
Abstract
Circadian rhythms enable organisms to mediate their molecular and physiological processes with changes in their environment. Although feeding behavior directly affects within-organism processes, there are few examples of a circadian rhythm in this key behavior. Here, we show that Daphnia have a nocturnal circadian rhythm in feeding behavior that corresponds with their diel vertical migration (DVM), an important life history strategy for predator and UV avoidance. In addition, this feeding rhythm appears to be temperature compensated, which suggests that feeding behavior is robust to seasonal changes in water temperature. A circadian rhythm in feeding behavior can impact energetically demanding processes like metabolism and immunity, which may have drastic effects on susceptibility to disease, starvation risk, and ultimately, fitness.
Biological rhythms enable organisms to coordinate their molecular and physiological processes with the daily and seasonal changes that occur in the environment. This coordination is well understood in organisms that undergo dormancy and in plant leaves that reposition throughout the day (Ruby, 2003; McClung, 2006). Biological rhythms that coordinate daily (24 h) processes are called circadian rhythms. For example, the vertebrate and invertebrate immune systems show circadian expression (Scheiermann et al., 2013; Curtis et al., 2014; Rund et al., 2016; Baxter and Ray, 2020). Behavioral traits often mediate many molecular and physiological processes; for example, metabolic rates are linked with foraging behavior (Biro and Stamps, 2010). As such, circadian rhythms in behavioral traits like foraging could affect or even drive circadian rhythms in other traits.
Circadian (daily) rhythms are maintained by endogenous clocks that cycle with a period around 24 h in length. Circadian rhythms are characterized by 3 main components: (1) they are entrained to environmental cues (or zeitgebers); (2) they oscillate under constant conditions, that is, if zeitgebers vary or are missing on a particular day, the rhythm will persist; and (3) the period of a circadian rhythm does not vary significantly with temperature fluctuations (Sorek and Levy, 2012). The most prominent environmental stimuli used to entrain these rhythms are photoperiod, light-dark cycles (Foster and Helfrich-Forster, 2001; Stelzer et al., 2010), but other environmental stimuli are also used as zeitgebers (temperature: Glaser and Stanewsky, 2005, 2007; Lahiri et al., 2005; species interactions: Levine et al., 2002, 2004).
Here, we evaluate whether there is a circadian rhythm in feeding behavior in Daphnia dentifera, a dominant primary consumer in North American temperate lakes. Prior studies have indicated that Daphnia have core clock genes and circadian expression of melatonin, immune, and sensory genes (Cellier-Michel et al., 2003; Bentkowski et al., 2010; Bernatowicz et al., 2016; Schwarzenberger and Wacker, 2015; Schwarzenberger et al., 2020, 2021). Daphnia are a model system for studying circadian rhythms in feeding behavior because they reproduce clonally, making individuals genetically identical, and quantifying individual feeding rates is relatively simple (Hite et al., 2020). In their natural habitat, they experience many daily fluctuations that could be used as environmental stimuli for entraining a circadian rhythm. For example, many species of Daphnia have a well-documented rhythm in locomotor behavior (diel/diurnal vertical migration, DVM; Haney and Hall, 1975) where individuals spend the day at the bottom of the water column and migrate to the top at dusk. This diurnal pattern of migration is hypothesized to be a defense strategy against UV (Leech and Williamson, 2000), fluctuations in temperature (Kessler and Lampert, 2004; Cooke et al., 2008), and predation (Dawidowicz and Loose, 1992). In addition, DVM has been linked to differences in feeding rate (Haney and Hall, 1975; Makinol et al., 1996). These daily environmental changes, which entrain DVM, may entrain a circadian rhythm in feeding behavior in Daphnia that could have downstream effects on their metabolic and immune functions. Here we show, under laboratory conditions, that D. dentifera have a robust nocturnal circadian rhythm in feeding behavior consistent with the DVM observed in Daphnia in natural systems.
Materials and Methods
We used a standard genotype of D. dentifera, originally collected in Southern Michigan (USA), that has been studied extensively in other contexts (Auld et al., 2012; Clay et al., 2019). Daphnia were fed the green alga Ankistrodesmus falcatus, batch-cultured in 500 ml of high nitrogen COMBO under constant light at 22 °C (Kilham et al., 1998). Prior to the start of the standardization protocol for D. dentifera, we froze aliquots (to provide 1 mgC/l of food per day) of A. falcatus from one 14-day-old algal culture to ensure a consistent chlorophyll:carbon ratio for the course of the experiment.
Prior to the experiment, we raised individual clonal D. dentifera in vials of 15-ml COMBO medium (Kilham et al., 1998) and fed daily aliquots of A. falcatus. We maintained D. dentifera under a 15:9 h light-dark photoperiod at 22 °C for 3 generations to standardize any potential maternal effects. Experimental animals were taken from the third clutches of third-generation standardized mothers on 5 consecutive days so that, at the outset of the circadian rhythm measurement, we had animals of 5 different ages (between 6 and 10 days old); on each day, all experimental animals were born within a single 24-h period to ensure that they were all the same age at the start of the experiment. At birth, animals were assigned to 1 of the 2 temperature treatments (16 or 22 °C). For each temperature, the experimental generation contained twelve 6-, 7-, 8-, 9-, and 10-day old individuals (n = 120). Using genetically identical individuals of different ages allowed us to run all feeding rate assays in one 24-h period, decreasing experimental error related to temperature, light, and machine fluctuations that can occur when experiments are run over multiple days.
To determine whether feeding rates were cued by the presence/absence of light, 9- and 10-day-old individuals experienced 24-h darkness during the feeding rate assay. Ten-day-old individuals were maintained in 24-h darkness for an additional day prior to the feeding assay to determine whether feeding rate patterns persisted without light cues. To quantify the feeding rate, we isolated individuals in 12-well plates (CELLTREAT, Pepperell, USA) in 5 ml of COMBO media with 1 mgC/l of A. falcatus. Individuals fed for 3 h, after which they were moved to a new well plate for the next feeding rate assay. At the end of each 3-h assay, we measured fluorescence on a plate reader (Tecan, Maennedorf, Switzerland), following Hite et al., 2020, to compare the relative quantities of chlorophyll-a in wells with D. dentifera to control wells that did not contain D. dentifera. We calculated individual feeding rates as:
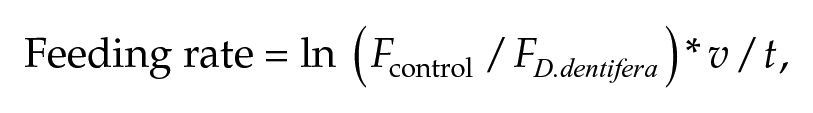
where Fcontrol is the average fluorescence of control wells, FD.dentifera is the fluorescence of an animal well, v is the volume of COMBO and algae in milliliters, and t is the time D. dentifera fed in hours (Sarnelle and Wilson, 2008). We measured 8 consecutive 3-h feeding rates (24 h) for individuals in each age group to assess the rhythmicity of D. dentifera feeding behavior.
Rather than measuring the feeding rate of a group of individuals across 5 consecutive days, we measured the feeding rate of genetically identical individuals of 5 different ages (6-10 days old) across a single 24-h period at 22 and 16 °C. To generate the data for analysis, we randomly chose 1 of the 12 individuals for each of the 5 days and concatenated those feeding rate measurements to construct a single 5-day feeding rate trajectory. We then analyzed the period and amplitude of feeding behavior across 12 randomly assembled trajectories. To generate bootstrap estimates for period and amplitude, we repeated this process 100 times. We verified that our protocol had similar period estimates and standard errors to what we observed when tracking the same individuals across 4 consecutive days (see Suppl. Fig. S1). To estimate the period and amplitude, we implemented MFourFit and Maximum Entropy Spectral Analysis (MESA) procedures with linear detrending in Biodare2 (Zielinski et al., 2014). Briefly, MFourFit is a curve-fitting method that assumes a single period and returns the best-fitting waveform for each cycle. MFourFit accounts for fit error and model complexity when testing periods and gives preference to simpler models (Edwards et al., 2010). Whereas MESA uses a stochastic modeling approach that assumes each value is a combination of the previous time point(s) value(s) and noise. This method uses a frequency spectrum to identify the strongest period within the data (Burg, 1972). To confirm whether the feeding rate rhythm was diurnal or nocturnal, we ran an analysis of covariance (ANCOVA) with phase (day: 0-16 ZT and night: 16-24 ZT) as a predictor and age as a covariate for each temperature treatment separately (R Core Team, 2021).
Results
At 22 °C, D. dentifera show a clear circadian rhythm of feeding behavior with a period around 24 h, which is shown across multiple ages and persists without light cues (Figure 1); 9- and 10-day-old individuals exhibited the same rhythm in feeding behavior under 24 and 48 h of constant darkness, respectively, indicating that the rhythm is endogenous and can free-run in the absence of light cues. At 16 °C, there is a similar overall pattern in feeding behavior across the 5 ages which persists without light cues (Figure 2), indicating that feeding is temperature compensated.
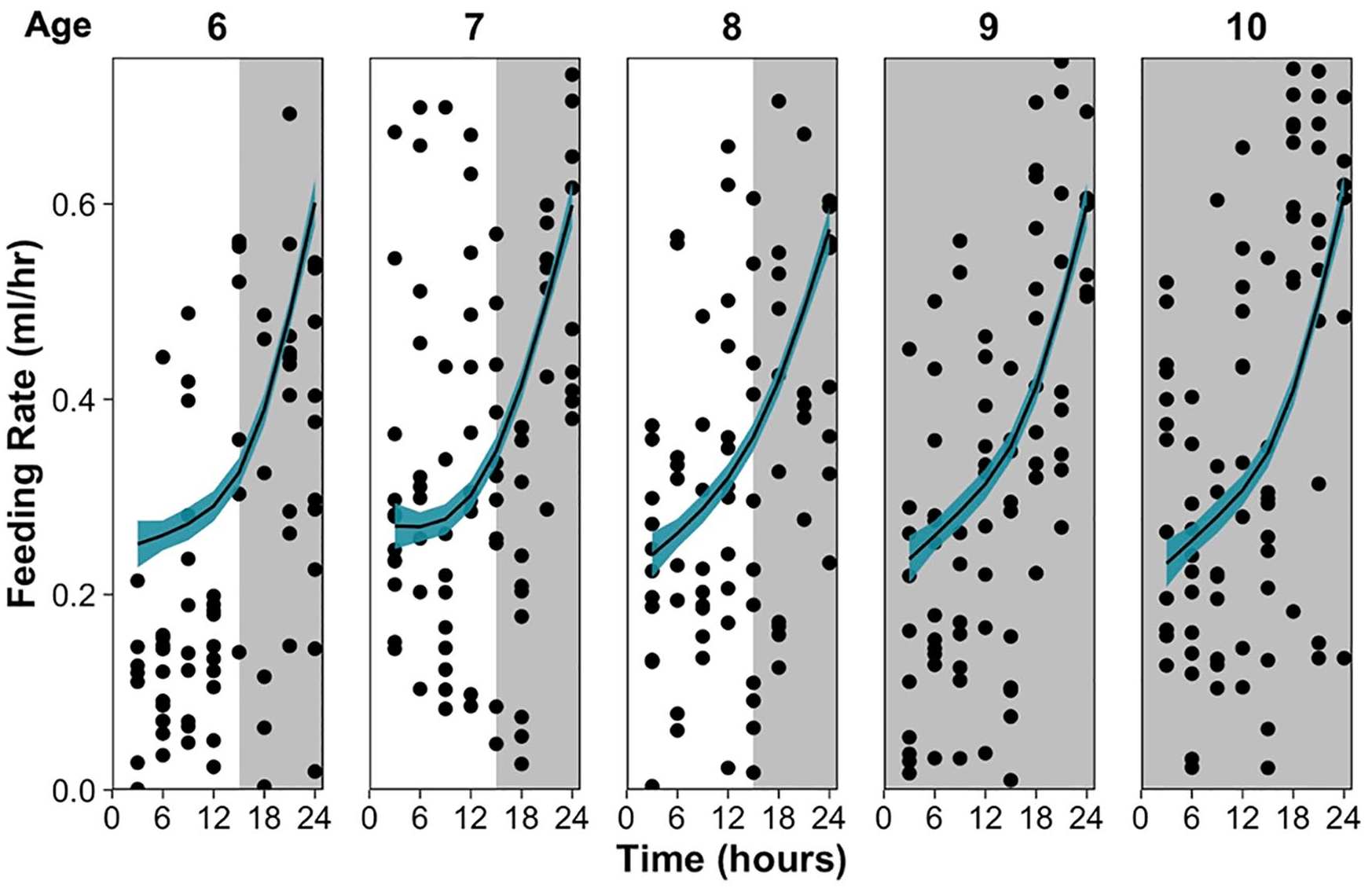
Feeding rate (ml/h) every 3 hours a 24-h period for individuals at ages 6-10 (from left to right) at 22°C. Plot shading indicates the light-dark cycle, with 6 - to 8-day-old individuals under a 15:9 light-dark cycle, 9-day-old individuals under 24-h darkness, and 10-day-old individuals under their second day of 24-h darkness. Points indicate individual feeding rates and the line is a generalized additive model fit per age with line shading indicating the 95% confidence intervals.
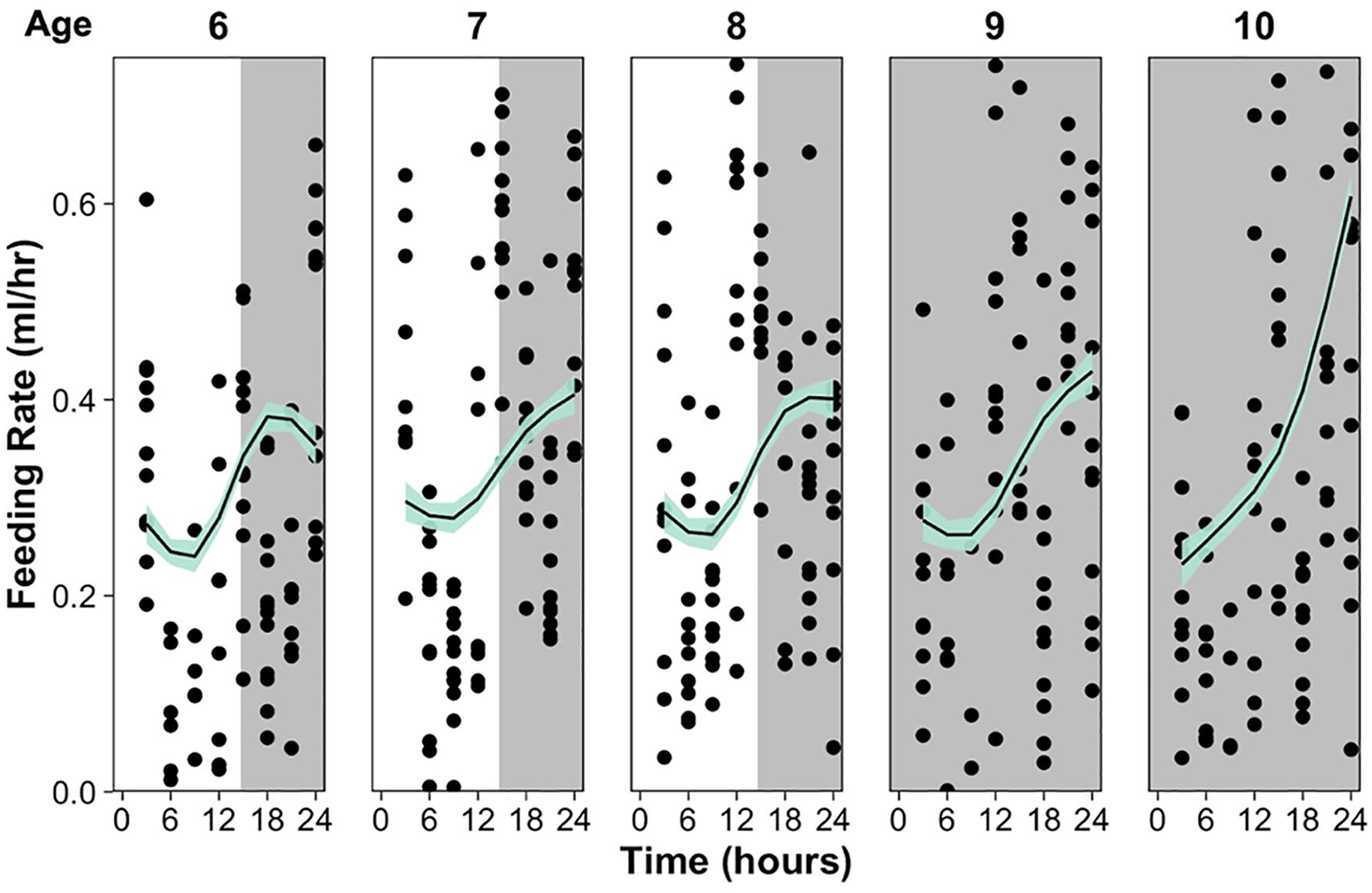
Feeding rate (ml/h) every 3 hours a 24-h period for individuals at ages 6-10 (from left to right) at 16°C. Plot shading indicates the light-dark cycle, with 6 - to 8-day-old individuals under a 15:9 light-dark cycle, 9-day-old individuals under 24-h darkness, and 10-day-old individuals under their second day of 24-h darkness. Points indicate individual feeding rates and the line is a generalized additive model fit per age with line shading indicating the 95% confidence intervals.
At both 22 and 16 °C, the feeding rhythm in D. dentifera has a period of about 24 h. Using the MESA approach, at 22 °C, the period is 23.8 ± 0.1 h and at 16 °C, it is 22.6 ± 0.1 h (Figure 3). Similarly, the MFourFit approach estimated a period of 24.2 ± 0.1 h at 22 °C and 23.9 ± 0.2 h at 16 °C (Figure 3). However, the amplitude (the height of the rhythm or the daily fluctuation in feeding rates) of the feeding rate rhythm was larger at warmer temperatures. For both approaches, the amplitude of the feeding rate rhythm was 0.06 ml/h lower at 16 °C compared to 22 °C (Figure 3), meaning the difference in feeding rates between the peak and the trough of the rhythm is much smaller at colder temperatures. In general, these results suggest that the circadian rhythm in D. denitera feeding rate is temperature compensated. Temperature compensation is likely very important in aquatic individuals’ circadian rhythms as water temperatures may fluctuate throughout the day and vary across seasons. For instance, the temperate freshwater lakes that D. dentifera inhabits have been shown to vary up to 3 °C within 14 days (Shocket et al., 2018).
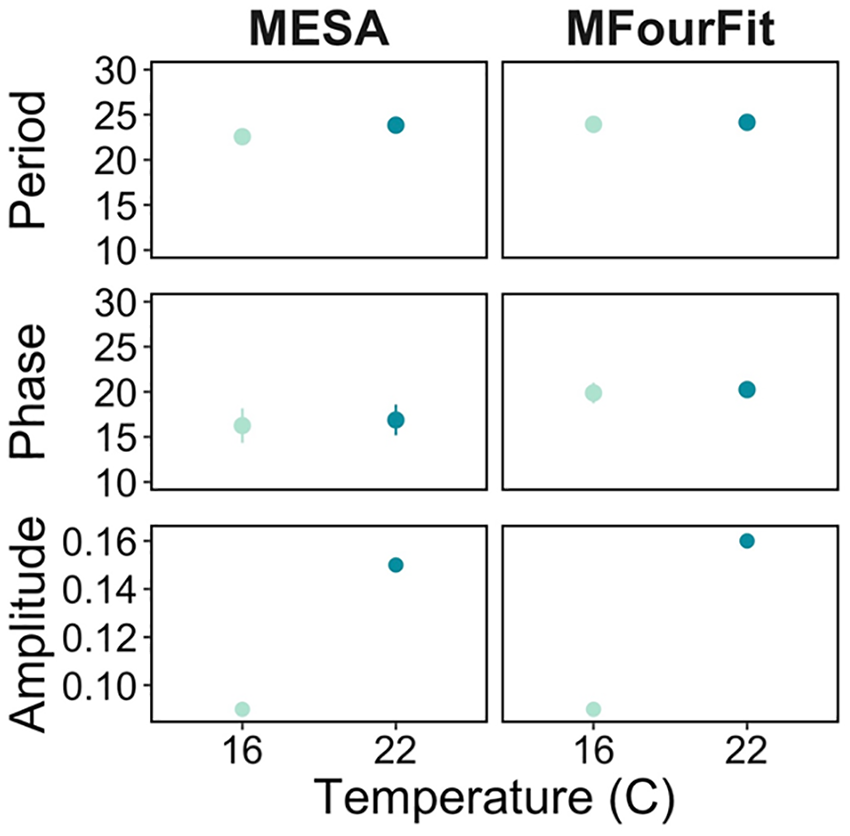
Period, phase, and amplitude estimates from Biodare2 analyses for the MESA and MFourFit methods expressed in hours with confidence intervals at 95% for both temperatures (16 and 22°C) of the study.Abbreviation: MESA = Maximum Entropy Spectral Analysis.
In Figures 1 and 2, it appears that D. dentifera have higher feeding rates at night compared to the day. To confirm that the circadian rhythm in feeding was nocturnal, we fit an ANCOVA with phase (day: 0-16 ZT and night:16-24 ZT) as a predictor and age as a covariate for each temperature treatment. At both temperatures, D. dentifera ate significantly more during the night (in the presence and absence of light cues) compared to the day, ANCOVA, 16 °C: F3,944 = 43.76, p < 0.001; 22 °C: F3,941 = 43.54, p < 0.001.
Discussion
The observed circadian pattern in feeding behavior observed here, with higher feeding rate at night, matches the pattern of DVM that Daphnia undergo in nature, suggesting that there is an association between swimming behavior and feeding. Given that Daphnia consume phytoplankton that are mostly found in the upper regions of the water column, it follows that as they migrate to the top of the water column at night, they also consume more food. This circadian rhythm in feeding behavior could affect metabolic and immune pathways which are energetically demanding processes, that is, at night when feeding and active levels are high might also be when metabolic and immune pathways are upregulated. In addition, times of high feeding and activity (night) are likely correlated with times of high exposure to pathogens, particularly food-borne pathogens. The interplay between a circadian rhythm in feeding that can affect an individual’s exposure to pathogens and their immune functioning is difficult to discern.
This is the first study, to our knowledge, to determine a circadian rhythm in feeding behavior in the keystone aquatic herbivore Daphnia. The nocturnal feeding rhythm shown in D. dentifera is temperature compensated and is consistent with the well-studied DVM behavior of many zooplanktons. Circadian rhythms in Daphnia have been shown to impact behavior and life history traits (Coldsnow et al., 2017; Gust et al., 2019; Schwarzenberger et al., 2020). The observed circadian rhythm in feeding behavior could alter the timing of energetically demanding processes like metabolism and immunity. Understanding these effects should be a major focus of future research, given the importance of Daphnia as a model organism in fields like ecotoxicology (Shaw et al., 2008), limnology (Seda and Petrusek, 2011), population ecology (Lampert, 2006), and disease ecology (Wale and Duffy, 2021).
Supplemental Material
sj-docx-1-jbr-10.1177_07487304211054404 – Supplemental material for Circadian Rhythm in Feeding Behavior of Daphnia dentifera
Supplemental material, sj-docx-1-jbr-10.1177_07487304211054404 for Circadian Rhythm in Feeding Behavior of Daphnia dentifera by Alaina C. Pfenning-Butterworth, Kristina Amato and Clayton E. Cressler in Journal of Biological Rhythms
Footnotes
Acknowledgements
We would like to thank Dr. Jessica L. Hite, University of Wisconsin-Madison, for assistance during pilot experiments and David Nguyen, University of Nebraska-Lincoln, for his advice in the statistical analysis. The authors received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.