
Abstract
Circadian clocks keep organisms in synchrony with external day-night cycles. The free running period (FRP) of the clock, however, is usually only close to—not exactly—24 h. Here, we explored the geographical variation in the FRP of the linden bug, Pyrrhocoris apterus, in 59 field-lines originating from a wide variety of localities representing geographically different environments. We have identified a remarkable range in the FRPs between field-lines, with the fastest clock at ~21 h and the slowest close to 28 h, a range comparable to the collections of clock mutants in model organisms. Similarly, field-lines differed in the percentage of rhythmic individuals, with a minimum of 13.8% and a maximum of 86.8%. Although the FRP correlates with the latitude and perhaps with the altitude of the locality, the actual function of this FRP diversity is currently unclear. With the recent technological progress of massive parallel sequencing and genome editing, we can expect remarkable progress in elucidating the genetic basis of similar geographic variants in P. apterus or in similar emerging model species of chronobiology.
Keywords
Circadian clocks orchestrate organisms’ metabolism, physiology, and behavior with the periodic alternation of night and day. Locomotor activity under constant dark conditions (DD) is frequently used as the readout for determining the free running period (FRP) of the circadian clocks in animals. The FRP is rarely exactly 24 h. For instance, the FRP is 24.5 to 25.9 h in Homo sapiens (Aschoff and Wever, 1962); ~23 h in the Mediterranean flour moth, Ephestia kuehniella (Kobelkova et al., 2015); ~24 h in the housefly Musca domestica (Codd et al., 2007); and ~23.9 h in Drosophila melanogaster (Petersen et al., 1988). A detailed study of the previously identified variability in the period gene (Costa et al., 1992) further explored the impact of these variants on temperature compensation and thus extended the FRP to 23.1 h to 24.5 h for naturally occurring D. melanogaster strains (Sawyer et al., 1997). For more examples of insect rhythmicity, see the 3rd edition of Insect Clocks (Saunders et al. 2002). Only a few additional studies have addressed the variability of the FRP within the same or very closely related species: the FRP in different field-lines of Nasonia giraulti ranged from 22.5 h to 23.7 h, and the FRP in N. vitripennis ranged from 24.5 h to 26.4 h (Bertossa et al., 2013). The locomotor activity of the mosquito Culex pipiens ranged from 21 h for the Iranian population to 24.4 h in the Japanese strain (Shinkawa et al., 1994). The pupal eclosion rhythm of Drosophila littoralis was explored in particularly great detail. The distribution of this fly ranges from the northern polar circle to Iran, and field-populations from this area were studied with regard to the FRP and photoperiodic conditions, inducing adult reproductive diapause (Lankinen, 1986). The FRP was quickly dampened at DD, particularly in the northern field-lines; yet, a clear cline in the FRP was observed, in which the southern field-lines had slower clocks (longest FRP ~24.3 h in line Kuatisi 7 from Caucasus Mts.), whereas the northern field-lines had faster clocks (shortest FRP ~18.8 h in line Oulu 1 from Finland). A similar comparison of two Himalayan field-lines of Drosophila helvetica revealed a ~26.1 h FRP for the high-altitude strain, originating from 4,121 m above sea level, whereas the low-altitude strain (1,132 m.a.s.l.) showed a remarkably faster circadian clock with an FRP of ~21.7 h (Vanlalhriatpuia et al., 2007).
Given this observed geography-dependent FRP in various insects, we decided to explore the geographic variability in the FRP of Pyrrhocoris apterus, an emerging model of chronobiology. This species has served as an excellent organism of insect physiology for ~50 years (Sláma and Williams, 1966), including in the regulation of adult reproductive diapause (Hodkova, 1976; Smykal et al., 2014), diapause mutants (Dolezel et al., 2005; Socha and Hodkova, 1994), circadian genes (Dolezel et al., 2007, 2008), and in studies addressing the tissue-autonomous role of circadian genes (Bajgar et al., 2013a, b). Adults of P. apterus enter diapause under short days at room temperature, and the photoperiodic response curve is steep (Dolezel, 2015; Numata et al., 1993; Saunders, 1983). Given the large distribution range of this species, from Morocco to Sweden to the north and to the Altai mountains and Mongolia in the east (Socha, 1993), we wondered how stable the FRP is between the field-lines originating from this area. Specifically, we wanted to test whether the northern lines tended to have a shorter FRP, as was shown for D. littoralis (Lankinen, 1986), or a longer FRP, as was found in C. pipiens (Shinkawa et al., 1994). Importantly, any hypothesis stemming from field-line comparisons can be functionally tested in P. apterus, either via RNA interference or with recently established gene editing (Kotwica-Rolinska and Dolezel, unpublished data). In this study, we analyzed 5187 individuals from 59 P. apterus field-lines and found remarkable variability in the FRP, which depended on the latitude of their origin.
Material and Methods
Animals
P. apterus males and females were collected in specified localities (see Table S1 for the location and the year of collection). One field-line was established for each locality, with the exception of Lyon, Chelcice and Marseille, where two lines were used. Lyon-yellow was isolated from the original Lyon-red field-line (Socha and Hodkova, 1994) in generation 130, and these two lines were then kept separately for an additional 100 generations. All field-lines were kept in diapause-preventing constant light conditions (LL, 500–1000 lux) at 24°C to 26 °C on dry linden seed (Tilia cordata) and were supplemented with water.
Activity Monitoring and Analysis
Diapause P. apterus show markedly lower activity than the reproductive individuals (Hodkova et al., 2003; Urbanova et al., 2016). Therefore, only adults kept under diapause-preventing conditions of the Long Day photoperiod (18 h of light, 500–1000 lux, 6 h of darkness) were used. Males (2 to 3 days after adult ecdysis) were individually housed in test tubes (2.5 cm diameter, 15 cm in length), supplemented with water on one side and dry linden seeds on other side, and were positioned in Locomotor Activity Monitors (LAM25; TriKinetics Inc., Waltham, MA) at 25 °C in incubators with constant darkness (DD). The FRP was determined for each individual using the Lomb-Scargle periodogram analysis in ActogramJ (Schmid et al., 2011) under the Fiji platform (Schindelin et al., 2012) for at least 10 consecutive days in DD. Bugs were scored as rhythmic if the PN value reached 15 (see Supplementary material for explanation on the PN value). If multiple periodic components were identified or if the FRP changed during the recording, the animals were considered as rhythmic with complex FRP. See Figure S1 for typical actograms. At least 40 males were monitored for each line tested. Correlations statistics (Pearson’s r) between the FRP, the latitude, and the percentage of rhythmic bugs in the field-line were calculated in R (R Core Team, 2012). In four field-lines, female behavior was determined and compared by t test (Kolomogorov-Smirnov) in GraphPad Prism (GraphPad Software, La Jolla, CA). See Table S1 for a comparison of the rhythmicity between males and females.
Our pilot comparison between individual males and groups of males indicates comparable activity profile during the day; however, the activities of individually housed males were noisier (data not shown). Therefore, we have measured LD activity of 5 males housed together in one glass tube (60 mm in diameter, 30 cm in length), which was horizontally positioned in a LAM60 (TriKinetics Inc., Waltham, MA). The average daily activity profile of the selected field-lines was calculated from at least three tubes (three groups of five males), when each tube was recorded for 10 to 14 days at an LD of 18:6 and at a constant temperature of 25 °C.
Phylogeography
Approximately 1 kb intronic region of the period gene was PCR amplified using forward 5’ACAGCTAGTGGTGGTGAAGAGG 3’ and reverse 5’CTACTGCTCTTACTGAACAACTTTT 3’ primers (Dolezel et al., 2005) and was directly sequenced with an amplification primer. At least three independent animals (and PCR) per line were used to avoid the amplification of artifacts. Chromatograms were visually controlled, and the resulting proofread sequences were aligned using Mafft with the G-INS-i algorithm (Geneious 8.1, Biomatters Ltd., Auckland, NZ). A phylogenetic tree was constructed using the RAxML algorithm (model: GTR CAT+I, 1000 bootstrap replicates). The alignment is available on request from D.D.
Results
Rhythmic Activity of P. apterus
In this study, we measured the locomotor activity of 5187 individual bugs from 59 field-lines, representing 56 localities (Table S1). In case of Lyon, Chelcice and Marseille, two field-lines were established for each locality.
The percentage of rhythmic animals with a clear FRP differed remarkably between the lines, with a minimum of 13.8% in the Skarzysko-Kamienna line and a maximum of 86.8% in the Lyon-red line (Figure 1C and Table S1). The FRPs also differed between individuals within the same field-lines (Figure 1B). Although some field-lines comprised a narrow distribution of FRPs (Lyon-red, Lyon-yellow and Roana), many others covered much larger ranges of FRPs (Toila, Orenburg and Szeged).
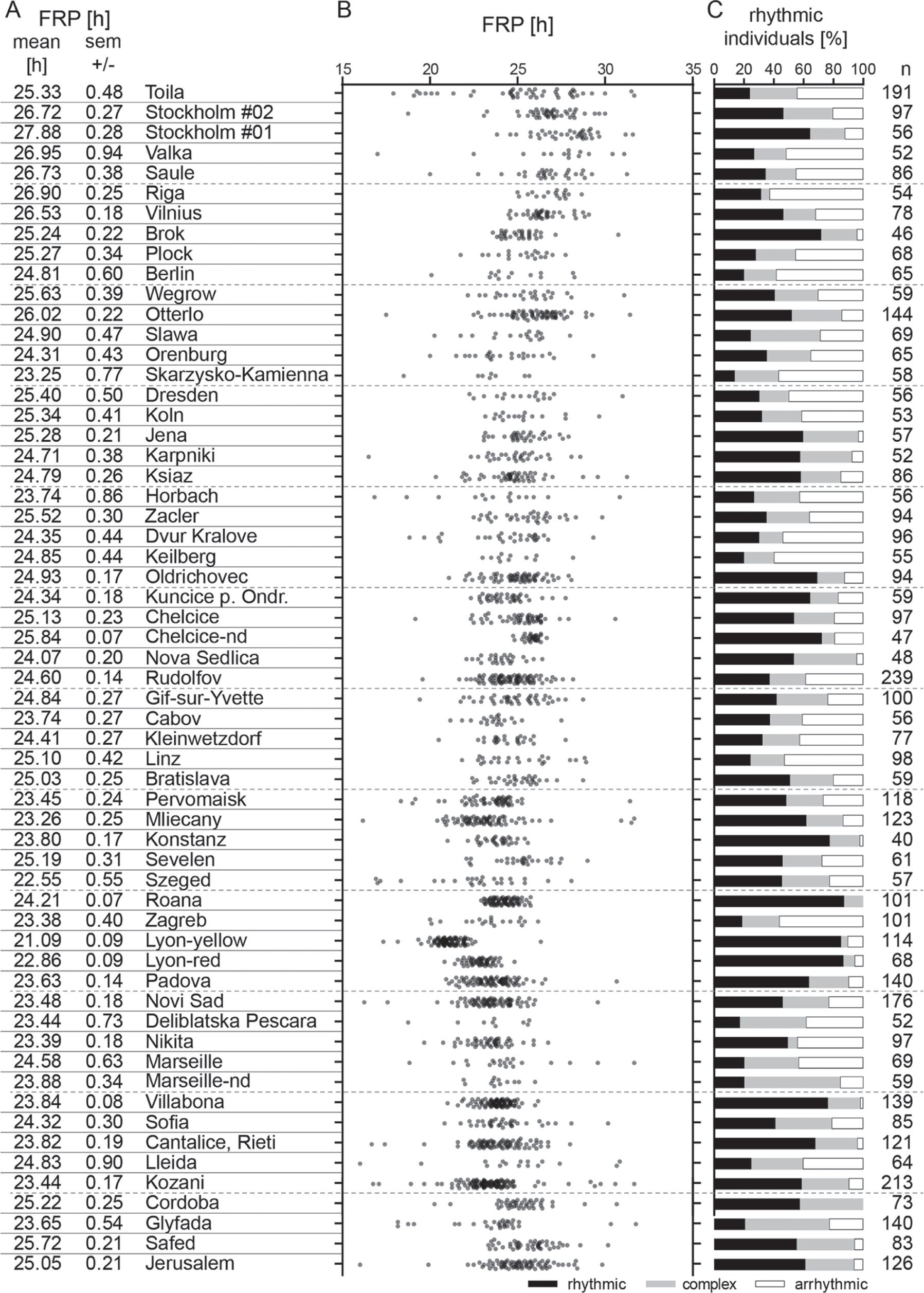
Locomotor activity of P. apterus males recorded at constant dark conditions at 25 °C. (A) List of localities from North (top) to South with the mean free running period (FRP) and the standard error of the mean. (B) The FRP of each individual male is represented by one dot. (C) The percentage of males with rhythmicity with one clear stable period (rhythmic), rhythmicity with more components or unstable period (complex), and males whose behavior did not show a significant period (arrhythmic). The last column shows the total number of males measured in each field-line.
Three pairs of sub field-lines (Lyon-red and Lyon-yellow; Marseille and Marseille-nd; and Chelcice and Chelcice-nd) were analyzed to assess the possible impact of genetic drift on the FRP. The most remarkable difference in the FRP was found in Lyon (1.77 hr, Table S1). Lyon-yellow was isolated from the original Lyon-red field-line (Socha and Hodkova, 1994) in generation 130, and these two lines were then kept separately for an additional 100 generations. We expected that genetic drift would be responsible for the observed differences in the FRP between individual sub-lines (Table S1), yet we cannot rule out the occurrence of spontaneous mutations in some individual lines, such as the extremely short FRP in Lyon-yellow.
We have tested a possible connection between the percentage rhythmicity, the free running period, and locality: the percentage rhythmicity of field-lines did not correlate either to the FRP of these field-lines (Pearson’s r = −0.083, P = 0.136, Figure S2) or to the latitude of the locality from which the given field-lines originated (Pearson’s r = −0.150; P = 0.237, Figure S3). The field-lines with a higher percentage of arrhythmicity also showed a larger standard error of the mean in the FRP (Pearson’s r = 0.7680535, P < 10−6, Figure S4). The FRP correlates with the latitude of the field-line origin (Pearson’s r = 0.508; P < 10−3, Figure 2) and the correlation is stronger, if non-diapause lines are excluded (Pearson’s r = 0.763; P < 10−8, Figure 2). In general, northern field-lines had the longest FRPs (maximum ~28 h in Stockholm #1), whereas central European FRPs were shorter than or close to 24 h (Figure 3). The inbred Lyon-yellow line was exceptional, with an FRP ~21 h, and the very southern lines, including two Israeli field-lines (Safed and Jerusalem) and Cordoba, also displayed a period longer than the 24 h (Figure 1B, Table S1).
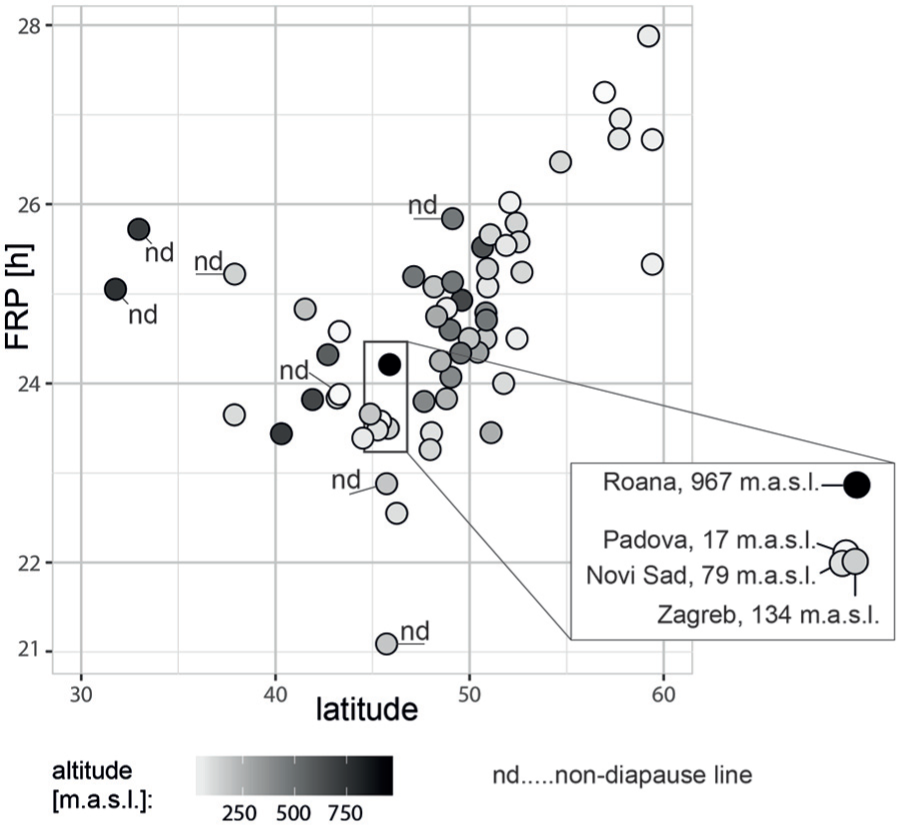
Free running period (FRP; y-axis) plotted over the latitude of the field-line origin (altitude; x-axis). Grey-scale shading for the altitude of locality are in meters above sea level (m.a.s.l.). The enlarged panel shows the details of the field-lines originating from similar geographical areas that differ mainly in altitude. For phylogenetic clustering of field-lines, see also Supplementary Figures S6 and S7.
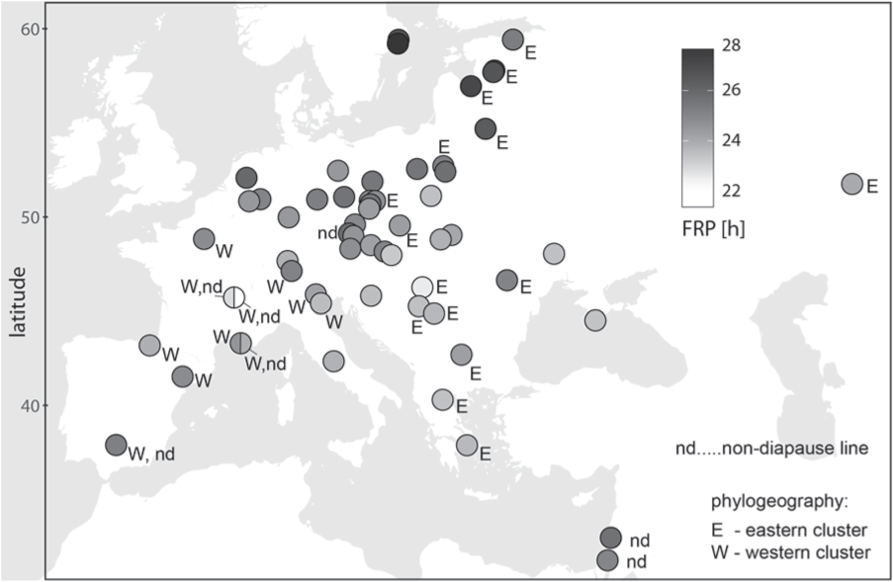
Map depicting the origin of field-lines. Grey-scale indicates the free running period (FRP); W and E indicates the western and eastern phylogenetic clustering, respectively, of a particular field-line. Note the diversity in the FRP is independent of the phylogenetic origin of P. apterus. Field-lines that do not undergo diapause under a short day at 25 °C are labelled with “nd”. Two field-lines from Israel and one from Spain do not enter the diapause from the very beginning after the lines were established. In contrast, three nd lines (Lyon, Marseille and Chelcice) were selected from the wild-type (wt) strains that originally entered diapause.
Male vs. Female Locomotor Activity
Diapause/reproduction status is usually recognized in the females (Smykal et al., 2014) because the male-specific diapause morphology assessment is more laborious (Urbanova et al., 2016). Therefore, we have compared the locomotor activity of males and females in four representative field-lines. Our data indicate that the FRP is indistinguishable between males and females in three of the field-lines. There was a significant difference only between the sexes of the Lyon-yellow field-line (t test, P < 0.05, Figure S5).
Activity in LD Regime
According to Aschoff’s rule, pacemakers with periods longer than 24 h are more efficient in tracking the dawn (Pittendrigh and Takamura, 1989). Indeed, the connection between the circadian phase and the FRP was reported for plants; moreover, these two circadian parameters were latitude-dependent (Michael et al., 2003).
To investigate whether this hypothesis is applicable to P. apterus, 10 field-lines with remarkably different FRPs were assayed for locomotor activity under LD 18:6. In general, all field-lines displayed one activity peak during the photophase, and two lines (Vilnius and Lyon-yellow) were also active in the scotophase (Figure 4). The total activity level (plotted as a number of a beam crosses per hour) also differed remarkably between field-lines (see the y-axis on Figure 4). Overall, there seems to be no clear connection between the FRP in DD and the activity peak in LD (for easier orientation, lines are ordered according to their FRP in Figure 4). The activity peaks were also quite broad, which undermines the adaptive value of precise activity timing in this species.
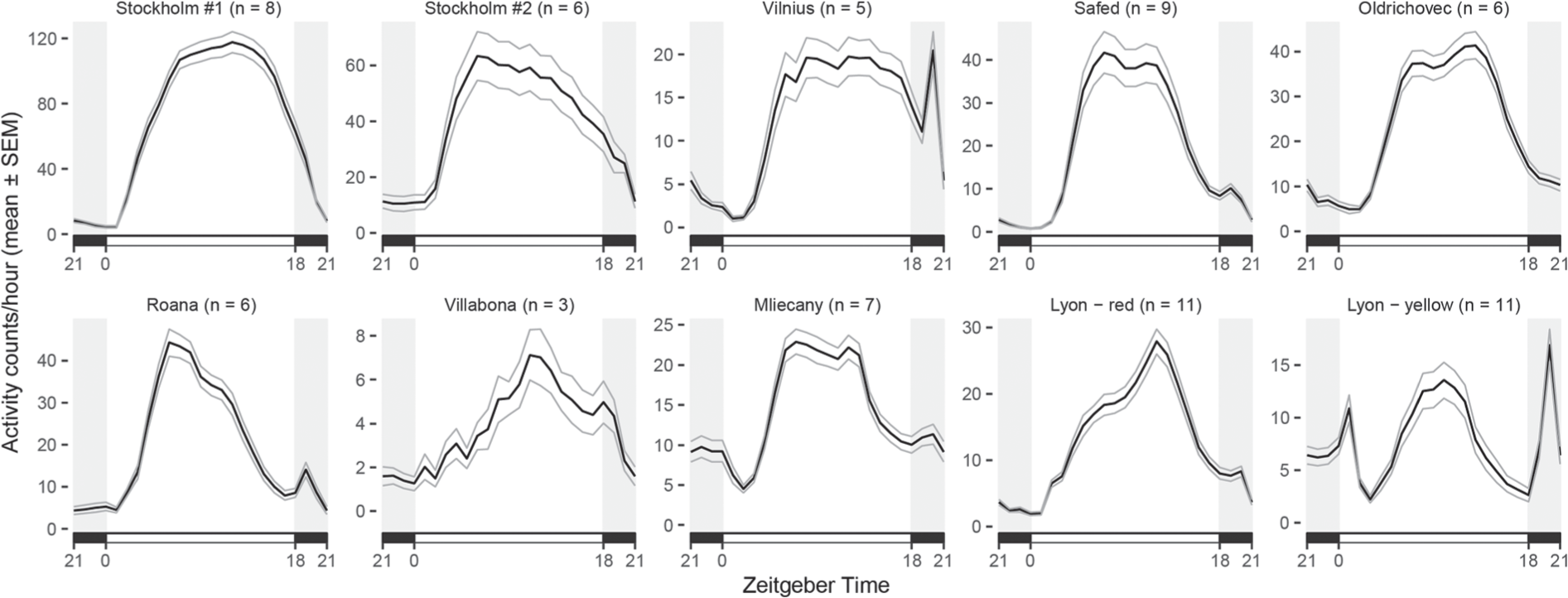
Daily locomotor activity pattern in representative field-lines. Five males were housed in a tube (dia 6 cm × 30 cm) in a LAM 60 (Trikinetics) and were recorded for 10–14 days in LD 18:6. The thick line corresponds to the mean from independent groups (number of groups is indicated as “n”) ± SEM (thinner lines). Field-lines are ordered according to the free running period (FRP) determined at DD: Stockholm #01 (27.9 h), Stockholm #2 (26.7 h), Vilnius (26.5 h), Safed (25.7 h), Oldrichovec (24.9 h), Roana (24.2 h), Villabona (23.8 h), Mliecany (23.3 h), Lyon-red (22.9 h) and Lyon-yellow (21.1 h). Note that the total activity levels (shown on the y-axis as a beam crosses per 1-h interval) differ remarkably between field-lines.
Phylogenetic Relationship between the Field-lines
We performed phylogenetic analysis between selected field-lines to test whether differences observed in the FRP reflect possible close relationships between field-lines, or whether FRP is independent of the evolution of P. apterus. Although the phylogenetic analysis of the intronic region of the period gene resulted in a poorly resolved tree (Figure S6), three clearly distinct clades were identified: 1) Western Europe, 2) Eastern Europe, and 3) two field-lines from Israel (Safed and Jerusalem). The “western clade” contains field-lines with remarkably different FRPs (field-lines from Cordoba ~25 h, Lyon ~21 h and Roana ~24 h). Similarly, the eastern clade contains field-lines with very long FRPs in Baltic field-lines (26 to 27 h, Table S1) and the field-lines with faster clocks (i.e., Szeged ~22 h and Orenburg ~24 h). Undoubtedly, a gradient of FRPs independently evolved (at least twice) in P. apterus (Figure 3 and Figure S6).
Discussion
In this study, we have described surprisingly large differences (up to 7 h) in the FRP between the different geographic field-lines of P. apterus. The observed span nearly corresponds with the differences found in the FRP of various fruitfly clock mutants (Hall, 2003) and, to our knowledge, exceeds the range of the FRP reported for any field-lines. Some extreme phenotypes of individual strains might result from unique mutation. Indeed, exploration of wild-caught lines led to the identification of a unique timelessritsu allele in D. melanogaster (Matsumoto et al., 1999; Murata et al., 1995), and spontaneous mutations in laboratory strains resulted in periodSLIH (Hamblen et al., 1998) and pdf0 (Renn et al., 1999).
Although either genetic drift or the occurrence of spontaneous mutations in some field-lines cannot be excluded, geographic variability in the FRP between ~23 h and ~28 h is supported by several independent field-lines, suggesting that this variability truly reflects the wild-type diversity in the P. apterus circadian clock. Similar to the FRP values, the field-lines also dramatically differ in the percentage of rhythmic individuals. This percentage of rhythmicity is independent on the FRP or on the altitude of their origin. Comparable naturally occurring arrhythmicity was reported for the eclosion of Lucilia cuprina (Smith, 1987).
The actual role of observed large geographic variability is unclear. The periodicity under constant conditions serves as a practical experimental readout of the circadian clock in the laboratory; however, the real selection pressure in environmental conditions has to be different. No clear connection between the FRP in DD and the activity peak in LD undermines the adaptive value of precise activity timing in P. apterus. However, these activity records were obtained from constant temperature conditions. Ideally, the locomotor activity should be obtained under natural conditions, including a temperature profile. This approach revealed a remarkably different behavior in D. melanogaster (Green et al., 2015; Vanin et al., 2012).
The geographic variability in the FRP might reflect an additional role for circadian genes. Indeed, a strong north-to-south cline is observed (Figure 2), with generally longer FRPs in the northern field-lines and a faster circadian clock in the southern field-lines, with the exception of the very southern field-lines (Cordoba, Safed, Jerusalem). Although the phylogenetic analysis of the intronic region of the period gene (Figure S6) resulted in a poorly resolved tree, three clearly distinct clusters were identified: 1) Western Europe, 2) Eastern Europe and 3) Israel (Figure S7 and Figure 3). The existence of western and eastern clades is generally consistent with the colonization trends of northern Europe from southern refugia after deglaciation (Taberlet et al., 1998). Undoubtedly, a gradient of FRPs independently evolved (at least twice) in P. apterus (Figure 3), suggesting some role for these differences in the FRP.
A connection of the photoperiodic timer and circadian clock genes is one possible explanation for the latitudinal gradient in the FRP, provided that these two time-related devices are (at least partially) linked, which is still under intense debate (Bradshaw and Holzapfel, 2010; Dolezel, 2015; Hut and Beersma, 2011; Kostal, 2011; and Saunders and Bertossa, 2011). If that is the case, then P. apterus might “sacrifice” the precision of the circadian clock and use circadian genes to adjust the functioning of the photoperiodic timer. Indeed, recent data confirmed the role of circadian genes in the diapause/reproduction switch of male development and the reproductive behavior in P. apterus (Urbanova et al., 2016). Notably, a latitudinal cline was observed in the eclosion rhythm of Drosophila subobscura (Lankinen, 1993) and D. littoralis (Lankinen, 1986). However, the trend was opposite in both species; the northern field-lines had shorter FRPs, whereas the southern field-lines showed slower eclosion rhythms. D. littoralis also displayed a robust adult reproductive diapause, with a clear gradient from a short critical photoperiod (CPP, the condition when 50% of animals enter diapause) in the south to a long CPP in the north (Lankinen, 1986). Importantly, a selection experiment confirmed a dissociation of the FRP and CPP in D. littoralis (Lankinen and Forsman, 2006).
Although the photoperiodic timer properties were beyond the scope of this study, our future effort will address this particular direction. The steep photoperiodic response curve (PPRC) with clear CPP described in P. apterus (Saunders, 1983; Numata et al., 1993) will allow testing the discussed connection between the photoperiodic timer and circadian clock in this insect. Classical genetics experiments are necessary for testing a possible connection between circadian rhythmicity and photoperiodic timer in P. apterus. Alternatively, very specific and delicate reverse genetic manipulations of circadian clock genes will change the FRP, and the impact on PPRC will be unambiguously assessed.
Further experiments that might shed light on the variability of the FRP will use the field-lines originating from comparable latitudes but with different altitudes, where a longer CPP is necessary to initiate the earlier entry to diapause in high-altitude field-lines. Indeed, flightless P. apterus, with a small dispersal capacity, seems to be an optimal organism for addressing the impact of these factors or to study any local adaptations (Hut et al., 2013). Unfortunately, the dataset presented here contains an insufficient number of altitudinal field-lines, preventing a rigorous analysis; yet, a few examples indicate a possibly positive correlation between the altitude and FRP (Figure 2, Table S1). For instance, the population from Roana, a mountain village of almost 1000 m.a.s.l., has a slower clock when compared to Padova, which is a close locality but is at sea level (17 m.a.s.l.). Another example comes from the Czech/Poland border, separated by the Krkonose mountains, where both the southern and northern field-lines have a shorter FRP than the central Zacler population, originating from 620 m.a.s.l. Notably, a comparable influence of altitude on the FRP was reported for D. helvetica, where the difference between low and high altitudes was 4.4 h (Vanlalhriatpuia et al., 2007), with a slower clock in the high-altitude line. Unfortunately, no data correlating the latitude and FRP are available for D. helvetica, and, similarly, missing altitudinal field-lines of D. littoralis prevent any comparison of altitudinal/latitudinal influence.
Taken together, P. apterus shows remarkable variability in the FRP, which is comparable to the phenotypic span of laboratory-induced circadian mutants in model organisms. The role of this variability is unclear but can be unambiguously addressed with emerging technologies, including population genomics tools (Bastide et al., 2013) and genome editing recently established in P. apterus (Kotwica-Rolinska and Dolezel, unpublished).
Footnotes
Acknowledgements
We thank many colleagues for their generous and enthusiastic help during the collection of live P. apterus: Carl-Cedric Coulianos, Manuel Baena, Rodolfo Costa, Lucia Salis, Marketa Ondrackova, Adam Bajgar, Milena Damulewicz, Enrico Bertolini, Petra Sekyrova, Radka Zavodska, Lukas Cizek, Lukas Drag, Dora Nagy, Plamen Kalushkov, Jana Pavlova, Magda Hodkova, Julius Lukes, Iva Fukova, Xanti Pagola-Carte, Penelope Mavragani-Tsipidou, Elene Drosopoulou, Frantisek Marec, Matilde Eizaguirre, Ramon Albajes, Stanislav Rada, Vladimir Kostal, Tomas Ditrich, Martin Kaltenpoth, Jaromir Cihlar, Daniel Jablonski and many others (we apologize to anyone not mentioned). L. P. was supported by INsecTIME (FP7-PEOPLE-2012-ITN, grant no. 316790) and J. K-R by Postdok_BIOGLOBE (CZ.1.07/2.3.00/30.0032), and the complete work was funded by the National Science Foundation (GACR, grant no. 14-32654J) to D.D. The work of Petr Kment was supported by the Ministry of Culture of the Czech Republic to the National Museum (DKRVO 2016/13, 00023272).
Conflict of Interest Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.