
Abstract
The Arabian oryx inhabits an environment where summer ambient temperatures can exceed 40 °C for extended periods of time. While the oryx uses a suite of adaptations that aid survival, the effects of this extreme environment on inactivity are unknown. To determine how the oryx manages inactivity seasonally, we measured the daily rhythm of body temperature and used fine-grain actigraphy, in 10 animals, to reveal when the animals were inactive in relation to ambient temperature and photoperiod. We demonstrate that during the cooler winter months, the oryx was inactive during the cooler parts of the 24-h day (predawn hours), showing a nighttime (nocturnal) inactivity pattern. In contrast, in the warmer summer months, the oryx displayed a bimodal inactivity pattern, with major inactivity bouts (those greater than 1 h) occurring equally during both the coolest part of the night (predawn hours) and the warmest part of the day (afternoon hours). Of note, the timing of the daily rhythm of body temperature did not vary seasonally, although the amplitude did change, leading to a seasonal alteration in the phase relationship between inactivity and the body temperature rhythm. Because during periods of inactivity the oryx were presumably asleep for much of the time, we speculate that the daytime shift in inactivity may allow the oryx to take advantage of the thermoregulatory physiology of sleep, which likely occurs when the animal is inactive for more than 1 h, to mitigate environmentally induced increases in body temperature.
The Arabian oryx is the smallest member of the genus Oryx and is native to the gravel and hard sands of the deserts and steppes of the Arabian Peninsula. In the summer months, peak ambient temperatures in the Arabian Desert regularly surpass 40 °C for prolonged periods of time. To cope with this hot and dry environment, the Arabian oryx uses a range of behavioral and morphophysiological responses to high temperatures. During the summer, Arabian oryx have been observed to increase the time spent inactive in the shade, where they appear to dig shallow depressions in which to rest (Stanley Price, 1989; Seddon and Ismail, 2002), creating a cooler microclimate (Hetem et al., 2012b). In addition to these behavioral strategies, the Arabian oryx has morphophysiological capacities that assist its survival, such as reduced fasting metabolic rates, lower total evaporative water loss rates, lower water influx/turnover rates compared with other similar-sized artiodactyls (Williams et al., 2001), adaptive heterothermy (Ostrowski et al., 2003; Hetem et al., 2010), and selective brain cooling (Hetem et al., 2012a). This suite of behavioral and morphophysiological features undoubtedly assists the Arabian oryx to survive in the desert of the Arabian Peninsula, although these features appear to be common to all species of oryx living in arid environments and are not specific adaptations of the Arabian oryx (Taylor, 1966, 1968, 1969, 1970a,b; Taylor and Lynn, 1972; King et al., 1975; King, 1979; Stanley Price, 1985).
Seddon and Ismail (2002) and Hetem et al. (2012b) indicated the possibility of an altered daily activity pattern between the cooler and hotter months in the Arabian oryx, although this was not explored in detail in either of those studies. Among many mammals, the dominant, or primary, zeitgeber for entraining circadian rhythms, including daily activity patterns, is ambient light (e.g., Daan and Aschoff, 1975; LeGates et al., 2014); however, under natural conditions, activity patterns may not be driven exclusively by the endogenous circadian clock but can be influenced by environmental conditions that bypass or act downstream of the clock (“masking”). Intricately linked with the entrained daily activity pattern is the timing of sleep and wakefulness, as sleep mostly occurs during the phase of general and prolonged inactivity (Monk, 1991).
The Arabian oryx can undergo substantial seasonal changes in the amplitude of its daily body temperature rhythm, although the timing of the rhythm does not appear to change seasonally (Hetem et al., 2010). To mitigate the effects of seasonal changes in ambient temperature, the Arabian oryx might alter its daily activity pattern, but it is unclear whether the timing of inactivity, and potentially of sleep, also changes in relation to the seasons (Seddon and Ismail, 2002; Hetem et al., 2012b). To determine whether daily inactivity patterns change seasonally or remain the same in the Arabian oryx, we used actigraphy (Ancoli-Israel et al., 2003) to analyze the timing of daily inactivity patterns during both summer and winter months.
Materials and Methods
Animals
In the current study, a total of 10 adult Arabian oryx were used. These animals formed 2 groups: Group 1 consisted of 9 adult males (mean body mass 79.8 ± 11.0 kg) being used for fine-grain actigraphy and body temperature recording in the Mahazat as-Sayd Protected Area, northeast of Taif, Saudi Arabia (22° 12′12″N, 41° 49′36″E) in February and July 2014, and group 2 consisted of 1 adult male (85 kg body mass) being used for fine-grain actigraphy near Richmond in the Karoo region of South Africa (31° 32′29″S, 23° 41′34″E) during February 2015. Ethical clearance for the current study was obtained from the University of the Witwatersrand Animal Ethics Committee (clearance certificate number 2014/53/D), and all animals were treated according to the guidelines of this committee, which parallel those of the National Institutes of Health (NIH) for the care and use of animals in scientific experimentation.
Anesthesia of the Arabian Oryx
All animals were remotely injected using a Dan-Inject dart gun (Daninject, Børkop, Denmark) with etorphine hydrochloride (Captivon 98, Wildlife Pharmaceuticals Ltd., White River, South Africa; 19 µg/kg; a semisynthetic opioid that has an analgesic effect and is an opioid receptor antagonist); ketamine (Ketaminol Vet., MDS Animal Health, Intervet International B. V., Boxmeer, The Netherlands; 0.3 mg/kg; a dissociative anesthetic agent that is an NMDA receptor antagonist); midazolam (Wildlife Pharmaceuticals Ltd., White River, South Africa; 0.13 mg/kg; a benzodiazepine class anesthetic agent that acts by enhancing the effect of GABA on GABAA receptors); and medetomidine (Zalopine 10 mg/ml, Orion Pharma, Espoo, Finland; 5 µg/kg; a sedative analgesic that is an α2-adrenergic agonist). All animals received oxytetracycline (Terramycin LA, Pfizer, Brazil; 20 mg/kg IM; a broad-spectrum antibiotic) and ketoprofen (Ketovet, Vemedim Animal Health, Can Tho City, Vietnam; 2 mg/kg IM; a propionic acid class of nonsteroidal anti-inflammatory that acts to inhibit the production of prostaglandin). After the procedure, anesthesia was reversed using naltrexone hydrochloride (Naltrexone, APL, Kungens Kurva, Sweden; 40 mg IM; an opiate antagonist) and atipamezole hydrochloride (Antisedan, Orion Pharma; 2 mg IM; a synthetic α2-adrenergic receptor antagonist). After the recording period, the animals were again immobilized using a similar drug combination, and the implants were removed. Again, oxytetracycline and ketoprofen were administered, and the animals were allowed to recover in a shaded pen prior to release to their normal environment.
Surgery for Fine-Grain Actigraphy and Body Temperature Recording
Following recumbency, the animals were moved to a shaded area for surgery. With aseptic techniques, a subcutaneous pocket was created on the left lateral aspect of both the neck and thigh, and an Actiwatch embedded in an inert wax (SasolWax 1276, Sasol, Johannesburg, South Africa) was placed into each pocket. The Actiwatch ambulatory device is commonly used for measuring sleep in humans. Within each Actiwatch is a piezo accelerometer device connected to a microchip that sums and records the number of acceleration events for each minute. The Actiwatch Spectrum (Philips Respironics, Philips) was used in the current study, which has a weight of 25 g and approximate dimensions of 35 × 35 × 12 mm. Each Actiwatch, factory calibrated, was programmed (data acquisition rate set at 1-min intervals) with Philips Respironics Actiware 5 software (Actiwatch communication and sleep analysis software) prior to implantation. The wristbands from the Actiwatches were removed, and the watches were insulated with standard electrical insulation tape and covered with 2 coats of biologically inert wax. The implantation sites of the Actiwatches (i.e., the side of the neck and the hindleg) were shaved, washed with chlorhexidine, and disinfected with alcohol. Sterile drapes were taped in place over the respective surgical sites to isolate the disinfected area. A small incision (less than 50 mm in length) was made at each of the respective implantation sites, and a subcutaneous pocket extending approximately 100 mm ventrally from the incision site was created. The Actiwatch, sterilized within a container with formalin pellets for 48 h prior to implantation, was inserted into the respective pockets, and the incisions were closed in an everting pattern with interrupted sutures, sterilized once again, and liberally sprayed with Necrospray (Bayer Animal Health). A data logger that measured temperature was inserted intra-abdominally (AIC001: Abdominal implant, Africa Wildlife Tracking, South Africa). For recording body temperature, the device was calibrated against an accurate thermometer in an insulated water bath, providing an accuracy of better than 0.06 °C. A small incision (less than 80 mm in length) was made through the skin at the implantation site, and the muscle layers of the abdominal wall were separated using blunt dissection. The implant, coated in inert wax and sterilized within a container with formalin pellets for 48 h prior to implantation, was inserted untethered into the abdomen; the incisions were closed and liberally sprayed with Necrospray.
Following reversal of the anesthesia, the animals were released into a 2 × 1-km fenced area of natural oryx habitat and were not provided with supplementary food or water for the entire recording period. For the single animal studied in South Africa, the body temperature logger was not implanted, but a radio transmitter collar was attached to the neck of the animal and the animal was released back to the wider game reserve (which measured 25 × 25 km) and not provided with supplementary food or water. After the recording period, the implanted animals were relocated and reanesthetized, and the Actiwatches and body temperature loggers were retrieved.
Data Analysis for Fine-Grain Actigraphy
Phillips Respironics Actiware 5 was used to retrieve the recorded data from each of the implanted Actiwatches. The raw data were exported to Microsoft Excel, where they were manually scored and analyzed. Data obtained from the neck and leg actigraphs were scored concurrently in 1-min epochs as either active or inactive. For an epoch to be scored as active, either the neck or leg actigraphs had to have an activity score greater than zero. Inactive epochs were scored when both the neck and the leg actigraphs had an activity score equal to zero. From the 1-min scored data, the modal state for 5 min was calculated and used to determine total active-inactive time, number of active-inactive episodes, and the average duration of active-inactive episodes. The data presented in the Results section is the grouped data for all individuals, with the standard deviation provided in parentheses following averages.
Data Analysis for Body Temperature
The temperature data obtained from the abdominal implants were downloaded into Microsoft Excel files for processing. The animals from each season were analyzed individually and the data were then pooled. Body temperature from each 10-min recording period was averaged across the recording period, and the data were then pooled to create an overall average body temperature.
Recording of Weather Conditions
A permanent weather station within the 1 × 2-km pen at the Mahazat as-Sayd Protected Area recorded weather parameters throughout the recording periods in Saudi Arabia. This weather station provided a reading of ground temperature, air pressure, air temperature, humidity, wind speed, wind direction, rainfall, and solar radiation every 15 min throughout both recording periods. The date and time stamps on the data were used to match the different recordings. In Richmond, South Africa, a portable solar-powered weather station (Environdata, www.environdata.com.au/easidatamark4), which recorded, every 10 min, air temperature, black globe temperature, relative humidity, wind speed, wind direction, rainfall, solar radiation, and illuminance (lux), was placed within the game reserve during the recording period. Sunrise and sunset times were obtained from various freely accessible databases.
Results
The current report outlines our findings regarding seasonal effects on daily inactivity patterns in Arabian oryx studied in 2 different environments. Initially, we used fine-grain actigraphy (1-min resolution, 2 devices per animal, subcutaneous neck and hindleg) for 28 days in both a cold month (February) and a hot month (July) in the Mahazat as-Sayd Protected Area, northeast of Taif, Saudi Arabia, to reveal that there was a seasonal shift in the pattern of daily inactivity from essentially nocturnal in the cold month to bimodal (with the additional appearance of inactivity during the daytime) in the hot month. In addition, we used fine-grain actigraphy to examine an oryx from the Karoo region of South Africa (28 days) in February (i.e., exposed to a photoperiod comparable to the Saudi Arabian summer but ambient high temperatures resembling the Saudi Arabian winter); this individual expressed a daily inactivity pattern similar to the animals recorded in the Saudi Arabian winter.
Seasonal Variation in the Daily Inactivity Pattern
Nine male oryx (mean body mass 79.8 ± 11.0 kg), 4 animals in February 2014 (winter) and 5 animals in July 2014 (summer), were used for this portion of the study (see Table 1 for environmental data for each month). During the winter recording period, the average daily inactivity measured was 5.01 h (±1.71 h). The animals displayed polyphasic inactivity with an average of 18 (±4) bouts of inactivity per day. The average daily onset of the main inactive bout (that being the longest period of inactivity) occurred between 0330 and 0400 h, with this bout having an average length of 127 min, just over 2 h (±63.5 min). Thus, during the winter recording period, the oryx displayed a nocturnal inactivity pattern, with the vast majority (94% ± 7.25%) of inactivity occurring during the night (Figs. 1 and 2).
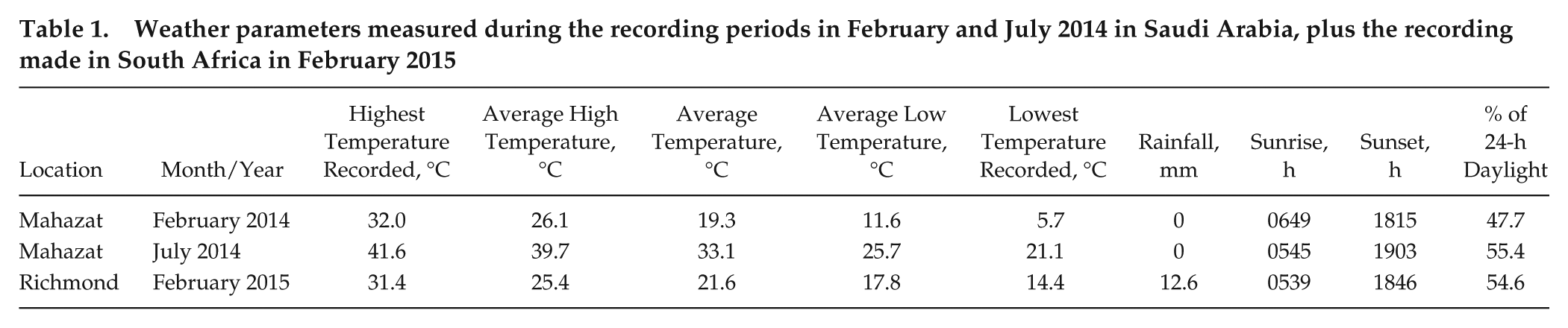
Weather parameters measured during the recording periods in February and July 2014 in Saudi Arabia, plus the recording made in South Africa in February 2015
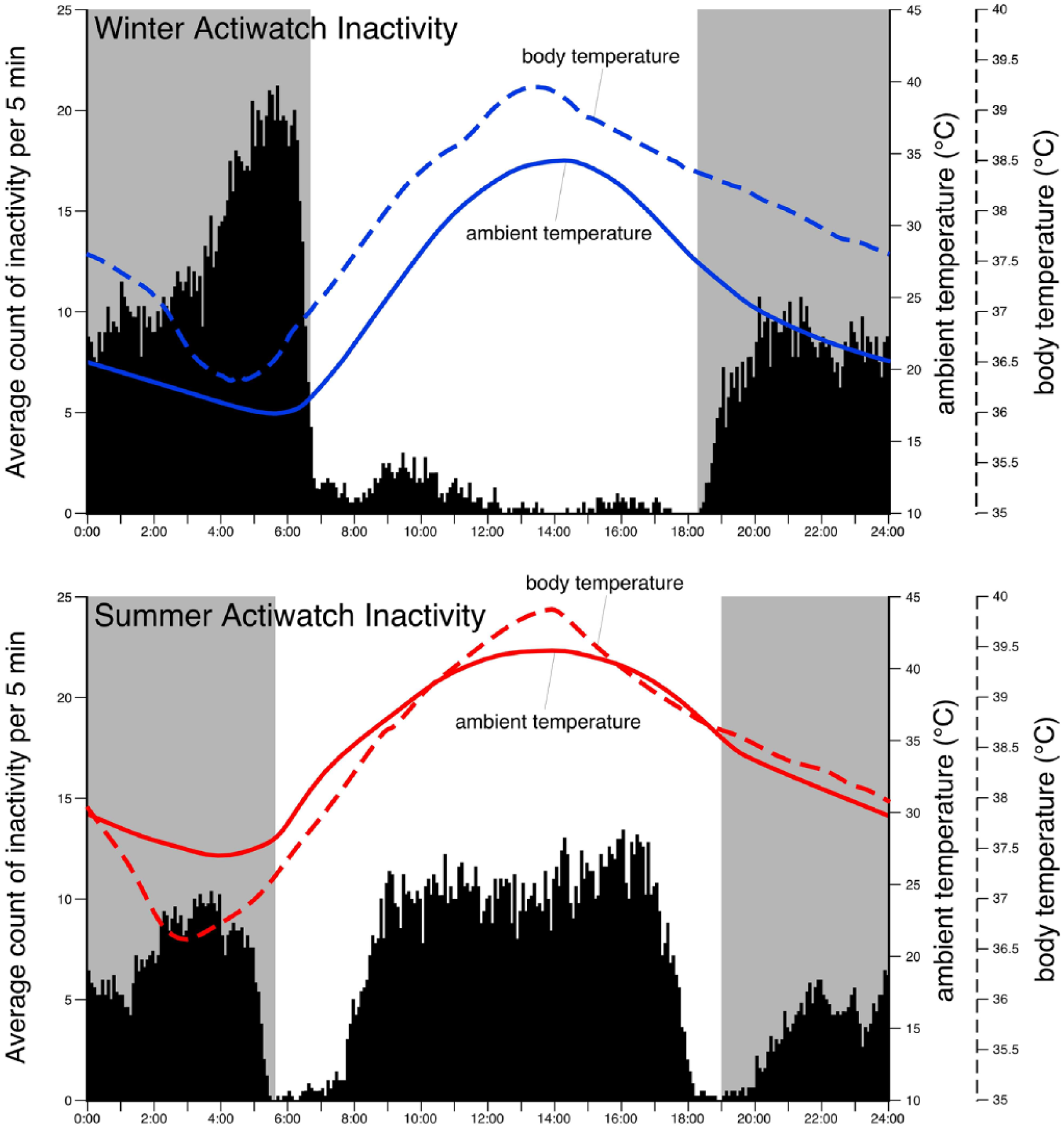
Graphs illustrating the average count of inactivity for any given 5-min period scored over 28 days in the winter month (upper graph, February 2014, Saudi Arabia, pooled from the 4 animals recorded) and the summer month (lower graph, July 2014, Saudi Arabia, pooled from the 5 animals recorded). Note the nocturnal pattern of inactivity during the winter month, with little inactivity occurring during the daytime. In contrast, in the summer month we observed a bimodal inactivity pattern, with the additional appearance of inactivity during the daytime. The average ambient (solid lines) and body (dashed lines) temperatures, recorded every 10 min, across the month of recording are overlain on the activity graphs (online, blue for winter, red for summer). Note the substantially higher average ambient temperatures in the summer month. Note that the timing of the daily rhythm of body temperature remains unaltered between seasons. The gray regions represent the period between dusk and dawn.
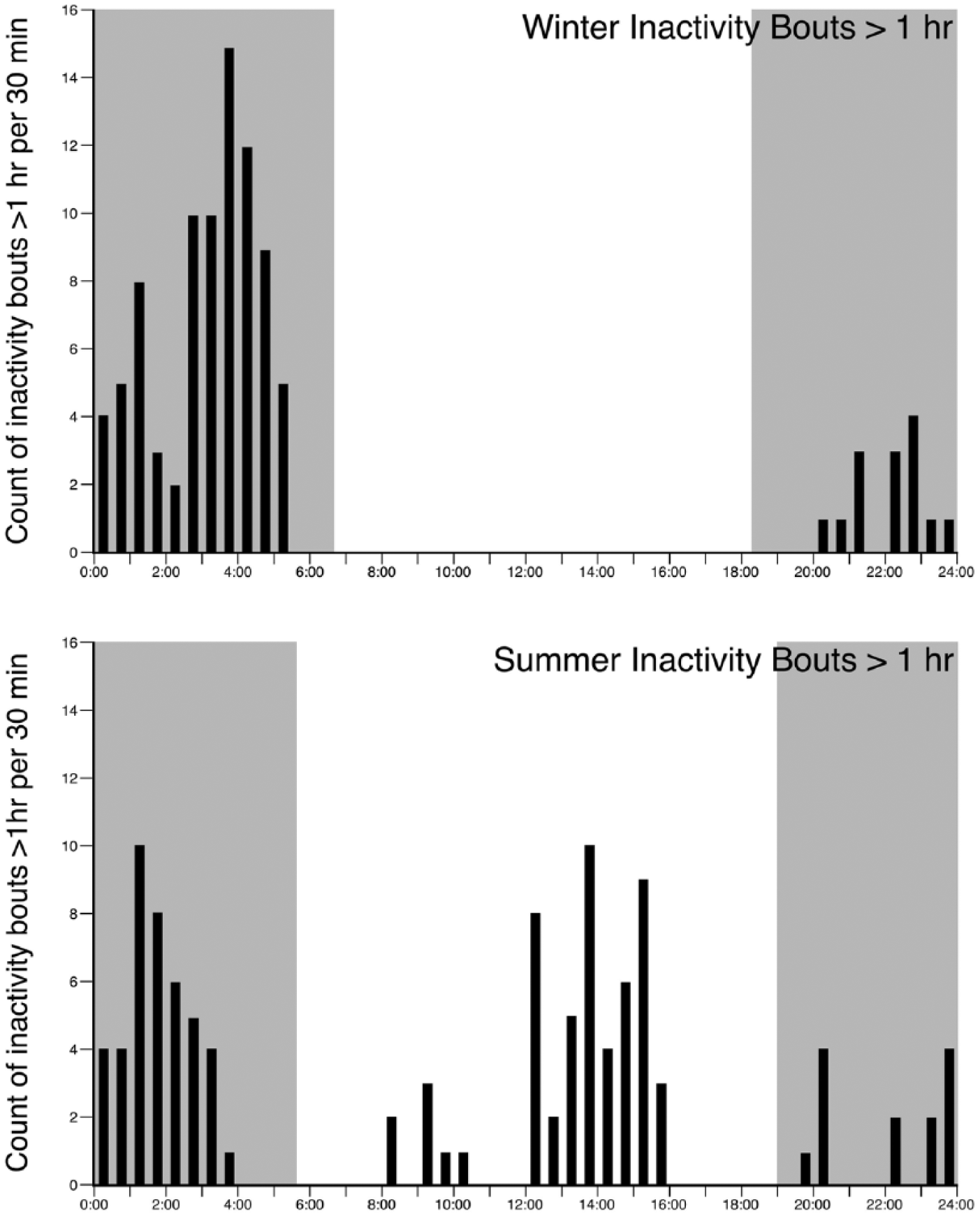
Frequency distribution graphs showing the number (in 30-min intervals) of inactive bouts of greater than 1 h duration during the winter and summer recording periods for the Arabian oryx measured with fine-grain actigraphy in Saudi Arabia. Note that during the winter, all bouts of inactivity greater than 1 h occurred between sunset and sunrise (gray-shaded regions), with the vast majority (97/111 bouts, 87.4%) occurring in the early morning hours between midnight and dawn. In contrast, in summer, the majority of inactive bouts (55/110 bouts, 50%) occurred during daytime, with 42/110 bouts (38.2%) occurring between midnight and dawn. The number of inactive bouts occurring between dusk and midnight was similar between seasons (winter: 14 bouts, 12.6%; summer: 13 bouts, 11.8%). These differences are statistically significant (daytime summer vs. daytime winter, Mann-Whitney, nonparametric test, P = 9.14 × 10−5; nighttime summer vs. nighttime winter, Mann-Whitney, nonparametric test, P = 2.15 × 10−5).
During the summer recording period, the average daily inactivity measured was 6.62 h (±1.93 h). The animals displayed polyphasic inactivity with an average of 27 (±6) bouts of inactivity per day. In this summer recording session, the main inactive bouts were found to have onsets between 0100 and 0130 h or between 1330 and 1400 h. Thus, prolonged periods of inactivity were observed at 2 different times during the 24-h period (Fig. 2). These main inactivity bouts had an average length of 137 min, just over 2 h (±73.4 h). Thus, when compared with winter, during the summer the daily inactivity pattern of the oryx changed dramatically, showing a bimodal pattern and having an extra 1 h and 37 min of inactivity (Mann-Whitney, nonparametric test, P = 1.25 × 10−12). The additional inactivity occurred mostly during the afternoon but was also present in the predawn hours (Figs. 1 and 2). During the summer, 68% (±24.6%) of inactivity occurred during the daytime compared with 6% in the winter (statistically significant difference, Mann-Whitney, nonparametric test, P = 2.49 × 10−30), while only 32% (±24.6%) of inactivity occurred during the nighttime in summer compared with 94% in the winter (statistically significant difference, Mann-Whitney, nonparametric test, P = 4.21 × 10−8). Despite this seasonal variation in the inactivity pattern, we could find no seasonal change in the timing of the daily body temperature rhythm, although the amplitude did change (Fig. 1), confirming the results of an earlier study (Hetem et al., 2010).
Is Photoperiod or Ambient Temperature the Dominant Factor Changing the Daily Inactivity Pattern?
To begin to distinguish whether photoperiod or ambient temperature is the factor causing the seasonally switching daily inactivity patterns in the Arabian oryx recorded in Saudi Arabia, we examined a single adult male (body mass estimate = 85 kg) from a free-ranging population near Richmond, in the Karoo region of South Africa. The advantage of including this particular individual is that the weather in the Karoo, while hot in summer (February), does not reach the temperatures recorded in summer in Saudi Arabia but matches the temperatures seen in winter in Saudi Arabia (Table 1); however, the day lengths in South Africa during the summer are similar to those seen in Saudi Arabia during the summer (Table 1).
With this individual animal we used fine-grain actigraphy recording and found that the Arabian oryx in South Africa displayed a predominantly nocturnal inactivity pattern (Fig. 3), similar to that seen in the winter recordings of oryx in Saudi Arabia (Fig. 1). During the summer recording period in South Africa, the average daily inactivity measured was 5.20 h (±1.7 h) (not significantly different from the average daily inactivity of the oryx measured during the Saudi winter, Mann-Whitney, nonparametric test, P = 0.052, but significantly less than for the oryx measured during the Saudi summer, P = 6.04 × 10−6). The animal displayed polyphasic inactivity with an average of 16 (±4) bouts of inactivity per day. The average daily onset of the main inactive bout occurred between 0330 and 0400 h, with this bout having an average length of 131 min, just over 2 h (±56 min). Thus, during the South African summer recording period, the single oryx displayed a nocturnal inactivity pattern, with the majority (81.6% ± 12%) of inactivity occurring during the night (Fig. 3). The only noted difference between the South African animal and the Saudi Arabian winter animals was an increase in inactivity in the late afternoon in the South African animal compared with the Saudi Arabian winter animals (Fig. 1). This may reflect some effect of increased summer temperatures but does not match the increase in summer afternoon inactivity levels observed in the Saudi Arabian summer animals.
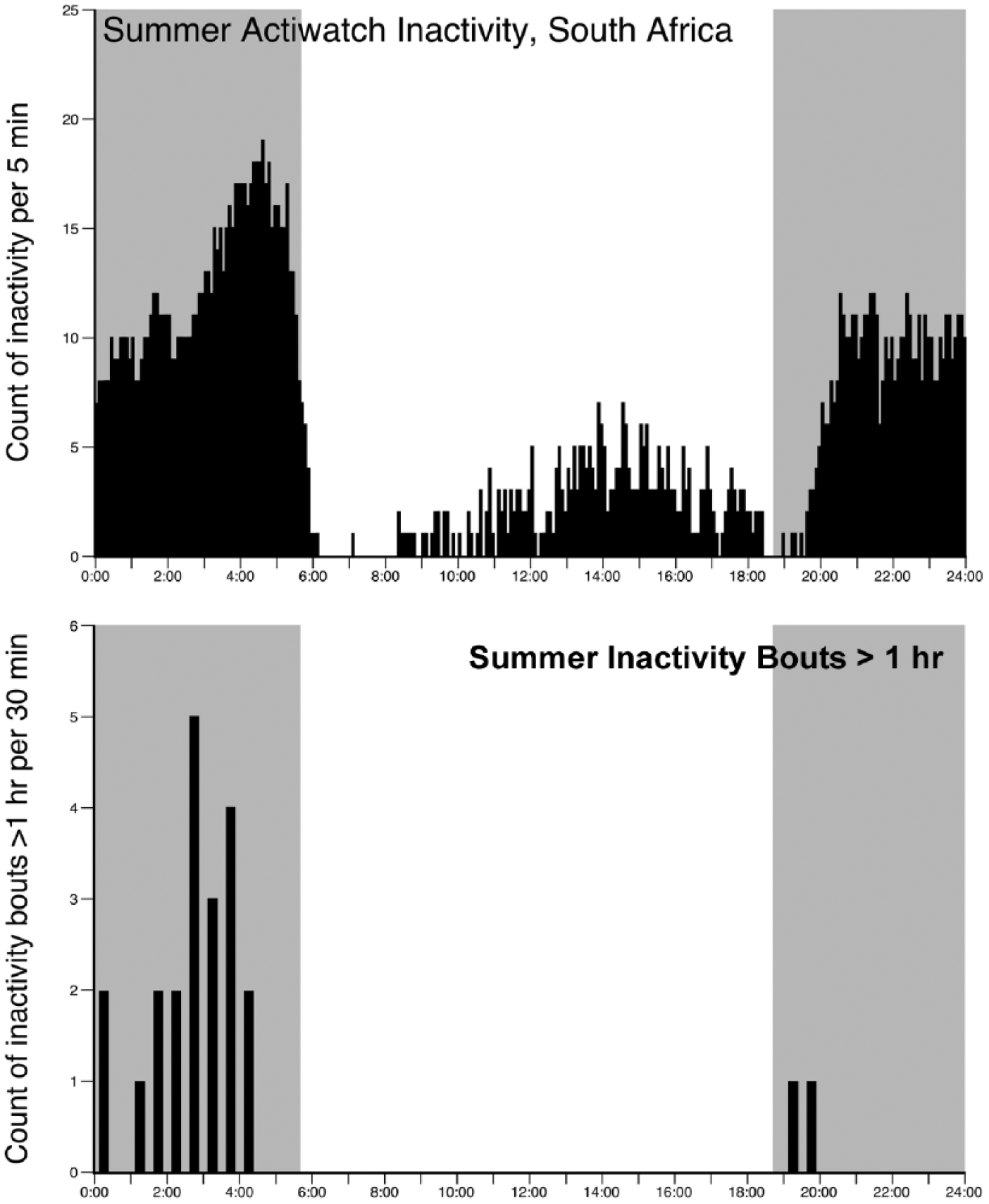
Upper graph illustrates the average count of inactivity for any given 5-min period scored over 28 days in the South African summer (February 2015) for a single male Arabian oryx. Note the nocturnal pattern of inactivity during this summer month, with relatively little inactivity occurring during the daytime. This contrasts with the recordings made in the summer month in Saudi Arabia (see Fig. 1). The lower graph shows the frequency distribution (in 30-min intervals) of inactive bouts of greater than 1 h duration during the South African summer recording period for the Arabian oryx measured. Note than 21 of the 23 (91.3%) bouts of inactivity greater than 1 h occurred in the period between midnight and dawn and that none occurred during the daytime, resembling closely the observations made on the Saudi Arabian oryx recorded in the winter.
Discussion
The current study reveals how the Arabian oryx manages daily inactivity patterns in relation to the dramatic seasonal conditions of a hot and inhospitable environment. First, we have observed that the Arabian oryx changes from nocturnal inactivity in the cooler months to a bimodal inactivity pattern during the hotter months. Second, the timing of the majority of inactivity also changes seasonally, from the coolest parts of the winter nights to the hottest parts of the summer days. Third, through comparison with a conspecific inhabiting a different environment, we suggest that ambient temperature is the dominant environmental condition driving these changes in daily inactivity patterns. Fourth, since the timing of the oryx body temperature rhythm did not show a seasonal change, the change in inactivity patterns likely represents a “masking” effect rather than a shift in the circadian clock’s endogenous oscillation.
The fine-grain actigraphy analysis revealed 2 different daily patterns of inactivity in the Arabian oryx, from nighttime inactivity in the cooler months to an additional daytime period of inactivity in the hotter months. Seddon and Ismail (2002) used focal 10-min observations from 0600 h to 1800 h and Hetem et al. (2012b) used a coarse grain method of actigraphy in their studies but were not able to clearly show this variation; this was because Seddon and Ismail’s observations did not include the full 24-h period and Hetem et al. did not make recordings throughout the entire year (the cooler months of December to March were not recorded). While many studies report seasonal variations of daily activity patterns, termed temporal niche switching (reviewed by Refinetti, 2008, and Hut et al., 2012) from a range of mammalian species, fewer examples under natural conditions have been reported. Of specific interest to the current study are those examples in which seasonal changes in ambient temperature appear to drive changes in activity patterns, for example, in the degu (Kenagy et al., 2002; Vivanco et al., 2010), blind mole rat (Oster et al., 2002), and bat-eared fox (Lourens and Nel, 1990). Under certain environmental conditions, daytime activity can be induced in the golden spiny mouse (Shkolnik, 1971) and Asiatic ibex (Fox et al., 1992). It is also possible that the dromedary camel, when it becomes an adaptive heterotherm under experimental dehydration conditions (El Allali et al., 2013), may show changes in daily activity patterns that correspond to the changes in the Arabian oryx under high ambient temperatures, but this has not yet been shown (El Allali et al., 2013; Bouaouda et al., 2014). Our analysis of the Arabian oryx in the South African summer suggests that temperature is the driving environmental factor behind the seasonal change in inactivity patterns, whereas day length and rainfall appear to have little or no role.
If we focus on the timing of the main bouts of inactivity (those greater than 1 h), many of which lasted for almost 2 h in both seasons in all the oryx studied in Saudi Arabia, it is reasonable to assume that during these periods of inactivity the oryx were likely to be asleep for much of the time. In this sense, during the winter/cooler months, the majority of sleep would have occurred between approximately 0300 and 0500 h (when ambient and body temperature are at their lowest), whereas during the summer/hotter months, the majority of sleep would have occurred in 2 periods between approximately 0300-0500 h and 1230-1430 h (the latter when ambient and body temperature are at their highest). The pattern of polyphasic inactivity in the Arabian oryx, with the main inactive bout occurring mostly during the night in the winter and an overall amount of inactivity being approximately 5 h, represents a typical inactive-sleep pattern in terms of timing, phasing, and duration for an artiodactyl of approximately 80 kg body mass (e.g., Campbell and Tobler, 1984; Siegel, 2005). In contrast, the timing of inactivity observed in the summer is unlike that observed in any artiodactyl studied to date, with a major inactive bout during the hottest part of the daytime and an increase of 1.5 h per day spent inactive. This contrasts with many other mammals, including humans, which are more inactive in the winter than the summer (e.g., Erriksson et al., 1981; Everts et al., 2004; Lariviere and Messier, 2009; Yetish et al., 2015). Because during slow-wave sleep, the body and brain temperature of all mammals studied to date decrease significantly (reviewed in Kräuchi and Deboer, 2010), we speculate that the daytime shift of inactivity-sleep in the Arabian oryx may be an adaptive response to increased ambient temperatures, taking advantage of the thermoregulatory physiology of sleep to lessen the impact of the summer heat.
Our observations in the Arabian oryx demonstrate a seasonal alteration in the phase relationship between inactivity and the body temperature rhythm. How this changing phase relationship is brought about by the neural systems involved in the control of behavioral and physiological rhythmicity is unknown. It is possible that ambient temperature might act on the downstream circuitry of circadian control, such as the ventral subparaventricular zone of the hypothalamus (Saper et al., 2005), and thus modify the timing of inactivity, and presumably sleep, although this remains to be investigated in more detail.
Footnotes
Acknowledgements
This project was funded by the National Plan for Science, Technology and Innovation (MAARIFAH), King Abdulaziz City for Science and Technology, Kingdom of Saudi Arabia, Award Number 11-ENV1918-02 (A.N.A., O.B.M., N.C.B. and P.R.M.). The authors are extremely grateful to Prince Bander bin Saud Al-Saud, President of the Saudi Wildlife Authority (SWA), for his unlimited and enthusiastic support to undertake these studies on the Arabian oryx managed by the SWA. We are also grateful for the tremendous help provided by our colleagues at the Prince Saud Alfaisal Research Center in Taif, Saudi Arabia, especially Mr. Ahmed Boug, Dr. Saud Anagariyah, Mr. Ghazi Raziman, Mr. Raed Aljuaid, and Mr. Ali Aljuaid. We also extend our sincerest thanks to Mr. Jan Pickard at Ratelfontein, South Africa, who allowed us to use one of his Arabian oryx in South Africa.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.