
Abstract
Within the human population, there is large interindividual variability in the timing of sleep and circadian rhythms. This variability has been attributed to individual differences in sleep physiology, circadian physiology, and/or light exposure. Recent experimental evidence suggests that the latter is necessary to evoke large interindividual differences in sleep and circadian timing. We used a validated model of human sleep and circadian physiology to test the hypothesis that intrinsic differences in sleep and circadian timing are amplified by self-selected use of artificial light sources. We tested the model under 2 conditions motivated by an experimental study (Wright et al., 2013): (1) a “natural” light cycle, and (2) a “realistic” light cycle that included attenuation of light due to living indoors when natural light levels are high and use of electric light when natural light levels are low. Within these conditions, we determined the relationship between intrinsic circadian period (within the range of 23.7-24.6 h) and timing of sleep onset, sleep offset, and circadian rhythms. In addition, we simulated a work week, with fixed wake time for 5 days and free sleep times on weekends. Under both conditions, a longer intrinsic period resulted in later sleep and circadian timing. Compared to the natural condition, the realistic condition evoked more than double the variation in sleep timing across the physiological range of intrinsic circadian periods. Model predictions closely matched data from the experimental study. We found that if the intrinsic circadian period was long (>24.2 h) under the realistic condition, there was significant mismatch in sleep timing between weekdays and weekends, which is known as social jetlag. These findings indicate that individual tendencies to have very delayed schedules can be greatly amplified by self-selected modifications to the natural light/dark cycle. This has important implications for therapeutic treatment of advanced or delayed sleep phase disorders.
Keywords
Sleep timing in the real world is highly variable across individuals, as has been documented using large-scale surveys (Roenneberg et al., 2007; Walch et al., 2016). The term “chronotype” is used to describe an individual’s preferred sleep timing, typically measured on nonwork (free) days. Sleep timing is under the control of both a sleep homeostatic process and a circadian pacemaker (Daan et al., 1984). The phase of entrainment of an individual’s circadian pacemaker depends both on physiological factors (e.g., intrinsic circadian period) and the pattern of light to which the individual is exposed (Duffy et al., 2001; Gronfier et al., 2007). These relationships are understood in terms of the circadian clock’s phase response curve and entrainment (Winfree, 1980). Currently, however, we lack a theoretical understanding of the relationships between an individual’s physiology, the light patterns he or she selects in the real world, and how this interaction affects sleep timing. This understanding would be helpful in developing effective personalized interventions for individuals who desire to shift their sleep timing, or for evaluating potential policy changes designed to modify sleep patterns at the societal level.
Sleep timing plays a crucial role in human health and physiology. Individuals with a midsleep time later than 5:30 AM have higher caloric consumption and a higher body mass index (Baron et al., 2011). Habitually sleeping after 1 AM is associated with smaller hippocampal volumes in healthy university students (Kuperczkó et al., 2015). Individuals who sleep late are also more likely to experience social jetlag, which is the mismatch in sleep timing between work days and free days (Wittmann et al., 2006). Social jetlag involves misalignment between sleep/wake and circadian cycles and is associated with poor outcomes, including obesity (Roenneberg et al., 2012), worse academic performance (Asarnow et al., 2014), increased resting heart rate (Rutters et al., 2014), and emotional distress (Asarnow et al., 2014).
Several physiological factors have been identified that modify an individual’s preference toward early or late sleep timing. The best-studied example is the intrinsic period of the circadian pacemaker,
The importance of environment in determining an individual’s chronotype was demonstrated by a 2-week experiment in which individuals were studied both when they were living at home with access to electric lights and when they went camping without access to electric light for a week (Wright et al., 2013). When camping, individual sleep times shifted much earlier and differences in sleep timing between the individuals were greatly reduced compared to when they had access to electric light. These findings suggest that an individual’s chronotype could be considered an evoked trait, and that the wide range of interindividual differences found in sleep timing across the population may be primarily a function of self-selected light patterns. Indeed, this has been previously proposed (Roenneberg and Merrow, 2007), and the concept is intuitive, since individuals who have a physiological tendency toward later schedules are more likely to be exposed to light later in the day, further delaying their sleep timing due to circadian phase delay. Similarly, individuals who have a physiological tendency toward early schedules are more likely to be exposed to light earlier in the day, further advancing their sleep timing. However, this idea is yet to be explored quantitatively.
Mathematical models have been developed to explain human sleep timing in terms of circadian and homeostatic processes. These models include the 2-process model (Daan et al., 1984) and more recent models of sleep physiology (Rempe et al., 2010; Kumar et al., 2012; Fulcher et al., 2014), some of which include the effects of light (Phillips et al., 2010, 2011; Gleit et al., 2013), which is the strongest zeitgeber for the human circadian pacemaker (Mistlberger and Skene, 2005; Duffy and Czeisler, 2009). Recorded patterns of self-selected light in time-free laboratory conditions have been used as inputs to a model of the circadian pacemaker to explain the circadian period observed under those conditions (Klerman et al., 1996). To date, however, models have not been applied to understanding the patterns of light exposure that individuals self-select in the real world and how these patterns interact with an individual’s physiology to potentially evoke their particular chronotype. Roenneberg and Merrow (1999) coined the term “zeitnehmer” for a circadian system that self-selects the timing of its exposure to a zeitgeber, which they implemented in a simple model of photosynthesis. The zeitnehmer concept can potentially explain the effects of social zeitgebers on the circadian clock and sleep timing, because social behaviors (e.g., when we choose to sleep) shape patterns of exposure to light (Roenneberg et al., 2013). Passive avoidance of light at certain times (e.g., during sleep) has been implemented in models of the human circadian clock and sleep (Phillips et al., 2010, 2011), but not as an active process in which the model can choose to switch lights on, as humans routinely do in the real world.
Here, we implemented simple decision-making rules in a previously validated model of human sleep and circadian rhythms to understand how self-selection of light exposure influences sleep timing and social jetlag. Using this approach, we investigated the effects of varying 1 physiological parameter (the intrinsic circadian period) to test our hypothesis that self-selection of light exposure will tend to evoke a greater range of chronotypes across the population, leading to amplification of individual differences relative to a fixed light/dark cycle. Specifically, the results of our simulations are compared to the results from the Wright et al. (2013) camping study. It is natural to begin by investigating the effects of changes in 1 physiological parameter; we chose the intrinsic circadian period, due to the large existing literature on circadian period in humans and other species.
Materials and Methods
We use a mathematical model of human sleep and circadian physiology to simulate the effects of realistic self-selected light patterns on sleep and circadian timing. The model is based on physiology and has been used previously in the same or similar form to investigate the effects of different physiological parameters on sleep and circadian timing, given either a 24-h periodic light stimulus (Phillips et al., 2010) or constant light during all hours of wakefulness (Phillips et al., 2011). Below, we describe the model equations, the rules used for generating sleep and light schedules, the model parameter values (Table 1), and the methods used for simulations and analyses. The mathematical model used in this article is freely available to others via e-mail or GitHub.
Parameter values used in the model.
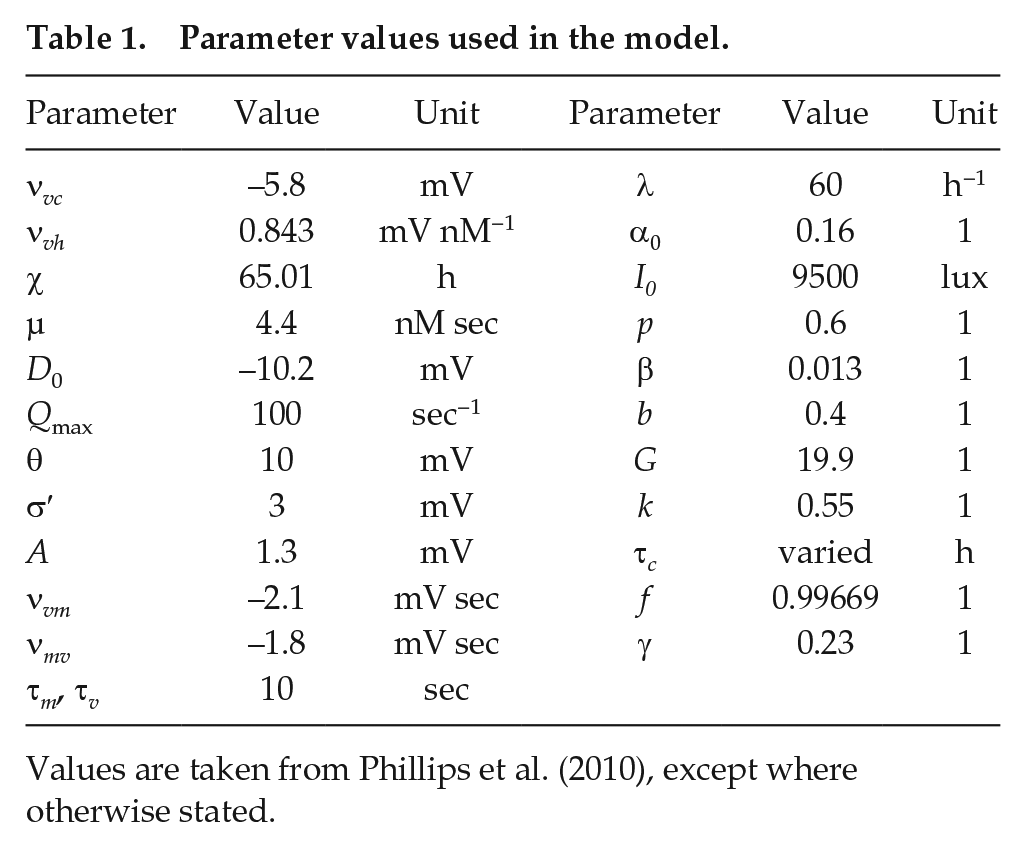
Values are taken from Phillips et al. (2010), except where otherwise stated.
Model of Human Sleep and Circadian Physiology
The model includes the following components: circadian phototransduction, the central circadian pacemaker, the sleep homeostatic process, and the sleep/wake switch (Figure 1). Each of these model components has previously been fit and tested against human data, both individually and in combination (Forger et al., 1999; Jewett et al., 1999; Kronauer et al., 1999; Phillips and Robinson, 2007, 2008; Phillips et al., 2010). As inputs, the model takes light levels (in lux) and times at which sleep is allowed. As outputs, the model gives the circadian phase, homeostatic sleep pressure, and sleep/wake state as functions of time.
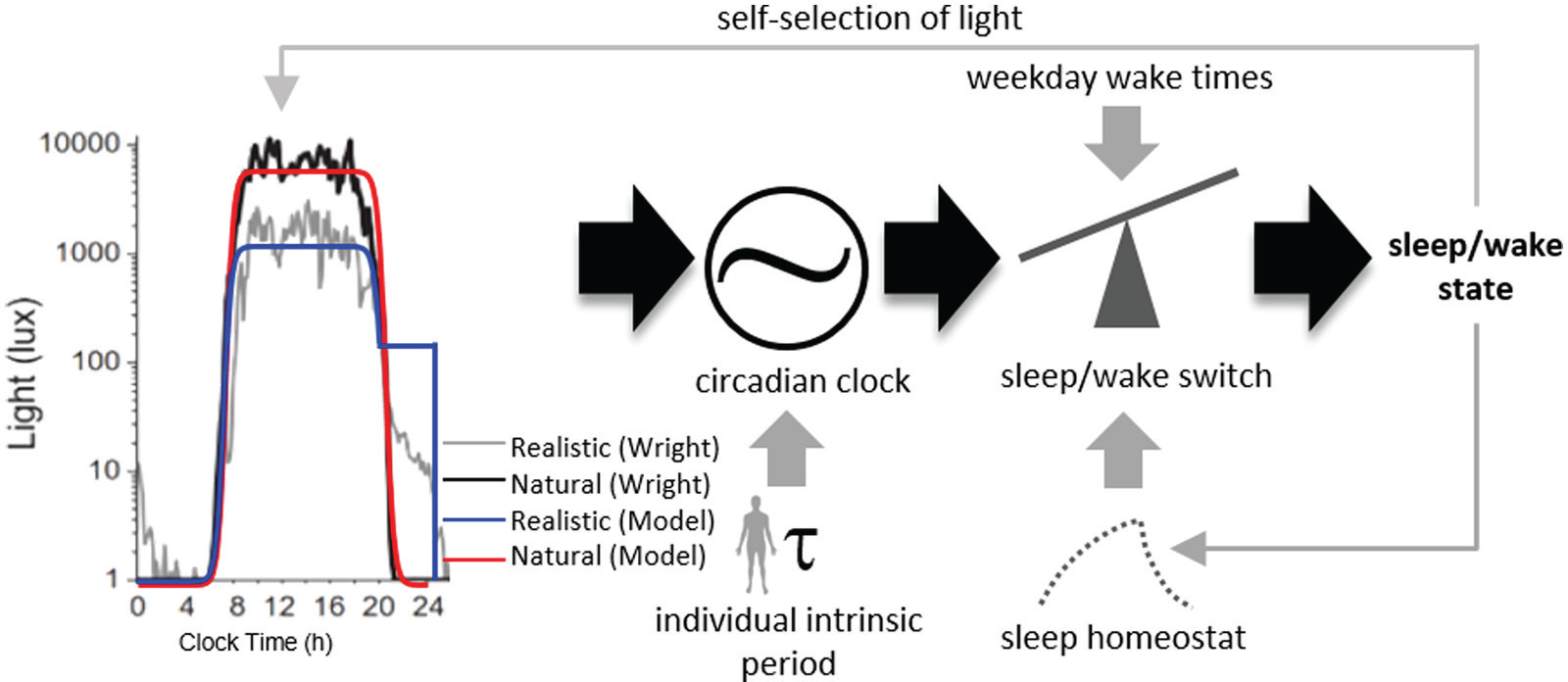
Model schematic with different light exposure patterns. (Left) Light exposure patterns from Wright et al. (2013). (Right) Model schematic. The model alters the light profile based on its sleep/wake state, which is then used as input into the circadian clock. The circadian clock’s phase is influenced by the light profile as well as the intrinsic circadian period. The circadian phase, along with the preset wake and sleep times and sleep homeostat, then drives the sleep/wake switch, which feeds back into the light profile pattern.
Photoreceptors are converted from a ready state to an activated state at a rate
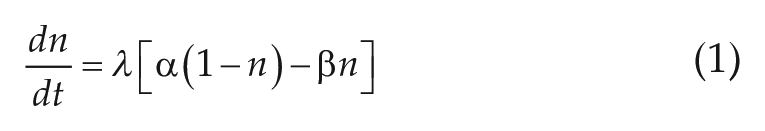
where
The photic drive
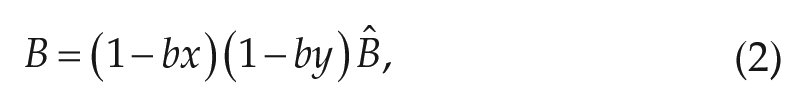

where
The circadian pacemaker is modeled by 2 first-order differential equations, based on a van der Pol oscillator:
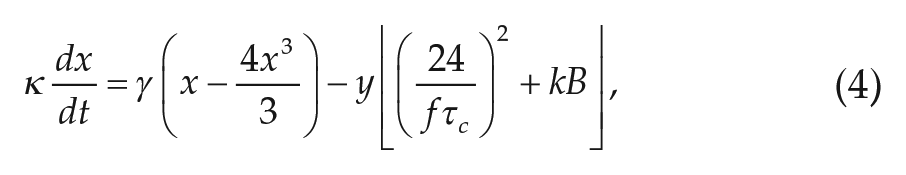

where
To model the ascending arousal system and homeostatic drive, we use the Phillips and Robinson (2007) model that includes the mutually inhibitory wake-promoting monoaminergic (MA) and sleep-promoting ventrolateral preoptic (VLPO) neural populations. Circadian (C) and homeostatic (H) inputs drive the VLPO neural population, while the MA neural population receives excitatory input A from cholinergic and orexinergic sources. For each population
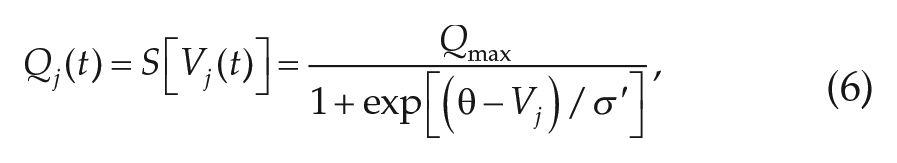
where
The differential equations for the 2 populations are
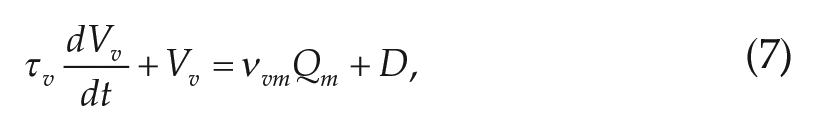
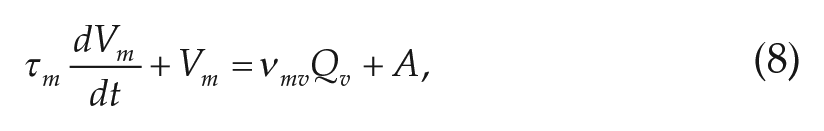
where
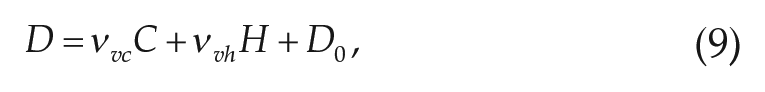
where
The homeostatic drive H accumulates during wake and dissipates during sleep via

where
Simulated Schedules
In previous work, the mathematical model passively selected light, receiving available light when awake and blocking available light when asleep. Here, we use some simple rules for self-selection of light, intended to mimic human behaviors in the real world.
First, we model a natural light profile,
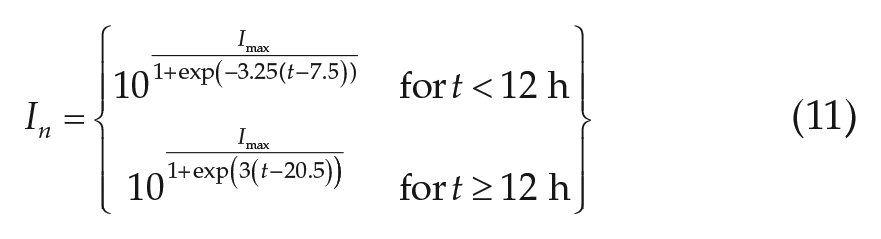
The values of −7.5 and −20.5 were chosen to give sunrise and sunset at the appropriate times, and the values of −3.25 and −3 to match the rate of change of light levels at sunrise and sunset for the camping study environmental conditions. The maximum light parameter,
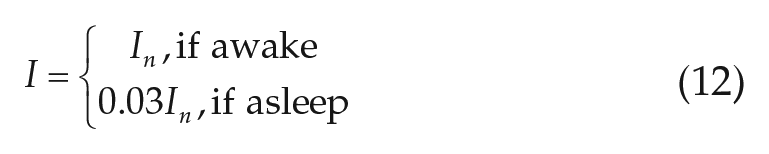
For “realistic” light simulations, we include use of electric light at a room light intensity of 200 lux. Whenever the model is awake and available natural light is below 200 lux, lights are turned on; during sleep, lights are turned off and 97% of available natural light is blocked as in the natural light case. In addition, the maximum daytime light intensity is reduced to 1000 lux by setting
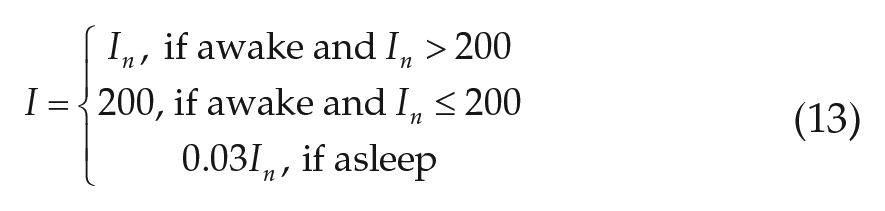
We note that simulations of individuals with different parameter values will generate different profiles, due to different amounts of time spent awake after sunset. While one example of the realistic light profile is shown in Figure 1, model sleep onset times and therefore the timing of artificial light usage can vary greatly with parameter values or with day of week (details below). The level of evening light in the model is therefore not directly comparable to the levels plotted in Wright et al. (2013), which were averaged across multiple participants, some of whom might be awake and some asleep. This averaging resulted in an average level of approximately 20 lux, which is much dimmer than that from most commonly used artificial light sources (e.g., overhead lights and television/computer screens). We assume that individuals use a relatively constant level of artificial light after the sun sets and have negligible exposure to light while asleep. The results we find here are specific to our choice of 200 lux, but similar trends could be found for other choices of evening light levels. In general, simulations with lower artificial light levels will tend toward the natural light case.
For some simulations, we also introduce weekdays and weekends into the model. This was done by forcing the model to be awake from 7:30 AM to 6:30 PM from Monday to Friday (assuming a 9 AM to 5 PM workday and 1.5 h for transit and other activities) and having no sleep restrictions on Saturday or Sunday. This was achieved by forcing
Model Parameter Values
For simulations in this article, most model parameter values are taken directly from the previously published values in Phillips et al. (2010). Here, we analyzed how sleep and circadian timing depend on the value of the intrinsic circadian period,
Two model parameters are updated here to better fit sleep data collected from the human population. These are
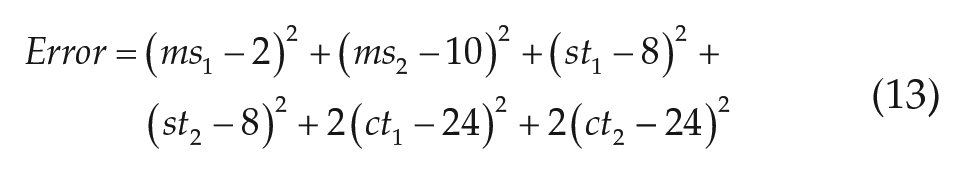
In the above expression,
Simulations and Analysis
To analyze the interaction between the intrinsic circadian period and electric light, we simulated weekday/weekend schedules under either the natural light condition or “realistic light” condition. The dependence of sleep/wake patterns on the value of the intrinsic period was assessed across the physiological range (23.7-24.6 h). For all simulations, the model was allowed 120 days to reach a stable, entrained sleep/wake pattern, which was defined as a pattern that repeated every 7 days to within 0.01 h. Two metrics were used to summarize the outputs of the model: midsleep time and social jetlag. Midsleep time was calculated as the midpoint of sleep onset and wake time, which is a commonly used sleep parameter. Social jetlag was calculated by taking the absolute difference in average midsleep time between weekends and weekdays.
In addition, we performed a sensitivity analysis of the model’s results with respect to the parameters
To compare model predictions with the results of Wright et al. (2013), we also computed sleep times for a population distribution of intrinsic periods. This was achieved by running the simulation 120 times, with the intrinsic period sampled for each simulation from a normal distribution with the mean and standard deviations previously reported for healthy human adults in Duffy et al. (2011). Average sleep onset and offset times were calculated in each case by averaging across the 7 days of the week. Mean and standard deviations from sleep onset and offset times were then computed by averaging across all cases.
To assess circadian timing, we tracked differences in the timing of minimum of the circadian drive
Results
Self-selected Light and Sleep Patterns
For realistic light patterns, different patterns of self-selected light exposure emerged depending on values of the intrinsic circadian period, ranging from 23.7 h to 24.6 h. At the lower end of the physiological range of the period (Figure 2A), the model predicted sleep would occur very early, waking before the 7:30 AM enforced wake time on both weekdays and weekends, and electric light was used both before sunrise and after sunset. Since the model’s sleep times were unaffected by weekday work constraints in these cases, it predicted wake at the same time on both weekends and weekdays (Figure 3A).
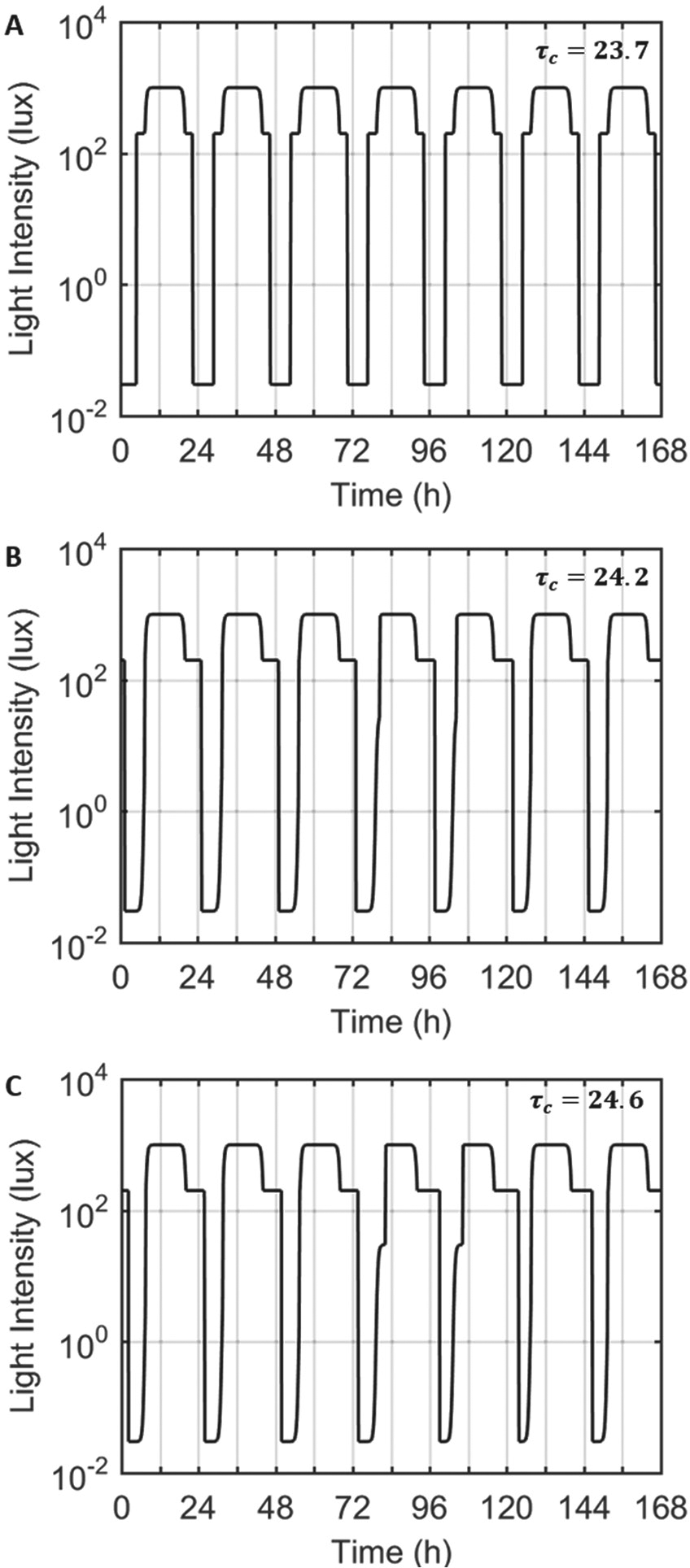
Light profiles input to the model with intrinsic circadian periods of 23.7 h (A), 24.2 h (B), and 24.6 h (C) for 1 week. The first cycle represents the light profile for Tuesday, such that the shaded regions indicate Saturday and Sunday for the model. In case A, with an intrinsic period of less than 24 h, the model turns on the lights both in the morning and in the evening, reflective of an early rise time. In cases B and C, with an intrinsic period longer than 24 h, the model turns on the lights only in the night time.
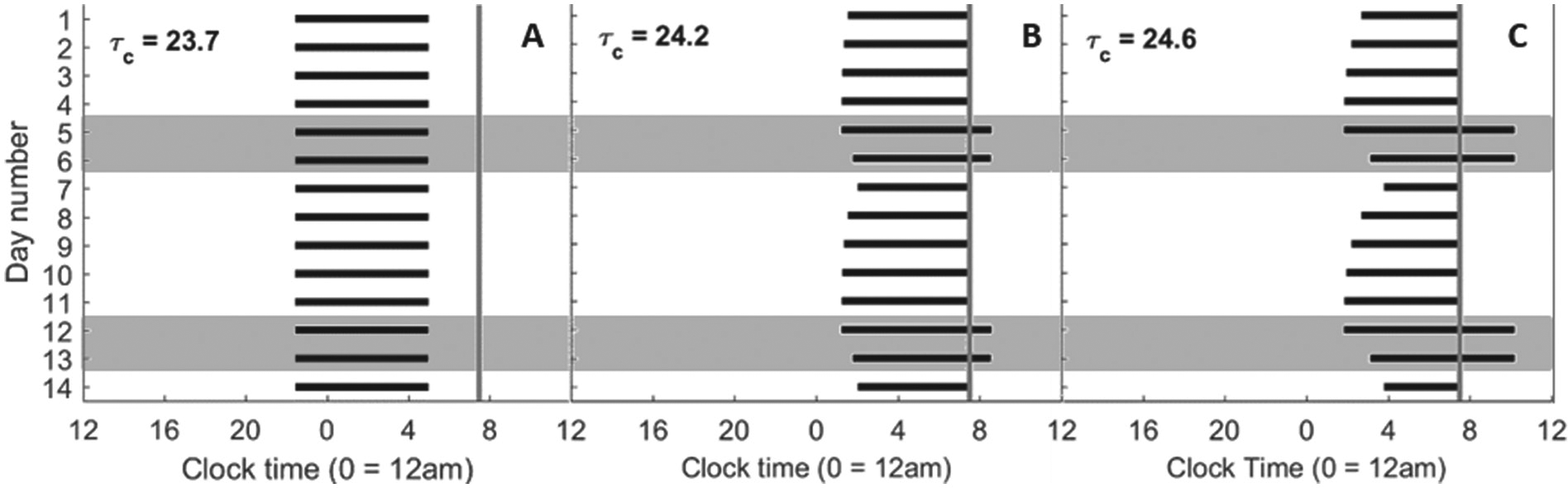
Raster plots of times of sleep over 2 weeks for different intrinsic circadian periods. The black bars indicate times of sleep. The shaded gray boxes indicate sleep on Friday and Saturday nights (weekends). The gray vertical lines indicate the 7:30 AM forced wake time on weekdays.
As the intrinsic circadian period was increased, the weekend rise and bed times moved progressively later. With intrinsic periods of 24.2 h (Figure 2B) or 24.6 h (Figure 2C), the model predicted wake at the enforced time of 7:30 AM on weekdays and predicted sleep to continue beyond 7:30 AM on weekends. Consequently, the model always predicted wake after dawn and only required electric light in the evenings (i.e., after sunset). The model’s predicted sleep/wake patterns differed significantly between weekdays and weekends, with the bed time shifting gradually earlier from Monday to Friday and slipping later during weekends. For an intrinsic period of 24.2 h (Figure 3B), the model’s predicted sleep times were later and sleep duration was longer on the weekend than during weekdays. On Monday morning, sleep was shortened to 5.5 h. This pattern was more exaggerated with an intrinsic period of 24.6 h (Figure 3C), with sleep shortened to only 3.7 h on Monday morning and extending to 8.4 h on Saturday morning.
Effects of Intrinsic Period on Sleep Timing Under Natural Versus Realistic Conditions
Simulations of natural versus realistic light conditions under the same weekday/weekend sleep constraints revealed significant differences in sleep timing. Figure 4 shows how midsleep times on weekdays and weekends differ between these conditions, when the simulation is performed at each
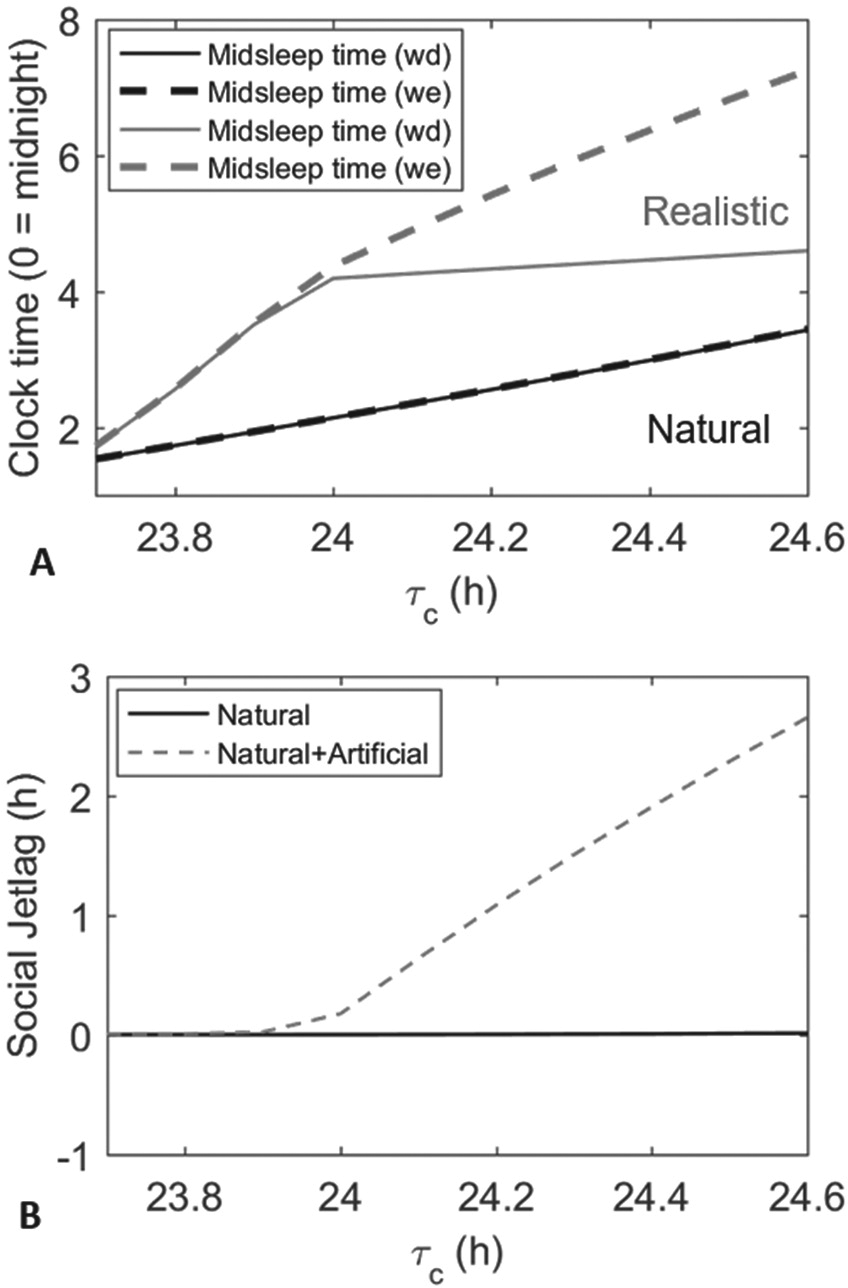
(A) Difference in midsleep timings between natural and realistic light profiles across different
Under a natural light pattern, the model predicted sleep at the exact same time on both weekdays and weekends. Even at the upper end of the physiological range, the model predicted wake before 7:30 AM each day. In addition, there was only a 1.9-h variation in midsleep timing across the full physiological range of intrinsic periods. In other words, without electric light, longer intrinsic periods still produced later sleep times, but only by a small amount compared to the degree of variation observed in the human population.
Under a realistic light pattern, sleep timing was shifted later, and the model produced much greater variation in sleep timing. The difference in midsleep time between the lower and upper ends of the physiological range for the intrinsic period was 2.9 h on weekdays and 5.5 h on weekends. In addition, the difference between midsleep time on weekdays and weekends (social jetlag) expanded as intrinsic period increased, with a maximal difference of 2.7 h for
Comparing Model Predictions to Empirical Data
Using the measured population distribution of intrinsic periods in healthy adults (Duffy et al., 2011), we compared model predictions for means and standard deviations of sleep onset and sleep offset to data published by Wright et al. (2013) for individuals living under natural conditions (no electric light) or realistic conditions (access to electric light), on which we based our simulated light profiles above. The model predictions are similar to the data, shown in Figure 5. With the addition of electric light, sleep times are pushed later and there is much greater population variability in the timing of sleep schedules in both the experimental and model data when compared with natural light sleep schedules. Under realistic lighting, standard deviations for model sleep and wake times were 1.2 h and 1.2 h, respectively, compared to experimental data values of 1.4 h and 0.8 h, respectively. Under natural lighting, these values reduced to 0.8 h and 0.6 h for the model data and 0.3 h and 0.4 h for the experimental data. The model’s sleep duration is shorter under realistic conditions than natural conditions due to forced awakenings on weekdays, whereas this is less apparent in the data.
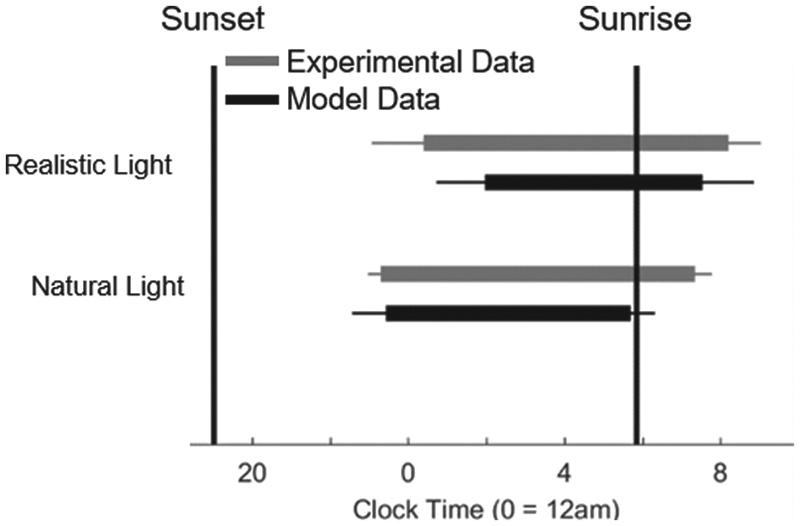
Raster diagram comparing the times of sleep averaged across the entire week between the natural and realistic conditions for both the experimental data and the model. The gray bars represent the data reported in the Wright et al. (2013) study. The black bars represent the data generated by the model. The error bars represent the standard deviation in sleep times for each condition.
Sensitivity of the model’s sleep timing and sleep duration were tested under realistic light conditions for changes of ±5% in the parameters
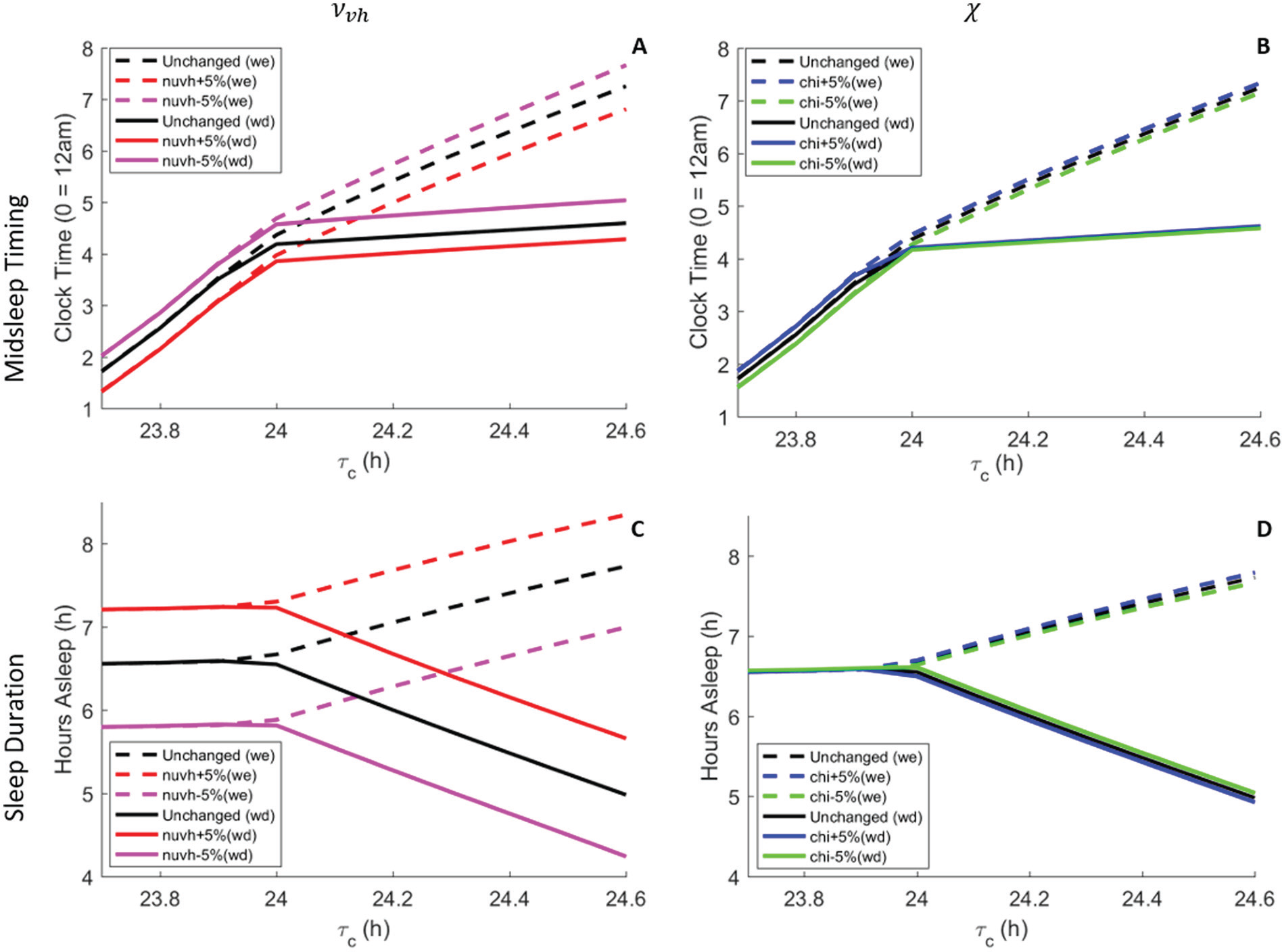
A sensitivity analysis of the model’s results with ±5% changes in the parameters and
Effects of Light Attenuation Versus Light Supplementation on Sleep Timing
Since realistic light profiles involve both attenuation of daytime exposure to natural light and use of supplementary electric light, we investigated the relative importance of these 2 factors by simulating light profiles that included none, one, or both of these factors. As shown in Figure 7, use of electric light is the dominant factor in delaying sleep timing, amplifying individual differences in sleep timing, and generating social jetlag. Attenuation of daytime light in combination with electric light significantly enhances these effects, but it has little effect on sleep timing on its own. With light attenuation only, weekend midsleep times range from 1.4 h to 3.7 h across the range of
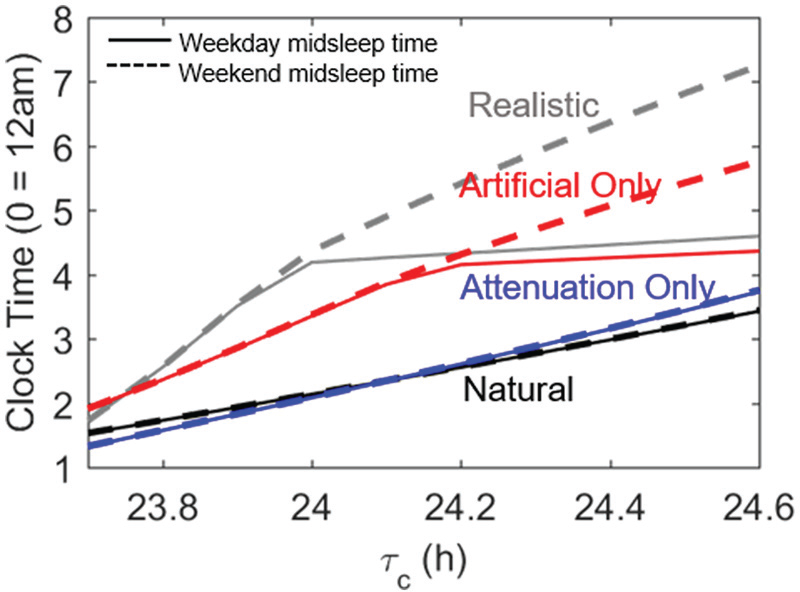
Midsleep time profiles across the natural range of
Discussion
Chronotype is frequently used for describing an individual’s typical sleep timing and is a useful predictive marker for health and other outcomes (Juda et al., 2013; Merikanto et al., 2013). In this article, we show explicitly how chronotype may depend on an interaction between an individual’s physiology, his or her self-selected patterns of light exposure, and constraints on sleep timing on weekdays. Specifically, we show that individual tendencies to sleep late are amplified by self-selected use of artificial light, resulting in a doubling in the range of interindividual differences in sleep timing compared to the same range under natural light conditions. This occurs because individuals with later sleep preference select increased exposure to artificial light in the evening, which delays the circadian clock and sleep onset, reinforcing the behavior by a positive feedback loop. The model prediction that chronotype is an evoked trait that is highly sensitive to environmental factors is supported by the fact that human populations exhibit very different average sleep times between countries (Walch et al., 2016), when camping (Wright et al., 2013), or in preindustrialized societies (Yetish et al., 2015).
A key challenge for our field is linking experimental results generated under highly controlled laboratory conditions to the complex phenotypes that emerge in the real world, where environmental stimuli are complex and there exist feedback loops between physiology and behavior. It should be noted that these challenges are not limited to human work. Drosophila exhibit very different activity patterns under natural light/dark patterns than under typical laboratory light/dark schedules (Prabhakaran and Sheeba, 2014). Mammalian species have also been found to exhibit very different activity patterns in laboratory conditions compared to field or field-analog conditions (Daan et al., 2011), or dependent on the presence or absence of a running wheel (Blanchong et al., 1999). The model we present here includes simple rules for simulating an individual’s modification of his or her environment through self-selection of light. While this is clearly an oversimplification of real human behaviors, it usefully captures empirical phenomena such as the delay of sleep timing on weekends and interindividual differences in sleep timing and it relates these phenomena to underlying physiology. In the future, this approach could be easily extended to include other empirically derived rules, such as individuals choosing to stay up late on weekends when they have no pressure to awaken early the next morning, or more complex social constraints, such as shift work or interactions with other individuals, including friends, roommates, partners, and children. Moreover, our approach could be applied to modeling other human behaviors in which there are interactions between physiology and self-selected behaviors, such as meal timing or drug use and addiction.
An additional consideration for future work is simulation of more realistic individual light profiles. The profiles used here provide relatively constant levels of light during the daytime and evening, reflecting average light profiles measured experimentally. For individuals, however, light profiles can fluctuate on short timescales (e.g., when individuals briefly walk outside), and levels of indoor light can significantly vary between rooms and individual workplaces. The spectral content of light can also differ, with differing effects on the circadian timing system (Lucas et al., 2014), which is a factor not currently implemented in our circadian model.
Outside of sleep and circadian physiology, psychological factors could also influence sleep timing, such an individual’s willingness to curtail use of electronic devices at nighttime. In the model presented here, sleep onset simply occurs when it is enabled by the combination of circadian and sleep homeostatic processes. In reality, individuals may attempt to sleep earlier or may attempt to prolong wakefulness by use of stimulating activities or factors such as social interactions, light, and caffeine. There is notably an association between late chronotype and reduced self-control (Hasler et al., 2013). In addition, studies have shown that intrinsic circadian period can be slightly changed by environmental stimuli, including light/dark patterns (Pittendrigh and Daan, 1976; Scheer et al., 2007). This plasticity is not accounted for in our circadian model but may be an important factor in reinforcing an individual’s evoked chronotype. For example, if consistently delayed schedules lead to a lengthening of the intrinsic period, this would create a positive feedback loop for delayed sleep, which would further interact with self-selection of light.
In summary, this model, which is originally based on laboratory data, replicates experimental data in the real world when we add simple rules for self-selection of light. These results therefore create a bridge between knowledge of sleep and circadian physiology gleaned in the laboratory and more complex sleep behaviors in the real world. Importantly, this provides a basis for quantitatively assessing potential intervention strategies in the real world and their likelihood of success, given knowledge of an individual’s physiology. One of the key predictions from our simulations is that curtailment of evening use of electric light is likely to be a more effective strategy in reducing social jetlag and advancing sleep timing than increasing exposure to daytime natural light.
Footnotes
Acknowledgements
The authors thank Drs. Céline Vetter, Charles Czeisler, Ken Wright Jr., and Till Roenneberg for helpful comments on preliminary versions of these results. This work was supported by the National Space Biomedical Research Institute (HFP02802, HFP0006, and HFP04201 through NASA Cooperative Agreement NCC 9-58), the National Institutes of Health (awards K24-HL105664, P01-AG009975, R01-GM-105018, R01-HL114088, R21-HD086392, K99-HL119618, and R00-HL119618), and a Harvard-Australia Fellowship.
Conflict of Interest Statement
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: EBK has received travel reimbursements from Brain Conference, Employer Health Benefit Congress, and Sleep Technology Council; and consulted for legal firms.