
Abstract
The suprachiasmatic nucleus (SCN) of the hypothalamus consists of a highly heterogeneous neuronal population networked together to allow precise and robust circadian timekeeping in mammals. While the critical importance of SCN neurons in regulating circadian rhythms has been extensively studied, the roles of SCN astrocytes in circadian system function are not well understood. Recent experiments have demonstrated that SCN astrocytes are circadian oscillators with the same functional clock genes as SCN neurons. Astrocytes generate rhythmic outputs that are thought to modulate neuronal activity through pre- and postsynaptic interactions. In this study, we developed an in silico multicellular model of the SCN clock to investigate the impact of astrocytes in modulating neuronal activity and affecting key clock properties such as circadian rhythmicity, period, and synchronization. The model predicted that astrocytes could alter the rhythmic activity of neurons via bidirectional interactions at tripartite synapses. Specifically, astrocyte-regulated extracellular glutamate was predicted to increase neuropeptide signaling from neurons. Consistent with experimental results, we found that astrocytes could increase the circadian period and enhance neural synchronization according to their endogenous circadian period. The impact of astrocytic modulation of circadian rhythm amplitude, period, and synchronization was predicted to be strongest when astrocytes had periods between 0 and 2 h longer than neurons. Increasing the number of neurons coupled to the astrocyte also increased its impact on period modulation and synchrony. These computational results suggest that signals that modulate astrocytic rhythms or signaling (e.g., as a function of season, age, or treatment) could cause disruptions in circadian rhythm or serve as putative therapeutic targets.
The suprachiasmatic nucleus (SCN) in mammals consists of approximately 20,000 neurons that act as circadian oscillators and about one-third as many astrocytes that have a poorly understood role in circadian system function (Silver, 2018). Neurons generating rhythms in gene expression and firing frequency are coupled together for robustness and synchronized to establish 24-h circadian rhythms for regulating various physiological and behavioral processes (Belle and Diekman, 2018; Hastings et al., 2014; Welsh et al., 2010). Impairment of the circadian clock has been implicated in numerous disorders, including sleep problems, mental illness, and metabolic diseases (Foster and Kreitzman, 2014; Liu et al., 2007). Neuronal coupling is mediated by neurotransmitters triggered by action potentials and released from presynaptic neurons (Allen et al., 2017; Colwell, 2011; Herzog et al., 2017). The primary coupling mechanism involves the neurotransmitter vasoactive intestinal polypeptide (VIP), which is necessary and sufficient for synchronization of the heterogeneous neural population (Aton et al., 2005; Herzog et al., 2017; Vosko et al., 2007), despite its wide range of endogenous periods (Aton et al., 2005; Honma, 2018). The role of other SCN neurotransmitters including γ-aminobutyric acid (GABA) and glutamate in circadian system function is less well understood (Albers et al., 2017; Chi-Castañeda and Ortega, 2018; Evans, 2016; Herzog et al., 2017; Ono et al., 2018).
Astrocytes are ubiquitous throughout the mammalian brain and have been shown to interact with neurons via spatial proximity and gliotransmission at synapses to regulate brain network function, behavior, and plasticity (Araque et al., 1999; Halassa et al., 2007; Volterra and Meldolesi, 2005). SCN astrocytes are rhythmically associated with neurons through intercellular coupling agents including VIP and arginine vasopressin (Becquet et al., 2008; Marpegan et al., 2009). Several recent studies have shown the crucial role of SCN astrocytes in tuning the SCN neuronal clock, contributing to circadian rhythms, and influencing neuronal physiology (Belle and Allen, 2018; Ruben and Hogenesch, 2017). The loss of astrocytic Bmal1 in the SCN lengthened circadian period and locomotor behavior (Tso et al., 2017), while deletion of astrocytic Bmal1 throughout the brain altered daily behavioral rhythms and GABA levels in the brain (Barca-Mayo et al., 2017). When SCN neurons expressed 1 copy of the tau mutation of CK1ε(which shortens their circadian period) and astrocytes did not express CK1ε(which lengthens their circadian period; Tso et al., 2017), the circadian period of the SCN and locomotor activity were lengthened.
Inhibition of GABA transporters expressed in SCN astrocytes resulted in an elevated tonic GABAA receptor–mediated current and affected the periodicity of Per1 expression in SCN neurons (Moldavan et al., 2017). Astrocytes also have been shown to synaptically communicate with and tune SCN neurons through glutamatergic transmission (Brancaccio et al., 2017; Scofield, 2018). Under constant darkness, the rhythmic release of glutamate via excitatory amino acid transporters (EAATs) enabled astrocytes to maintain higher intracellular calcium levels in presynaptic neurons through glutamate receptor activation, which in turn produced elevated inhibitory GABAergic tone in the dorsal SCN (Brancaccio et al., 2017, 2019). Although these experimental studies demonstrated that astrocytes affect circadian behavior, the individual contributions associated with GABA (Barca-Mayo et al., 2017), glutamate (Brancaccio et al., 2017, 2019), and VIP (Marpegan et al., 2009) are poorly understood.
Mathematical modeling has proven to be a powerful complementary tool in experimentation for understanding the roles of the molecular clock, electrophysiology, cell-to-cell coupling, and neural network topology on circadian behavior (Bernard et al., 2007; DeWoskin et al., 2015; Hafner et al., 2012; Kingsbury et al., 2016; Vasalou et al., 2009, 2011; Vasalou and Henson, 2011). To our knowledge, existing models are restricted to neuronal behavior and do not directly include the possible modulatory effects of SCN astrocytes, which are also self-sustained oscillators. Although there was a proposed model of distinct multioscillatory units in the circadian system, the model indirectly introduced the concept of neutral elements that did not exhibit self-sustained oscillation and were presumably possible glial cells in such a system (Diez-Noguera, 1994). Also, some astrocyte models have been developed for other brain regions such as the cortex and hippocampus (De Pittà et al., 2012; Manninen et al., 2018; Oschmann et al., 2018). These models span single astrocytes, the neuron-astrocyte tripartite synapse, and larger neuron-astrocyte networks but are not directly applicable to the SCN. The development of SCN-specific neuron-astrocyte network models has the potential to provide mechanistic underpinnings for observed experimental behavior, expand our understanding of circadian timekeeping, and provide experimentally testable hypotheses about astrocytic modulation of neuronal networks.
The goal of this in silico study was to develop a simplified but biophysically based multicellular model of neuronal-astrocytic SCN networks and to use the model to gain better understanding of the synaptic communication mechanisms that may have driven circadian behavior observed in several recent experimental studies (Barca-Mayo et al., 2017; Brancaccio et al., 2017, 2019; Ruben and Hogenesch, 2017; Tso et al., 2017). Our computational model was based on complex bidirectional communication between glial and neuronal cells reported in other brain regions (De Pittà et al., 2012; Manninen et al., 2018; Oschmann et al., 2018) specialized to the SCN to probe potential influences of astrocytes on the neuronal clock (Barca-Mayo et al., 2017; Brancaccio et al., 2017, 2019; Moldavan et al., 2015; Tso et al., 2017). By developing a novel model of an SCN astrocyte, we applied the concept of a tripartite synapse consisting of 1 astrocyte and 2 (pre- and postsynaptic) neurons (Vasalou et al., 2011; Vasalou and Henson, 2010) to construct an integrated SCN network model. The model accounted for VIP, GABAergic, and glutamatergic signaling between a single astrocyte and a heterogeneous population of 100 neurons. We focused our computational studies on unraveling the effects of neuronal and astrocytic endogenous periods, bidirectional neuronal-astrocytic signaling, intercellular coupling strength, and neural network topology on circadian rhythmicity, synchronization, and period of the neural population.
Materials And Methods
SCN Neuronal-astrocytic Network Model
The SCN network model was based on the hypothesis that astrocytes alter neuronal dynamics via bidirectional interactions mediated through the actions of VIP, GABA, and glutamate at tripartite synapses. The integrated model consisted of a neuronal model based on our previous studies (Vasalou et al., 2011; Vasalou and Henson, 2010) and an astrocytic model specifically developed for this study (Fig. 1A). The single-neuron model consisted of 3 interconnected modules (gene regulation, electrophysiology, and neurotransmitter signaling) following our previous modeling studies.
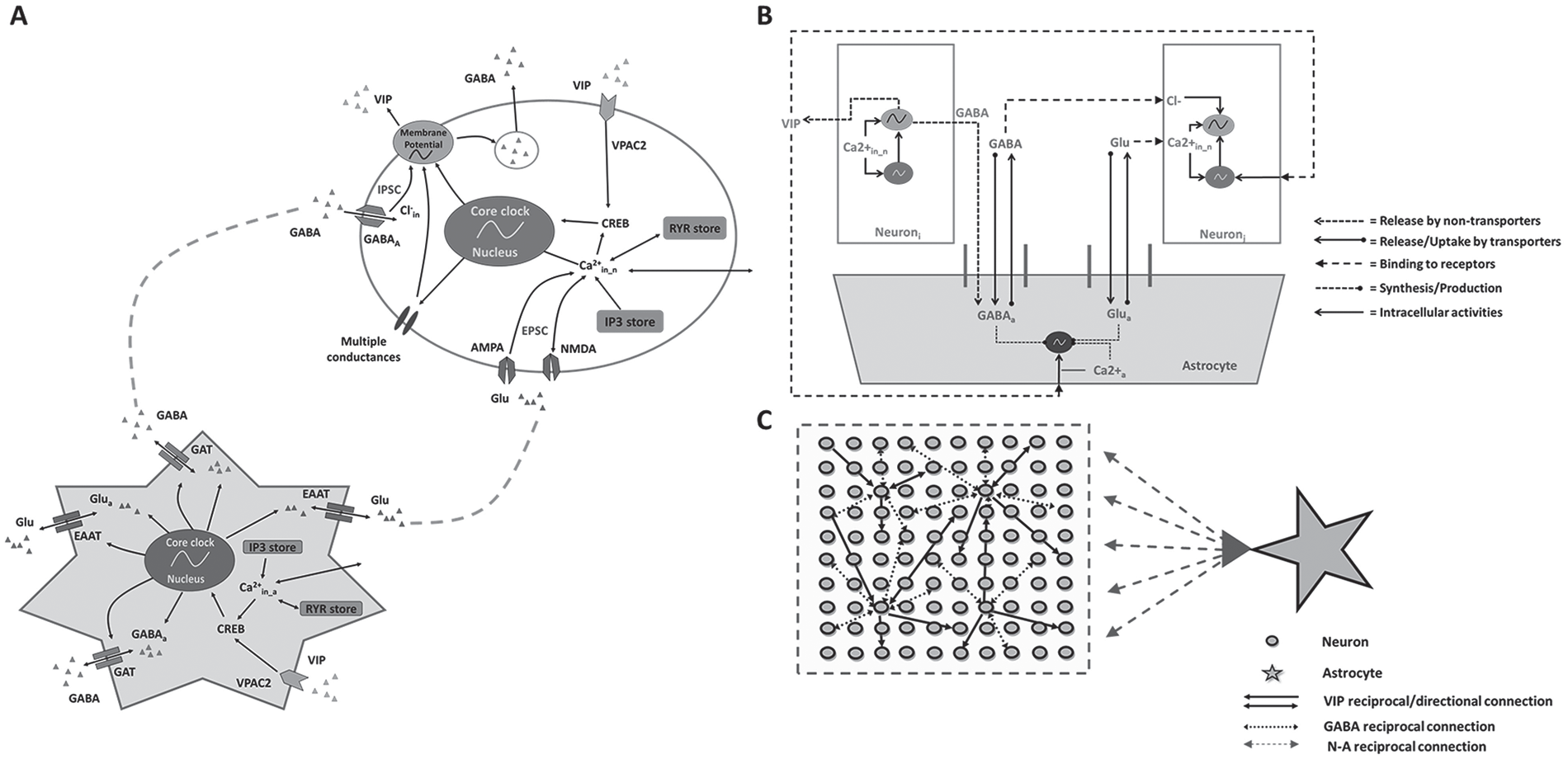
In silico SCN model architecture and neuron-astrocyte interactions. (A) Schematic representation of the coupled neural and astrocytic oscillators. The neuron model (oval) was modified from Vasalou and Henson (2010), whereas we developed the astrocyte model (star) in this study. The astrocyte model included core clock gene regulation, intracellular pathways (e.g., calcium and CREB), γ-aminobutyric acid transporters (GABA; GATs), glutamate transporters (excitatory amino acid transporters), and vasoactive intestinal polypeptide (VIP) receptors. (B) Schematic representation of the tripartite synapse model with cell-to-cell communication of both neurons and the astrocyte at synapses mediated through multiple intercellular signaling pathways (VIP, GABA, and glutamate). (C) Schematic representation of the SCN network model with 100 heterogeneous neurons synaptically interacting with a single astrocyte. Neuron-to-neuron connectivity followed a small-world network topology (Vasalou et al., 2009), while astrocyte-to-neuron connectivity followed a mean-field network topology. Inositol 1,4,5-trisphosphate (IP3) and ryanodine (RYR) are different calcium stores; Glu is glutamate; CREB is a cellular transcription factor; AMPA and NMDA are glutamate receptors; IPSC and EPSC are inhibitory and excitatory postsynaptic currents, respectively.
The astrocyte model retained the same gene regulation module since core clock genes identified in SCN neurons are preserved in SCN astrocytes (Brancaccio et al., 2017; Tso et al., 2017), while the neurotransmitter signaling module was modified as detailed below, and the electrophysiology module was eliminated since astrocytes do not exhibit firing behavior. Instead, intracellular calcium represented a proxy for astrocytic excitability (Dallérac et al., 2013). We modeled calcium dynamics and internal calcium pools (i.e., IP3 and ryanodine stores; Mesiti et al., 2015; Vasalou and Henson, 2010) and included VPAC2 receptors (Masmoudi-Kouki et al., 2007) for binding VIP released by SCN neurons (Marpegan et al., 2009). The astrocyte model also included GABA transporters, which potentially regulate GABA concentrations in synaptic and extrasynaptic regions (Moldavan et al., 2015, 2017) as well as glutamate transporters for glutamate uptake and release (Brancaccio et al., 2017, 2019). The astrocyte was treated as a GABAergic and glutamatergic cell (Brancaccio et al., 2017; Schousboe et al., 2013; Yoon et al., 2012) that produced glutamate and stored GABA with circadian variations. Hence, our single astrocyte model included core clock gene regulation, intracellular calcium signaling dynamics, glutamate/GABA synthesis, release and uptake, and VIP binding.
To construct the SCN network model, we applied the concept of the tripartite synapse (Perea et al., 2009; Pérez-Alvarez and Araque, 2013), in which an astrocyte can modulate communication between pre- and postsynaptic neurons (Fig. 1B). The tripartite synapse was scaled up to the network level by allowing a heterogeneous population of 100 neurons to be modulated by a single astrocyte (Fig. 1C). The assumption of a single astrocyte in the network was based on knowledge that astrocytes are highly interconnected through gap junctions, essentially sharing their intracellular content (Shinohara et al., 2000); that a single astrocyte can modulate as many as 100,000 synapses (Halassa et al., 2007); and that a neuron is unlikely to be contacted by more than 1 astrocyte as astrocytes typically form nonoverlapping domains (Sofroniew and Vinters, 2010). The astrocyte was assumed to modulate all neuron-to-neuron connections in the simulated network. Following our previous work (Vasalou et al., 2009, 2011), the neuronal network was constructed by placing an ensemble of 100 heterogeneous neurons on an equally spaced, 2-dimensional grid. Approximately 20% of the neurons were randomly selected to synthesize VIP, while all neurons were GABAergic (Vasalou et al., 2009, 2011). Because GABA is commonly recognized as the major inhibitory neurotransmitter in the SCN (Albers et al., 2017), we assumed that GABA signaling was strictly inhibitory following previous modeling studies (e.g., DeWoskin et al., 2015; Vasalou et al., 2011). Hence, our study did not explore the effects of excitatory GABA (Ono et al., 2018) as a function of intracellular chloride levels (Klett and Allen, 2017; Myung et al., 2015), time of day (Alamilla et al., 2014; Albus et al., 2005; Choi et al., 2008; De Jeu and Pennartz, 2002; Wagner et al., 1997), SCN region (Alamilla et al., 2014; Myung et al., 2015), or light stimulation (Farajnia et al., 2014; Myung et al., 2015). Individual neurons were coupled according to small-world (SW) network connectivity (Vasalou et al., 2009, 2011). In this work, 100 heterogeneous neurons were used to construct the SW network, which required a low density of long-range shortcut connections added to the local connections. We did not attempt to differentiate between the dorsal and ventral SCN or account for the effects of light entrainment on circadian behavior. A more complete description of the combined neuronal-astrocytic network including the model equations and parameters is contained in the Supplemental Material.
Computational Studies and Simulation Analysis
All simulations were performed within MATLAB by adapting code developed in our previous modeling studies (Vasalou et al., 2009, 2011). To account for heterogeneity among the SCN neurons, we assigned each neuron model randomly perturbed values of the Bmal1 mRNA transcription and degradation rates and the Per mRNA transcription rate as described elsewhere (Vasalou et al., 2011). These random perturbations yielded an uncoupled network with approximately 25% sustained rhythmic neurons with endogenous oscillation periods ranging between 22 and 26 h as experimentally observed (Aton et al., 2005). The single astrocyte was connected to all 100 neurons, while the neurons were interconnected according to an SW topology following our previous approach (Vasalou et al., 2009, 2011) by randomly assigning 5% of possible long-range connections. Because each simulation contained a different neural population and neural network topology, we performed 5 independent simulations for each scenario (see below) and reported mean values along with the standard error of the mean.
In addition to performing simulations with wild-type (WT) SCN cells, we studied the effects of neuronal and astrocytic mutations on circadian behavior to mimic experimental studies (Brancaccio et al., 2017; Tso et al., 2017; Fig. 2A). These mutations either varied the endogenous cell period or eliminated cell rhythmicity completely through Bmal1 knockout. The endogenous period was changed experimentally by genetic manipulations targeting posttranslational feedback loops. The tau mutation in casein kinase 1 (the kinase that phosphorylates PER proteins) accelerates the dynamics of circadian timekeeping; therefore, the CK1 tau mutation was used experimentally to shorten the endogenous cell period (Maywood et al., 2014; Meng et al., 2008; Takahashi, 2016). By contrast, the loss of F-box/leucine-rich-repeat protein or FBXL3 (the ubiquitin ligase for the CRY proteins; Godinho et al., 2007; Siepka et al., 2007) stabilizes CRY 1 and 2 and increases the endogenous cell period (Maywood et al., 2014). These mutations were implemented in silico by tuning parameters associated with the CKI-PER (k1 and vdPC) and FBXL3-CRY (v3PC and vdCC) pathways (Leloup and Goldbeter, 2003; St John et al., 2014) to achieve shorter or longer endogenous periods (Suppl. Table S3). To implement Bmal1 knockout (Bmal1-/- ) in astrocytes, we set the Bmal1 mRNA synthesis rate (vsB) to zero and removed the Michaelis-Menten terms governing the maximum rates of GABA/glutamate synthesis and transportation driven by the astrocytic clock genes (equations 52-55 in the Supplemental Material) such that those rates were purely constitutive (Leloup and Goldbeter, 2003; Mirsky et al., 2009).
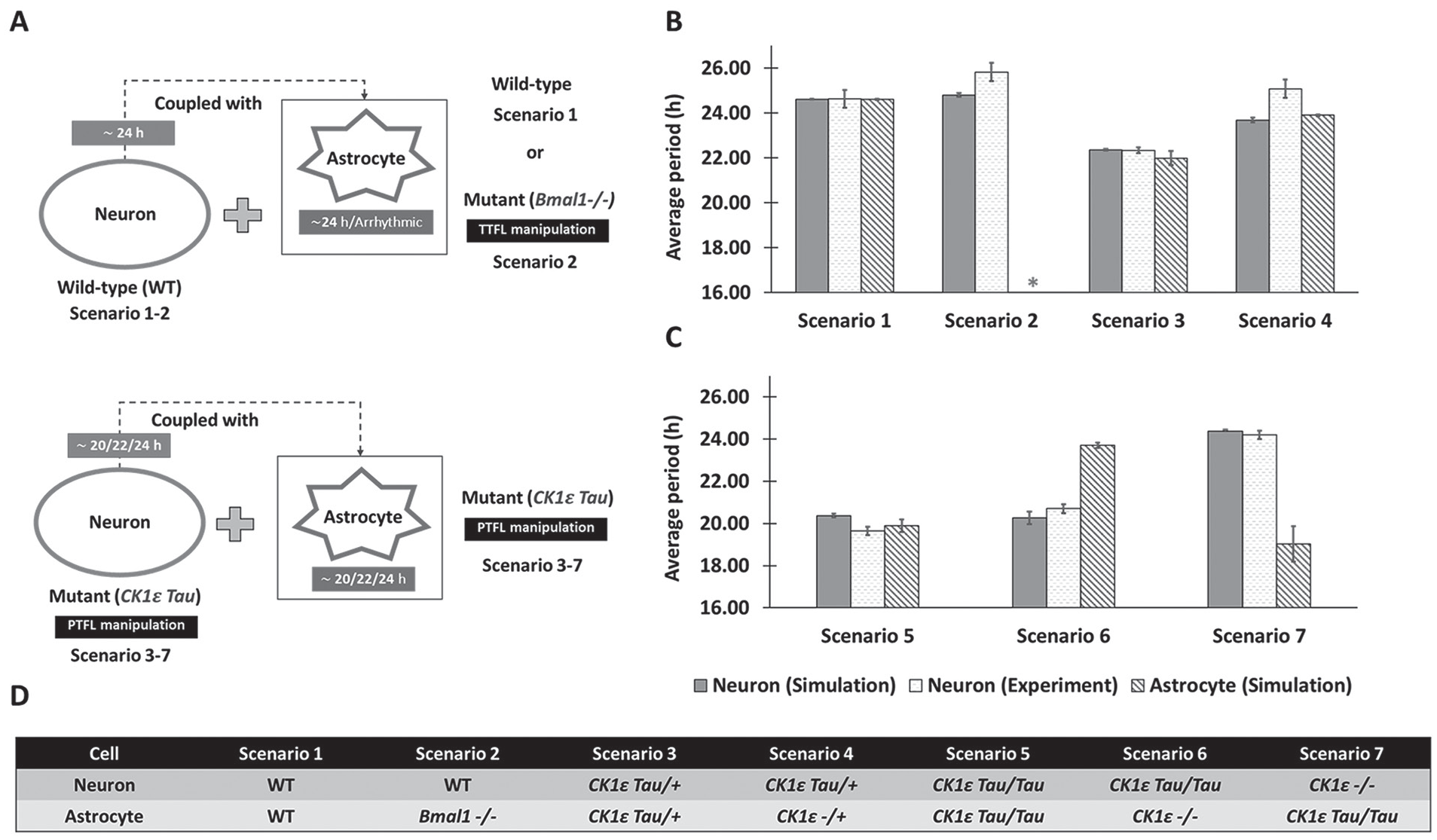
Comparison of neuronal periods from in vitro experiments and in silico simulations for 7 different cell combinations. (A) Schematic diagram showing 7 simulation scenarios that mimic recent experiments on SCN astrocytes We compared the average period (mean ± SEM) of neuronal Per mRNA from the model simulations with the neuronal PER2 from experimental observations reported in Tso et al. (2017) (B) and Brancaccio et al. (2017) (C). We also presented the simulated periods of the astrocyte. The 7 simulation scenarios involve different genetic manipulations that altered the endogenous periods of the astrocyte and the neuronal population as presented in (D). TTFL, transcription-translation feedback loop; PTFL, posttranslational feedback loop; WT, wild type (about 24-h period). *Arrhythmic (no period). The average periods corresponding to Figures 2B and 2C are also provided in Supplemental Tables S4 and S5. CK1εTau/Tau or CK1εTau/+ cells have about 20-h and 22-h endogenous periods, respectively, while CK1ε-/- or CK1ε-/+ cells have an about 24-h endogenous period (Meng et al., 2008).
Simulations were performed by initiating cell-to-cell coupling after about 7 cycles (t ≥ 150 h) and running for 25 total cycles (600 h) to assess circadian behavior. The oscillatory behavior of the Per mRNA rhythms was assessed by period, phase, and amplitude following the methods presented elsewhere (To et al., 2007; Vasalou et al., 2011; Vasalou and Henson, 2010). We also calculated measures of phase synchrony and amplitude coherence of the Per mRNA signal over a specified period using the synchronization index (Strogatz, 2000) and order parameter (R; Gonze et al., 2005), respectively. Both measures yielded values between 0 (no coordination) and 1 (perfect coordination).
Results
Astrocyte-neuron Network Model Predicts Genotype-dependent Circadian Periods
We performed simulations to mimic recent experiments aimed at deciphering the role of SCN astrocytes in circadian timekeeping (Brancaccio et al., 2017; Tso et al., 2017). These experiments have shown, counterintuitively, that SCN astrocytes with mutations in the Bmal1 or CK1ε genes (mutations that are presumed to abolish or lengthen circadian rhythms, respectively) produced a similar increase in period of the SCN and locomotor behavior (Tso et al., 2017). When we simulated circadian networks with different endogenous neuronal and astrocytic periods, we found that the circadian period of the astrocytes affects rhythms in the neurons.
We performed simulations for 7 distinct scenarios that differed according to the endogenous periods of the astrocyte and the neuronal population (Fig. 2A, D). Wild-type neurons were connected to a WT astrocyte (scenario 1) or an astrocyte rendered arrhythmic through Bmal1 knockout (scenario 2). As observed experimentally for scenario 1, the model predicted a small increase in the average period of the coupled neurons compared with their endogenous 24-h period (Fig. 2B). When the astrocyte was arrhythmic, the model correctly predicted a small neuronal period increase compared with scenario 1 (Fig. 2B). Next, we coupled mutant CK1ε Tau/+ neurons with a 22-h period to mutant CK1ε Tau/+ astrocytes with a 22-h period (scenario 3) or CK1ε -/+ astrocytes with a 24-h period (scenario 4). The neuronal period was correctly predicted to be 22 h for scenario 3 and was predicted to be close to the 24-h period of the astrocyte for scenario 4, while the experimentally determined period for this scenario was ~25 h (Fig. 2B).
To test the effects of even shorter circadian periods in astrocytes and neurons (Fig. 2C), we combined CK1ε Tau/Tau neurons with a 20-h period with either a CK1ε Tau/Tau astrocyte with a 20-h period (scenario 5) or a CK1ε -/- astrocyte with a 24-h period (scenario 6). As observed experimentally, both of these scenarios were predicted to generate neuronal periods of ~20 h, although the actual period was slightly higher for scenario 6 when the tau mutation was removed from the astrocyte. Finally, CK1ε -/- neurons with a 24-h period were coupled with CK1ε Tau/Tau astrocytes with a 20-h period (scenario 7), which produced a predicted neuronal period of 24 h in agreement with the experiment. Collectively, these model predictions suggested that astrocytic rhythmicity was not necessary for neuronal rhythmicity but that the coupled neuronal period was dependent on the endogenous periods of both neurons and astrocytes.
Astrocytes Can Entrain Neurons When Their Endogenous Periods Are Sufficiently Close
We sought to determine if differences between the endogenous periods of neurons and astrocytes could be used to predict how astrocytes affect daily rhythms in the SCN. We performed 16 distinct simulations containing cells with endogenous periods ranging from 20 h to 26 h by modifying the parameters associated with PER/CRY stability (Leloup and Goldbeter, 2008; Maywood et al., 2011; St John et al., 2014; Fig. 3A; see the Materials and Method section and Suppl. Table S3).
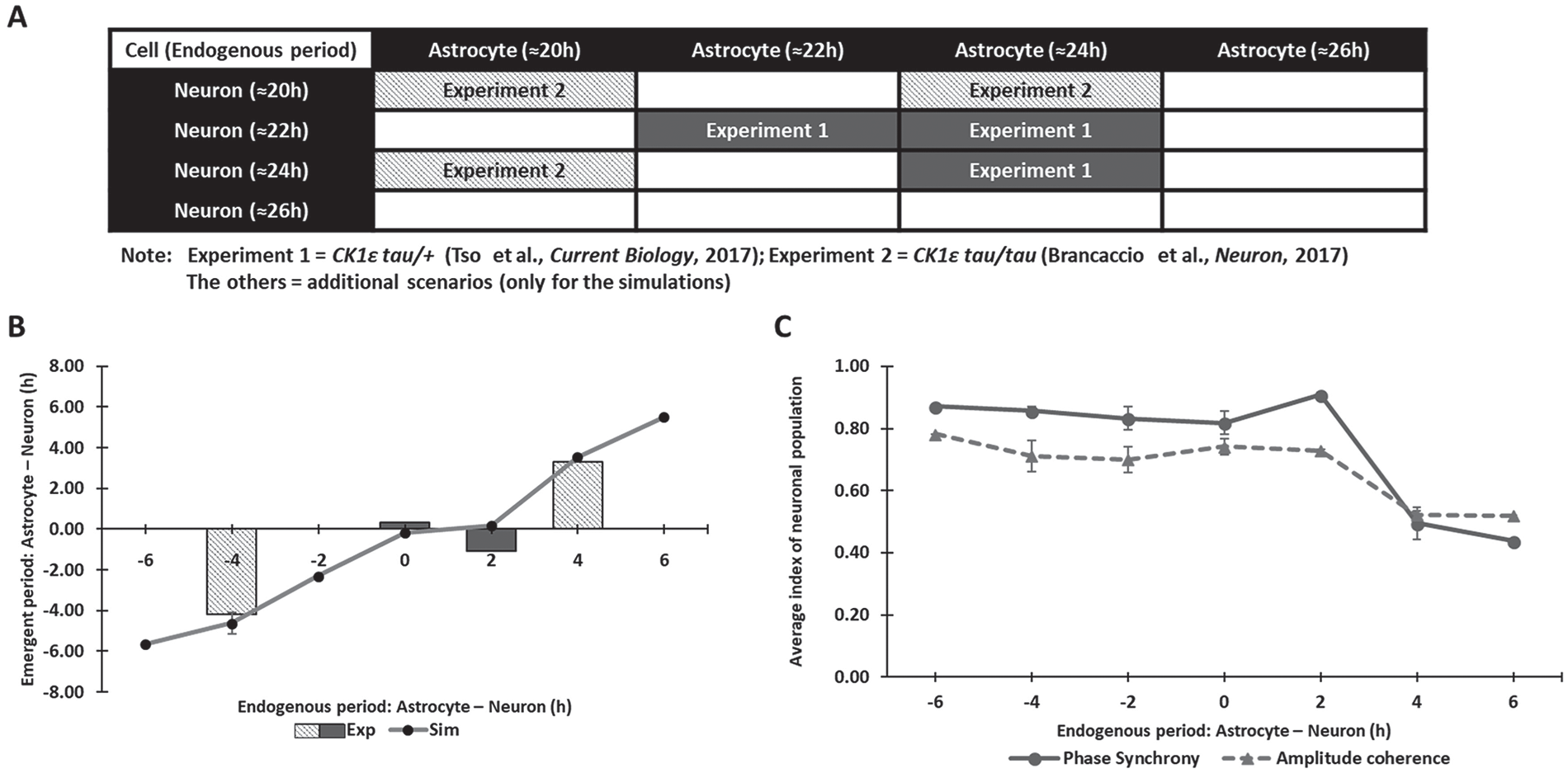
Astrocytic control of the neuronal period depends on the difference between the endogenous neuronal and astrocytic periods. (A) Sixteen simulation scenarios performed to investigate the effect of endogenous periods on the coupled neuronal period. Six scenarios could be compared with available data (Brancaccio et al., 2017; Tso et al., 2017), while the other 10 scenarios were constructed using endogenous periods reported in other studies (Maywood et al., 2011; Patton et al., 2016; St John et al., 2014). (B) The difference between the endogenous astrocytic and neuronal periods (x-axis) plotted against the difference between the coupled neuronal and astrocytic periods (y-axis) for all 16 simulation scenarios and the 6 scenarios for which comparable data were available (Brancaccio et al., 2017; Tso et al., 2017). (C) Neuronal phase synchrony and amplitude coherence measures (mean ± SEM) predicted for the 16 simulation scenarios plotted against the difference between the endogenous astrocytic and neuronal periods.
Our model predicted that differences between the endogenous periods and not the individual periods themselves were the key determinant as to whether the coupled neurons were entrained by the astrocyte (Fig. 3B). We found that the neurons produced an average period close to the astrocytic period when (1) the endogenous astrocytic period was greater than or equal to the endogenous neuronal period and (2) the difference between the 2 endogenous periods was approximately 2 h or less. These 2 conditions were satisfied by 7 of the 16 simulations that produced endogenous period differences of 0 h or +2 h (Fig. 3A, B). For the other 9 scenarios, the astrocyte was not able to entrain the neurons, and the coupled period of the neuronal population was close to its endogenous period. Thus, our model accurately simulated experimental results implicating astrocytes in modulation of the circadian period of the SCN. Furthermore, the model generated experimentally testable predictions that, when both SCN neurons and astrocytes are circadian, their phase synchrony and amplitude coherence will substantially decrease when (1) the endogenous astrocytic period is greater than the endogenous neuronal period and (2) the difference between the 2 endogenous periods is greater than 2 h (Fig. 3C).
Astrocytic Modulation of the Neuronal Population Depends on the Density of Astrocyte-neuron Connections
To investigate the putative impact of network structure on astrocytic modulation of neuronal rhythms, we performed additional simulations with WT cells in which neuron-to-neuron and astrocyte-to-neuron connectivities were varied. Two types of neuronal networks were considered: SW networks as used in Figure 2 and a nearest-neighbor network formulated as in our previous study (Vasalou et al., 2009). Six types of astrocyte-to-neuron networks were constructed by randomly allowing different percentages (0%, 5%, 25%, 50%, 75%, 100%) of all possible connections, with a mean-field connectivity of 100% used in Figure 2. Finally, 2 different period distributions of the uncoupled neuronal population were constructed by varying random perturbation of the Bmal1 transcription rate (vsB) and degradation rate (vmB; Vasalou et al., 2011). A 2% standard deviation produced relatively narrow distributions (PD1; also used in Fig. 2) with uncoupled periods of 22 to 26 h, while a larger 3% standard deviation generated a broader distribution (PD2) with periods of 21 to 29 h. The average number of neuron-to-neuron connections and the total number of neuron-astrocyte connections for all 24 network structures are shown in Supplemental Figure S1.
Our model predicted that the extent of astrocytic modulation depended on the density of neuron-to-neuron and astrocyte-to-neuron connections as well as the degree of neuronal heterogeneity. As the percentage of astrocyte-to-neuron connections was increased from 0% to 100% with the narrow period distribution PD1, the neuronal period was predicted to increase ~1 h for the SW network topologies (Fig. 4A). The nearest neighbor (NN) topologies did not exhibit a similar period increase, but the percentage of rhythmic neurons increased substantially as the percentage of astrocyte-to-neuron connections was increased (Fig. 4B). By contrast, astrocytic modulation was not required to achieve high rhythmicity in the SW networks. Phase synchrony (Fig. 4C) and amplitude coherence (Fig. 4D) of the neuronal population were increased in the SW networks as astrocyte-to-neuron connectivity increased, while the NN networks exhibited low synchrony and coherence independent of astrocytic modulation. While similar predictions were obtained when simulations were performed with the broader period distribution PD2 (Suppl. Fig. S2), astrocytic modulation generally had less impact on cell-to-cell coordination in terms of phase and amplitude. Unlike the increased period for the SW network, the neuronal period was predicted to decrease ~0.5 h for the NN network topologies as the astrocyte-to-neuron connectivity was increased. Uncoupled period distributions that are either narrow (PD1) or broad (PD2) and coupled period distributions for the 24 network structures are shown in Supplemental Figure S3. Collectively, our predictions suggested that astrocytic modulation of the neuronal population within an SW network depended strongly on the density of astrocyte-neuron connections but only weakly on the heterogeneity of endogenous neuronal periods.
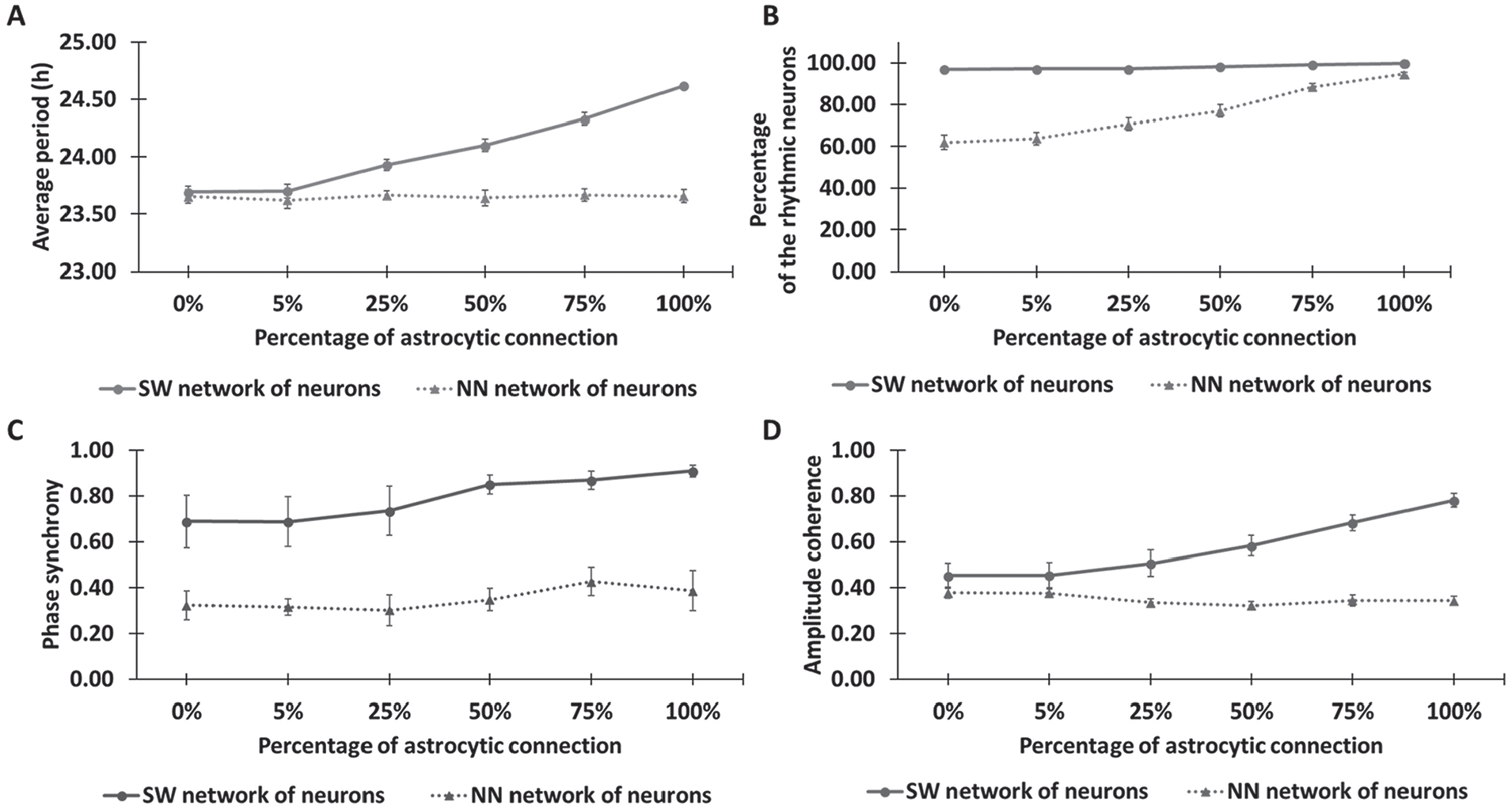
Astrocytic modulation of the neuronal population for different network topologies. SCN models with 2 types of neuronal networks (small world [SW] and nearest neighbor [NN]; Vasalou et al., 2009) and 6 types of astrocyte-to-neuron networks containing different percentages of all possible connections (0%, 5%, 25%, 50%, 75%, and 100%) were constructed and simulated. Four emergent system properties were calculated for each model (mean ± SEM): (A) average period of the neuronal population, (B) percentage of rhythmic neurons, (C) neuronal phase synchrony, and (D) neuronal amplitude coherence. The simulations were based on a narrow neuronal period distribution before coupling (≈22-26 h). Simulation results for a broader neuronal period distribution are shown in Supplemental Figure S2.
Astrocytic Modulation of Neuronal Populations Depends on Mutual Contributions of the Multiple Intercellular Signaling Pathways
To explore the importance of intercellular signaling pathways on astrocytic modulation, we performed simulations with individual VIP, glutamate, and GABA pathways blocked. More specifically, we generated 5 types of models containing either WT cells (no blockade), neurons unable to bind VIP (VIP_N), astrocytes unable to bind VIP (VIP_A), neurons unable to bind glutamate (Glu_N), or neurons unable to bind GABA (GABA_N). For each model type, simulations were performed with cells having no mutations and the 6 experimentally realized neuron/astrocyte mutations explored in Figure 2, generating 35 distinct scenarios.
Simulation results for the 5 models containing neurons and astrocytes without mutations are shown in Figure 5. The loss of VIP coupling between neurons was predicted to cause relatively large decreases in the average period and rhythmicity of the neuronal population (Fig. 5A), consistent with experimental results (Hastings et al., 2014). Interestingly, the loss of astrocyte-released glutamate that binds only to the respective receptors in neurons resulted in a decrease of ~1 h in the average period but had little effect on rhythmicity. The blockade of VIP signaling to the astrocyte or GABA signaling to the neurons had little effect on period or rhythmicity. Phase synchrony and amplitude coherence of the neuronal populations were affected similarly by loss of VIP or glutamate signaling to neurons or VIP signaling to the astrocyte (Fig. 5B). Neither phase synchrony nor amplitude coherence was affected by the loss of GABA signaling in neurons. For WT cells (scenario 1), the model predicted that neuronal VIP and glutamate receptors play dominant roles in the SCN neuron-astrocyte interaction at a synapse.
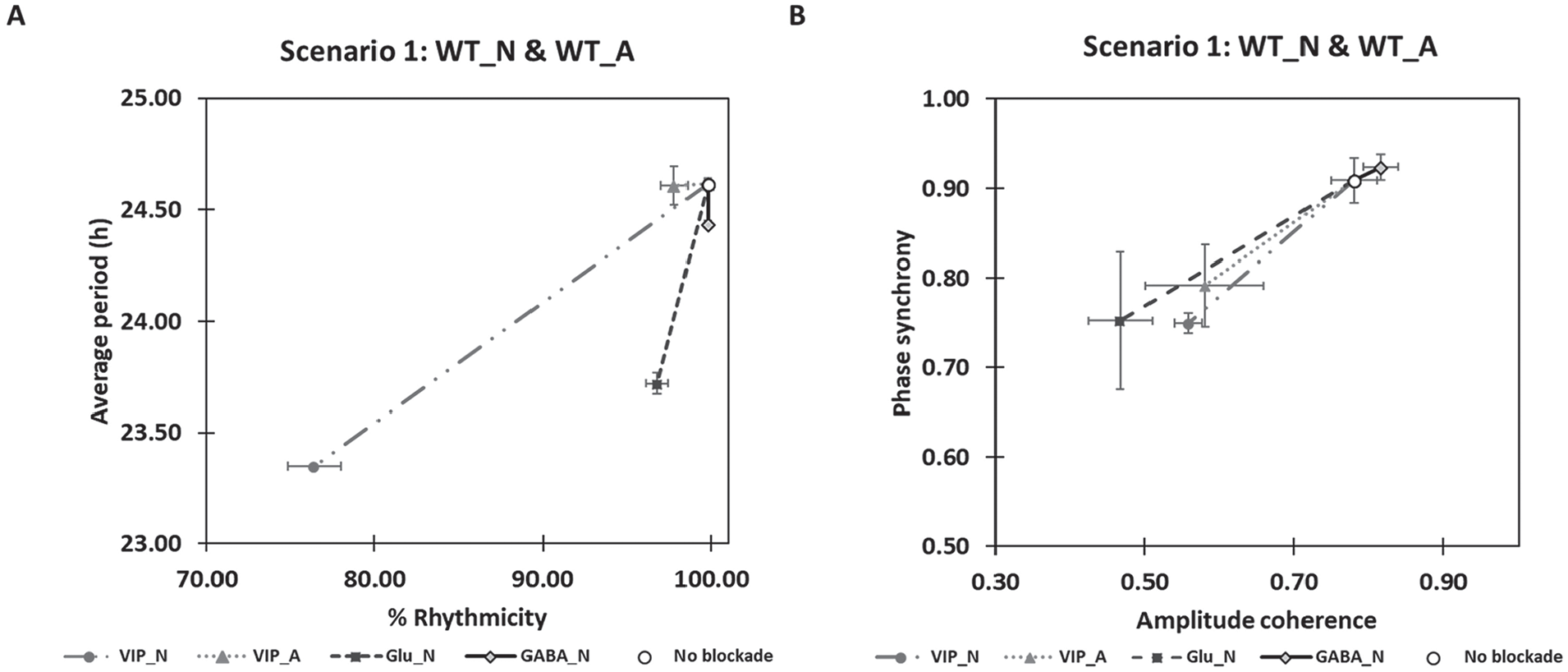
Astrocytic modulation of the neuronal population with different intercellular signaling pathways removed. SCN models containing wild-type cells (no blockade), neurons unable to bind vasoactive intestinal polypeptide (VIP_N), astrocytes unable to bind VIP (VIP_A), neurons unable to bind glutamate (Glu_N), or neurons unable to bind γ-aminobutyric acid (GABA_N) were constructed and simulated. (A) Average neuronal period (mean ± SEM) and percentage of rhythmic neurons (mean ± SEM). (B) Phase synchrony (mean ± SEM) and amplitude coherence (mean ± SEM) of the neuronal population. The simulation results for 6 experimentally realized neuron/astrocyte mutations (scenario 2-7 presented in Fig. 2) are shown in Supplemental Figure S4.
Corresponding simulation results for the 6 experimentally realized neuron/astrocyte mutations (Suppl. Fig. S4) revealed several trends regarding the mutual effect of endogenous periods and intercellular coupling on astrocytic modulation. An astrocyte with shorter endogenous periods (e.g., about 20 or 22 h) was predicted to generate lower percentages of rhythmic VIP-blockade neurons (Suppl. Fig. S4B, G, and I) than the WT astrocyte with a 24-h endogenous period (Fig. 5A). Interestingly, the absence of VIP, which binds to neuronal receptors, allowed neuronal period modulation by an astrocyte with a 4-h longer endogenous period. To illustrate, when VIP-blockade neurons with relatively short endogenous periods of ~20 h were completely coupled with a WT astrocyte with an endogenous period of ~24 h, the neuronal period was reset toward the astrocytic period (Suppl. Fig. S4H), and the neuronal population exhibited higher phase-amplitude synchrony (Suppl. Fig. S4K). By contrast, the blockade of neuron-released VIP that binds to the astrocytic receptors had little impact on neuronal period and rhythmicity (Figs. 5A, S4A-C, and S4G-I) but strongly affected phase synchrony and amplitude coherence of the neuronal population, especially when the endogenous period of the astrocyte was ~24 h (Figs. 5B, S4F, and S4K).
Glutamate and GABA signaling were also predicted to affect the ability of the astrocyte to entrain the neuronal population. Glutamate blockade consistently resulted in period shortening, reduced rhythmicity, and less phase and amplitude coordination in the neuronal population despite the presence of the astrocyte. GABA blockade of neurons caused the neuronal population to exhibit more variable period resetting that depended on the endogenous neuronal and astrocytic periods. Such GABA blockade slightly rescued desynchronization of the neuronal population when the endogenous neuronal and astrocytic periods were similar (Figs. 5B, S4E, and S4J). Taken together, these model predictions suggest that astrocytic modulation of neuronal behavior is dependent on both endogenous period and intercellular signaling; however, the activation of neuronal VIP and glutamate receptors likely has more impact on the overall neuronal rhythms and astrocytic modulation than GABA receptors.
Astrocytes Have Tunable Mechanisms Primarily Mediated by VIP and Glutamate for Modulating SCN Neuronal Period
We investigated the possible roles of signaling to and from astrocytes on SCN circadian rhythms beyond the effect of genetically manipulated neurons and astrocytes (especially for scenarios 2 and 4 in Fig. 2) to better understand neuronal period modulation (Tso et al., 2017). According to our previous computational findings related to the dominant signaling agents (Fig. 5; Suppl. Fig. S4), we selectively varied the coupling strengths of neuronal VIP or glutamate receptors as well as the mechanistic rates related to astrocytic glutamate signaling to resimulate scenarios 2 and 4 in addition to modifying parameters associated with astrocytic Bmal1 for arrhythmicity or PER/CRY stability for different endogenous periods. For example, we increased the degree of coupling strengths (the extent of receptor saturation modeled in To et al., 2007; Vasalou and Henson, 2010, 2011) associated with either neuronal VIP (VIPR2_N) or neuronal glutamate (iGluRs; Fig. 6A). We also specifically enhanced the glutamate synthesis/transport rate regulated by the astrocyte (EAATs) from its nominal values used in this work (Fig. 6A). In the absence of astrocytic rhythmicity (scenario 2, Fig. 6C), our model predicted that these 3 model parameters could be independently tuned to yield similar neuronal period increases, including the ~1-h increase observed experimentally (Tso et al., 2017) when Bmal1-/- astrocytes were combined with WT neurons (Fig. 6A). Simultaneous increase of VIP and glutamate receptor strengths in networks with CK1ε -/+ astrocytes and CK1ε Tau/+ neurons (scenario 4, Fig. 6B) could also produce large neuronal period increases, including the ~2-h increase observed experimentally (Tso et al., 2017). Thus, it is possible that loss of Bmal1 or heterozygous CK1ε Tau mutation of astrocytes may indirectly contribute to enhanced VIP and glutamate signaling activity toward the lengthened neuronal period. Collectively, these model predictions suggest that the astrocytes may have tunable mechanisms potentially mediated by VIP (Marpegan et al., 2009) and glutamate (Brancaccio et al., 2017) for modulating SCN neuronal period.
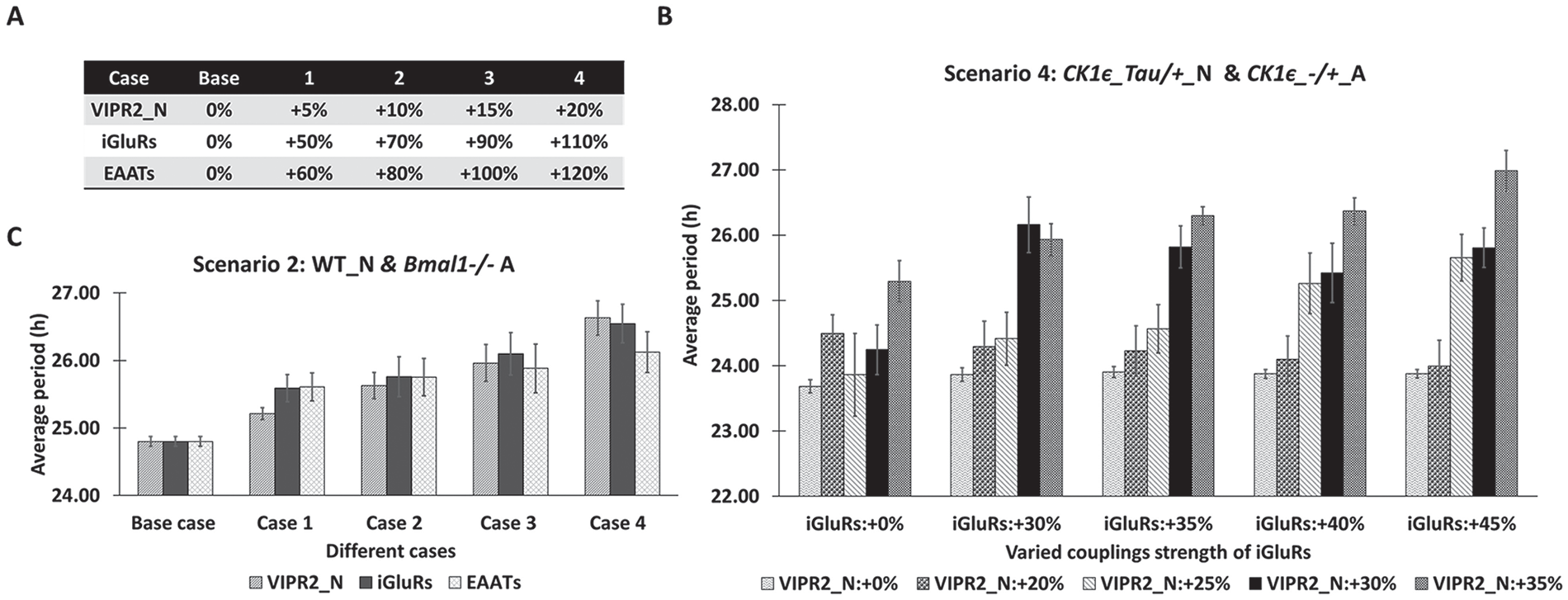
Effect of increased network coupling strengths on the average period of the neuronal population. Model parameters for network coupling strengths associated with neuronal vasoactive intestinal polypeptide (VIPR2_N) and glutamate (iGluRs) receptors and the glutamate synthesis/transport rate regulated by the astrocyte (excitatory amino acid transporters) were increased from their nominal values. (A) Percentage increases in each parameter used to perform simulations of scenario 2 (wild-type neurons and Bmal1-/- astrocytes). (B) Period increases (mean ± SEM) of the simulated neuronal populations for scenario 2. (C) Period increases (mean ± SEM) of the simulated neuronal populations for scenario 4 (CK1ε Tau/+ neurons and CK1ε-/+ astrocytes) and simultaneous increases of VIP and glutamate receptor strengths.
Discussion
Our multicellular model was able to qualitatively recapitulate recent experimental findings (Brancaccio et al., 2017; Tso et al., 2017) demonstrating that astrocytic rhythmicity can alter daily rhythms in the SCN and behavior (Fig. 2). Model simulations with WT and mutant cells helped rationalize the contexts under which astrocytes were dominant and reset the average period of the coupled neurons away from their endogenous periods and specifically toward the endogenous astrocytic period. Based on these simulations, we concluded that astrocytes were able to entrain neuronal populations when the endogenous astrocytic period was greater than or equal to the endogenous neuronal period and the difference between the 2 endogenous periods was approximately 2 h or less (Fig. 3). Our model predictions were generated over a wider range of neuronal and astrocytic endogenous periods than experimentally studied to date (Brancaccio et al., 2017; Tso et al., 2017) and could be additionally evaluated using established methods to generate short-period and long-period mutant cells (Godinho et al., 2007; Maywood et al., 2014; Meng et al., 2008; Siepka et al., 2007; Takahashi, 2016) for future experiments.
Our simulations suggested that the density of astrocyte-to-neuron connections plays a functional role in tuning neuronal dynamics and may augment neuron-to-neuron coupling in circadian timekeeping. More specifically, denser astrocyte-to-neuron connectivity was predicted to result in increased rhythmicity, lengthened period, and enhanced coordination of the neuronal population (Figs. 4; Suppl. Figs. S1-S3). Our model predicted that a denser connectivity also allowed the astrocyte to entrain neuronal populations with broader endogenous period distributions (Suppl. Fig. S3), although the entrainment capability remained limited by the endogenous periods of the 2 cell types. In addition, our simulations indicated that a well-coordinated neuronal network achieved through a combination of neuron-to-neuron coupling and astrocyte-to-neuron connections allowed greater astrocytic modulation of neuronal outputs. Since SCN neurons are known to display a wide range of endogenous periods (Gu et al., 2009), astrocytic modulation through adjustable numbers of astrocytes and connections with the neuron population provides a putative mechanism for enhancing neural coordination and enhancing network plasticity. The predicted effect of astrocytic modulation of neuronal circadian rhythms depending on the network density is also consistent with the experimental finding that a daily synaptic rearrangement of the astrocytic network contributes to a change in rhythmic in the degree of astrocyte-to-neuron connection and facilitates functional alteration of neuronal activity in the SCN (Bosler et al., 2015). When combined with an expanding experimental literature (Barca-Mayo et al., 2017; Brancaccio et al., 2017, 2019; Ruben and Hogenesch, 2017; Tso et al., 2017), our model predictions support the hypothesis that astrocytes provide a complementary means via their synaptic coupling for adjusting SCN neuronal outputs to the extensively studied mechanisms associated with interneuronal coupling (Bernard et al., 2007; Vasalou et al., 2009) and external cues (Komin et al., 2011; Tsumoto et al., 2011; Xu et al., 2012).
Our model predicted that the astrocytic modulation of SCN neurons depended on mutual contributions from multiple intercellular signals (VIP: Marpegan et al., 2009; GABA: Barca-Mayo et al., 2017; Moldavan et al., 2017; and glutamate: Brancaccio et al., 2017; Scofield, 2018) regulated not only by neurons but also by astrocytes. Our signaling-blockade simulations suggested that the astrocytic binding of neuronally released VIP is an essential mechanism for establishing bidirectional communication between the 2 cell types. For instance, when a network was formed with an astrocyte that did not sense VIP but had an endogenous period 2 h larger than that of mutant neurons, the astrocyte was not able to drive the neuronal population to higher periods and synchrony (scenario 4 in Suppl. Fig. S4.) These results suggest that astrocytic sensing and response to rhythmically released VIP by neurons might modulate the neuronal population. Furthermore, our model predicted that loss of rhythmicity in the VIP-blockade neuronal populations cannot be compensated for by bidirectional coupling with an arrhythmic astrocyte because of Bmal1 knockout (scenario 2 in Suppl. Fig. S4), supporting the hypothesis that astrocytic Bmal1 may be essential for astrocytic-modulated neuronal rhythms (Barca-Mayo et al., 2017).
We hypothesized that astrocytes are capable of tuning the circuit-level dynamics of SCN neurons via complex synaptic interactions mediated mainly through excitatory signaling (i.e., VIP and glutamate; Brancaccio et al., 2017, 2019), with inhibitory signaling (i.e., GABA; Semyanov et al., 2004; Yoon et al., 2012) playing a less significant role. Analogous to the functional contribution of VIP, glutamate was predicted to play a dominant role in the astrocytic modulation of SCN neurons (Brancaccio et al., 2017, 2019). For example, an astrocyte having an endogenous period 2 h larger than that of glutamate-blockade neurons was not able to drive the neuronal population to higher periods and synchrony (scenario 4 in Suppl. Fig. S4). Inhibitory GABAergic activity that can be mediated by astrocytes through transformed glutamatergic excitation (Héja et al., 2009, 2012) was predicted to have a less direct role in the astrocytic tuning of neuronal outputs. Together, our computations suggested that multiple intercellular signals functionally coordinate to mediate astrocyte-to-neuron communication at synapses and generate circadian rhythms. Nevertheless, VIP (Marpegan et al., 2009) and glutamate (Brancaccio et al., 2017, 2019; Scofield, 2018) signaling tend to be the primary coupling mechanisms contributing to astrocyte-modulated neuronal rhythms. Our model omits other intercellular coupling mechanisms that may influence astrocytic-neuronal interactions, including excitatory GABA signaling (Albers et al., 2017; Ono et al., 2018) and arginine vasopressin signaling in the shell region of the SCN (Hastings et al., 2018; Honma, 2018).
Our simulations then further demonstrated that increased coupling strengths of neuronal VIP or glutamate receptors or increased glutamate-related activity of genetically modified astrocytes can be possible mechanisms leading to the increased period of neuronal populations modulated by the astrocyte. While arrhythmic or ~24-h period astrocytes did not produce a considerable change in neuronal period when nominal values of coupling strengths were used in our model (Fig. 2), we found that loss of Bmal1 or CK1ε Tau mutation in astrocytes could indirectly enhance coupling strength associated with neuronal receptors and mechanistic rates related to astrocytic signaling activity (Fig. 6). These results possibly rationalized the finding of the lengthened neuronal period observed in recent experiments (Tso et al., 2017). We hypothesized that astrocytic Bmal1 might be functionally associated with changes in glutamate synthesis and transport in astrocytes or the coupling strength of VIP (Barca-Mayo et al., 2017) or glutamate receptors in WT neurons. Furthermore, our model suggested that removal of the CK1ε Tau/+ mutation in astrocytes may increase cellular activities by promoting stronger VIP or glutamate-mediated coupling in mutant neurons. Collectively, we hypothesized that loss of tau mutation or Bmal1 knockout in astrocytes may result in the lengthened neuronal period as experimentally observed (Tso et al., 2017) because such genetic manipulations may be associated with enhanced coupling activities mediated by neuronal VIP and astrocytic glutamate and therefore alter daily rhythms of the neuronal population.
In summary, our computational model supports the emerging view that astrocytes encode circadian information via bidirectional synaptic interactions based on the entrainment of VIP released by neurons and then influence daily neuronal rhythms through release and uptake of GABA and glutamate to adjust the neuronal core clock. We have shown that SCN astrocytes can modulate neuronal populations through multiple mechanisms depending on distinct factors, including intercellular coupling, network topology, and endogenous periods. We believe that such mechanistic models are essential for generating testable predictions that can guide the design of experiments for investigating interactions between SCN neuronal and astrocytic populations and the functional role of these interactions in circadian timekeeping. For example, astrocytes may broadly interact with neurons across the SCN or specifically communicate with an SCN subpopulation (e.g., VIPergic neurons). Furthermore, whether gliotransmitters such as GABA and glutamate affect SCN neuronal activity at the synapse, cell, and circuit level remains obscure. Larger simulated networks containing multiple, heterogeneous astrocytes may be worth considering in future studies. Finally, the effects of external stimuli such as light on the astrocytic modulation of neuronal populations and SCN plasticity require further study (Leone et al., 2015). It would also be intriguing to know whether astrocytes vary their number or neuronal contacts as a function of, for example, age or season, to alter their modulation of neurons.
Supplemental Material
Fig.S1_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S1_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S2_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S2_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S3A-S3C_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S3A-S3C_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S3D-S3G_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S3D-S3G_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S3H-S3J_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S3H-S3J_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S3K-S3N_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S3K-S3N_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S4A-S4C_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S4A-S4C_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S4D-S4F_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S4D-S4F_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S4G-S4I_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S4G-S4I_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S4J-S4L_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S4J-S4L_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S5_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S5_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Fig.S6_no_color – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Fig.S6_no_color for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
JBR19_SUPP_MAT_2nd_Revision – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, JBR19_SUPP_MAT_2nd_Revision for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Supplemental Material
Table_S1_(2nd_revision) – Supplemental material for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling
Supplemental material, Table_S1_(2nd_revision) for Astrocytic Modulation of Neuronal Activity in the Suprachiasmatic Nucleus: Insights from Mathematical Modeling by Natthapong Sueviriyapan, Chak Foon Tso, Erik D. Herzog and Michael A. Henson in Journal of Biological Rhythms
Footnotes
Acknowledgements
The research was supported by the National Institutes of Health (grant U01-EB021956-01).
Conflict Of Interest Statement
The authors have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.