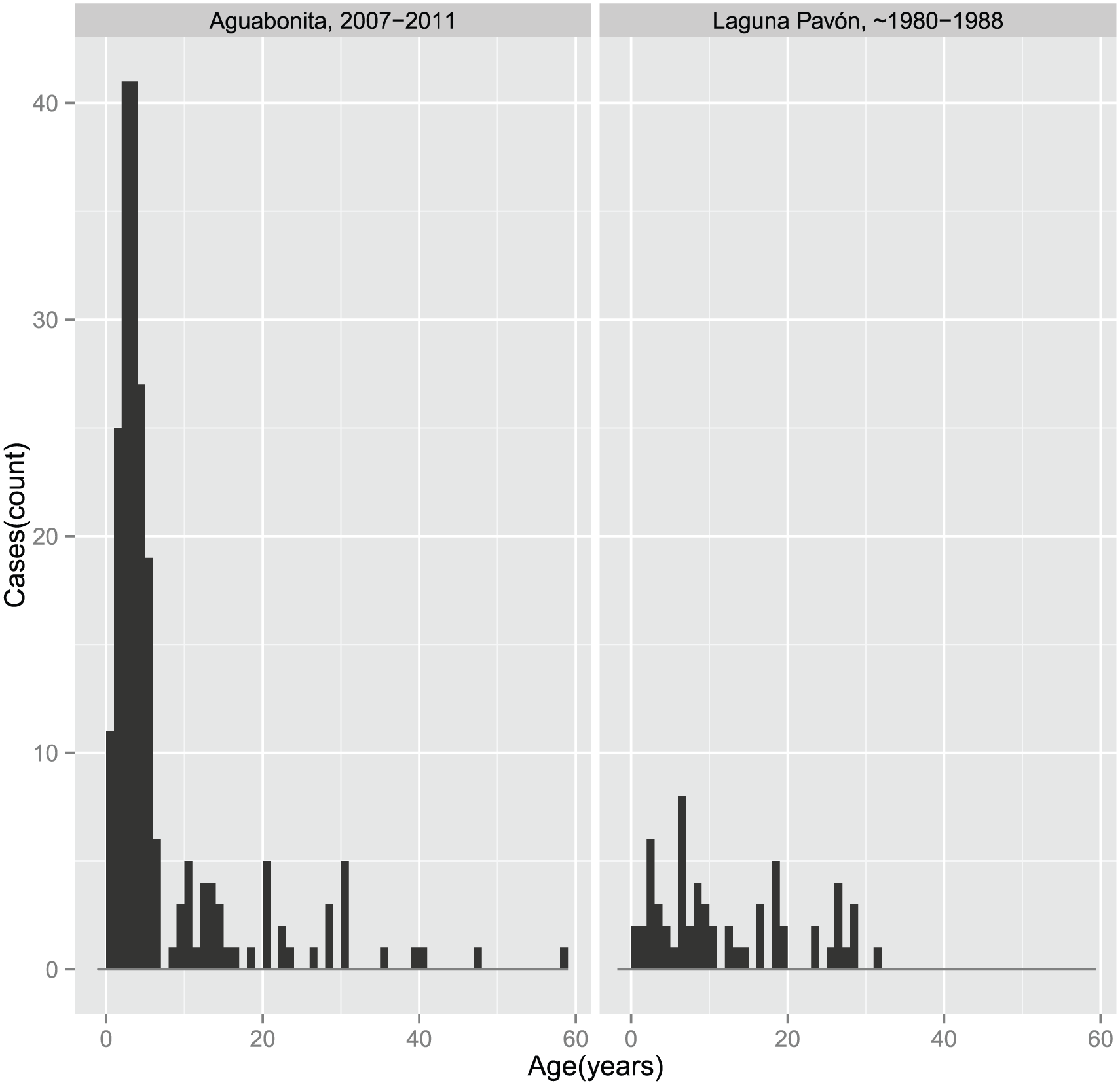
Malaria prevalence for the Nükak populations of Aguabonita (2007–2011), and the population studied by Diehl in Laguna Pavón II
c. Demography and epidemiology
No association was detected between demographic variables and malarial outcome at an individual level, in the model that has malarial cases among offspring as the outcome, and individual and population demographic values for each woman (e.g. parity, mean and standard deviation of interbirth interval [IBI]). However, after controlling for demographic variables, we find a significant effect of population on malarial outcome. Table 2 shows different models used to compare demographic variables and malarial outcome.
Summary table for all models, including different parameters
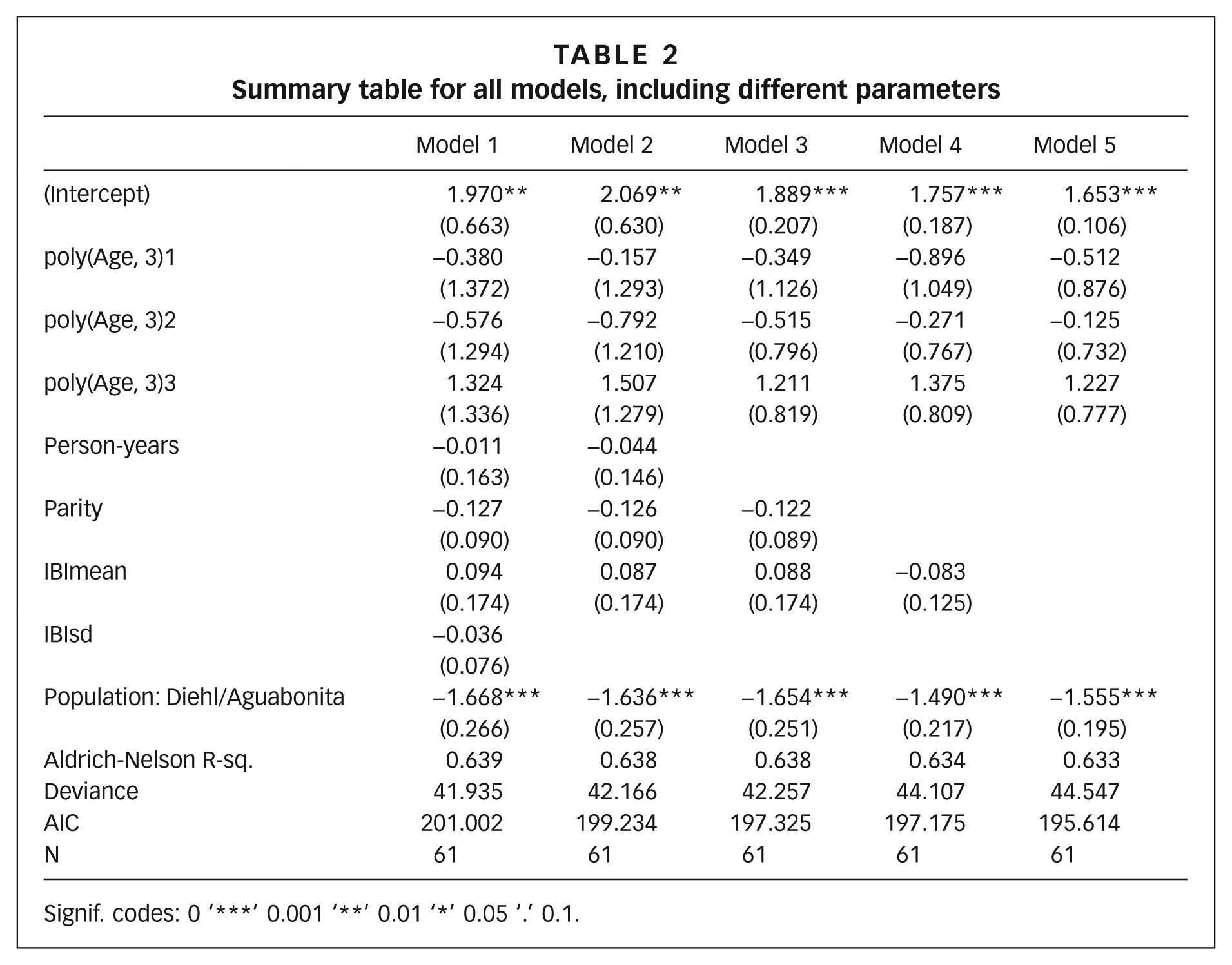
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1.
d. Model verification
To evaluate the generality of this model, we conducted a case study to find the 20 communities of Colombia where malaria was most endemic. We examined malaria case reports for 2008–2010 for 1,123 municipalities. In each municipality, we calculated the density of cases of individuals <5 years of age, for indigenous, Afro-Colombian, and mestizo (majority) populations, separately. We sorted the populations by density of cases <5 years of age, for the 515 populations examined (only populations with more than 10 cases in the <5 age class were considered). Results for the top 20 communities (0.4 per cent) are discussed.
If our forest malarial model is generalizable to other communities, we should observe the following patterns among populations experiencing malaria endemicity:
1) Population size and densities higher than those of typical Amazonian indigenous populations but lower than in large cities (more than 50,000 inhabitants), in locations where the urban population is among the fastest growing in the country;
2) Recent changes in subsistence patterns and increasing energy balance in females;
3) Faster reproduction rates (short IBI values).
Table 3 summarizes the values for the 20 populations where malaria is most endemic, with 18 indigenous and 2 Afro-Colombian individuals. Malaria does not appear to be endemic among the majority population, which is mostly of mixed heritage, and is most endemic in ethnic minorities. Furthermore, wherever a non-indigenous (i.e. Afro-Colombian) population experiences endemicity, the local indigenous population experiences endemicity at higher intensity (Pueblo Rico and Alto Baudó). The indigenous population of San José del Guaviare, where the fieldwork for this study took place, was the third most endemic population of Colombia.
Malaria intensity in the 20 most endemic municipalities and populations of Colombia
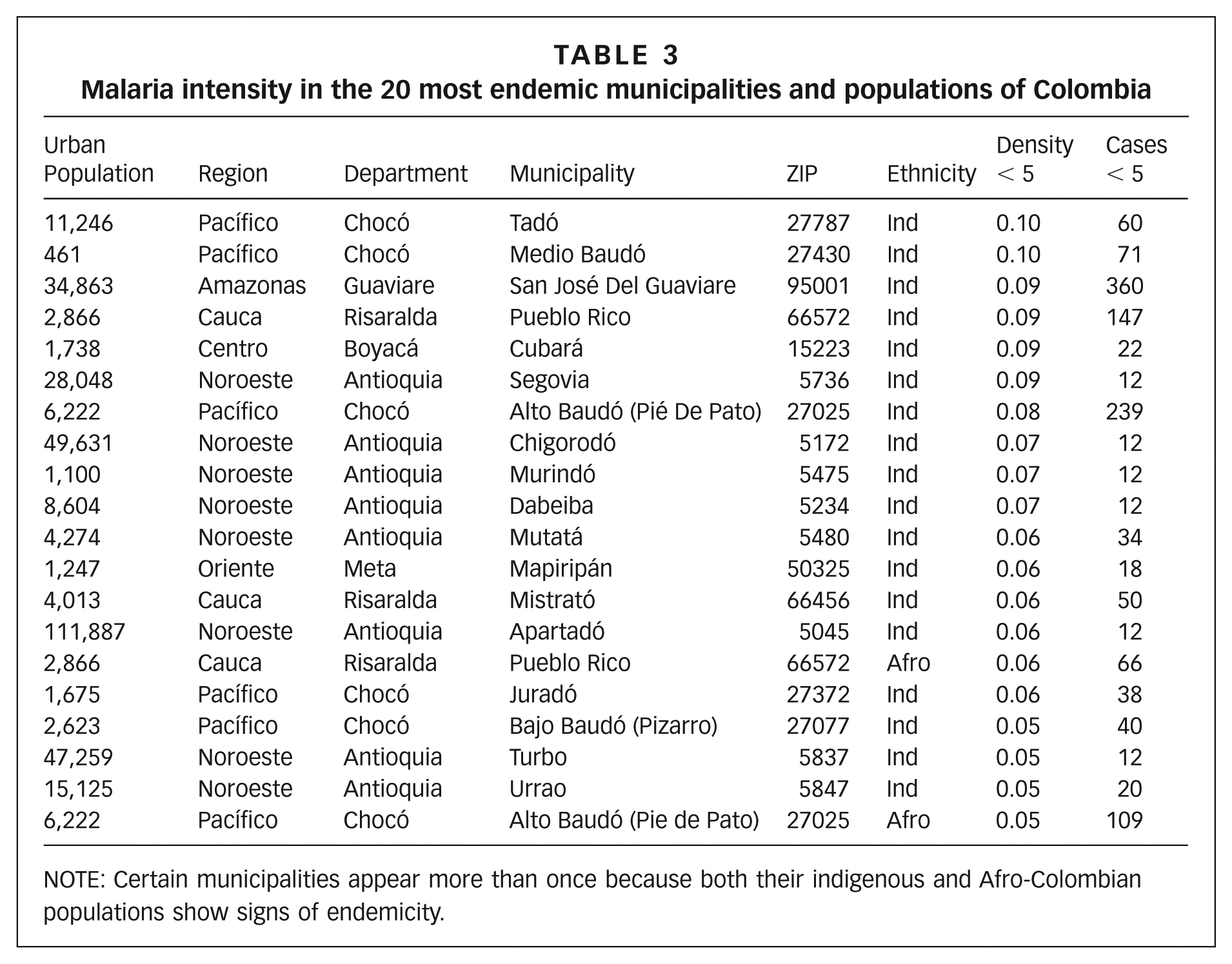
NOTE: Certain municipalities appear more than once because both their indigenous and Afro-Colombian populations show signs of endemicity.
One striking aspect that came to light with this simple analysis applied to the whole country was that the Embera population emerges as one of the most vulnerable communities in Colombia to malaria. They inhabit and outnumber most ethnic minorities in 15 of the 20 most endemic municipalities. They have also been particularly affected by the armed conflict and have experienced forced displacement to urban areas.
Embera inhabited several of the most endemic municipalities of Colombia, and were the most numerous ethnic minority in most cases. Municipalities with Embera populations included Tadó, Alto, Medio and Bajo Baudó, Juradó, Segovia, Mutatá, Chigorodó, Murindó, Dabeiba, Mistrató, Apartadó, Turbo and Urrao. The total population consisted of 71,412 individuals in 2005.(95) Pueblo Rico, in Risaralda, also houses an Embera population, and has experienced malarial endemicity.
The Embera have been subject to extreme violence in the past years. Since 2000, the most violent year experienced by the community, three massacres have terrorized their population and they ranked 6th place in homicides against ethnic minorities in Colombia (55 out of 1,126).(96) In 2009, when our data suggest malaria was endemic, a massive migration of 2,000 people occurred, precipitated by the armed conflict.(97)
Other non-Embera indigenous populations also highlighted by malarial endemicity are the indigenous population of Mapiripán, in Meta, who are mainly Jiw, as well as the population of Cubará in Boyacá, who are mainly U’wa. These two groups have also experienced forced displacement out of their territory.
Most of the populations where malaria is hyperendemic, or where malaria is constantly present at a high incidence, and where malarial infection affects all age groups equally,(98) with the exception of Apartadó, are urban settlements with populations of fewer than 50,000 inhabitants. This is the fastest-growing urban class in Colombia, and Apartadó’s is the single fastest-growing urban population in Colombia since 1993.(99)
V. Conclusions
The most important finding of this study is empirical support for what we believe is one of the first reports of holoendemic, peri-urban malaria in the Americas. Second, we characterize rapidly growing peri-urban locations as hotspots for malarial transmission, with potential holoendemicity, in terms of individual and population-level demographic risk profiles. When malaria is holoendemic, population-level risk factors prevail over individual risk factors. This finding is consistent with epidemiologists stressing the need to use multilevel approaches to study disease patterns, and they can provide numerous insights relevant for public health interventions. Third, we have shown that the effect of migration upon growth of urban populations is twofold: 1) by increasing the number of urban dwellers, and 2) by increasing the fertility rate (for a window of time, probably not permanently) of at least some populations, like the case of the Nükak.
When our research began, local public health officials questioned the relevance of this research by stating that Nükak populations rarely contract the disease. Our findings indicate that this is not only a community where malaria is extremely prevalent, but likely one of the most holoendemic populations in Colombia.
Holoendemicity is associated with population-level behavioural risk factors from a demographic perspective. Contrary to most socioeconomic studies, we find no evidence of household size or fertility rates of females as a risk factor for malaria infection. However, at a population level, we find evidence that malaria prevails where the population has a higher proportion of children and infants. We suggest this is evidence that underlying demographic patterns are crucial for the epidemic characteristics of malaria, whereby a population that increased the reproductive rate of females, by becoming sedentary, crossed a population threshold that enabled the parasite to persist. Naturally, factors such as habitat modification and mobility also play an important role in the persistence of malaria, and further studies of populations living under different conditions of sedentism and anthropogenic habitats will confirm the degree to which combinations of these forces interact to maintain endemicity.
a. Epidemic thresholds of malaria in the Amazon basin
From the epidemiological literature, it is known that a critical fraction of susceptible individuals in a population must be greater than the ratio of the transmission rate and the recovery rate for an epidemic to occur.(100) Similarly, in a non-stationary population, the ratio of rate of entry (in this case, births) and the rate of exit (deaths) must be greater than the critical population of susceptible individuals for the epidemic to persist, if births and death rates are disease independent.
We find evidence that births have increased, without evidence of disease-related deaths. We also detected asymptomatic and endemic malaria among the Nükak. The epidemic parameters affected by demographic transformations described in this study are: First, the birth rate increases in the demographic transition, thus decreasing the threshold parameter for endemicity. Second, the recovery rate is affected by asymptomatic cases, causing adults to remain infective for prolonged periods, thus decreasing the critical threshold parameter for the existence of an epidemic.
We were unable to determine the typical trajectory of the epidemic process from the gathered data and the mentioned literature. We have no evidence of an interaction between demographic dynamics and the pathogen, in the sense that no evidence suggests that population densities or growth were influenced by disease-induced mortality. We suggest that the relationship is the exact opposite, actually – demographic processes being one of the underlying factors driving epidemic change, and not the other way around. The other factor is the change in mobility patterns, which we observe happening in Colombia mainly as a result of forced displacement due to armed conflict.
b. Implications for malarial dynamics
The proposed model of malaria has some practical implications. First, urban expansion and settlement patterns in the Amazon basin coupled with deforestation(101) are cause for concern in the context of epidemics, and particularly for malaria, especially after outbreaks resulting from public policies of migration to the Amazon region.(102) Those migrations, and how the colonization process of the Amazon rainforest continues, will likely influence malaria dynamics. If malaria intensity varies as a response to both population size and the proportion of susceptible individuals in the population, and is more abundant where forest is cleared for various human uses, the colonization process will make Amazonian populations more vulnerable to malaria each day.
Our findings are consistent with the hypothesis laid down by Reichel-Dolmatoff(103) and by Durham,(104) which suggests that demographic patterns are an adaptation to reduce pathogen load in Amazonian populations. We have documented the impact of recent transformations of Amazonian populations, moving towards urban settlements, on malaria epidemics. Both Reichel-Dolmatoff and Durham suggest that by living in dispersed conditions and with slow life histories (e.g. late reproductive age), Amazonian populations lived below an epidemic threshold. Our study showed that by increasing the reproductive rate, and settling in more dense, peri-urban locations, indigenous groups such as the Nükak are altering their conditions to produce malarial endemicity.
Our study has potential implications for malarial control policies. Mainly, our findings suggest that the intensity of malaria can be affected by demographic dynamics. Phase transitions of endemicity and life history are likely occurring simultaneously in indigenous Amazonian populations. The presence of asymptomatic malaria in adults, as well as faster reproduction, alters the epidemic conditions by affecting the fraction of susceptible (e.g. children with naïve immune systems) and infected persons (e.g. asymptomatic adults who are infective for prolonged periods of time).(105) Thus, malaria control strategies for populations experiencing endemicity should include components of reproductive health and treatment of adult asymptomatic individuals, particularly among the fastest-growing urban populations.
Supplemental Material
feged-rivadeneira-supplement – Supplemental material for Demographic and epidemic transitions in peri-urban areas of Colombia: a multilevel study of malaria in the Amazonian city of San José del Guaviare
Supplemental material, feged-rivadeneira-supplement for Demographic and epidemic transitions in peri-urban areas of Colombia: a multilevel study of malaria in the Amazonian city of San José del Guaviare by Alejandro Feged-Rivadeneira, Carlos Del Cairo and William Vargas in Environment & Urbanization
Footnotes
Acknowledgements
The authors would like to thank Stanford University, the Zaffaroni family, and the Morrison Institute for their financial support. We would also like to thank Alison Maassen and Ariel Domlyn for their contributions as research assistants in San José del Guaviare, in particular Alison’s contributions to understanding Nükak family trees during the first phase of ethnographic work. We thank Erik Vergel-Tovar for his comments on a draft of this manuscript. Most importantly, we are grateful to the Nükak people for receiving us in their community and sharing their time with us. We hope this document will contribute to the understanding and mitigation of their hardships.
6.
8.
10.
14.
28.
33.
39.
41.
50.
59.
72.
73.
Conducted by Browder and Godfrey (1990, 1997); and ![]() .
.
75.
80.
81.
86.
88.
Diehl (1992, ![]() ).
).
89.
Diehl (1992, ![]() ).
).
90.
Diehl (1992, ![]() ).
).
91.
Diehl (1992, ![]() ).
).
92.
93.
94.
Diehl (1992, ![]() ).
).
105.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.