
Abstract
Many human action sequences, such as speaking and performing music, are inherently rhythmic: Sequence events are produced at quasi-regular temporal intervals. A wide range of interindividual variation has been noted in spontaneous production rates of these rhythmic action sequences. Dynamical theories of motor coordination suggest that individuals spontaneously produce rhythmic sequences at a natural frequency characterized by minimal energy expenditure and maximal temporal stability, relative to other frequencies. We tested this hypothesis by comparing the temporal variability with which musicians performed rhythmic melodies at their natural spontaneous rate with variability in their performances at faster and slower rates. Musicians’ temporal variability was lowest during performances at their spontaneous rate; in addition, performers’ tempo drift during trials at other rates showed bias toward their spontaneous rate. This study provides the first direct evidence that spontaneous rates of motor coordination represent optimally stable natural frequencies of endogenous rhythms.
Keywords
Many human action sequences, such as speaking and music performance, are inherently periodic; sequence events, such as syllable onsets in speech, or tone onsets in music, are produced at semi-regular temporal intervals. These periodicities are produced spontaneously for a range of actions, including finger tapping (Fraisse, 1982; Collyer et al., 1994; McAuley et al., 2006), walking (Murray et al., 1964), speech (Jungers et al., 2003), and music performance (Loehr and Palmer, 2011; Zamm et al., 2015, 2016). Evidence suggests that spontaneous periodicities of human action sequences vary widely across individuals and can change over the life span (McAuley et al., 2006). These patterns are consistent with several endogenous human rhythms, from body temperature to metabolic rate and day-sleep cycles (Crowley and Eastman, 2015; Rising et al, 1992; Roenneberg et al, 2003a, 2016), suggesting that spontaneous periodicities, or natural frequencies, present in human action sequences may be considered a type of endogenous rhythm. Unlike the attention given to endogenous rhythms such as metabolism or sleep cycles, little research has investigated natural frequencies of human action sequences.
Dynamical systems theories of temporal coordination posit that the natural frequency of a given system is the frequency at which minimum energy is required to coordinate motion between parts of the system, such as knee and ankle joints during walking (Bernstein, 1967; Haken et al., 1985; Kelso, 1997; von Holst, 1973). Evidence suggests that when a biological system is perturbed away from its natural frequency, motion variability increases and the system eventually returns to its natural frequency (von Holst, 1973; Kelso, 1997). These predictions have been widely tested in the context of simple biological motion such as fish fin movement (van Holst, 1937) and spontaneous human limb movement (Rosenblum and Turvey, 1988). However, it is not clear whether dynamical accounts can explain natural frequencies of more complex motor behaviors.
Natural frequencies can be measured in the context of music performance, a complex motor behavior that is inherently periodic. Musicians show natural frequencies of tone onset production that vary widely across individuals but are stable within individuals over time, limbs, and simple melodies (Zamm et al., 2016). Although musicians are trained to perform music at a range of frequencies, natural frequencies appear to constrain musicians’ ability to perform with others; duet partners matched for natural frequency, as measured by spontaneous rates of solo performance, are more synchronous than mismatched partners (Zamm et al., 2016), and the faster partner tends to anticipate the tone onsets of an external stimulus relative to the slower partner (Loehr and Palmer, 2011; Zamm et al., 2015). This increased coupling between individuals or organisms with similar natural frequencies is seen in many biological systems, ranging from fireflies (Hanson, 1971) to humans walking side-by-side (Nessler and Gilliland, 2009), as well as between individuals’ endogenous rhythms and rhythms present in the environment (Dodd et al., 2005; Ouyang et al., 1998).
The current study tests the dynamical hypothesis that musicians’ natural frequencies—the rate at which they spontaneously produce tones in a given music performance—represent an optimal mode of coordination relative to performance at other frequencies. Optimality in neural computation refers to the optimal satisfaction of simultaneous constraints (Smolensky and Legendre, 2006). An optimal mode of coordination is empirically measured here by temporal stability of performance. Trained pianists performed a familiar melody at their natural frequency and at faster and slower frequencies in two tasks; an uncued self-paced task (No Cue) and an initially cued task (Initial Cue). In the No Cue task, pianists performed at a comfortable, natural rate (their natural frequency) and then at self-determined faster and slower rates. In the Initial Cue task, pianists performed the melody following the presentation of an initial metronome-cued rate, set to the rates that they had performed in the No Cue task; the metronome cue turned off when pianists started performing. If pianists’ natural frequencies represent an optimal mode of motor coordination, then the temporal stability of pianists’ tone onset production—defined in terms of minimal temporal variability and temporal drift—should be greatest in performances at the natural frequency relative to other frequencies. Furthermore, comparisons of temporal stability across No Cue and Initial Cue tasks allow us to assess whether natural frequencies represent an optimal coordination state independent of whether the action sequence is spontaneously (intrinsically) generated or externally cued.
Method
Participants
Twenty-four trained pianists (20 female, 22 self-reported right-handed) between the ages of 18 and 30 (mean age = 20.6 years, range = 18-27 years) were included in the study. Criteria for inclusion were at least 6 years of private instruction on the piano (mean years of training = 9.7, range = 6-17 years), no history of psychiatric or neurological conditions, and normal hearing (≤20 dB binaurally), as assessed in a standard pure tone audiometry screening (125-750 Hz). The McGill Research Ethics Board approved the study protocol, and participants provided written informed consent according to the Declaration of Helsinki. All participants reported prior familiarity with the stimulus melody with the exception of two individuals for whom the question was accidentally omitted. Additional participants failed the audiometry screening (n = 2) or failed to perform the melody correctly (n = 3) and were therefore excluded from the study.
Stimulus Materials
Pianists performed a simple familiar melody, Frère Jacques (Brother John). The melody was primarily isochronous (20 quarter notes, 8 eighth notes, 4 half notes) and was 8 measures in duration (in binary 4/4 meter). The melody was notated to be performed with the right hand. The musical notation was emailed to pianists prior to their arrival in the laboratory with the instruction to memorize the melody according to the provided notation, using the right-hand, as is customary for this melody performed on the piano. Participants were also administered a brief musical background questionnaire asking for their age, years of training, age of onset of musical training, and other questions about their past and current involvement in musical activities.
Equipment
Pianists performed the melody on an electronic keyboard (Roland RD-700SX, Shizuoka, Japan) in a sound-shielded room. Software used to control auditory feedback associated with pianists’ keystrokes (Finney, 2001) was run on a Linux operating system (Fedora 14, Raleigh, North Carolina) and recorded Musical Instrument Digital Interface (MIDI) keystroke information from the keyboard. Tones associated with pianists’ keystrokes were sounded with a tone generator (Roland SD-50) using a piano timbre (GM2 sound bank). A metronome pacing cue was sounded with a woodblock percussion timbre (Rhythm 001 from the Roland SD-50 tone generator). Participants heard their performances and the metronome cues over studio headphones (AKG K271), delivered from a headphone amplifier (Behringer Powerplay Pro8, Austraße, Weilheim an der Teck).
Study Design
Two independent variables were manipulated in a 2 × 5 within-subject design: Performance Task (No Cue, Initial Cue) and Rate (5 levels). The two performance tasks were No Cue (pianists performed the stimulus melody at a self-determined pace) and Initial Cue (pianists performed the stimulus melody at the pace of a metronome cue that was calibrated to their rates from the No Cue task). In each of these tasks, pianists’ performance rate was manipulated in 5 conditions: spontaneous production rate (SPR), during which pianists were instructed to perform the stimulus melody at their most natural and comfortable performance rate; Fast (pianists were instructed to perform at a rate faster than the SPR); Faster (pianists were instructed to perform at a rate faster than the Fast condition); Slow (pianists were instructed to perform at a rate slower than the SPR); and Slower (pianists were instructed to perform at a rate slower than the Slow condition). The order of the rate conditions was counterbalanced across pianists: Half of the pianists completed order A (SPR, Fast, Faster, Slow, Slower), and half completed order B (SPR, Slow, Slower, Fast, Faster). For each pianist, the order of rates was fixed across No Cue and Initial Cue tasks.
Study Procedure
Screening tests
Participants were tested in the Sequence Production Lab at McGill University during daytime hours (0900-1700 h); the testing time of day is examined in subsequent analyses. Pianists first completed an audiometry screening test to ensure normal hearing. Pianists who passed the audiometry screening test subsequently completed a melody memory test to ensure that they had memorized the stimulus melody sent to them in advance of their appointment. Pianists were instructed to perform the melody from memory without making pitch errors and were given up to two attempts to pass the memory test.
Experimental tasks
Pianists who passed the screening tests completed two performance tasks. In the No Cue task, pianists were instructed to perform the melody at a natural and comfortable rate, as a measure of their spontaneous rate (SPR-01). Then, pianists who completed order A were instructed to perform the melody at a rate faster than their SPR (Fast) and then at an even faster rate (Faster); pianists who completed order B were instead instructed to perform the melody at a rate slower than their SPR (Slow) and then at an even slower rate (Slower). After completing the No Cue conditions, pianists took a 5-min break to complete the musical background questionnaire. After the break, pianists were instructed to perform the melody again at their spontaneous rate (SPR-02 condition). Then, pianists who completed order A performed the Slow and Slower conditions, and pianists who completed order B performed the Fast and Faster conditions. Within each condition, pianists completed 1 practice trial and 3 test trials, where each trial comprised 4 continuous repetitions of the stimulus melody. All trials were performed from memory (without the music notation).
In the Initial Cue task, the cued metronome rates were based on each pianist’s mean performance rate computed for each corresponding rate condition in the No Cue task. Each performance rate was defined as the mean tone interonset interval (IOI) computed across the middle 2 repetitions of No Cue trials that did not contain pitch errors, given previous work indicating maximal temporal stability in the middle of a performance (Loehr and Palmer, 2011; Palmer et al, 2013; McAuley et al., 2006). In the Initial Cue task, pianists were initially cued by a metronome that sounded for 8 pulses and then stopped, indicating the signal for the participants to begin performing. Because the metronome cue for each Initial Cue task was calibrated to match each participant’s mean rate in the equivalent No Cue Rate condition, the only difference between tasks was the source of pacing cue (internal or external). Pianists again completed 1 practice trial and 3 test trials for each rate condition. Pianists completed the Initial Cue rate conditions in the same order as in the No Cue task.
Data Analysis
Performance errors
Analyses were conducted on the MIDI piano keystroke measures. Pitch errors were first assessed by comparing pianists’ performances with an ideal performance in MATLAB (The MathWorks, Natick, MA) (Large and Rankin, 2007). Because music performance timing is disrupted by pitch errors such as added and deleted tones, melody repetitions containing added or deleted tones were removed from analyses (3.85% of total repetitions were excluded across all participants; mean pitch error rate = 0.16%, SD = 0.15%). After pitch errors were removed, timing errors were identified. First, IOIs were computed at the quarter-note level by excluding eighth notes that occurred between beats and interpolating half notes. Half notes were linearly interpolated and rounded to the nearest integer. This procedure yielded 32 onsets per melody repetition (the last melody repetition in each trial contained 30 onsets due to the final half note, which was not interpolated). Pianists’ performances were then analyzed for timing errors, defined as IOIs exceeding 3 standard deviations of the mean within a given condition. Identified timing errors were excluded from analyses (mean timing error rates = 0.52%, SD = 0.27%).
Dependent variables
Three measures of pianists’ temporal coordination were assessed. Performance rate was measured by the mean IOI at the quarter-note level, computed for each trial. The mean IOI provides a measure of the natural frequency of each performance and allows us to assess whether pianists followed experimental instructions to perform at different rates across conditions. The temporal stability of performance was measured by the coefficient of variation (CV = standard deviation (IOIs) / mean IOI). The CV, commonly used in motor tasks, provides an inverse measure of temporal stability (Hamilton et al., 2004; Richardson et al., 2007; Keller and Repp, 2005); the smaller the CV, the greater the stability.
Finally, the drift in performance rate within each trial was measured by the slope of IOIs regressed over melody serial positions, from beginning to end of each performance (n = 126). The slope measures change in IOI duration (milliseconds) across successive tone onsets in a music performance and indicate whether pianists drift over time toward their spontaneous rates. Positive slope values indicate that IOIs increase in duration over a performance, as would be expected when pianists perform at rates faster than their spontaneous rate; negative values indicate that IOIs decrease in duration over a performance, as would be expected when pianists perform at rates slower than their spontaneous rate. Because temporal drift is common in spontaneous production of auditory sequences by humans (Madison and Delignieres, 2009; Yamada, 1995) as well as by birds (Chi and Margoliash, 2001; Tachibana et al, 2015), the drift recorded in each individual’s SPR condition was treated as a baseline for slope analyses, and mean slope values in all other rate conditions for each pianist were adjusted by the mean slope in their SPR condition (slope in given condition – slope in SPR condition). To ensure that the time windows were sufficiently long to measure temporal drift, slope was computed only for error-free trials (n = 126 serial positions per trial); 4 subjects’ data contained pitch errors across trials of at least one rate condition and were therefore excluded from slope analyses (n = 20 participants in all slope analyses). Finally, participant means for each condition were obtained by averaging across trial means for each dependent measure.
Results
Spontaneous Production Rates
Figure 1 shows the range of pianists’ spontaneous performance rates in the No Cue task (mean IOI, averaged across SPR-01 and SPR-02); values ranged by more than 100% from the fastest to the slowest spontaneous rates, consistent with previous literature indicating a wide range of natural frequencies in music performance (Loehr and Palmer, 2011; Zamm et al., 2015, 2016; Zamm et al., 2017). To confirm that each pianist’s SPR was stable over the experiment, we computed a paired-sample t test comparing spontaneous rates across SPR-01 and SPR-02 conditions: Results indicated that means in these two conditions did not differ significantly (t23 = 1.15, p = 0.26). We also correlated pianists’ rates in the SPR-01 and SPR-02 conditions: Rates were highly correlated across these conditions (r22 = 0.75, p < 0.001). These results confirm the stability of SPRs within individuals, and IOIs were averaged across the SPR-01 and SPR-02 conditions in subsequent analyses.
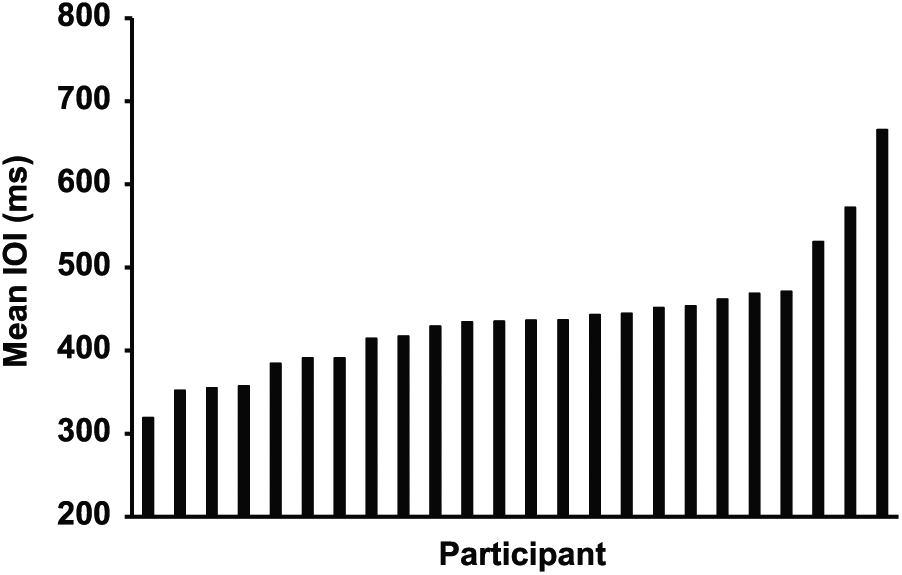
Range of pianists’ mean interonset intervals (IOIs) in the spontaneous production rate condition in the No Cue task.
Individual differences in spontaneous rates were examined in terms of experiment time of day, age, and sex, as these differences have been shown to influence biological rhythms (Duffy and Czeisler, 2002; Duffy et al., 2011; Santhi et al., 2016). Because the experimental sessions were conducted either in the morning or in the afternoon, the 24 participants were divided into two groups (early vs. late test session) based on a median split by testing time (early median = 1045 h; late median = 1330 h). A 1-way between-subjects analysis of variance (ANOVA) conducted on the spontaneous rates in the No Cue task by time of day yielded no significant effect of testing time on SPR (F1,22 = 0.46, p = 0.50), suggesting that spontaneous rates did not differ across the testing time of day. Sex differences could not be statistically compared due to unequal sample sizes of male and female pianists (4 males, 20 females). The range of spontaneous rates in the No Cue task for the male participants (390.87-665.65 msec), however, was encompassed by the range of rates observed for the female participants (318.80-572.13 msec), suggesting that sex differences in spontaneous rates did not account for the observed distribution. Finally, age differences were addressed by correlating each participant’s age with his or her mean SPR value in the No Cue task; this correlation was not significant (r22 = 0.16, p = 0.45), suggesting that age did not contribute substantially to the distribution of SPR values.
The mean rates at which pianists performed in each rate condition are shown for the No Cue and Initial Cue tasks in Figure 2. Mean IOIs increased monotonically from Fastest to Slowest conditions in both tasks, indicating that pianists followed instructions to perform at a range of rates around their SPR. An ANOVA on mean IOIs with Performance Task and Rate condition as factors confirmed a significant main effect of Rate on mean IOI (F4,92 = 101.50, p < 0.001). There was also a significant main effect of Task on mean IOI (F1,23 = 15.17, p = 0.001); pianists performed on average 18.98 msec faster in the Initial Cue task (second task) than in the No Cue task (first task). No significant interaction was observed.
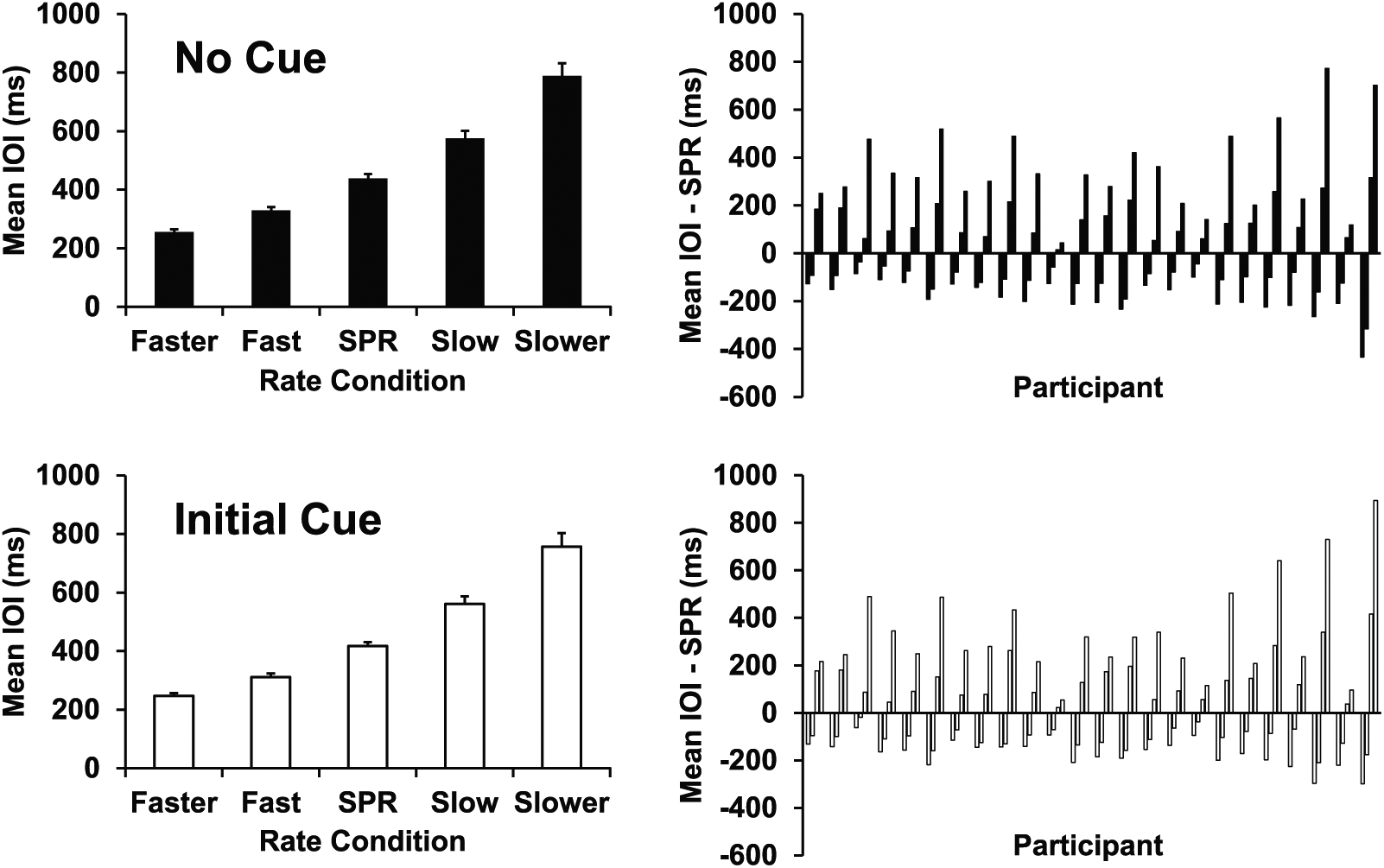
Left panel: Pianists’ mean interonset intervals (IOIs) by Rate condition for the No Cue task (top) and Initial Cue task (bottom). Right panel: Each set of 4 bars represents the difference in each pianist’s mean IOI per rate condition (faster, fast, slow, and slower) minus the mean IOI in their spontaneous production rate (SPR) condition (from left to right). Participants are ordered by mean SPR IOI in the No Cue task (fastest on the left).
Temporal Variability
The effects of spontaneous rates on temporal stability were assessed by the mean coefficient of variance measures. Pianists’ mean CVs (SD (IOI) / mean IOI) are shown in Figure 3a for all conditions. A two-way ANOVA on mean CVs revealed a main effect of Rate condition (F4,92 = 16.94, p < 0.001), with lowest CV (greatest temporal precision) in the SPR condition. There was no effect of Task (F1,23 = 0.31, p = 0.58) or interaction of Task with Rate (F4,92 = 0.336, p = 0.85). Post hoc Tukey comparisons (alpha = 0.05) were conducted to assess whether CVs significantly differed between the SPR condition and other Rate conditions, shown in Figure 3b. Results indicated that the mean CV in the SPR condition was significantly lower than mean CVs of all other Rate conditions except for the Slow condition. Figure 3b shows mean CVs for individual participants by Rate condition. Three CV outliers (values >3 SD from the mean CV across conditions and tasks) were identified in Fast/Faster rate conditions. To confirm that these outliers did not drive the observed differences between Rate conditions, the same ANOVA on CV values was repeated without the two participants who produced these three outlying values. Results confirmed the original findings of a main effect of Rate on CV (F4,84 = 17.27, p < 0.001) and no effect of Task (F1,21 = 1.78, p = 0.20). Post hoc Tukey comparisons (alpha = 0.05) confirmed the previously observed pattern of differences in means between Rate conditions.
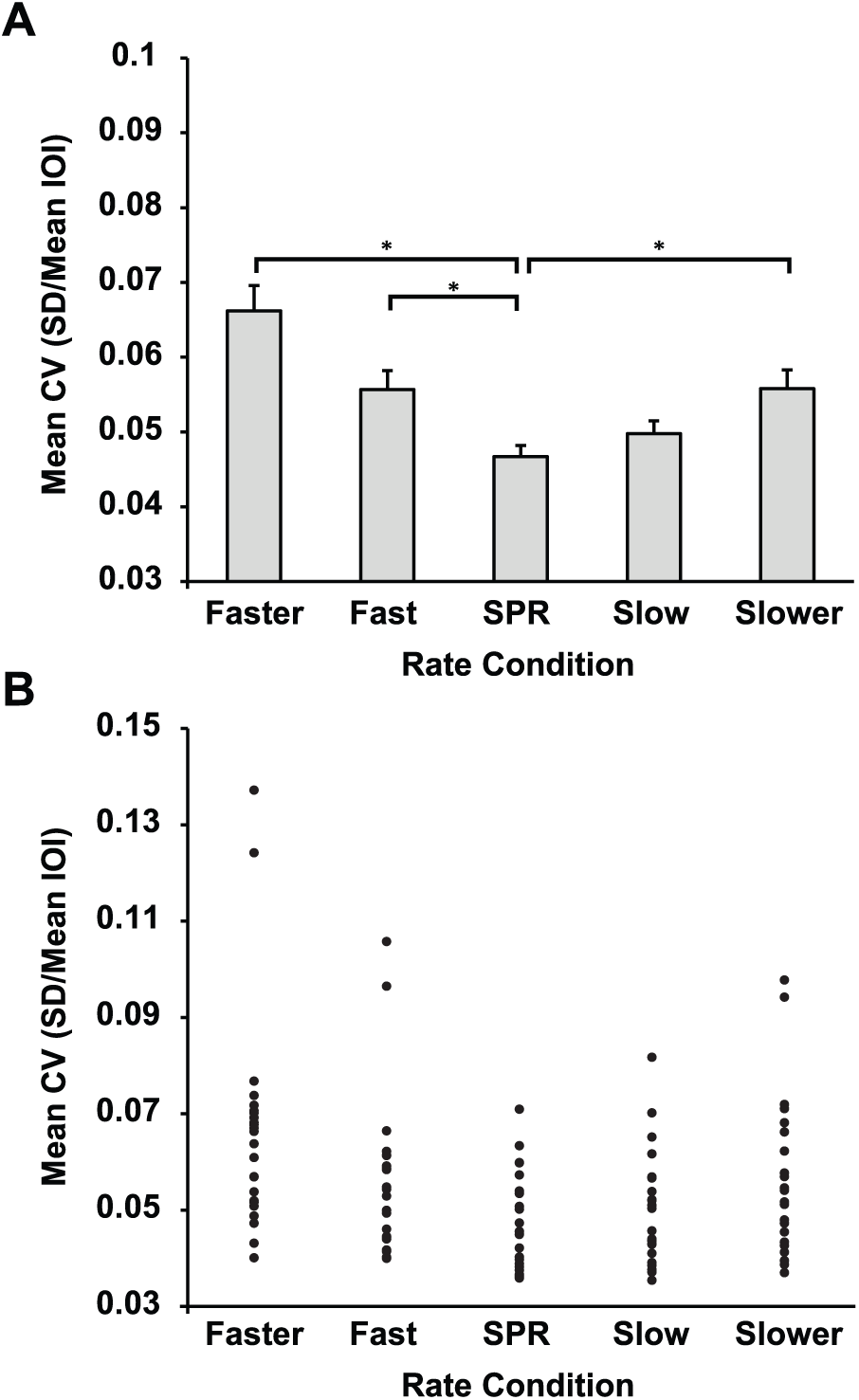
(A) Pianists’ mean coefficient of variation (CV) by Rate condition averaged across tasks. Brackets indicate significant differences between the spontaneous production rate (SPR) condition and other Rate conditions (p < 0.05). (B) Individual participant mean CVs by Rate condition.
Temporal Drift
Pianists’ tendency to drift in rate toward their spontaneous rates within each trial was assessed with the IOI adjusted slope values, shown in Figure 4a for all conditions. The slope value (IOI in milliseconds per serial position) measures change in IOI duration across successive melody tones. Participants’ mean slope value in the SPR condition (change in IOI in milliseconds across sequence events) was −0.060, suggesting that, on average, successive tone onsets decreased in duration by 0.060 msec per tone (No Cue task slope = −0.053; Initial Cue task = −0.066). A 2-way ANOVA on adjusted slope values showed a significant main effect of Rate (F3,57 = 27.75, p < 0.001); as shown in Figure 4, positive values in Fast/Faster conditions and negative values in Slow/Slower conditions indicated drift in the direction of the SPR. There was no significant effect of Task (F1,19 = 0.60, p = 0.45) or interaction of Task with Rate (F3,57 = 0.056, p = 0.983). Post hoc Tukey tests (alpha = 0.05) between all Rate condition means, shown in Figure 4a, indicated significant differences in adjusted slope values between all pairs of Rate conditions except for the Fast condition–Slow condition comparison.
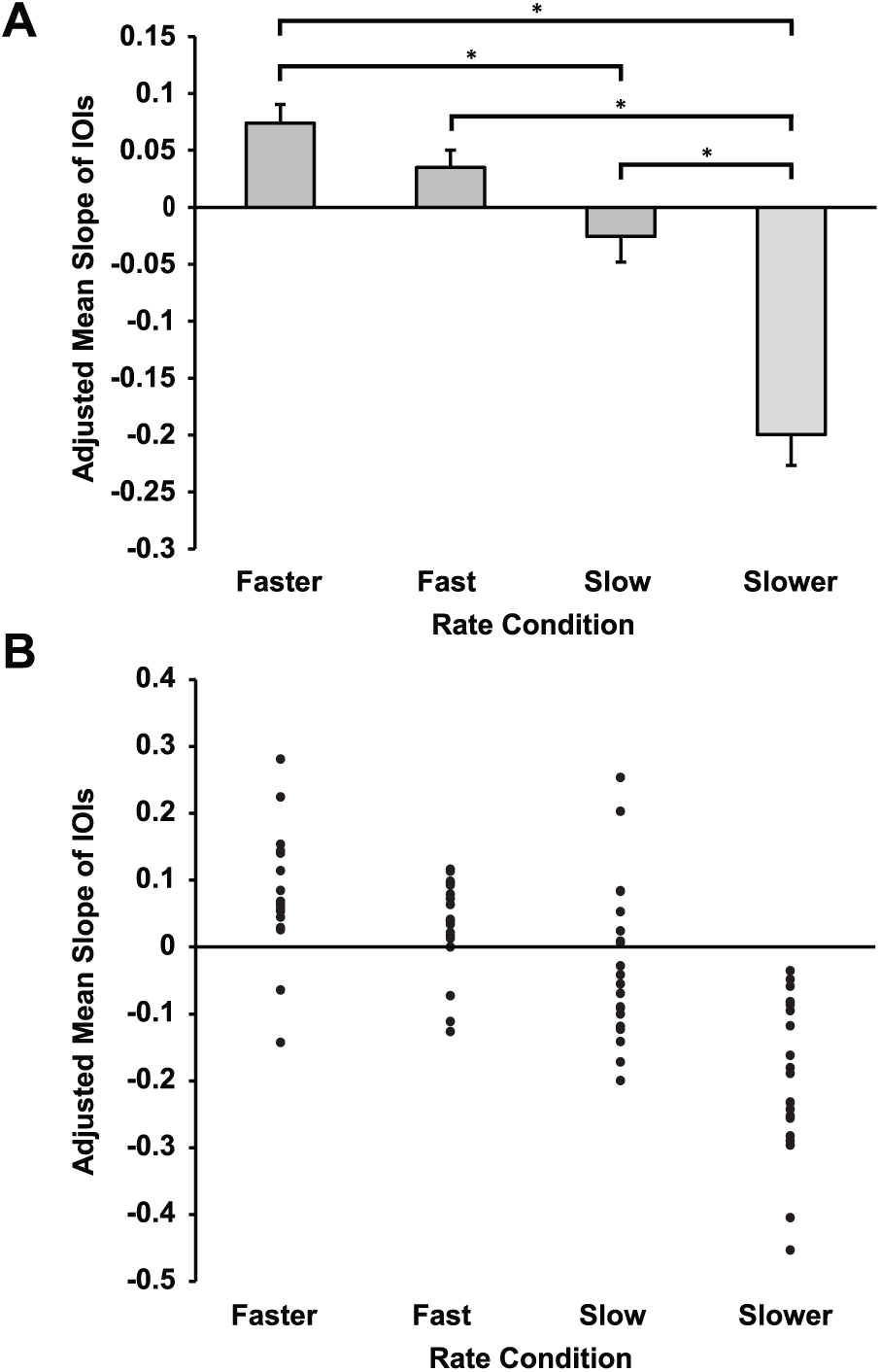
(A) Pianists’ mean adjusted slopes by Rate condition, averaged across No Cue and Initial Cue tasks. Asterisks indicate significant differences (p < 0.05). (B) Individual participant mean adjusted slopes by Rate condition.
Figure 4b shows that individual participants’ adjusted slope values were positive when participants performed at rates faster than their spontaneous rate and negative when participants performed at slower rates. To assess the consistency of this pattern, across participants, each participant’s adjusted slope values in the Faster, Fast, Slow, and Slower rate conditions were regressed on the values 1 through 4 (representing categorically increasing IOI durations across Rate conditions) within each task (No Cue, Initial Cue). Resulting values were then averaged across tasks for each participant. Nineteen of 20 participants showed a negative mean slope of adjusted slopes, a rate higher than expected by chance (binomial probability: p < 0.001). Thus, the observed pattern of adjusted slopes in Figure 4b was consistent across participants.
To test whether the amount of asynchrony across Rate conditions was related to participants’ SPR values, the slope of adjusted slope values were correlated with mean IOIs in the SPR condition for each Task. This correlation was not significant in either the No Cue or Initial Cue tasks (Initial Cue: r18 = 0.21, p = 0.38; No Cue: r18 = 0.19, p = 0.43).
Discussion
The current study tested the dynamical systems hypothesis that natural frequencies of human motion reflect optimal modes of coordination, defined by maximum temporal stability. We tested this hypothesis in the context of piano music performance, an inherently rhythmic task for which humans display large interindividual variation in their natural frequencies (Loehr and Palmer, 2011; Zamm et al., 2015, 2016). Pianists’ temporal coordination of tone onsets was maximally stable at their natural frequency of music performance, captured by SPRs, relative to performances at faster and slower frequencies. The optimal mode of coordination was also supported by lower coefficients of variation of IOIs and by temporal drift in the direction of the natural frequency.
Perturbations of a biological system from its natural frequency have been shown to result in increased behavioral variability and an eventual return to the natural frequency in both human movement tasks (Rosenblum and Turvey, 1988) and other animal forms of movement (von Holst, 1973). Dynamical system perspectives suggest that the natural frequency represents a state of minimal energy expenditure; increased motion variability occurs when the system is perturbed from its natural frequency because more energy is required to coordinate motion at other frequencies, and drift toward the natural frequency occurs because systems cannot sustain the increased energy expenditure required for coordination at other frequencies (von Holst, 1973; Bernstein, 1967; Kelso, 1997; Haken et al., 1985; Kelso, 1997). This perspective has received wide support from studies of simple rhythmic movements such as interlimb coordination (for a review, see Richardson et al., 2008). Our findings provide the first direct evidence that natural frequencies may also constrain coordination of more complex motor behaviors, such as music performance, even in the presence of large intentional rate changes.
Interestingly, pianists’ performances exhibited maximal temporal stability at their natural frequency across both No Cue (self-paced) and Initial Cue tasks, indicating that the natural frequency represents an optimal coordination mode regardless of whether the task and rate are internally or externally cued. Consistent with dynamical theories of motor coordination, principles of spontaneous (self-paced) motor coordination reflect a point of reduced energy expenditure (von Holst, 1973; Bernstein, 1967; Kelso, 1997; Haken et al., 1985). Our findings indicate that these spontaneous modes of coordination may also constrain movement in contexts in which timing is externally controlled. The self-paced task (No Cue) required greater working memory demands than the Initial Cue (externally-paced) task; to produce slower and faster self-paced performances, the pianists had to first recall their spontaneous rate. Although temporal stability measures did not differ between No Cue and Initial Cue tasks, pianists performed slightly slower on average in the (first) No Cue task than in the (second) Initial Cue task, consistent with reduced demands on working memory, as well as possible order effects. Different mechanisms contributing to self-paced and externally paced movement timing are suggested by activation of different brain networks during production of self-paced and externally paced motor sequences (Gerloff, 1998; Wessel et al., 1997). The potential influences of working memory and natural frequencies on movement pose an interesting avenue of future study.
In sum, these findings provide the first direct evidence for constraints of natural frequencies on individuals’ temporal coordination of complex rhythmic motor behavior such as music performance. Findings are also consistent with previous observations indicating that natural frequencies influence interpersonal synchrony between duet musicians, and our findings support intrinsic oscillator mechanisms (i.e., Loehr et al., 2011) that can explain why partners with similar natural frequencies perform better together than partners with different natural frequencies (Zamm et al., 2016) and why a performer with a faster natural frequency tends to anticipate the tone onsets of an external stimulus relative to an external stimulus at a slower natural frequency (Loehr and Palmer, 2011; Zamm et al., 2015). The ability to oscillate with a self-sustained amplitude in the absence of external cues and to entrain that oscillation to external rhythms is not limited to music performance; it is also a defining feature of circadian clocks (Roenneberg et al., 2003b). An open question is whether these natural frequencies in musicians’ performance correspond to other endogenous rhythms such as sleep-wake cycles, body temperature, and metabolic rate (Rising et al, 1992: Roenneberg et al., 2003a; Santhi et al., 2016). The current research provides a bridge for investigating these questions about the origins of endogenous rhythms in human action sequences.
Footnotes
Acknowledgements
The authors thank Frances Spidle and Jamie Dunkle for technical assistance. The research was supported by an FQRNT-PBEEE award to A. Zamm and by a Canada Research Chair and NSERC Grant 298173 to C. Palmer.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.