
Abstract
The suprachiasmatic nucleus (SCN) drives circadian rhythms in locomotion through coupled, single-cell oscillations. Global genetic deletion of the neuropeptide Vip or its receptor Vipr2 results in profound deficits in daily synchrony among SCN cells and daily rhythms in locomotor behavior and glucocorticoid secretion. To test whether this phenotype depends on vasoactive intestinal polypeptide (VIP) neurons in the SCN, we ablated VIP SCN neurons in vivo in adult male mice through Caspase3-mediated induction of the apoptotic pathway in cre-expressing VIP neurons. We found that ablation of VIP SCN neurons in adult mice caused a phenotype distinct from Vip- and Vipr2-null mice. Mice lacking VIP neurons retained rhythmic locomotor activity with a shortened circadian period, more variable onsets, and decreased duration of daily activity. Circadian hormonal outputs, specifically corticosterone rhythms, were severely dampened. In contrast, deletion of neonatal SCN VIP neurons dramatically reduced circadian gene expression in the cultured SCN, mimicking the effects of global deletion of Vip or Vipr2. These results suggest that SCN VIP neurons play a role in lengthening circadian period and stimulating the daily surge in glucocorticoids in adults and in synchronizing and sustaining daily rhythms among cells in the developing SCN.
Located in the ventral hypothalamus, the suprachiasmatic nucleus (SCN) is the dominant pacemaker that aligns daily physiological and behavioral rhythms to the local light-dark schedule (Coomans et al., 2015; Hastings et al., 2018; Herzog, 2007; Welsh et al., 2010). Individual SCN neurons express cell autonomous oscillations driven through a negative transcription-translation feedback loop involving the core clock genes, which include Bmal1, Clock, Period 1 and 2, and Cryptochrome 1 and 2 (Takahashi, 2017; Webb et al., 2009; Welsh et al., 1995). Intercellular neurotransmission is necessary to couple single-cell oscillators and generate high-amplitude, precise, circuit-wide rhythms capable of driving behavioral circadian rhythms (Herzog et al., 2004; Tokuda et al., 2018; Welsh et al., 1995; Yamaguchi et al., 2003).
Vasoactive intestinal polypeptide (VIP) is sparsely expressed throughout the mammalian brain in the cortex, retina, superior colliculus, and SCN. Released by 10% of SCN neurons, VIP is necessary for the synchronization of single-cell circadian rhythms (Abrahamson and Moore, 2001; Aton et al., 2005). Mice lacking Vip or its receptor Vipr2 show an advanced phase angle of entrainment in a light cycle, and approximately 60% lose daily locomotor rhythms or exhibit multiple periodicities in constant darkness (Aton et al., 2005; Colwell et al., 2003; Cutler et al., 2003; Harmar AJ et al., 2002). Vip-null mice also lose rhythmicity in hormonal circadian outputs, for example, the daily rhythm in corticosterone (Loh et al., 2008). In vitro, SCN slices lacking VIP signaling (either through global genetic deletion of Vip or Vipr2 or pharmacologic blockade) show decreases in the number of rhythmic SCN neurons and loss of intercellular synchrony (Aton et al., 2005; Brown et al., 2007; Maywood et al., 2006). These results have led to the conclusion that VIP signaling is required to synchronize circadian cells and amplify daily rhythms in physiology and behavior. The relative roles of VIP neurons in the SCN, neocortex, and other brain areas in the development and sustenance of circadian rhythms have not, however, been fully clarified.
Furthermore, mice that lack Vip or Vipr2 display phenotypic heterogeneity in their circadian profile (only 60% of mice show behavioral arrhythmicity). This suggests that other non-VIP signaling pathways compensate for the loss of VIP signaling (Brown, 2005; Maywood et al., 2011). In Drosophila, the genetic knockout of pigment-dispersing factor (PDF; the functional homologue of VIP) yields a similarly disrupted circadian phenotype to the ablation of PDF-expressing neurons (Renn et al., 1999). But cell ablation and genetic knockout need not produce identical phenotypes. For example, deletion of the Opn4 (melanopsin) gene has mild effects on circadian entrainment compared with the complete loss of photic entrainment following ablation of Opn4-expressing intrinsically photosensitive retinal ganglion cells (Güler et al., 2008; Hattar, 2002). Therefore, we selectively ablated VIP SCN neurons in vivo and in vitro to test for their necessity in the development and adulthood for circadian rhythms in locomotor activity, glucocorticoid release, and gene expression.
Materials and Methods
Animals
Male mice were housed in a 12h:12 h light:dark cycle in the temperature- and humidity-controlled Danforth Animal Facility at Washington University in St. Louis with ad lib access to food and water. Combinations of the following genotypes were used in all experiments: VIP-IRES-Cre knock-in mice (VIPtm1(creZjh), Jackson Laboratories, RRID: IMSR_Jax:010908), Per2::Luciferase knock-in mice (founders generously provided by Dr. Joseph Takahashi, UTSW), tdTomato reporter mice (B6.Cg-Gt(ROSA)26Sortm9(CAG-tdTomato)Hze/J, Jackson Laboratories, #007909), Vip-/- (founders generously provided by Drs. Christopher Colwell and James Waschek, University of California, Los Angeles), Vipr2-/- (founders generously provided by Drs. Anthony Harmer and Michael Hastings, MRC Cambridge), and C57Bl/6JN mice. All procedures were approved by the Animal Care and Use Committee of Washington University and adhered to National Institutes of Health guidelines.
In Vivo Deletion of VIP Neurons
pAAV-flex-taCasp3-TEVp plasmid was obtained from Addgene (gift from Nirao Shah & Jim Wells, Addgene plasmid #45580, http://n2t.net/addgene:45580; RRID:Addgene_45580) and packaged into a high-titer AAV8 virus by the Washington University Viral Core. To ablate VIP SCN neurons in vivo, 6- to 10-week-old VIP-IRES-Cre heterozygous mice were injected with AAV8-Casp3-TEVP virus targeted to the SCN. Briefly, anesthetized mice (2% isofluorane) were placed into a stereotactic device, and the virus (0.5 µL/side) was injected bilaterally into the SCN with a 30-gauge Neuros syringe at +0.4 mm anterior, ±0.15 mm lateral, and 5.6 mm ventral to the bregma. Age-matched control mice were either VIP-IRES-Cre heterozygotes receiving artificial cerebrospinal fluid (ACSF; containing in mM: NaCl, 125; KCl, 2.5; NaH2PO4, 1.25; NaHCO3, 25; CaCl2, 2; MgCl2, 1; dextrose, 25-310 mOsmol/L) injections or mice lacking Cre expression (C57Bl6/JN or PER2::Luc/+) receiving AAV8-Caspase3-TEVP injections. Mice received analgesic treatment during recovery from surgery. At the end of the experiment, histologic examination of all mice revealed minimal damage to the bilateral paraventricular nucleus (PVN) and no damage to the SCN in control mice.
Locomotor Activity
After recovery, mice were placed in custom-built cages to monitor running-wheel activity (Clocklab, Actimetrics, Evanston, IL) in 12h:12 h light:dark for at least 7 days and in constant darkness for at least 14 days. Experiments were run on 4 separate cohorts of mice. Locomotor activity was analyzed from at least 1 week of data using Clocklab software (Actimetrics) and custom-written Python scripts. Circadian period, amplitude, and rhythmicity were calculated using chi-squared periodogram. A 8-h box filter was used to identify daily activity onsets and offsets to calculate cycle-to-cycle onset variability and the duration of daily activity. Custom Python scripts were used to quantify total daily activity counts. All data were blinded to treatment and genotype before analysis.
Circadian Corticosterone Analysis
Following locomotor activity recordings, a randomly selected subset of the mice exhibiting the range of locomotor phenotypes were transferred to custom-built cages for fecal matter collection. These cages consisted of a wire stage in which mice could comfortably sit with easy access to ad lib food and water. Most feces and urine fell through the wire stage onto a piece of filter paper at the bottom of the cage. A slot was cut out of the bottom front of the cage that allowed an investigator to easily collect and replace the filter paper. We estimate that 85% to 100% of feces per time point were collected. After mice habituated to the cages in constant darkness for 2 days, investigators collected fecal samples in either 3- or 4-h time bins for 48 to 72 h. Samples were immediately transferred to −80 °C. For steroid extraction, samples were baked at 50 °C for 3 to 4 h until completely dry before being individually ground into a powder using a mortar and pestle. Total weight per time point was calculated, and steroids were extracted from 25 mg of powdered fecal matter using 80% methanol. After agitation, the supernatant was transferred to new tubes, and methanol was evaporated inside a fume hood. The pellets were suspended in 500 µL of enzme-linked immunosorbent assay (ELISA) buffer and diluted to a concentration of 1:2500 in ELISA buffer. Samples were processed in duplicate for corticosterone concentration using the ELISA kit instructions (corticosterone EIA, Cayman Chemicals, Ann Arbor, MI). Final concentrations (in ng per mg of feces) were determined based on the standard curve, dilution factor, and total fecal weight per time point. There were no measurable differences in fecal weight between VIP neuron–ablated (VIPN) and control mice. Corticosterone collection and measurement were completed on 3 separate cohorts of mice.
Immunohistochemistry
After locomotor and corticosterone assessment, mice were reentrained to a 12 h:12 h light:dark cycle and then perfused during late day (zeitgeber time 7-10). Briefly, mice were anesthetized with 1.25% Avertin (2,2,2-tribromoethanol and tert-amyl alcohol in 0.9% NaCl; 0.025 mL/g body weight) and transcardially perfused with phosphate-buffered saline (PBS) and 4% paraformaldehyde. The brain was dissected and transferred to 30% sucrose the following day. Frozen coronal sections were cut at 40 µm on a cryostat (CM1850, Leica, Wetzlar, Germany) and placed in 3 separate wells. For VIP and arginine-vasopressin (AVP) immunofluorescence, free-floating sections were washed for 1 h in PBS and then blocked in PBSGT (5% normal goat serum, 0.3% Triton and PBS) for 1 h. Sections were incubated overnight in primary antibodies diluted in PBSGT (rabbit anti-VIP 1:2000, Immunostar [Hudson, WI], and mouse anti-AVP PS41 1:100, generous gift of Dr. Hal Gainer, National Institutes of Health). Slices were washed again and incubated for 2 h at room temperature with secondary antibodies (anti-rabbit 488 and anti-mouse 564, 1:500). Sections were briefly stained with DAPI, washed again in PBS, mounted, and cover-slipped with DABCO (1,4-diazobicycol[2,2,2]-octane) mounting medium. All sections from a specific cohort were simultaneously processed. Sections were imaged on an epifluorescent microscope (TE2000 inverted, Nikon, Tokyo, Japan). The SCN was identified using the DAPI staining, resulting in 2 to 5 SCN sections per animal. Images of VIP and AVP immunofluorescence were acquired using the same exposure and gain for every mouse in each round, allowing us to directly compare staining intensities. An investigator, blinded to the genotype and treatment of the mouse, drew boundaries around the SCN using ImageJ software and measured the mean intensity of VIP and AVP staining within those boundaries for each SCN section. Background staining for VIP and AVP was calculated as the mean intensity for each image from an area slightly lateral to the SCN that had no visible AVP or VIP labeling. To calculate the relative staining intensity of VIP or AVP, the respective background staining was subtracted from the SCN VIP or AVP mean staining intensity. Values reported are the highest relative staining intensities for an individual mouse. A subset (n = 7) of mice that received in vivo Cre-dependent caspase3 injections expressed tdTOMATO in VIP neurons. Instead of immunohistochemistry, these mice were sectioned and imaged using fluorescence microscopy. Of these 7 mice, 1 had no tdTOMATO expression indicating ablation of VIPNs, 3 retained VIP expression (VIPN present), and 3 lacked Vip-driven Cre expression (C57Bl/6JN controls). These 7 mice were included in Figures 1 and 2, but excluded from Figure 5.
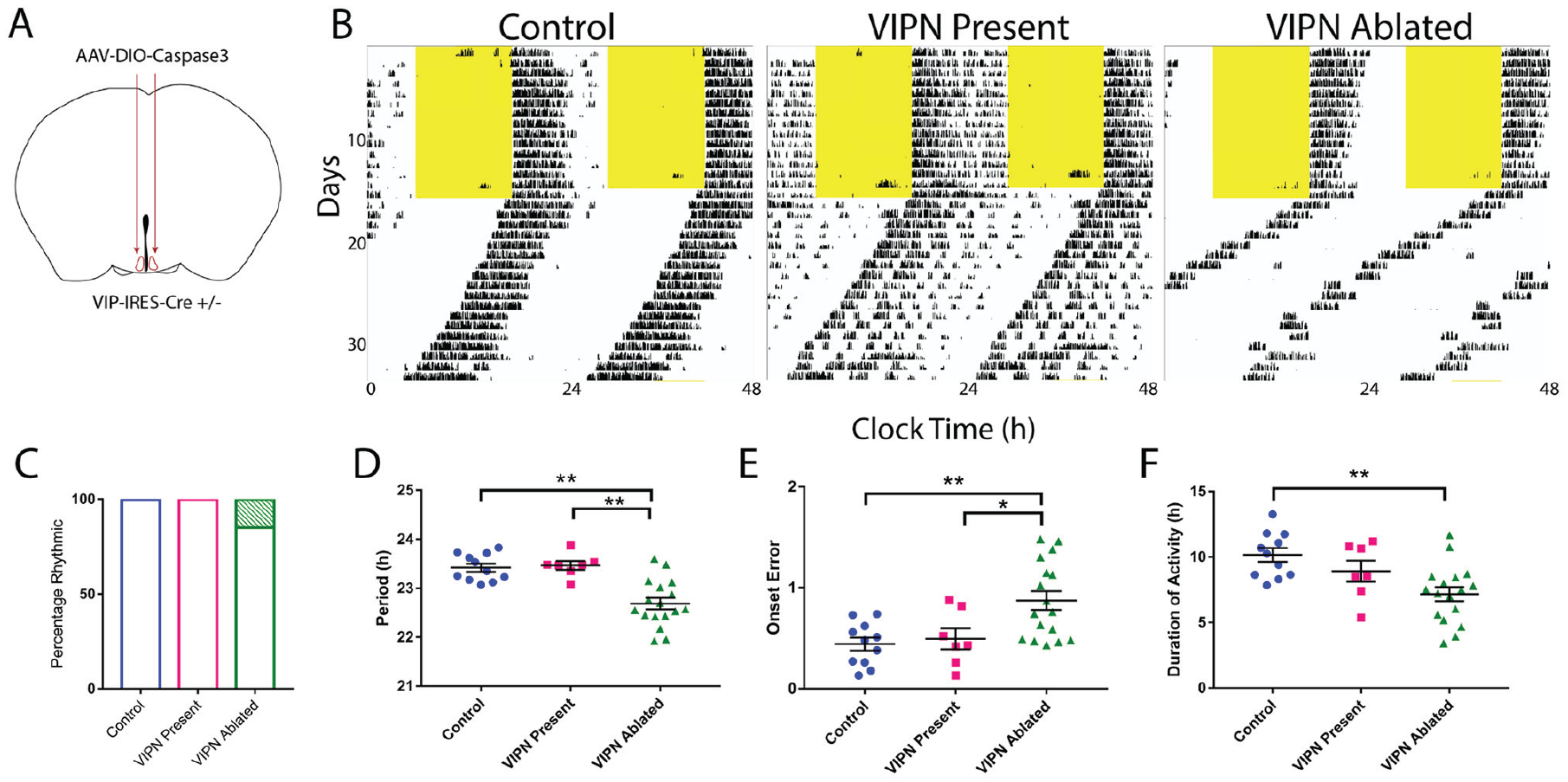
Ablation of SCN vasoactive intestinal polypeptide (VIPN) neurons in adult mice shortens the duration and period of daily locomotor activity. (A) Schematic showing injection of Cre-dependent Caspase3 virus bilaterally into the SCN of adult VIP-Cre heterozygous mice. (B) Representative actograms of locomotor activity in a light:dark cycle (LD) and constant darkness (DD) for the following 3 groups: control, VIP neuron (VIPN) present, and VIPN ablated. (C) Ablation of VIP SCN neurons had little effect on the percentage of circadian mice (3/20 VIPN ablated mice were arrhythmic compared with 0/11 control and 0/7 VIPN present mice). (D) Ablation of VIP SCN neurons shortened the circadian period, (E) increased onset variability, (F) and reduced the duration of daily activity. Error bars show mean ± SEM.
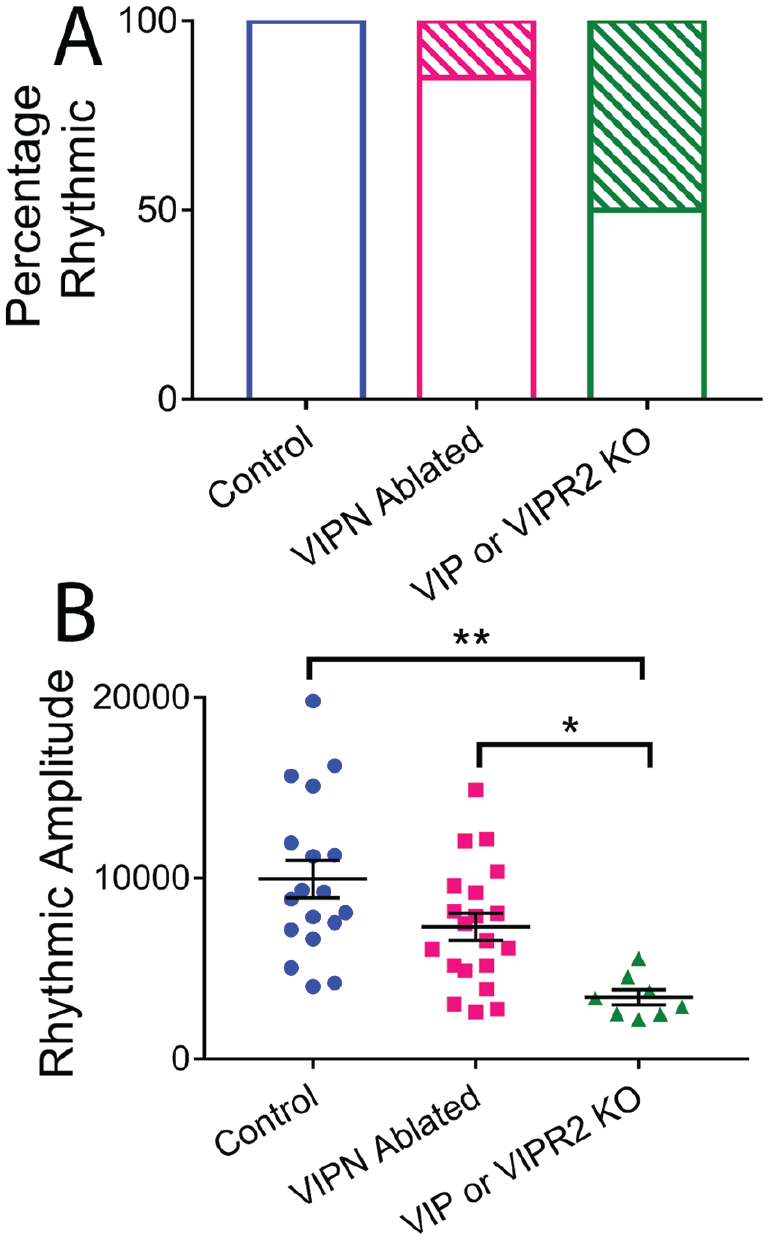
Comparison of vasoactive intestinal polypeptide (VIP) SCN neuronal ablation to genetic knockout of Vip or Vipr2. (A) Under constant darkness, only 15% of VIP neuron (VIPN)–ablated mice were behaviorally arrhythmic (17/20) compared with 50% of Vip or Vipr2 knockout mice (4/8). (B) Rhythmic amplitude was substantially decreased in Vip or Vipr2 knockout mice compared with control mice or mice lacking VIP neurons.
In Vitro Deletion of VIP Neurons
The SCN from triple transgenic mice heterozygous for VIP-IRES-Cre, Rosa-tdtomato, and PER2::Luc were used for these experiments. Briefly, between postnatal day 6 and 7 (P6-P7), these mice were decapitated, and brains were rapidly removed. The bilateral SCN was dissected from 300-µm-thick coronal brain slices, imaged to verify tdtomato fluorescence, and cultured on Millicell-CM inserts (Millipore, Billerica, MA) in prewarmed culture medium (DMEM supplemented with 10 mM HEPES and 10 µM beetle luciferin; Promega, Madison, WI). The following day, SCN were infected with 1 to 2 µL of either AAV8-Caspase-TEVP or AAV9-DIO-eYFP viruses. The virus was applied directly on top of the slice. Each SCN explant in its sealed 35-mm Petri dish (BD Biosciences, San Jose, CA) was placed under a photomultiplier tube (HC135-11, Hamamatsu Corp., Hamamatsu City, Japan) in a light-tight incubator kept at 36 °C. Bioluminescence counts were integrated every 10 min for up to a month using custom software. Each slice was imaged on an epifluorescent microscope (TE2000 inverted, Nikon) once per week to visualize the changes in the number of VIP neurons, and media were replaced every 2 weeks. All data analyzed were blinded to genotype and treatment. Circadian period was characterized using a cosine fit on detrended data (Chronostar software, generous gift of Dr. Achim Kramer, Charité University). Amplitude was calculated using a custom-written Python script that measured the daily peak-to-trough difference from the raw data. Epifluorescent images for tdtomato-positive neurons were quantified using ImageJ’s cell counter plug in.
Experimental Design and Statistical Analyses
All statistical tests were performed using Prism version 7.00 for Windows 7 (GraphPad Software, San Diego, CA). For circadian locomotor experiments, 1-way analyses of variance (ANOVAs) were calculated to compare 3 groups (control, VIPN present, and VIPN ablated). When comparing locomotor behavior between 4 groups (control, VIPN present, VIPN ablated, and Vip or Vipr2 knockout), we used either a 1-way ANOVA or Kruskal-Wallis test (e.g., when standard deviations were significantly different between the groups in comparing rhythmic amplitudes in Figure 2 and as noted in Table 1). When comparing corticosterone measurements, VIP/AVP staining intensity, and bioluminescence, we used an unpaired Student t test. Welch’s correction was applied when standard deviations were significantly different (corticosterone maximum and amplitude, AVP staining intensity, in vitro VIP neuron count, mean bioluminescence, correlation coefficient). We calculated the Pearson correlation to test the relationship between locomotor phenotype and VIP staining intensity.
Light-dark and constant-darkness analyses.
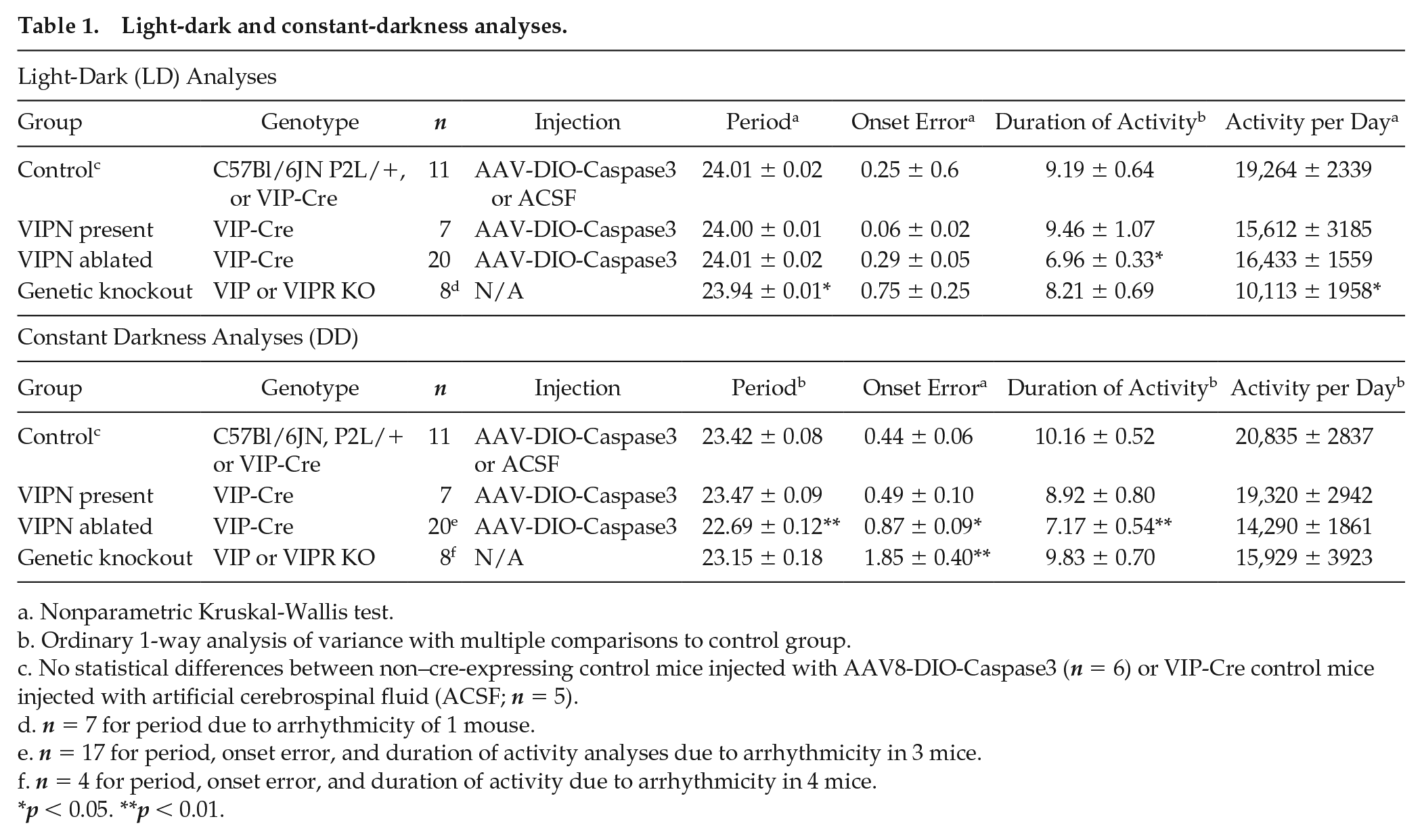
Nonparametric Kruskal-Wallis test.
Ordinary 1-way analysis of variance with multiple comparisons to control group.
No statistical differences between non–cre-expressing control mice injected with AAV8-DIO-Caspase3 (n = 6) or VIP-Cre control mice injected with artificial cerebrospinal fluid (ACSF; n = 5).
n = 7 for period due to arrhythmicity of 1 mouse.
n = 17 for period, onset error, and duration of activity analyses due to arrhythmicity in 3 mice.
n = 4 for period, onset error, and duration of activity due to arrhythmicity in 4 mice.
p < 0.05. **p < 0.01.
Results
Ablation of VIP Neurons in Adult Mice Alters Circadian Locomotor Activity
To test whether VIP SCN neurons are necessary for rhythmic locomotor behavior, we targeted them for ablation in adult mice and monitored running-wheel activity. To selectively ablate VIP SCN neurons, we bilaterally injected an adeno-associated virus expressing Cre-dependent Caspase3, AAV8-Casp3-TEVP (Yang et al., 2013) into the SCN of VIP-IRES-Cre heterozygous mice (Fig. 1A). Expression of the Caspase3 construct cell-autonomously triggered the apoptotic pathway, resulting in selective ablation of VIP SCN neurons. At the conclusion of the behavioral experiments, mice were sacrificed to confirm the ablation of VIP SCN neurons. Hereafter, experimental mice with confirmed ablation of VIP neurons are referred to as “VIPN ablated” (n = 20). Virus-treated mice that retained wild-type levels of VIP expression are referred to as “VIPN present” (n = 7). Control mice were either VIP-IRES-Cre mice injected with ACSF (n = 5) or non–Cre-expressing mice injected with AAV8-Casp3-TEVP (n = 6).
Starting at least 1 week after surgery, we recorded wheel-running behavior in entrained conditions (12 h:12 h light:dark [LD] cycle) for 1 to 2 weeks and then in constant darkness (DD) for 2 to 3 weeks (Fig. 1B and Fig. 3). Mice lacking SCN VIP neurons showed normal entrainment in a light cycle (Figs. 1C, 2, and 3) but differed from controls in constant conditions, running for approximately 3 h less per day (7.17 ± 0.54 h compared with 10.16 ± 0.52 h, F2,32 = 7.24, p = 0.0025), with a 0.7-h shorter circadian period (22.69 ± 0.12 h compared with 23.42 ± 0.08 h, F2,32 = 16.59, p < 0.0001) and doubled variability in day-to-day activity onset (0.87 ± 0.09 h compared with 0.44 ± 0.06 h, F2,32 = 7.374, p = 0.0023; Table 1; Fig. 1D-F). Compared with Vip- or Vipr2-null mice, VIPN-ablated mice had higher circadian amplitudes (Vip-/- or Vipr2-/-: 3409 ± 1166; VIPN-ablated mice: 7304 ± 3363, H[2] = 16.58, p = 0.0003) and were less likely to become arrhythmic in constant darkness (n = 4 arrhythmic of 8 Vip-/- or Vipr2-/-; n = 3 of 20 VIPN-ablated mice; Fig. 2A, B). Loss of VIP SCN VIP neurons in adulthood or VIP signaling from birth resulted in a similar shortened circadian period in those mice that retained rhythmicity in constant darkness. In contrast, global loss of VIP signaling (Colwell et al., 2003; Harmer et al., 2002) from birth results in phenotypes not seen with SCN-specific deletion of VIP neurons in adults: advanced phase angle of entrainment in a light cycle and a reduced circadian amplitude and fraction of rhythmic mice in constant darkness.

Locomotor phenotype in constant darkness. Locomotor actograms during constant darkness ordered by level of vasoactive intestinal polypeptide (VIP) staining (top-left to bottom-right; outline color indicates control, VIP neurons present). Note that the predominant phenotype among mice with the lowest level of VIP staining was a shortened period and a decreased duration of daily activity.
Ablation of VIP Neurons Dampens the Daily Surge in Corticosterone
Following locomotor activity recordings, a subset of mice was placed in custom-built cages that allowed frequent, noninvasive measurement of fecal corticosterone levels. Mice habituated to the cages for 2 days in total darkness before fecal collection every 4 h for 2 more days (n = 13 VIPN ablated, n = 10 control). When we aligned individual corticosterone rhythms to the time of their first daily peak, we found that mice lacking VIP neurons had dampened circadian corticosterone levels compared with controls, with roughly half the average (29.77 ± 2.43 vs. 41.86 ± 4.20 ng corticosterone per mg feces, t[21] = 2.63, p = 0.0157), maximum (48.32 ± 3.84 vs. 94.36 ± 9.93 ng corticosterone per mg feces, t[11.69] = 4.32, p = 0.001), and peak-to-trough daily amplitude of corticosterone levels (30.37 ± 3.25 vs. 74.06 ± 9.50 ng/mg, t[11.12] = 4.35, p = 0.0011; Fig. 4). We found no difference in the daily minimum of corticosterone (17.04 ± 2.19 vs. 20.18 ± 3.11 ng/mg, t[21] = 0.85, p = 0.4042) and conclude that SCN VIP neurons are necessary for the daily surge in corticosterone.
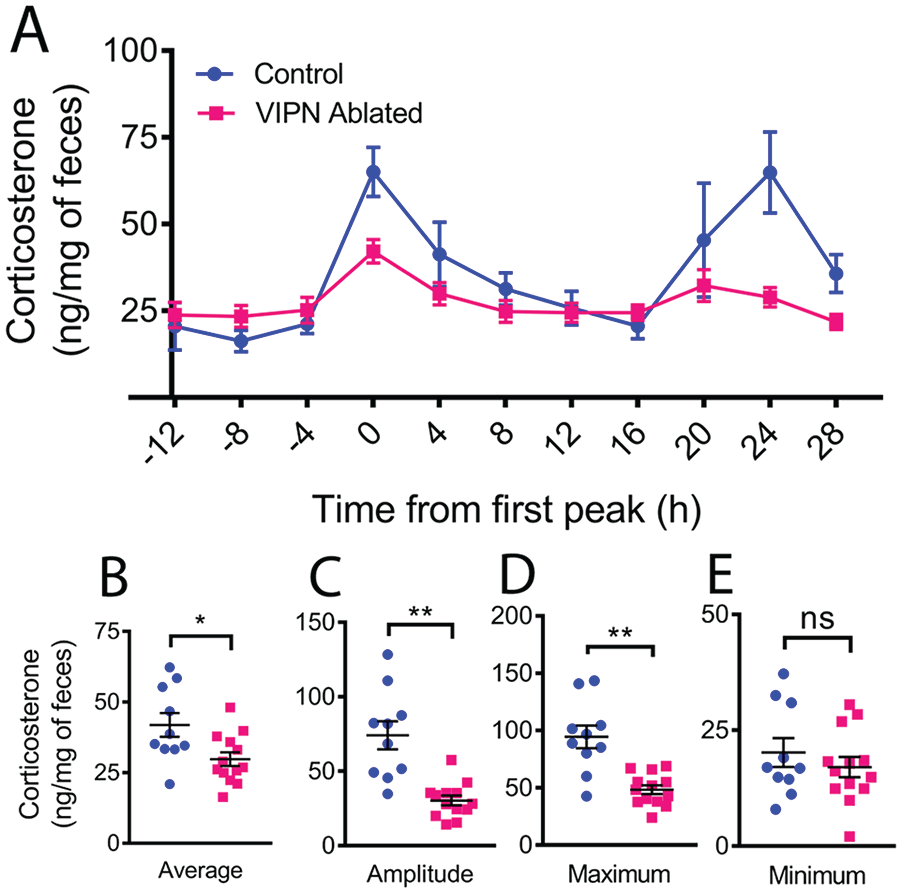
Ablation of vasoactive intestinal polypeptide (VIP) neurons dampens the circadian rhythm in corticosterone. (A) Average profile of fecal corticosterone levels over 2 days in constant darkness from mice with and without SCN VIP neurons. Individual traces were phase aligned to their first daily peak. (B) Loss of VIP neurons the reduced daily average, (C) peak-to-trough amplitude of corticosterone, and (D) maximum but did not change the daily minimum (E). Color version of the figure is available online.
Circadian Phenotype Correlates with VIP Staining Intensity
To measure the extent of VIP neuron ablation, we quantified VIP immunofluorescence staining. All mice were sacrificed during entrained conditions (12 h:12 h light-dark cycle) during late subjective day (CT 7-10) when AVP staining is high (Jin et al., 1999; Silver et al., 1999; Tominaga et al., 1992). On average, VIP-IRES-Cre mice injected with Cre-dependent Caspase3 had significant reductions in the intensity of VIP staining compared with control mice (110.9 ± 43.6 compared with 483.4 ± 98.9 relative VIP staining intensity, t[27] = 3.768, p = 0.0008) but no changes in AVP intensity (791.3 ± 8.3 compared with 770.6 ± 57.2 relative AVP staining intensity, t[23.65] = 0.2081, p = 0.8369; Fig. 5A, B). Among the mice that received the virus into the SCN, we found a subset (n = 4) with VIP staining levels comparable with controls (VIPN-present mice). Across all mice, the staining intensity for VIP in the SCN was strongly correlated with increasing period (r[24] = 0.40, p = 0.0005; Fig. 5C) and decreasing onset variability (r[24] = 0.3485, p = 0.0015; Fig. 5D) of daily locomotor rhythms in constant darkness. The 3 VIPN-ablated mice that were behaviorally arrhythmic had comparable levels of VIP staining to other VIPN-ablated mice (see orange asterisks, Fig. 5C, D). AVP staining intensity did not correlate with locomotor period (r[24] = 0.01, p = 0.55) or onset variability (r[24] = 0.09, p = 0.14). We conclude that reducing the number of VIP neurons in the SCN results in reduced circadian period and reduced cycle-to-cycle precision in locomotor rhythms.
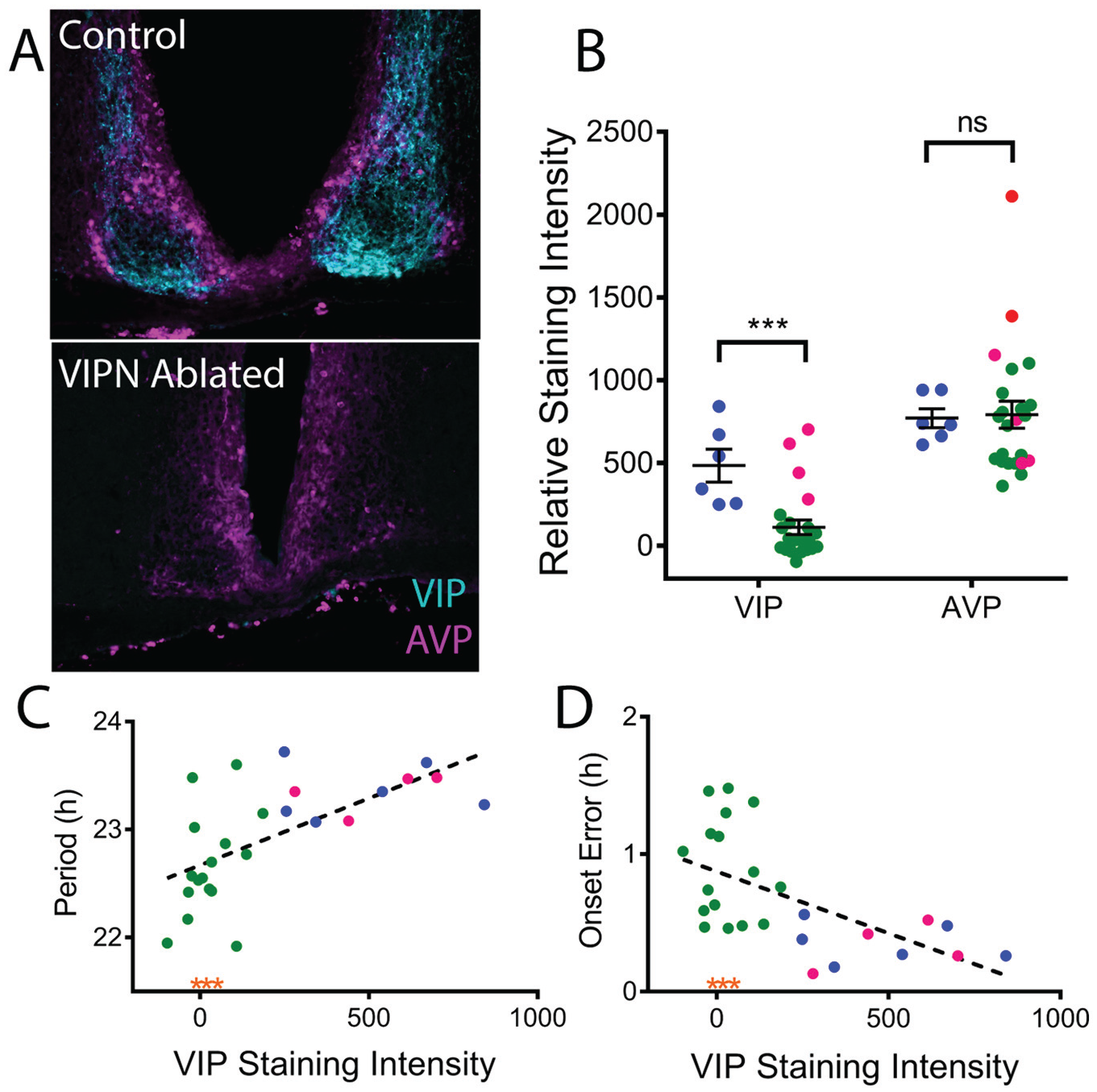
Locomotor phenotype correlates with level of vasoactive intestinal polypeptide (VIP) expression. (A) Representative coronal sections of the SCN from control and VIP neuron (VIPN)–ablated mice double labeled for VIP (cyan) and arginine-vasopressin (AVP; magenta). Note the loss of VIP immunoreactivity in the mouse treated with AAV-Casp3. (B) Quantification of VIP and AVP immunostaining revealed lower average VIP, but not AVP, immunostaining in the SCN of mice treated with AAV8-Casp3 (green: VIPN ablated; magenta: VIPN present) compared with controls (blue dots). (C) Circadian period length correlated with VIP staining intensity, while (D) day-to-day onset variability inversely correlated with VIP staining intensity. Note that VIPN-ablated mice exhibiting arrhythmic circadian behavior have VIP staining intensities in line with other VIPN-ablated mice (orange asterisks).
In Vitro Deletion of VIP Neurons Impairs Circadian Rhythmicity in the Neonatal SCN
To further compare the effects of loss of VIP to loss of VIP neurons, we generated triple transgenic mice that expressed a reporter of the clock protein, PERIOD2 (PER2), in all cells and red fluorescence only in VIP neurons (VIP-IRES-Cre/+, floxed-ROSA-tdTOMATO/+, PERIOD2::Luciferase/+). At postnatal day 6, we dissected the SCN and infected SCN slices with either AAV-DIO-Caspase3 to kill VIP neurons or a control virus (AAV-DIO-eYFP). We then recorded PER2-driven bioluminescence every 10 min for up to 1 month while imaging fluorescence once a week. By 3 weeks after viral expression, caspase-treated SCN slices showed significantly fewer VIP neurons than control slices (18 ± 5 compared with 268 ± 28, t[5.327] = 8.71, p = 0.0002; Figure 6). In VIPN-ablated SCN slices, both the average (127,813 ± 7770 compared with 229,559 ± 20,699 counts/10 min, t[13] = 5.8, p < 0.0001) and daily peak-to-trough amplitude of PER2 expression (36,641 ± 8897 compared with 132,085 ± 14,425 counts/10 min, t[13] = 5.794, p < 0.0001) were dramatically reduced. Notably, the quality of the rhythm was also reduced (0.91 ± 0.03 compared with 0.99 ± 0.00, t[7.026] = 2.46, p = 0.0434, as assessed by the correlation coefficient of a cosine fit to each bioluminescence trace), although the circadian period was not (24.04 ± 0.21 h compared with 23.64 ± 0.17 h, t[13] = 1.42, p = 0.1776). We conclude that loss of VIP neurons shortly after birth reduces the amplitude and precision of circadian rhythms in the SCN. These results are consistent with the reduced synchrony of circadian cells in the SCN of Vip- and Vipr2-null mice.
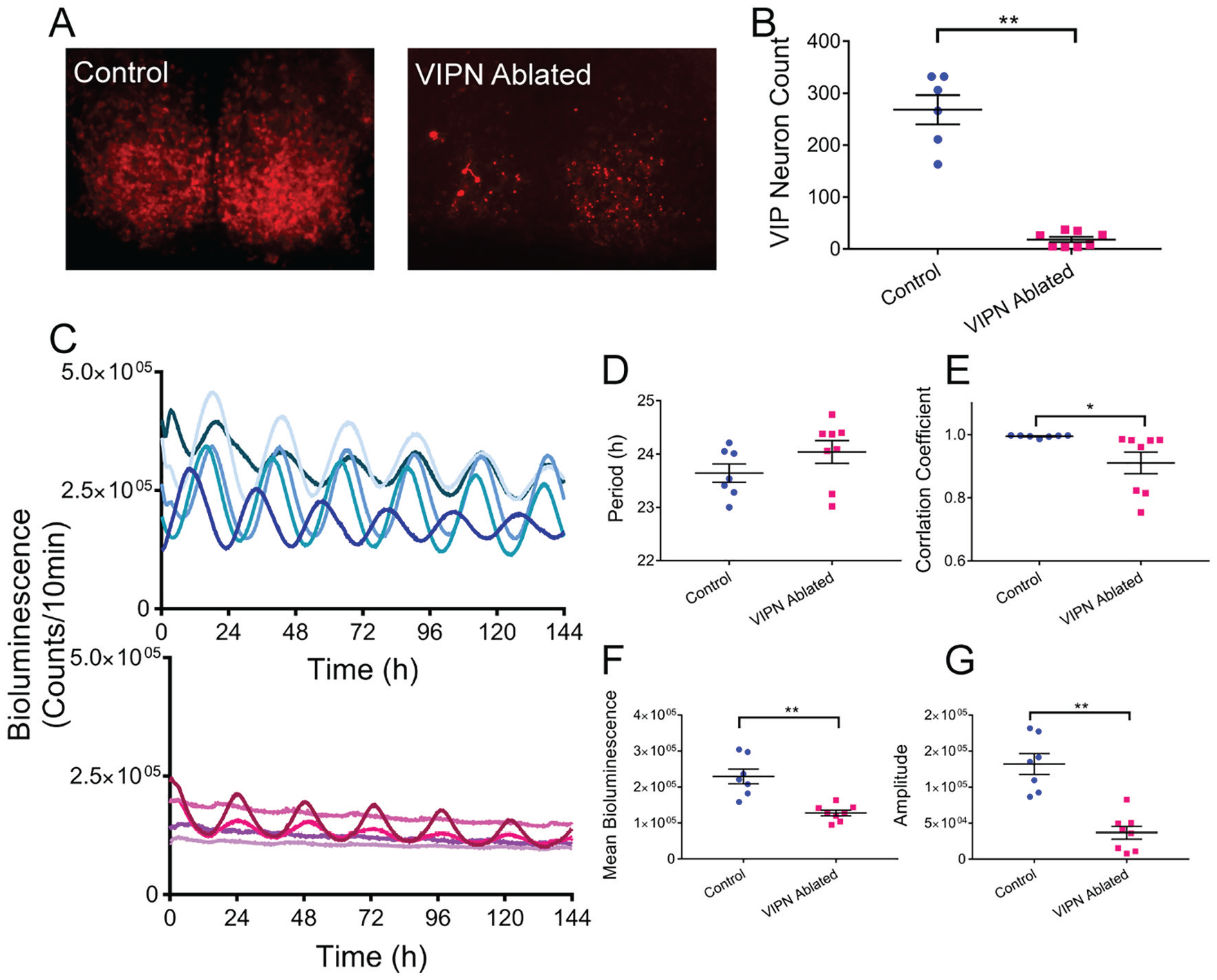
Ablation of vasoactive intestinal polypeptide (VIP) neurons in neonatal SCN slices in vitro decreases circadian rhythmicity. (A) VIP-specific tdTOMATO fluorescence was substantially decreased in SCN slices treated with AAV-DIO-Caspase3 (VIPN ablated) compared with those treated with an eYFP-expressing virus (control). (B) Quantification of the number of VIP neurons remaining in the SCN slices 3 weeks after treatment. (C) PER2 expression over days from individual SCN slices 3 weeks after treatment with control (top blue traces) or caspase3 (bottom magenta traces) viruses. (D) SCN slices lacking VIP neurons showed no change in circadian period (E) but did show reduction in rhythm quality as measured by the correlation coefficient of a cosine fit, (F) mean bioluminescence, and (G) peak-to-tough circadian amplitude. Color version of the figure is available online.
Discussion
We found that ablation of VIP SCN neurons in adult mice caused a circadian phenotype distinct from that seen in Vip- or Vipr2-null mice. Whereas adult loss of VIP neurons has relatively little effect on circadian rhythms in locomotion compared with the arrhythmicity seen in mice deficient for VIP signaling from birth, both manipulations result in a blunting of glucocorticoid rhythms. Combined with the striking decreases in circadian gene expression and rhythmicity seen when VIP neurons are ablated in the neonatal SCN, we conclude that VIP SCN neurons may have differential roles in development and adulthood.
Viral targeting of VIP neurons allowed the comparison between mice with variable numbers of SCN VIP neurons. The remaining numbers of VIP neurons correlated highly with the locomotor phenotype: fewer cells, shorter circadian period, and greater cycle-to-cycle variability. The procedure of injecting the virus bilaterally into the SCN produced minimal damage to the SCN and surrounding brain, so the variability in VIP neuron ablation likely resulted from the precision of the SCN targeting during the stereotaxic surgery and the limited diffusion of the AAV8 serotype virus. We quantified the relative loss of VIP neurons based on maximal immunofluorescence. Although this does not resolve whether reduced fluorescence resulted from loss of processes or cell bodies, it provides a conservative indicator of the presence of any VIP neurons. The strong correlation between reduced VIP immunofluorescence and shortened circadian period and increased cycle-to-cycle variability suggests that these phenotypes depend on VIP signaling. It remains to be seen whether the period-lengthening effect of VIP neurons seen in this study and others is similar or inverted in nocturnal versus diurnal animals.
The absence of a severe circadian locomotor phenotype in VIPN-ablated, adult mice suggests that VIP signaling plays a distinct role in the development of the circadian system. VIP neuropeptide expression within the SCN is first detected in the early postnatal days followed by retinal innervation of the SCN (Carmona-Alcocer et al., 2018; Carmona-Alcocer et al., 2020; Sernagor, 2005). VIP may therefore be necessary for the correct formation of retinal input to the SCN, intra-SCN connectivity, and SCN output to hypothalamic areas. It is possible that disruption of VIP signaling during this critical postnatal period contributes to the severe circadian disruption seen in Vip- or Vipr2-deficient mice. This appears to be different from findings in Drosophila, in which deletion of the neuropeptide PDF gene or cells produces an identical circadian behavioral phenotype, although deletion of these neurons in the adult has not yet been performed (Renn et al., 1999). In mammals, VIP neuropeptide signaling may play a role in establishing circadian circuit features during development, as it does in modulating the excitatory/inhibitory balance in visual cortex (Batista-Brito et al., 2017).
Because we found that deletion of VIP neurons in the neonatal SCN results in reductions in circadian rhythms in the SCN that are similar to previously published genetic or pharmacological abrogation of VIP signaling (Aton et al., 2005; Brown et al., 2007; Colwell et al., 2003; Harmar et al., 2002; Maywood et al., 2006), we conclude that these neurons are important for synchrony among circadian cells early in development but not in the adult. This is consistent with evidence that other signals, including the neuropeptides AVP or gastrin-releasing peptide, can act to synchronize circadian cells in the SCN (Brown, 2005; Herzog et al., 2017; Maywood et al., 2011). Furthermore, mice that lose VIP neurons in adulthood share features with the subset of mice born deficient for VIP signaling that retain circadian rhythms such as significantly shortened periods. This indicates that SCN VIP neurons normally serve to lengthen the circadian period, consistent with results showing that chronic stimulation of VIP receptors lengthens the circadian period in vivo (Pantazopoulos et al., 2010), VIP largely phase delays the in vitro SCN (An et al., 2011; Reed et al., 2001), activation of SCN VIP neurons can phase delay behavior in vivo (Mazuski et al., 2018), and the phase-delaying effects of light require activation of SCN VIP neurons (Jones et al., 2018). Our results also suggest that VIP neurons are necessary for circadian locomotor activity in the second half of the circadian night. Ablation of VIP neurons compresses circadian locomotor activity to the first part of the subjective night.
We found reduced corticosterone levels in mice lacking SCN VIP neurons, suggesting that they underlie stimulation of the daily surge in glucocorticoids. Prior results showing loss of glucocorticoid rhythms in VIP-null mice (Loh et al., 2008) can now be interpreted as likely caused by the disruption of VIP SCN neurons and not VIP in other areas (such as the adrenal glands) or arrhythmic circadian behavior. Neuronal projections from the SCN to the PVN of the hypothalamus are necessary for the daily rhythm in glucocorticoids (Buijs et al., 1993), and SCN VIP neurons project to the PVN (Abrahamson and Moore, 2001). It remains to be tested whether activity in these VIP projections is sufficient and necessary for daily regulation of glucocorticoid release. Future work should further elucidate the relative roles of VIP in establishing and maintaining the many daily rhythms in physiology and behavior.
Footnotes
Acknowledgements
This work was supported by National Institutes of Health grants UO1 EB02195601 (E.D.H.) and F31-GM11517 (C.M.). The authors thank members of the Herzog lab, especially Drs. Matt Tso and Daniel Granados-Fuentes, for technical contributions.
Conflict of Interest Statement
The authors have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.