
Abstract
Recent mathematical results for the noisy Kuramoto model on a 2-community network may explain some phenomena observed in the functioning of the suprachiasmatic nucleus (SCN). Specifically, these findings might explain the types of transitions to a state of the SCN in which 2 components are dissociated in phase, for example, in phase splitting. In contrast to previous studies, which required additional time-delayed coupling or large variation in the coupling strengths and other variations in the 2-community model to exhibit the phase-split state, this model requires only the 2-community structure of the SCN to be present. Our model shows that a change in the communication strengths within and between the communities due to external conditions, which changes the excitation-inhibition (E/I) balance of the SCN, may result in the SCN entering an unstable state. With this altered E/I balance, the SCN would try to find a new stable state, which might in some circumstances be the split state. This shows that the 2-community noisy Kuramoto model can help understand the mechanisms of the SCN and explain differences in behavior based on actual E/I balance.
Keywords
All life on earth is adapted to the external 24-h light-dark cycle, in which mammals normally have one bout of activity each cycle and one bout of sleep. However, when hamsters are placed in constant light conditions, this regular pattern of sleep and wakefulness is disturbed, and splitting of the activity bout may occur. In this case, the single period of activity is dissociated into 2 components that ultimately settle in antiphase, effectively producing sleep-wake cycles of approximately 12 h (Pittendrigh and Daan, 1976). It has been shown that this behavioral phase splitting in hamsters and mice has its origin in the suprachiasmatic nucleus (SCN), which is the location of the master clock that regulates the 24-h rhythms in physiology and behavior (de la Iglesia et al., 2000). The SCN is a bilaterally paired nucleus, in which splitting of the behavioral rhythm, caused by exposure to continuous light conditions, induces the left and right nucleus to activate in antiphase (de la Iglesia et al., 2000; Mendoza and Challet, 2009).
Between species, there are differences in phase-splitting behavior. Hamsters show splitting when put in constant light conditions (Pittendrigh and Daan, 1976; de la Iglesia et al., 2000). In a mutant mouse strain, called the CS mouse, rhythm splitting occurs in constant darkness (Abe et al., 1999). Rats and mice can also show split behaviour when subjected to so-called forced desynchrony protocols (Campuzano et al., 1999; de la Iglesia et al., 2004; Casiraghi et al., 2012). Rats were subjected to a 22-h light-dark cycle and mice to a chronic jet lag protocol. It was shown that in the forced desynchrony protocol, the dissociation arises between the ventral and the dorsal part of the SCN. Here, the ventral part follows the external light-dark cycle, and the dorsal part has a period close to the endogenous free-running period of the animal. It is unknown whether these different types of phase dissociation, being a dissociation in phase of the left and right SCN or of the ventral and dorsal SCN, have different underlying mechanisms.
The interaction within the ventral part is mainly based on the neurotransmitter vasoactive intestinal polypeptide, which is excitatory. In the dorsal SCN, the interaction mainly occurs through arganine vasopressin, which is also excitatory in nature. The communication between the ventral and dorsal part of the SCN occurs predominantly through γ-aminobutyric acid (GABA), which can be inhibitory but in the SCN also excitatory (Choi et al., 2008; Farajnia et al., 2014; Olde Engberink et al., 2018). For the left-right distinction, less is known about the communication mechanisms, let alone whether these are excitatory or inhibitory (Michel et al., 2013).
Recently, a variety of models has been proposed to explain why the phase-split state of the SCN occurs. These models have typically tried to modify the standard 2-oscillator models for the SCN by including time delays in the coupling (Indic et al., 2008), assuming large variation in the coupling strengths (Schroder et al., 2012) or taking the intracommunity coupling to be negative (Oda and Friesen, 2002). The model proposed by Oda and Friesen (2002) models each community of the SCN as a single oscillator and connects these using coupled Pittendrigh-Pavlidis equations. They consider coupled identical oscillators as well as nonidentical oscillators, making a distinction between morning and evening oscillators. Indic et al. (2008) considered a model most similar to the one we will consider. They included the possibility of a delay in the interaction between oscillators and also observed that a nonuniform network structure, which has at least 2 communities, is essential to realize the phase-split state. Their work included analysis of the stability of the phase-split state. The article by Schroder et al. (2012) used a model proposed by Leloup et al. (1999) and Gonze and Goldbeter (2006) and applied this to 2 groups of 100 oscillators. The results were based on numerical simulations and showed that the split state can arise without a change in the structure or strength of the interactions between oscillators but rather by a change in the circadian properties of individual oscillators. Only Schroder et al. (2012) addressed different transitions to the phase-split state, but they did this by looking at the interplay between light intensity and the interaction strength parameters of the individual oscillators. We use a different model, more similar to the model used in Indic et al. (2008). In our model, the external driving is implicit in the change of interaction strength parameters. This model allows us, for the first time, to fully identify the phase diagram to show which transitions are possible for which parameter values.
In this article, we interpret recent findings on the phase diagram of the noisy Kuramoto model on a 2-community network (Meylahn, 2019) in the context of the phase-split SCN, in which the left and right SCN, or the ventral and dorsal part of the SCN, dissociate into 2 antiphasic neuronal communities, instead of using a 2-oscillator model. In contrast to previous studies, in which additional parameters were necessary to obtain a phase-split state, here only the 2-community structure of the network is required. This enriches the model significantly in comparison with the original 1-community version. This is a surprising finding in itself, as the modification to consider the model on a 2-community network seems almost trivial a priori. Not only does this model exhibit the phase-split state, but it also exhibits a bifurcation point in the possible states it can take (shown as separate lines in the phase diagram in Fig. 1), which determines the existence of a nonsymmetrically synchronized state. This might explain the different transitions to the phase-split state observed in the experiments. We investigate the stability properties of the various states by using simulations of the system and find that the system might have to pass through the nonsymmetrically synchronized solution when it is above the bifurcation point.
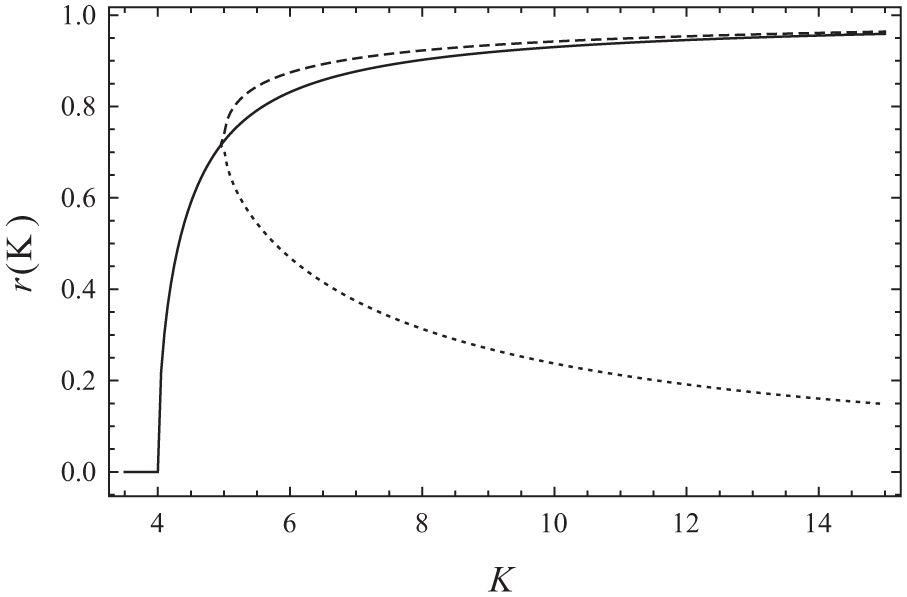
Solutions of the self-consistency equations (5) and (6) for different values of K while
Model
To model the SCN, we modified the noisy Kuramoto model by placing it on a 2-community network structure. Each community consists of N oscillators, which correspond to neurons in the SCN. Oscillators in the same community interact with a strength
We will denote the phase of the oscillators (which can be between 0 and 2π) in the first community by
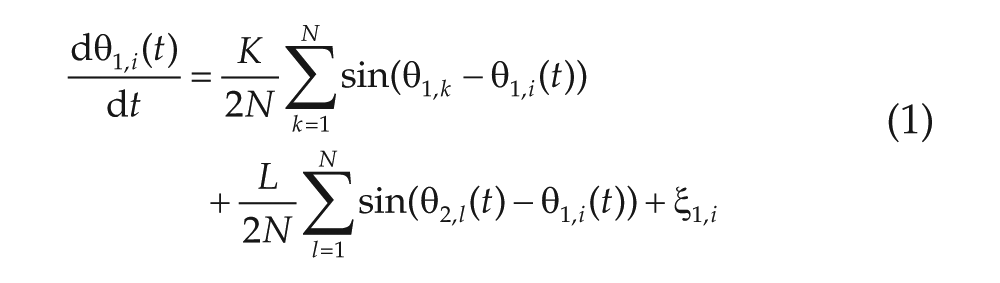
and
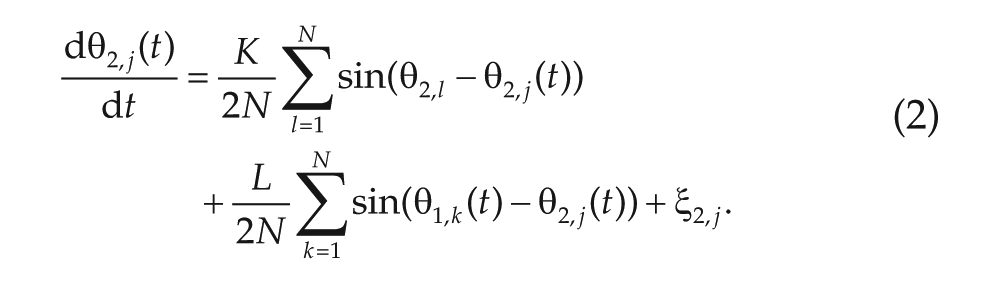
Here,
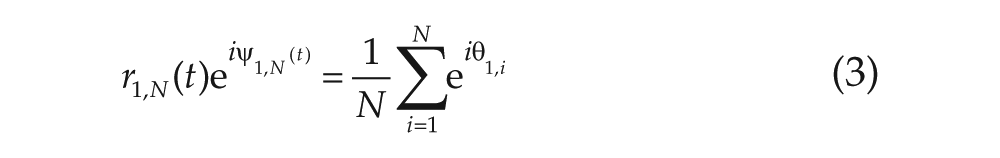
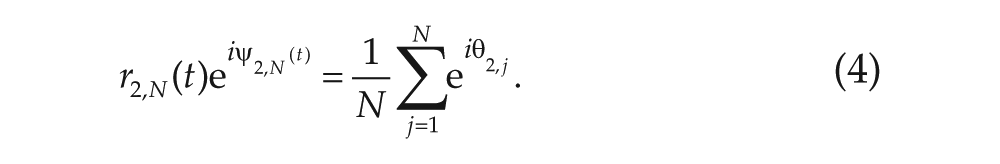
The synchronization levels
When taking the limit of the number of oscillators going to infinity, we see that the system can be described by a probability distribution in each community, namely,
Note in this respect that in the circadian field, the synchronization term is often treated differently. Where mathematical phase indicates the state of one oscillator at a specified time point, in the circadian field, the time of a certain state is taken for each oscillator, such as peak time. Thus, synchronization in the mathematical sense indicates a synchronization in oscillator state, while in the circadian sense, it indicates a synchronization of the oscillators in time. Thus, we define the circadian synchronization here as time synchronization.
In the long-time limit, the distributions reach a steady state (a state in which the order parameters are stationary) that can be described analytically (see Meylahn, 2019). Which values the order parameters
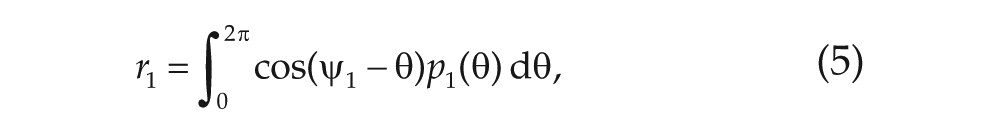
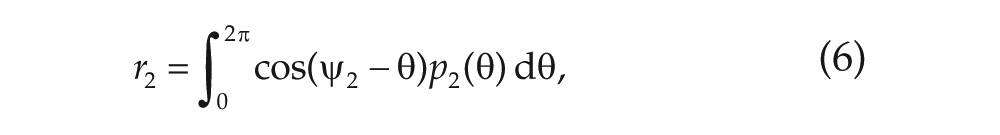
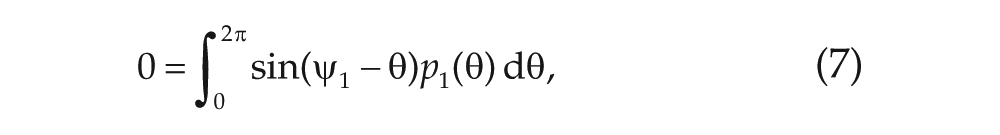
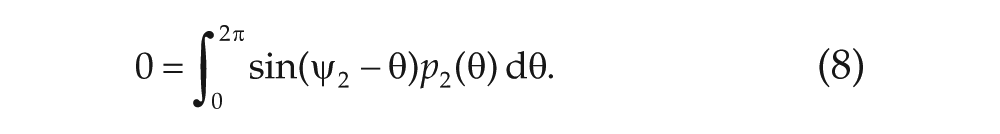
Because of the invariance of the system under rotations, one of the average phases can be set to zero (i.e.,
The point at which the nonsymmetric solution bifurcates from the symmetric solutions gives a line in the phase diagram. In Figure 2, we plot the phase diagram given that the average phases are aligned (i.e.,
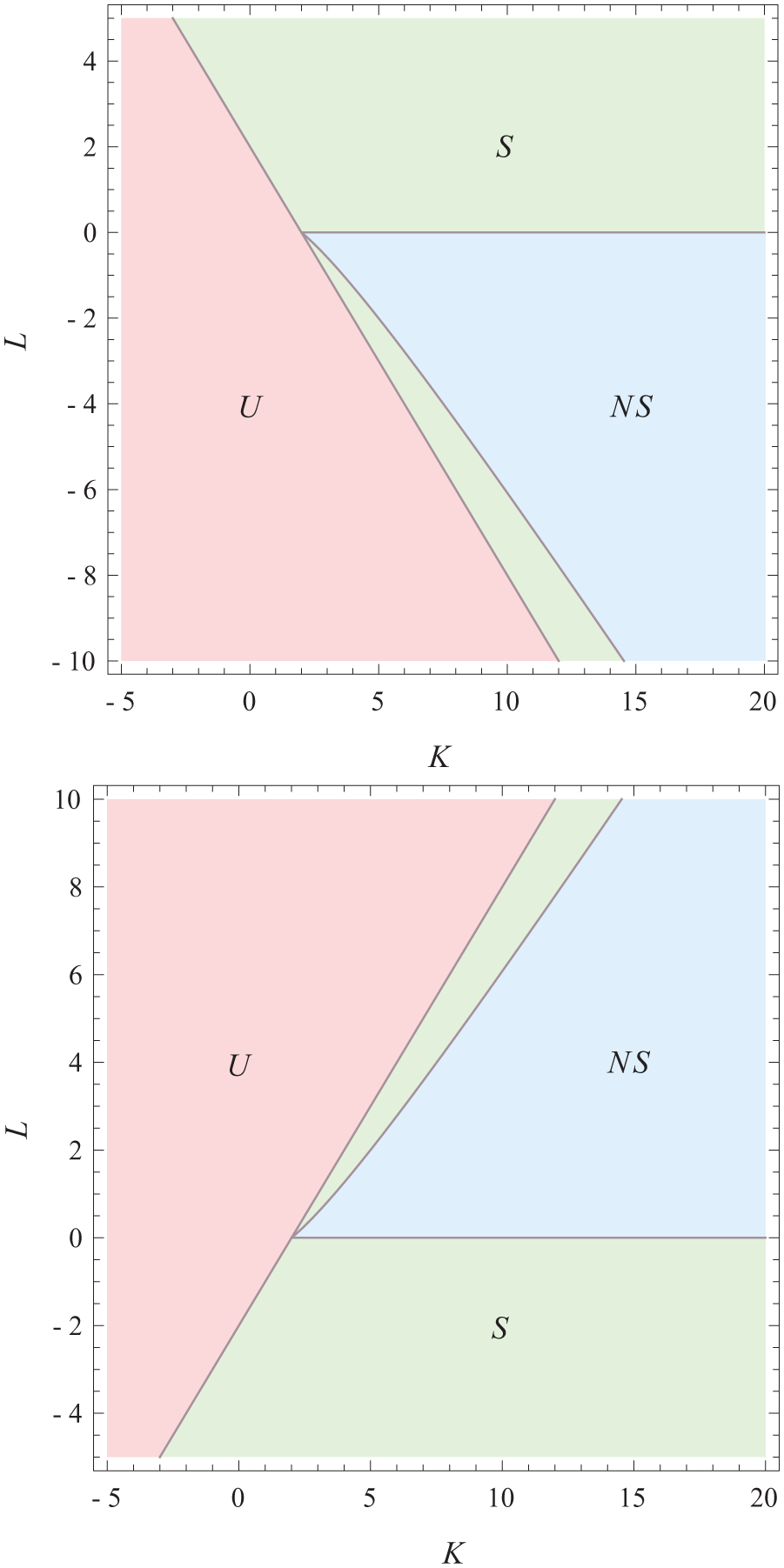
Phase diagram in which the average phases are aligned (top) and in which they are anti-aligned (bottom). In the region labeled
Note that, depending on external conditions, the circadian system can move through this phase diagram since the interaction strength parameters might change because of extreme external conditions. Because of the conjecture in Meylahn (2019) about the possible phase differences in the steady state, we assume that these are the only 2 phase diagrams for the steady state.
The analysis done in Meylahn (2019) and summarized here concerns the stationary points of the system of Eqs. (5) to (8) for the order parameters, in which each solution corresponds to a pair of stationary distribution profiles,
Simulations
We can simulate the system to investigate the stability properties of the various solutions. We summarize the observed stability properties in Table 1. What is meant by metastable here is that the system can, if prepared correctly, stay in this state for a long time but will eventually move to a stable stationary state (it does not refer to the mathematical definition of metastability). Intuitively, this table makes sense. If the interaction between communities is positive, we can, roughly speaking, say that the 2 communities attract one another. If the system is in a state in which the average phases are anti-aligned, with both communities sufficiently synchronized, it can stay there for a while but would ultimately prefer to be in a state with the phases being aligned. This is because the mean-field Kuramoto model corresponding to the aligned state has interaction strength parameter
Stability of possible solutions when

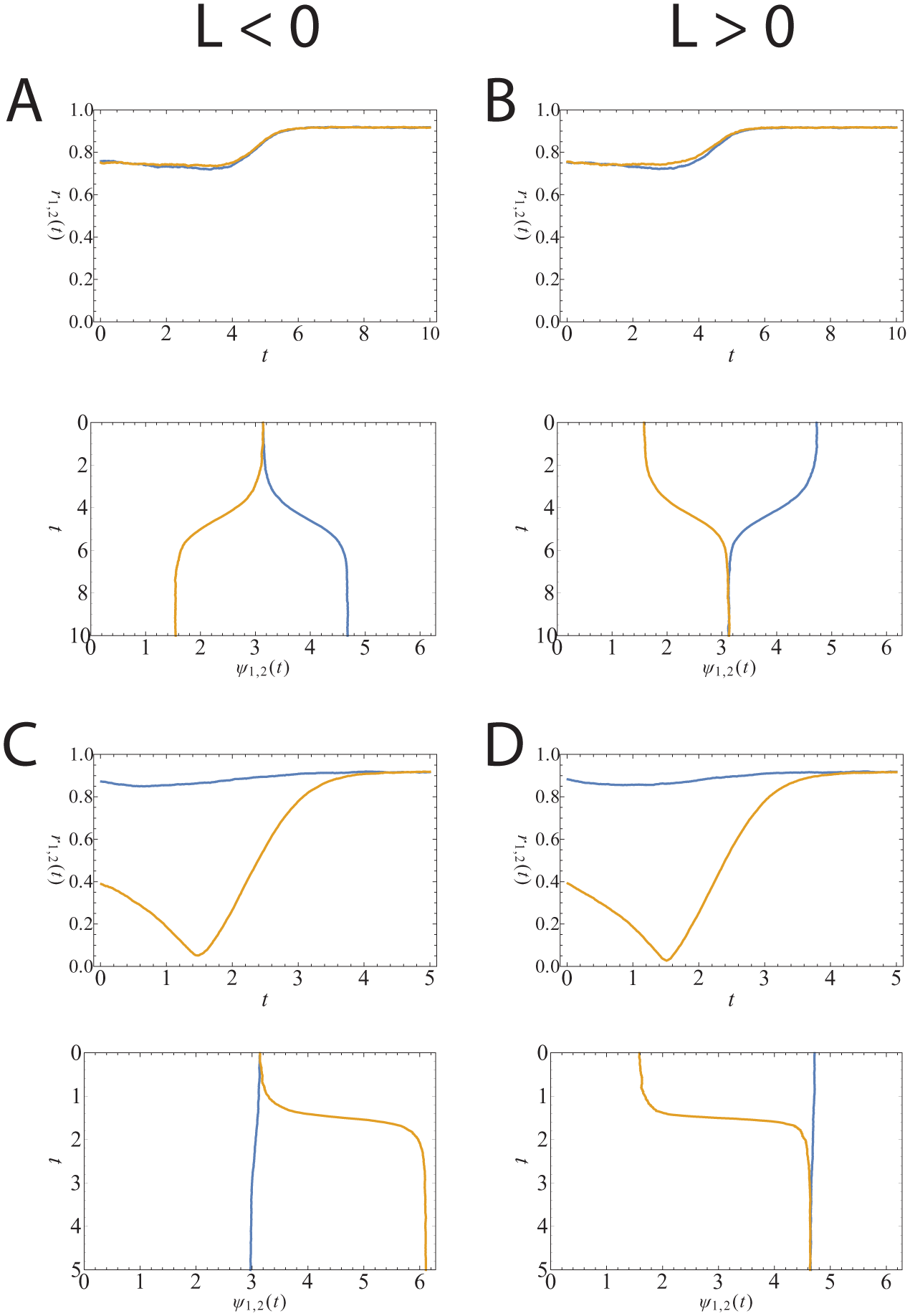
Simulation of 10,000 oscillators per community with
The simulations are of 10,000 oscillators per community. In the first simulations (Fig. 3A), we initialize the 2 communities to have approximately the same synchronization level (upper frame) and with the average phases of the 2 communities aligned (lower frame). We take
In the second simulation (Fig. 3C), we again initialize the average phases to be aligned, but this time, one community is more synchronized than the other (upper frame). We take
Discussion
The metastable anti-aligned state discussed in the previous section might be the state observed in hamsters displaying 2 periods of activity in a single 24-h cycle (Pittendrigh and Daan, 1976).
The underlying cause of the dissociation in phase, both phase splitting of the left and right SCN as well as through forced desynchronization of the ventral and dorsal SCN, seems to be that a 2-community structure must be present in the SCN. As these communities interact with each other, in normal circumstances they remain synchronized. However, in particular conditions, these 2 communities can arrive in the metastable anti-aligned state. This state is, as described, metastable, so after some time, it will return to the stable synchronized state. However, it appears that when keeping animals in a forced desynchronization protocol, this metastable state can be maintained, even though the SCN is inclined to return to the stable synchronized state.
Apparently, according to Figure 2, the circadian system of animals is normally either in the green (
Changes in external conditions may affect the ability to synchronize by changing the interaction strength
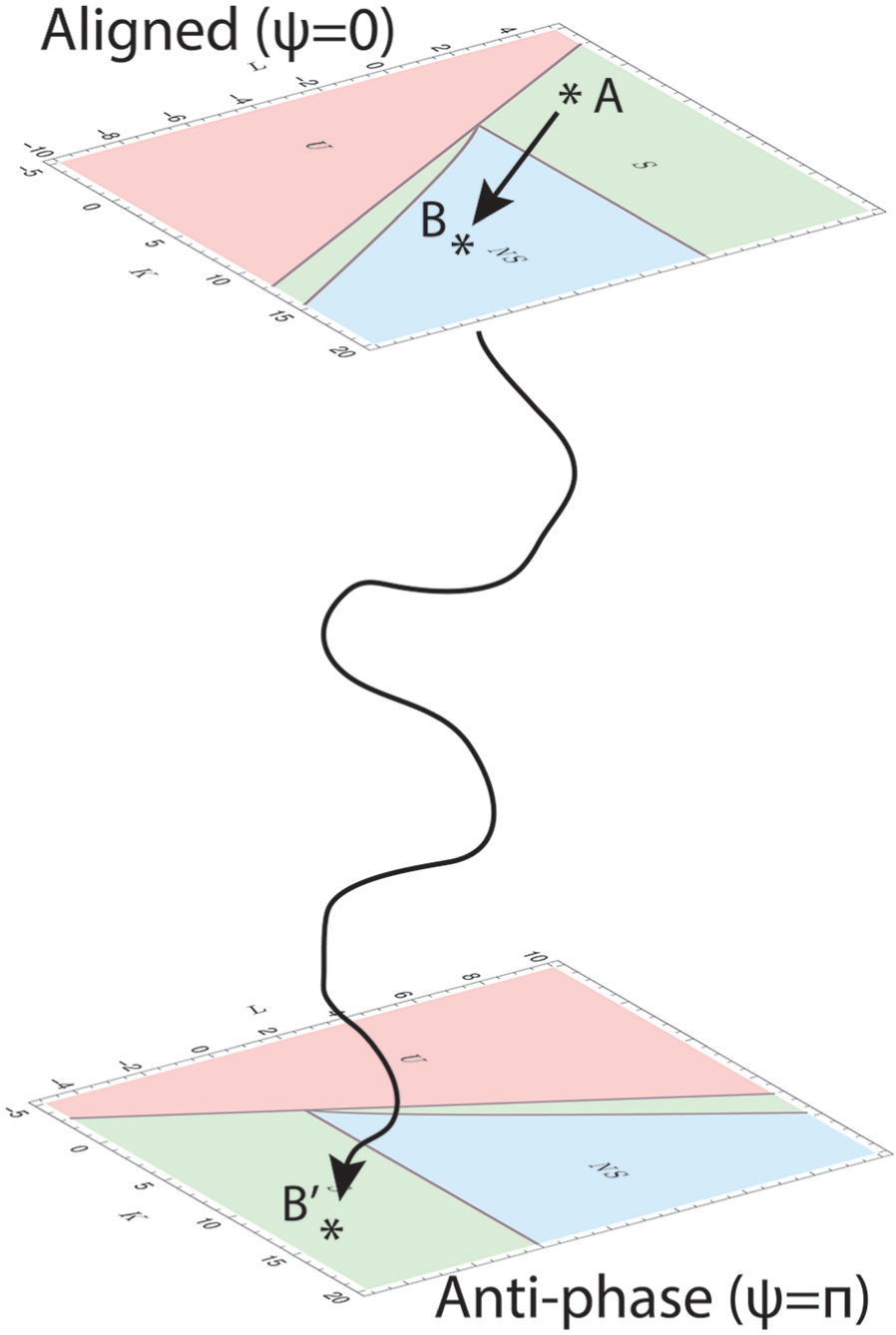
Mechanism of the suprachiasmatic nucleus changing states. A disturbance in the external conditions changes the excitation-inhibition (E/I) balance of the system, meaning that the parameters
Phillips et al. (2019) recently showed that there are interindividual differences in light sensitivity between subjects, which may affect sleep and circadian timing. These interindividual differences in light sensitivity may also be the reason that not all animals will arrive in the split state when subjected to constant light or a forced desynchrony protocol. The mechanism that we describe here may explain why some animals do end up in the split situation while others do not. The interindividual differences may cause the change in E/I balance to be a little different among different animals. Thus, each animal would arrive in a different location B in the aligned space in Figure 4. Some of these locations may be stable, while others may be unstable. When an animal arrives in an unstable state, there is a good chance it will end up going to the antiphase plane. Gaining a better understanding of the influence of external light on the E/I balance of individual animals will provide more details on the state space by which the animals are governed.
Studying the dynamics of the system between the various states mathematically is difficult. Simulations of the system give an impression for what might be typical behavior, and they might be relevant for understanding the unpredictable response of the SCN to the different transitions to and from the phase-split state when animals are exposed to constant light or forced desynchronization protocols. To better study the trajectories that the system may take, jet lag studies may be employed to collect data on single cells and review the dynamics between the ventral and dorsal communities. In jet lag experiments, a dissociation between the ventral and dorsal SCN is also observed. Albus et al. (2005) showed that when a phase shift is applied to the SCN, the ventral SCN shifts immediately with the new light-dark cycle, while the dorsal SCN lags behind. The dissociation observed between both communities is dissolved after 6 days (Rohling et al., 2011). Using the jet lag protocol, the dynamics of both communities could be studied in subsequent days.
Conclusion
This article is novel in the fact that it points out the existence of the bifurcation point in the 2-community noisy Kuramoto model and notes that this might be the reason for the observation of different transitions to the phase-split state of the SCN. To prove this is mathematically challenging, since that would require studying the dynamics between different states of the 2-community noisy Kuramoto model. There is also no experimental work measuring the activity of individual neurons in the phase-split state or during the transition into it to which one could compare the simulations done here. The conclusion of this article is then that the 2-community noisy Kuramoto model has more to offer in terms of explaining the behavior of the SCN.
The table predicts that the stable state of the system is the anti-aligned state when
This does not invalidate the current model, however, as the main message we want to bring forward is that a change in external circumstances brings about a change in the coupling variables within and between 2 communities, thus changing the E/I balance of the system. This change in E/I balance may move the system into an unstable or metastable state, and the system will search through its state space for a stable state based on the changed E/I balance.
Footnotes
Acknowledgements
We thank Stephan Michel for valuable comments on earlier versions of the article. This work was supported by Velux Foundation, project grant 1029. J. M. Meylahn is supported by NWO Gravitation Grant 024.002.003 NETWORKS.
Conflict of Interest Statement
The authors have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.