
Abstract
The fruit fly Drosophila melanogaster survives thermally stressful conditions in a state of reproductive dormancy (diapause), manifested by reduced metabolic activity and arrested ovarian development in females. Unlike insects that rely primarily on photoperiodic stimuli to initiate the diapause program, in this species dormancy is regulated by low temperature and enhanced by shorter photoperiods. Overwintering phenotypes are usually studied under simple laboratory conditions, where animals are exposed to rectangular light-dark (LD) cycles at a constant temperature. We sought to adopt more realistic diapause protocols by generating LD profiles that better mimic outdoor conditions. Experimental flies were subjected to semi-natural late autumn and summer days, while control females received the same amounts of light but in rectangular LD cycles (LD 8:16 and LD 15:9, respectively). We observed that semi-natural autumnal days induced a higher proportion of females to enter dormancy, while females in semi-natural summer days showed reduced diapause compared with their corresponding rectangular controls, generating an impressive photoperiodic response. In contrast, under rectangular light regimes, the diapause of Drosophila field lines exhibited minimal photoperiodicity. Our semi-natural method reveals that D. melanogaster diapause is considerably more photoperiodic than previously believed and suggests that this seasonal response is best studied under simulated natural lighting conditions.
Organisms are constantly exposed to predictable cyclical changes of environmental stimuli that reoccur on a daily and seasonal basis. Consequently, organisms have evolved adaptive mechanisms that allow them to predict these changes and enhance their fitness (Kreitzman and Foster, 2009). With the approaching winter, the fruit fly Drosophila melanogaster enters a dormant state, called diapause, in order to overwinter (Saunders et al., 1989; Tatar et al., 2001; Williams and Sokolowski, 1993). Diapause is a neurohormonally regulated switch to an alternative developmental program, characterized by marked changes in metabolism, growth, survival, lifespan, and stress resistance (Denlinger, 2002; Hahn and Denlinger, 2011; Kubrak et al., 2014; Schiesari et al., 2016). In many insects, dormancy is a robust photoperiodic response, initiated by short day lengths (De Wilde, 1962; Hodek, 1971; Numata and Hidaka, 1982; Enomoto, 1982; Watabe, 1983; Kreitzman and Foster, 2009). However, D. melanogaster exhibits a shallow diapause, regulated predominantly by temperature that can be enhanced by shorter photoperiods (Saunders et al., 1989; Saunders, 1990; Emerson et al., 2009; Tauber et al., 2007).
Despite the complexity of the external environment, most laboratory studies rely on basic simulations of outdoor lighting conditions, using rectangular light-dark (LD) cycles based on light on-off switching at constant temperatures. However, when studied in the wild (compared with the laboratory), mammals and flies reveal quite different circadian phenotypes at both molecular and behavioral levels (Gattermann et al., 2008; Daan et al., 2011; Vanin et al., 2012; Bywalez et al., 2012; Menegazzi et al., 2012; Menegazzi et al., 2013; Montelli et al., 2015).
Possible connections between the circadian clock and the diapause-governing seasonal timing system have been suggested in numerous studies on various insects (Pavelka et al., 2003; Goto et al., 2006; Tauber et al., 2007; Ikeno et al., 2011a, 2011b; Meuti et al., 2015). In D. melanogaster, two allelic variants of the circadian clock gene timeless (tim) were found to significantly affect the overwintering response (Tauber et al., 2007). Individuals carrying the ls-tim allele that generates both long (l) and short (s) TIM protein isoforms exhibit consistently higher diapause levels compared with females bearing the s-tim variant (Tauber et al., 2007; Kyriacou et al., 2008, Schiesari et al., 2016; Pegoraro et al., 2017). It was furthermore shown that LS-TIM attenuates the photosensitivity of the circadian clock due to its weaker interaction with the circadian photoreceptor CRYPTOCHROME (CRY) (Sandrelli et al., 2007). In northern latitudes, where shorter daylengths are associated with relatively lower temperatures compared with southern latitudes, possessing an attenuated circadian photosensitivity can be advantageous, allowing flies to enter diapause more readily (Sandrelli et al., 2007). Since the fly’s circadian clock appears to be highly influenced by natural conditions (Vanin et al., 2012), diapause may similarly be altered under biologically more realistic circumstances.
An improved diapause protocol has recently been introduced in which both the temperature and rectangular LD cycles were gradually changed in order to mimic the shortening days of an upcoming winter and the lengthening days of an approaching spring (Zonato et al., 2017a). This winter-spring scenario was found to enhance the diapause response of flies and brought about more stable phenotypes (Zonato et al., 2017a). Although this protocol recognized the importance of changing the conventional simplified photoperiod and temperature settings, it still operates entirely with rectangular profiles for both light and temperature parameters.
We have therefore further improved our experimental protocol by using simulated semi-natural light profiles to better mimic autumn and summer days while also considering light patterns at dawn and dusk. Here we report features of diapause in 4 European Drosophila field lines (in both ls- and s-tim genetic backgrounds), tested under either rectangular or more realistic semi-natural light conditions.
Materials and Methods
Fly Strains
The D. melanogaster strains Hu-S and Hu-LS originated from an isofemale line obtained from a natural population of Houten (Holland, 52.02° N; 5.16° E) (Fig. 1A), collected in 2004. PCR genotyping was used to make these lines homozygous for either the s- or ls-tim variant (Tauber et al., 2007; Sandrelli et al., 2007). WTALA-S and WTALA-LS are Northern Italian isofemale lines, collected in 2004. WTALA-S (bearing s-tim) was originated from Postal (46.61° N; 11.19° E), while WTALA-LS (carrying ls-tim) was derived from Foresta (46.67° N; 11.12° E) (Fig. 1A). Both lines, together with several others sampled in the same area, were used to establish the natural wild-type strain WT-ALA (Wild Type Alto Adige) (Vanin et al., 2012).
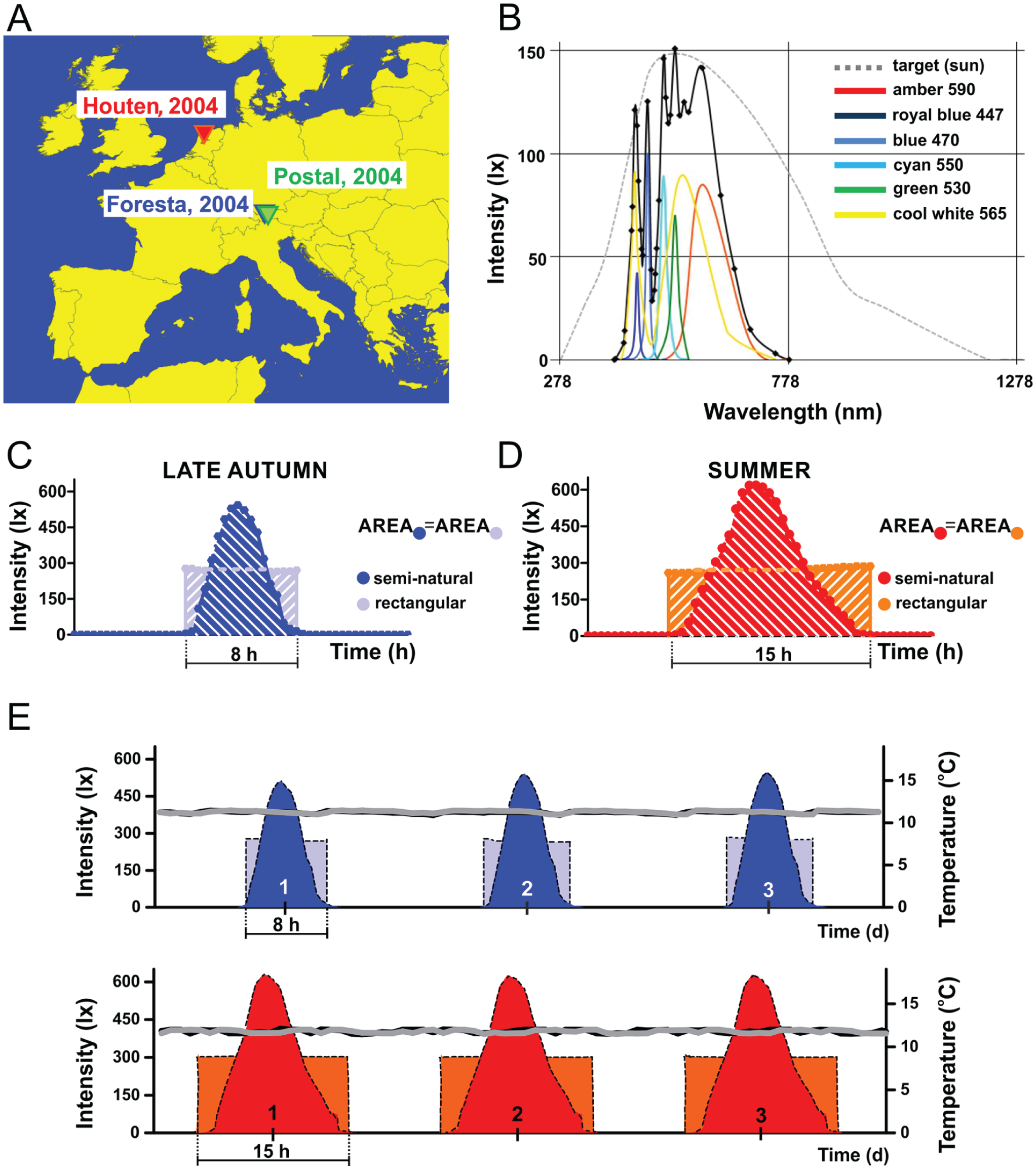
Simulated profiles to mimic light conditions of consecutive fall and summer days. (A) D. melanogaster field lines used in this study: Houten (Holland), Postal (Italy, designated as WTALA-S), and Foresta (Italy, designated as WTALA-LS). (B) Color composition of the generated light profiles. Various selected LED sources are used to approximate as much as possible the target profile of the sun (gray dotted line). The different color components and their wavelength are listed on the right. The sum of the contribution of all the individual colors is marked by a black continuous line. (C, D) A representative late autumnal and summer day and the corresponding rectangular short (LD 8:16) and long (LD 15:9) day. The integrals of the semi-natural and rectangular profiles are equal in the case of both fall and summer conditions. (E) Three representative days of the simulated late autumnal (upper panel) and summer profiles (lower panel). Gray and black curves indicate the recorded temperature data during rectangular and semi-natural light settings, respectively.
Flies were reared on a standard cornmeal/agar medium, at 23 °C, 70% relative humidity, in 12-h light/12-h dark cycles (LD 12:12; zeitgeber time (ZT): lights-on ZT0 at 0900 h, lights-off ZT12 at 2100 h; 480 lux) until their eclosion.
Simulation of Fall and Summer Profiles
To investigate the effect of photoperiod on diapause induction, we simulated late autumn and summer days in the laboratory based on a real dataset collected in Northern Italy (Treviso, 45.67° N; 12.23° E) (Vanin et al., 2012). The experimental setup takes into consideration the intensity changes of natural sunlight throughout the day and better mimics outdoor light conditions in term of spectral composition. To generate natural-like light profiles according to outdoor conditions, we constructed a custom-built programmable simulator that produces the light regimes in an incubator (Sanyo Electric Co., Osaka, Japan, MLR-351 versatile environmental test chamber). The adopted device combines outputs of 6 groups of light-emitting diodes (LEDs) with different emission spectra (Fig. 1B) and dictates sophisticated light profiles with progressively changing light intensity over the day (described also in Vanin et al., 2012; Green et al., 2015a, 2015b). Twilight data were accessed from the online database of the United States Naval Observatory (USNO) Astronomy Application Department, “Rise, Set, and Twilight Definitions” (http://aa.usno.navy.mil/faq/docs/RST_defs.php). To mimic upcoming winter days, light conditions of a late November day were simulated, adopting a day length of ~9 h (8 h 59 min, 30 November 2014). Because fruit flies have been shown to favor shaded conditions in nature (Rieger et al., 2007), 510 lux maximum light intensity was chosen, a value close to what has been recorded by the Trikinetics (Waltham, MA, USA) environmental sensor when placed in shadow in the wild (Vanin et al., 2012). To ensure that individuals under the rectangular LD cycles (used as controls) received equal amounts of light, the integral of the continuous light curve was calculated, yielding 290 lux constant light intensity to be used during the 8 h of light phase of rectangular cycles (Fig. 1, C and E). In the case of summer days, a photoperiod of 15 h was adopted for the rectangular profile, maintaining 290 lux light intensity. Considering that on the longest summer day in Treviso the photoperiod is ~15.5 h (15 h, 42 min, 21 June 2015) and that areas determined by the light profiles must be equal between rectangular and semi-natural conditions, we used 620 lux as maximum light intensity to mimic a real summer’s day. All experiments were performed at constant 12 °C, and both temperature and light data were continuously monitored (Fig. 1, D and E).
Diapause Assays
Newly eclosed flies (5 h post eclosion) were collected in plastic vials and placed under diapause-inducing conditions. After 11 days, flies were killed in absolute ethanol and immediately analyzed in PBS. The ovaries of females were dissected under 40× zoom with LeicaMZ6 stereomicroscope (Leica Microsystems GmbH, Wetzlar, Germany), and diapause levels were scored. Each female was considered being in diapause in the complete absence of vitellogenic oocytes in its gonads, when yolk deposition is not yet visible (Saunders et al., 1989). Diapause levels were later presented as the proportion of diapausing females among all the dissected individuals. At least 60 females per replicate were analyzed, and a minimum of 11 replicates per condition were performed.
Statistical Analyses
All data were plotted with GraphPad Prism 6 software, whereas the statistics were computed with R statistical software (version 3.0.1, www.r-project.org). Diapause data were transformed to arcsine value prior to be statistically analyzed. Normal distribution was controlled by Shapiro-Wilk normality test. Differences between groups were determined by ANOVA, while pairwise post hoc comparisons were conducted with Tukey honest significant difference (HSD) tests.
Results
Flies were subjected to short autumnal and long summer days through use of semi-natural and rectangular light profiles (LD 8:16 and LD 15:9, respectively). We first observed that the adopted light condition (semi-natural or rectangular) has a strong season-dependent effect on the percentage of diapausing females (ANOVA, significant light profile × season interaction; F1,252 = 219.6, p < 0.001). To further investigate the effect of the newly generated profiles, we compared results of semi-natural and rectangular conditions for both seasons. When individuals were exposed to semi-natural light under short, autumnal days to mimic the forthcoming winter, a higher proportion of females entered the dormant state compared with their corresponding control flies kept in short rectangular cycles (Tukey HSD test, p < 0.001) (Fig. 2A). However, when flies were subjected to semi-natural long summer days, the effect of the light profile used was the opposite: Females showed reduced diapause levels compared with those subjected to long rectangular LD cycles (Tukey HSD test, p < 0.001) (Fig. 2B). Consequently, semi-natural conditions enhance the difference in diapause levels between the two photoperiods.
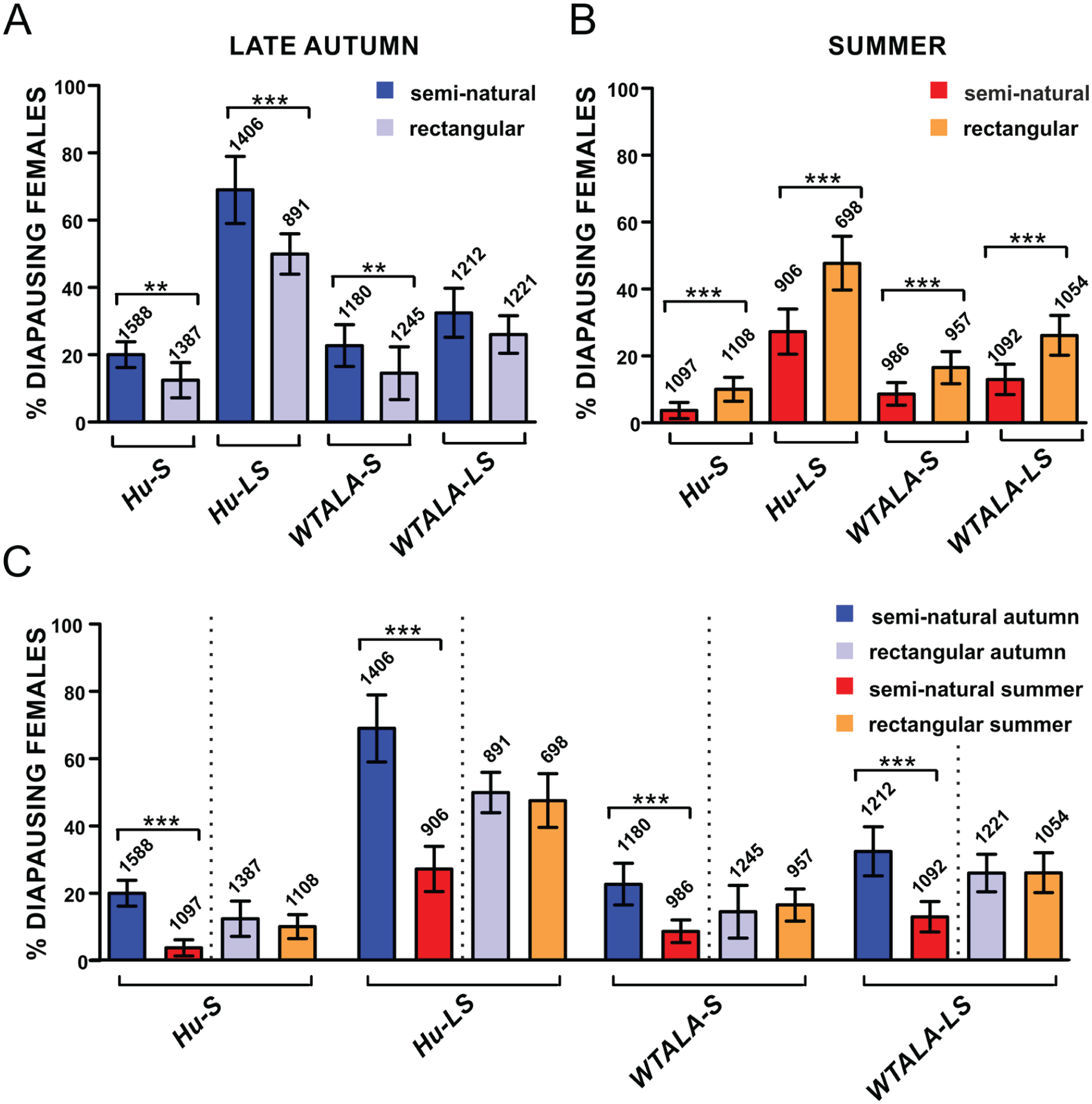
Semi-natural light profiles largely affect the incidence of diapause in Drosophila field lines. (A) Simulation of consecutive late autumnal days induced a higher proportion of females to enter diapause (dark blue bars) compared with controls kept in short rectangular LD cycles (LD 8:16; light blue bars). (B) Generation of summer light conditions promoted diapause response in a significantly smaller proportion of females (red bars) compared with controls exposed to long rectangular LD cycles (LD 15:9; orange bars). (C) Data are replotted from panels A and B. Robust photoperiodic diapause emerged when more realistic light profiles were used, highlighting significantly higher levels of dormancy during late autumnal days (red vs. blue bars). When flies were subjected to rectangular LD cycles, diapause levels did not differ when long and short days were compared (orange vs. light blue bars). Data are presented as mean ± SD. Numbers above each column indicate the number of dissected females. ANOVA after arcsine transformation, followed by post hoc Tukey HSD test. ***p < 0.001.
Natural populations of European D. melanogaster have been reported to be photoperiodic when tested under rectangular LD cycles (Tauber et al., 2007; Zonato et al., 2017a), while American wild-caught lines were found to enter diapause regardless of the photoperiod (Emerson et al., 2009). To analyze the photoperiodic response of flies in our experimental setup, diapause inducibility between long and short days was compared in both semi-natural and rectangular conditions. Surprisingly, approximately the same proportion of females entered the dormant state in rectangular short and long days, suggesting that the natural populations used in this experiment do not exhibit a photoperiodic diapause when exposed to these simplified light conditions (Tukey HSD test, p = 0.82, not significant) (Fig. 2C). However, a robust photoperiodic diapause response was observed when individuals were exposed to semi-natural light profiles, with a significantly higher diapause incidence during simulated autumnal days (Tukey HSD test, p < 0.001) (Fig. 2D).
We additionally found, as expected, a significant timeless influence on the incidence of dormancy (ANOVA, F1,252 = 878.7, p < 0.001), with higher levels of diapause in the case of Hu-LS and WTALA-LS females, both carrying the ls-tim allele. This result corroborates the data obtained by Tauber et al. (2007), who first described the diapause-promoting action of the recently arisen natural ls-tim allele. The effect of tim was irrespective of whether rectangular or semi-natural conditions were used, as ANOVA showed no interaction for tim polymorphism × light profile (F1,252 = 0.7, p = 0.42, not significant).
Discussion
Circadian and seasonal responses are crucial adaptive mechanisms enabling organisms to synchronize their behavior and physiology with their external environment. In their study of Drosophila circadian behavioral rhythms in nature, Vanin et al. (2012) first pointed out some unexpected features, emphasizing the importance of studying organisms under more natural environmental stimuli. Here we provide evidence that seasonal diapause levels are highly influenced by more realistic environmental conditions. Our results highlight that, unlike insects that exhibit photoperiodic dormancy even under commonly used rectangular LD laboratory cycles (De Wilde, 1962; Hodek, 1971; Numata and Hidaka, 1982; Enomoto, 1982; Watabe, 1983; Tauber et al., 1986; Kreitzman and Foster, 2009), the two populations of European fruit flies that we have studied seem not to be as photoperiodic under these simplified light conditions. However, when outdoor conditions were better mimicked by semi-natural light profiles, a robust photoperiodic diapause response appeared in the two European Drosophila lines tested.
Diapause does not occur in summer in D. melanogaster but rather in late autumn, when daily average light levels are generally reduced compared with their peak in summer. Furthermore, when flies diapause they do so under manure heaps, in garden sheds, at the base of fruit trees just below the grass cover, or in holes within the tree trunks (C.P. Kyriacou, unpublished observations). Light levels are low under these conditions, so we used light intensities that were less than those reported by Vanin et al (2012) in their locomotor activity studies under natural conditions and more realistic for diapausing environments.
Despite the very simplified rectangular lights-on/lights-off cycles, photoperiod-driven diapause remains robust in some Drosophilids, as in D. littoralis, D. montana, and D. ezoana (Lankinen, 1986; Salminen et al., 2015). However, in D. melanogaster diapause appears to be a shallow response that is predominantly regulated by temperature (Saunders et al., 1989; Saunders, 1990; Emerson et al., 2009; Anduaga et al., 2018), and shares elements of both diapause and quiescence (Tatar et al., 2001). The exit from this dormancy occurs soon after flies are transferred to warm temperatures, regardless of the photoperiod (Saunders et al., 1989; Tatar et al., 2001). Nevertheless, Saunders et al. (1989) also reported a photoperiodic diapause response in the wild-type Canton-S strain of D. melanogaster. Females exposed to short days at 12 °C entered reproductive diapause, while those maintained in long days underwent ovarian maturation at the same low temperature (Saunders et al., 1989). However, it is important to stress that the Canton-S strain has been domesticated in laboratory conditions for about 100 years (Bridges, 1916), and the lack of key external stimuli in the environment of captive animals may modify behavioral phenotypes (Price, 1999; Stanley and Kulathinal, 2016). Indeed, marked behavioral and genetic differences have been documented between the typical laboratory-reared Drosophila strains (Canton-S, Oregon-R, w1118) and wild populations (Stanley and Kulathinal, 2016). Therefore, photoperiodic diapause should also be studied in fly populations, which have been introduced to the laboratory from the wild more recently. For this reason, all the lines used in this study were wild-caught in 2004, but even so, this amount of time in the laboratory may induce some “domestication.” It might therefore be interesting and informative to examine whether freshly collected lines show similar photoperiodic effects.
Natural populations of European D. melanogaster have been reported to exhibit photoperiodic diapause both at 12 °C and 13 °C when kept in rectangular LD cycles (Tauber et al., 2007; Zonato et al., 2017a). However, in our hands, all the 4 Drosophila populations failed to distinguish between long and short days when exposed to similar LD cycles. The likely explanation for the contrasting results might be the thermal difference between the two photoperiodic conditions in the different studies. Biological experiments are commonly performed in incubators that generate the desired light regime relying on their own built-in lighting system containing fluorescent light tubes. However, one needs to consider that the majority of the electrical energy input (~60%-90%) dissipates as heat production; thus, long photoperiods very often couple to slightly higher temperatures, creating difficulties in studying clear photoperiodic effects. Indeed, recent experiments on Drosophila chill coma recovery times (CCRt) revealed that the difference between the CCRt of females reared under short and long photoperiods can be explained, at least partially, by a low-amplitude thermoperiod generated by the light system (Pegoraro et al., 2014). Light boxes are also frequently used to keep the samples under the chosen photoperiod, although they usually operate with more energy-efficient LEDs. However, heat generation occurs also within the LED, and in the relatively small, closed light box these temperature fluctuations could be problematic.
One way of overcoming such small temperature changes is to place flies of the same genotype in the same light box, but covered in foil, so the flies are exposed to any photoperiodically induced temperature fluctuations, but in DD (Tauber et al., 2007, Pegoraro et al., 2014, Zonato et al., 2016). Under these conditions, these covered flies show no photoperiodic diapause so the temperature cycle on its own does not create one. However, even this may be an insufficient control because the experimental flies may also experience a “greenhouse” effect as they are maintained in glass vials, so they may be exposed to slightly higher temperatures during the photoperiod than the control flies that are covered. Indeed, this has recently been demonstrated in a very carefully controlled study of thermal effects on diapause under such LD conditions (Anduaga et al., 2018). Therefore, when one is studying the specific effect of different photoperiods on diapause, it is important to ensure that the observed phenotype is due to the photoperiod and is not influenced by any temperature cycle. However, realistically there is no light without heat, so in nature one comes with the other. Nevertheless, our natural light simulations have therefore “rediscovered” a true photoperiodic effect on diapause. Perhaps some further subtle changes in rate of change of illumination during the day, for example, which is likely to differ among latitudes, might even enhance this response and would certainly be worth attempting in future. In the experimental setup, no light-related temperature oscillations were detected, as any temperature fluctuations were similarly present during the dark phases. Emerson et al. (2009) examined dormancy in American natural populations of D. melanogaster, maintained at 4 different low temperatures (10, 11, 12, and 14 °C) in both short and long days (LD 10:14 and LD 18:6, respectively). The investigators found that flies entered the dormant state irrespective of the photoperiod (Emerson et al., 2009). Importantly, thermal conditions of the different photoperiods were also registered, highlighting no marked temperature differences between long and short days in their experimental setup (Emerson et al., 2009). The authors concluded that low temperature is the leading environmental cue that governs the reproductive dormant state in this species, a conclusion mirrored by Anduaga et al., 2018. Our results are in accordance with these data, since all the 4 European lines tested entered diapause regardless of the length of the rectangular photoperiod.
We additionally found significant differences between the diapause inducibility of flies carrying the ls-tim allele (Hu-LS and WTALA-LS) and those expressing the s-tim variant (Hu-S and WTALA-S): ls-tim flies exhibited consistently higher levels of dormancy under all light profiles tested. These results corroborate the data of Tauber et al. (2007), who first documented the diapause-enhancing effect of the recently evolved ls-tim allele. European fly populations originating from more northern areas compared with southern flies have never been observed to show the kind of steep latitudinal cline in diapause inducibility present in populations from North America, and any cline seems quite shallow in the Old World (Schmidt et al., 2005; Tauber et al., 2007; Pegoraro et al., 2017; Zonato et al., 2017a). While the more northern ls-tim line (Hu-LS) exhibits higher diapause levels compared with the southern ls-tim population (WTALA-LS), this was not observed when comparing Hu-S with WTALA-LS that carry the ancient s-tim allele. Nor was this the case in two southern Italian population studied by Tauber et al. (2007) when compared with the northern Houten line, again suggesting the absence of any strong cline in diapause in Europe (see Pegoraro et al., 2017). This is to be expected as the highly diapausing ls-tim allele originated very recently in southern Italy (Zonato et al., 2017b) and is spreading northward under natural selection (Tauber et al., 2007), thereby obscuring any diapause cline.
In conclusion, our results demonstrate that none of the tested European D. melanogaster populations show photoperiodic diapause under simplified laboratory conditions. Indeed, reports of photoperiodic effects reported by us and others may have been due to a “greenhouse effect” whereby small increases in temperature (<0.4 °C) associated with the lights-on phase could have cumulatively enhanced development over the 12-day diapause maintenance period (Anduaga et al., 2018). However, robust photoperiodicity appears when more realistic conditions are adopted. Flies perceive the difference between simplistic and more natural-like light simulations and use this information to regulate their diapause behavior. In our experimental setup, semi-natural autumn days enhanced while the summer conditions reduced the diapause response of females compared with flies subjected to rectangular LD cycles. Moreover, our results corroborate the observation that ls-tim populations display higher diapause levels than s-tim flies. Our work, both methodologically and biologically, contributes to making D. melanogaster a better model for diapause studies than previously believed, which now includes a robust photoperiodic component. This is important given the sophisticated genetic toolbox that is available for this species, which should allow the molecular dissection of this more natural seasonal response.
Footnotes
Acknowledgements
This project was supported by the INsecTIME Marie Curie Initial Training Network, grant PITN-GA-2012-316790 to R.C. and C.P.K. and ITN doctoral fellowships to D.N. and A.M.A. G.A. was funded by a postdoctoral fellowship from the University of Padova (Italy).
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.