
Abstract
In mammals, the pineal hormone melatonin is thought to be essential to process environmental photoperiodic information. In this study, we demonstrate in a circannual species, the European hamster Cricetus cricetus, the existence of a melatonin-independent second pathway. In 4 physiological parameters (reproduction, body weight, activity pattern, body temperature), a large majority of pinealectomized European hamsters were entrained to an accelerated photoperiodic regime. It compressed the natural variations in the photoperiod to a 6-month cycle, which allowed us to record up to 6 complete physiological cycles during the life span of the individuals. We show further that whether a pinealectomized animal is able to entrain to changes in the photoperiod is influenced by the season of pinealectomy. The results do not disprove that melatonin is capable of entraining a circannual rhythm, but they show clearly that melatonin is not necessary, demonstrating another melatonin-independent pathway for circannual entrainment by the photoperiod. In view of these new insights, a revision of the original literature revealed that probably the melatonin-independent pathway plays an important role in most circannual mammals but only a minor role in photoperiodic species. Thus, the present work provides also the first evidence for different synchronization mechanisms in photoperiodic and circannual species.
Keywords
Seasonal variations in the environment cause high physiological demands for organisms. Animal species that live in such environments need precise timing to ensure that their offspring are born at the most favorable seasons during spring and summer and to prepare for the harsh conditions of winter. Changes in the photoperiod (PP) are the most reliable markers for the time of year (Gorman et al., 2001). Basically, 2 strategies have evolved to keep the physiology in synchrony with the annual cycle: 1) seasonal rhythms that are directly controlled by the PP or 2) endogenously generated circannual rhythms that are entrained by the PP (Zucker, 2001).
In the former strategy, the PP directly drives the seasonal changes in physiology. For example, in Syrian hamsters (Mesocricetus auratus) or Siberian hamsters (Phodopus sungorus), a short PP (SP) induces gonadal atrophy, and animals become sexually active as soon as a long PP (LP) is re-established (Gaston and Menaker, 1967; Elliott, 1981; Hoffmann, 1982). However, when they are kept in a constant SP, an as yet unknown endogenous process (referred to as photorefractoriness) (Reiter, 1972; Stetson et al., 1976) induces sexual reactivation after 24 to 28 weeks, and the gonads will remain active for the lifetime of the hamsters unless photorefractoriness is broken by a new exposure to a long PP. In short, these photoperiodic species are unable to pass a complete annual cycle without changes in the PP (Reiter, 1972; Hoffmann, 1981).
Animals that use the circannual strategy express repetitive, complete seasonal oscillations in their physiology, with a period of approximately 1 year when they are kept for several years in constant environmental conditions (Pengelley, 1974; Gwinner, 1986; Boissin and Canguilhem, 1988). The seasonal changes in physiology are endogenously driven by a circannual clock. To keep such a circannual clock synchronized with the environmental period length of 1 year, the photoperiodic signal is not required during the whole year but only during certain time spans (Randall et al., 1998; Miyazaki et al., 2005; Monecke et al., 2009).
The European hamster (Cricetus cricetus) shows circannual rhythms (Masson-Pévet et al., 1994), which modulate at least 26 physiological parameters (for review, see Monecke et al. [In press]). Changes in the PP entrain the circannual clock during 2 phases of sensitivity around the solstices (Saboureau et al., 1999; Monecke and Wollnik, 2004). With transfers to artificial short PPs, it has been shown that the most important one is the so-called sensitive phase to short PPs from mid-May to mid-July (Saboureau et al., 1999), during which principal resetting of the circannual clock occurs (Monecke et al., 2009). During the sensitive phase around the winter solstice, the PP is also effective in entraining the circannual clock but to a minor extent (Monecke et al., 2009; Monecke et al., 2010). Both phases start anticipatory long before, in natural conditions, the resetting photoperiodic signal (shortening of the PP after the longest day or lengthening of the PP after the shortest day) is expected.
The nocturnal melatonin peak is the crucial element of the photoperiodic entrainment pathway of seasonal rhythms, since its duration varies proportionately with night length (Bartness and Goldman, 1989; Pévet et al., 1991; Steinlechner, 1992). Removal of the pineal gland severely impairs the perception of photoperiodic information, and virtually all reviews on melatonin conclude that pinealectomized mammals are not able to detect and process photoperiodic information (Goldman and Nelson, 1993; Reiter, 1993; Arendt, 1995; Malpaux et al., 2001). Furthermore, in pinealectomized mammals, short- or long-duration melatonin infusions induce summer or winter physiology, respectively (Goldman et al., 1981; Bittman et al., 1983; Bartness et al., 1993), or entrain circannual rhythms (Woodfill et al., 1994).
European hamsters show extremely strong seasonal variations not only in the duration but also in the amplitude of nightly melatonin production: Around the summer solstice (Vivien-Roels et al., 1992), or in an artificial long PP (Vivien-Roels et al., 1997), nightly melatonin levels barely exceed daytime values. A nightly melatonin peak reappears when the natural PP falls below 15.5 h of day length (critical PP) (Canguilhem et al., 1988) in mid-July when gonadal regression is initiated (Saboureau et al., 1999) and an abrupt change in the activity pattern observed (Wollnik et al., 1991; Monecke and Wollnik, 2005; Monecke et al., 2006). Originally, it was assumed that this reappearing melatonin peak then transduces the inhibiting short photoperiodic message to the reproductive axis. However, it is known from Siberian hamsters that after a transfer from a long PP to a short PP, the first increase in the nocturnal melatonin peak is seen only after some days and that several weeks are needed to see its full extension (Illnerová et al., 1984). Thus, melatonin changes do not appear to be instrumental in the rapid response to a short PP, which is observed, for example, in the abrupt change in the activity pattern of European hamsters.
In European hamsters, 3 findings raised doubts that melatonin is the only internal transducer of the photoperiodic message: 1) The lack of circadian melatonin production which is endogenously driven (Vivien-Roels et al., 1997) coincides with the phase of sensitivity to short PPs (Monecke and Wollnik, 2005), and it is endogenously driven (Vivien-Roels et al., 1997). The melatonin rhythm seems absent precisely during the phase when the PP is measured and a short PP is able to synchronize the circannual clock (Monecke et al., 2009). 2) Furthermore, during the opposing sensitive phase around the winter solstice, that is, also during a season when the PP is measured, melatonin receptors are profoundly reduced in the pars tuberalis, which is thought to be essential in the translation of the photoperiodic message (Dardente, 2012). mRNA expression levels of the melatonin receptor MT1 are then close to zero (Dardente et al., 2003), and the number of melatonin binding sites is profoundly reduced (Skene et al., 1993). These observations suggest that European hamsters might be able to respond normally to photoperiodic changes in the absence of a melatonin rhythm. 3) Further support for this hypothesis is derived from data obtained on pinealectomized animals in a constant PP: properties of their circannual oscillations in reproduction differ greatly between animals in a constant long PP and constant short PP but not between intact and pinealectomized animals in the same PP (Masson-Pévet et al., 1994).
The aim of the present work was to demonstrate that European hamsters are able to entrain to photoperiodic changes even in the absence of pineal melatonin. We thus studied the effect of photoperiodic change on the entrainment capacities of pinealectomized and sham-operated European hamsters. An accelerated photoperiodic regime was applied, which mimics natural changes in the PP but compresses them to 6 months (182 days), to which European hamsters entrain without problems (Canguilhem, 1989). This zeitgeber regime was chosen because 1) it differs clearly from the endogenous circannual period length, 2) it overcomes problems due to the short life span of the hamster (maximum 2-3 years), and 3) it allows for following a larger number of cycles. To ascertain whether only particular functions or the whole physiology of an animal was entrained to the accelerated PP, we simultaneously recorded the reproductive state, body weight, daily activity rhythm, and body temperature, which are known to be expressed seasonally.
In the first experiment (2007) (Table 1), European hamsters were pinealectomized or sham operated at an age of 9 months around the maximal PP (LDmax). We chose that PP since in these conditions, a nightly elevation of melatonin is lacking either under natural or experimental conditions (Vivien-Roels et al., 1998; Monecke and Wollnik, 2005).
Experimental conditions
In the second experimental series (2009-1 to 2009-3) (Table 1), we investigated whether the age and/or the season at pinealectomy affects the ability to entrain to photoperiodic changes. Animals were split into 3 groups. Surgery was performed at 3 ages (3, 6, and 9 months) and 2 seasons (LDmax and minimal PP [LDmin]) (Table 1).
In the third experiment (2010) (Table 1), we checked the presence of plasma melatonin in pinealectomized animals. The experimental conditions were the same as for the 2007 group. Sampling seasons were shortly after the LDmin when the duration of the light phase of the accelerated PP was 9 h 26 min (SP) and shortly after the LDmax when the light phase lasted 14 h 16 min (LP).
Materials and Methods
Maintenance
A total of 132 male European hamsters were used. They were born in March or April in our breeding colony (Chronobiotron, CNRS-UMS3415) and raised in LD16:08, with continuous dim red light (<3.5 lux) during the night phase. They were transferred to the accelerated PP (6-month cycles) in early July, just when it corresponded to LD16:08 and was starting to decrease. The light phase in the accelerated PP ranged between 16 h 10 min (LDmax) and 8 h 15 min (LDmin), mimicking the natural range of a PP at 48°N around Strasbourg, and in the dark phase, dim red light was provided. The timer was adapted accordingly once per week during the light phase. Weekly photoperiodic steps ranged between 8 and 50 min. In all protocols, the animals received standard laboratory food (Safe005 diet for rodents, Safe, Augy, France) and water ad libitum and were individually maintained in type 3 Makrolon cages (Bioscape, Emmendingen, Germany) at a constant temperature (20 °C ± 2 °C) and humidity (55% ± 5%).
All experiments were performed in accordance with the National Institutes of Health guidelines for the care and use of animals in experimental procedures, with the European Communities Council Directive of November 24, 1986 (86⁄609⁄EEC) and with French laws.
Pinealectomy
For pinealectomy or sham surgery, the animals were anesthetized with an intraperitoneal injection of 0.2 mL/100 g body weight of a mixture of 4 volumes of Zoletil 20 (Virbac, Carros, France) and 1 volume of Rompun (Bayer HealthCare, Monheim, Germany). The pinealectomy was performed as described previously (Hoffman and Reiter, 1965). Briefly, after cutting out a circular bone disc around the lambda, the superior sagittal sinus below was perforated with pointed forceps and the pineal gland directly underneath seized and removed. A vacuum pump allowed surgery in spite of bleeding, which was stopped with gel foam (Bloxang, Bausch & Lomb, Montpellier, France). The bone disc was then returned to its place. For sham surgery, the circular bone disc was removed and replaced. Two to 4 mL of warm Ringer solution was injected to replace blood loss.
To verify the absence of melatonin in pinealectomized animals, blood samples were taken in dim red light in a short isoflurane flush shortly after the LDmin or LDmax at 3 circadian times from the retrobulbar venous plexus in each individual of the 2010 group. Since a volume of 1 to 1.5 mL of blood was needed for the analysis of melatonin in the plasma, the 3 sampling times were separated for each individual by 77 to 108 h to allow recovery between samplings. Blood was sampled by means of microcapillaries. Plasma was obtained via heparin. It was stored at −20 °C until analysis. Plasma melatonin was measured by radioimmunoassay (RIA) following the protocol of Vivien-Roels et al. (1992). Briefly, we used a rabbit antiserum (R19540, INRA, Nouzilly, France) to label 2-[125I]iodomelatonin, previously verified for European hamsters (Pévet et al., 1989). The minimum detection level of the assay was between 5 and 10 pg/mL of plasma. At the levels of 20, 100, and 200 pg/mL, the interassay coefficients were 4%, 8%, and 12%, and the intra-assay coefficients were 4%, 3%, and 3%, respectively (Vivien-Roels et al., 1992).
Markers Used to Follow the Circannual Rhythm
Reproductive state and body weight
Body weight and reproductive state were checked biweekly in a short isoflurane flush. Males were considered reproductive when the testis size exceeded the length of 1.8 cm, since testosterone levels are then maximal (Buijs et al., 1986; Masson-Pévet et al., 1994). Sexual quiescence was defined as the time when testes were no longer visible and ascended into the abdomen. As a seasonal marker for body weight, the maximum increase during a cycle was used (Fig. 1).
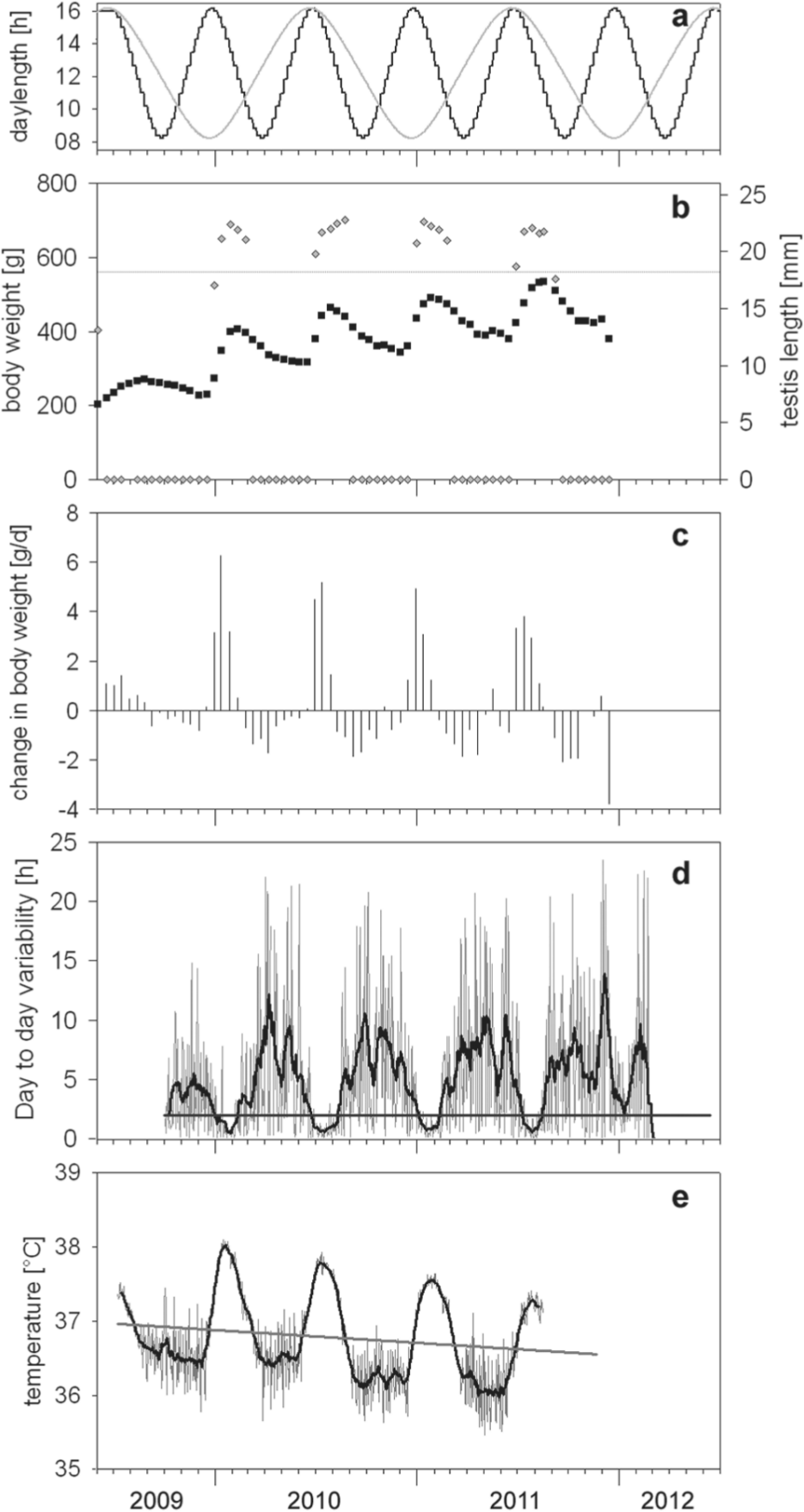
Reference points in seasonal physiological changes in 3 representative sham-operated European hamsters kept in an accelerated PP. (A) Accelerated photoperiodic schedule (black) and, for comparison, natural changes in the PP (gray) at 48°40′N. (B) Body weight (black squares) and testis length (gray diamonds). Onset or offset of the reproductive phase is defined as the crossing of the testis length and the threshold of 1.8 cm (horizontal line). (C) The maximum increase in body weight during one cycle marks the reference point for body weight (same animal as in B). (D) For the parameter activity, the crossings of the 15-day running average (black line) of the day-to-day variability in the activity onset (gray line) with a threshold set at 2 h (horizontal line) were defined as onsets or offsets of the precise activity pattern of summer. (E) For body temperature, it was considered as a winter pattern when the 15-day running average (black line) of the daily means in euthermic body temperature (mean of all values ≥34 °C, gray line) was below the regression line (gray) through the whole data set and as a summer pattern when it was above.
Activity and temperature
Locomotor activity was recorded using passive infrared (IR) captors mounted on the top of the cages. Data acquisition was performed with CAMS (Howard Cooper, Lyon, France). Further analysis was performed by ClockLab (ActiMetrics, Evanston, IL) and Microsoft Excel (Redmond, WA). We chose the day-to-day variations (absolute values) of the activity onsets as a seasonal marker (Fig. 1), since in the course of the year, its precision varies considerably (Wollnik et al., 1991; Monecke and Wollnik, 2005). The intersection of the 15-day running average and a threshold set at a variability of 2 h are used as reference points for the beginning and end of the precise (summer) phases.
Body temperature was measured in approximately 50% of animals by implanted iButtons (DS1922L, Thermochron iButtons, Maxim, Dallas, TX) (for further details, see Davidson et al. [2003]) embedded in a paraffin/Elvax coating (Respironics/Mini Mitter, Bend, OR). For each use, functionality was pretested in a 37 °C water bath. Temperatures ranged between 36.631 °C to 37.116 °C. Two iButtons were implanted simultaneously in the abdominal cavity under isoflurane anesthesia to allow recording at 20-min intervals for 3 months when iButtons were replaced. Data were read out by the software OneWireViewer (Maxim) and were processed by Microsoft Excel and Sigma Plot (SPSS ASC GmbH, Erkrath, Germany). As a marker, the alternating phases of euthermy and heterothermy (Wollnik and Schmidt, 1995) were taken. To make this method also applicable for animals that show only a few torpor bouts during winter, only euthermic values (20-min values of ≥34 °C) were considered for daily means. The crossings of the 15-day running average with the linear regression through the whole recording time were defined as changes between the summer and winter state of the animals (Fig. 1).
Data Analysis
An animal was considered to be completely entrained to the photoperiodic cycle when the number and timing of cycles in reproduction, body weight, activity, and body temperature corresponded to the number and timing of zeitgeber cycles. Additionally, in all 4 parameters, the period length tau between subsequent reference points should correspond to the zeitgeber period. These reference points were maximal increases in body weight and the onsets and offsets of periods characterized by reproduction, sexual quiescence, precise activity pattern, and elevated euthermic temperature. The means of these parameters were summarized to the overall tau for an animal. Statistics were performed by multifactorial ANOVA followed by least significant difference (LSD) post hoc comparison.
Results
Verification of Pinealectomy and Melatonin Absence
The success of surgery was checked twice for each animal 1) by histological investigation of the removed tissue and 2) by post mortem inspection of the cranial cavity. All experimental animals were successfully pinealectomized. However, in 2 sham-operated animals, the pineal gland was accidentally removed. They were thus assigned to the respective pinealectomized group.
Blood was sampled only in a separate group (2010 group), since the high blood loss (1.5 mL at each sampling point) might have interfered with seasonal entrainment. A nocturnal melatonin peak was observed only in sham-operated SP animals (Fig. 2). In 16 pinealectomized animals, no melatonin was detectable, whether they were able to entrain to the accelerated PP or not. Minimal amounts of melatonin could be detected only in 2 pinealectomized animals at one time point each, but this was neither during late night (the peak time in controls) nor exceeded these values the LP night-time levels of the control group. Moreover, they did not show seasonal variations. The multifactorial ANOVA revealed significant effects for the factors treatment (p < 0.0001), PP (p < 0.0001), and time (p < 0.05) as well as for all possible interactions (p < 0.05). The SP midnight (p < 0.05) and morning (p < 0.001) values of the control group differed significantly from all other values (LSD test of the interaction group*PP*time).
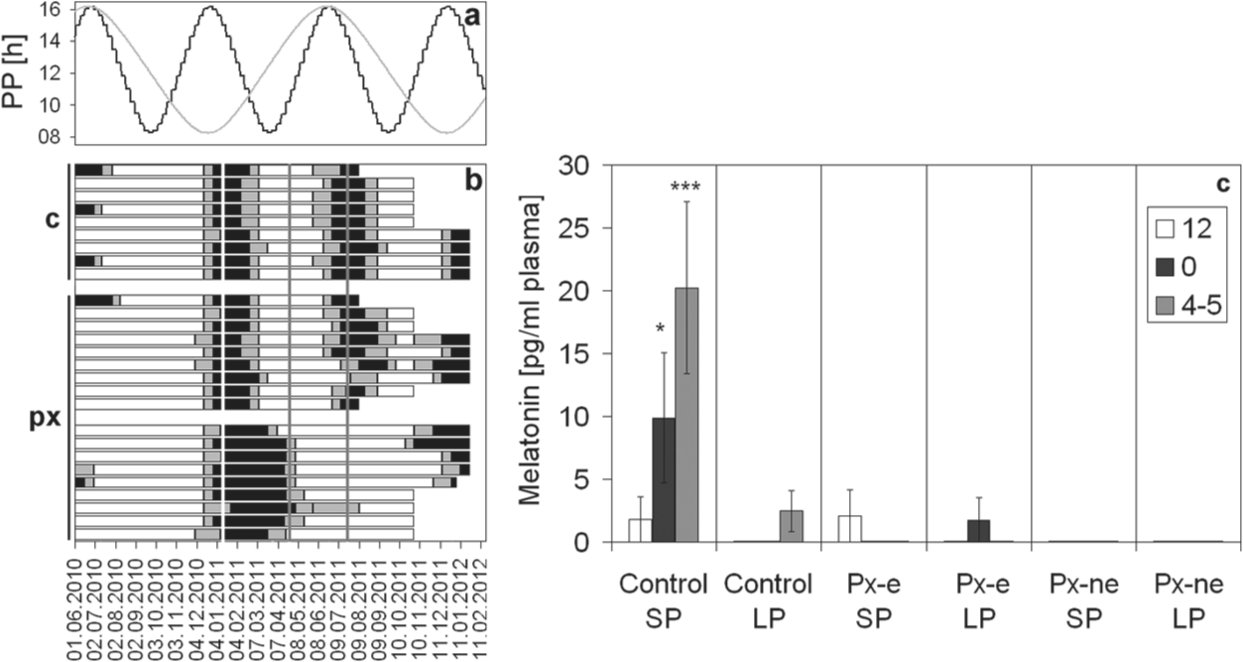
(A) Seasonal changes in plasma melatonin of male European hamsters kept in an accelerated PP (A) and pinealectomized (px) or sham operated (c) at an age of 9 months. (B) Reproductive state of individuals represented by horizontal bars, which are black when the animals are reproductive and white when gonads are regressed. Gray stands for the transition. Vertical lines indicate the time of surgery (white) or time of the 2 blood samplings (gray) shortly after the subsequent minimal and maximal PPs (SP and LP, respectively). (C) The mean (±SEM) plasma melatonin concentration at 3 different sampling times: white: midday; black: midnight; gray: 1 h before lights-on. One asterisk indicates a significant difference from all other values (p < 0.05, LSD test); 3 asterisks indicate a highly significant difference (p < 0.0001, LSD test) to all values without asterisks. Note that zero was assigned to samples in which melatonin could not be detected. Px-e and Px-ne stands for entrained or not entrained pinealectomized animals, respectively.
Do Pinealectomized European Hamsters Entrain to an Accelerated PP?
All control animals of the 2007 group entrained to the accelerated PP (Fig. 3). The ratio of the duration of reproductive competence and quiescence was similar to that observed in natural conditions. For each photoperiodic cycle, the animals showed one cycle in physiology in all 4 parameters. Reproductive phases started shortly before the LDmax was reached, and the increase in body weight peaked at its beginning. The precise phase in the activity pattern also started around the LDmax as well as the phase in which the animals showed the high temperatures of summer physiology. The mean circannual period length tau of the controls was 182.3 ± 3.0 days (range, 165.4-193.4 days) and reflects the length of the zeitgeber period of 182 days.
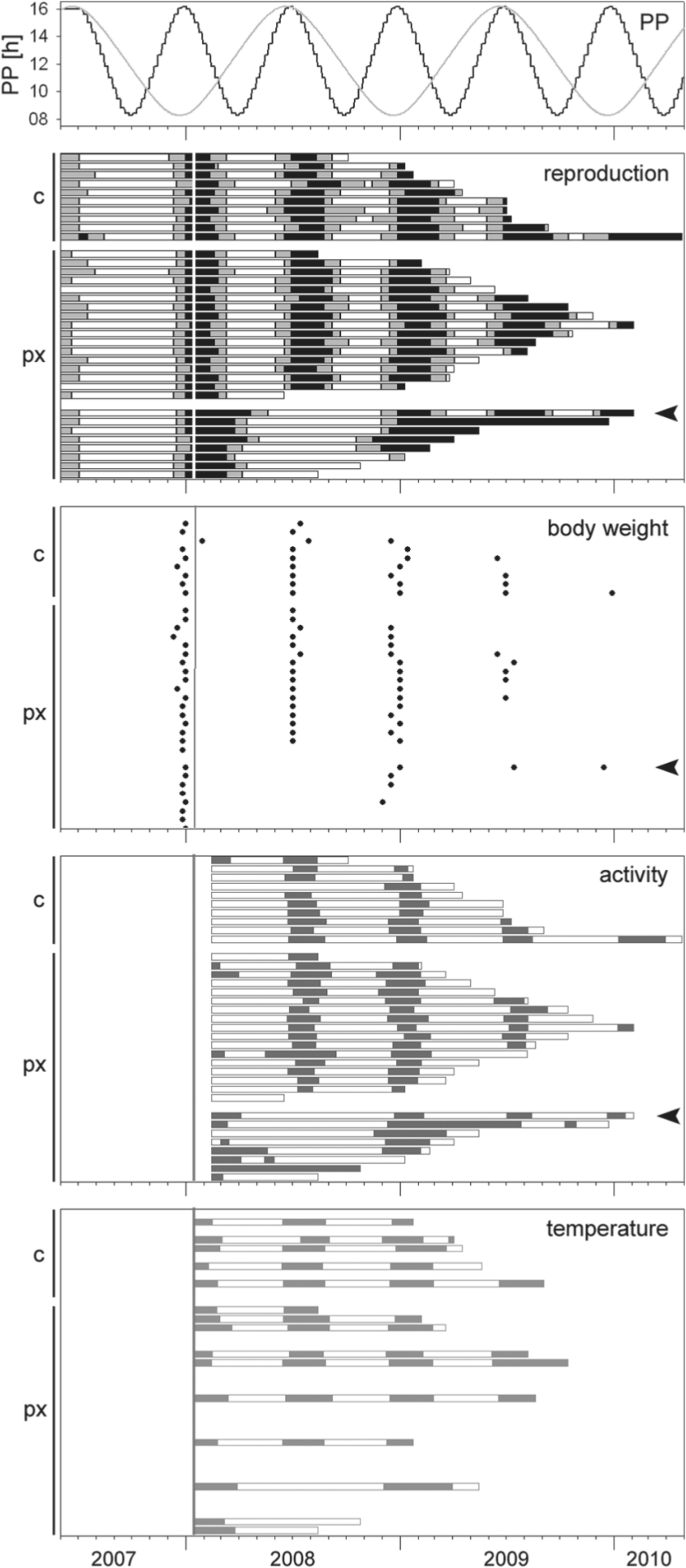
Seasonal changes in physiological parameters of pinealectomized (px) or sham-operated (c) male European hamsters kept in an accelerated PP. Individual animals are presented by horizontal bars or a horizontal succession of circles. Note that in each panel, the individuals are listed in the same order and further that this order differs from the random order in the animal facility. Filled bars indicate the summer physiology (reproductive, precise activity pattern, high euthermic body temperature), and open bars indicate the winter pattern (sexual quiescence, imprecise activity pattern, low euthermic body temperature). Gray bars in the reproduction panel indicate additionally the transition between the reproductive state and sexual quiescence. Black circles in the body weight panel indicate the date of the maximum increase in body weight during one cycle, and the vertical line indicates the date of surgery. The black arrowheads point to a re-entraining animal.
The majority of pinealectomized animals of the 2007 group also entrained to the accelerated PPs (Fig. 3). In up to 4 complete cycles after pinealectomy, 17 of 25 animals showed the same timing as the controls in all measured parameters. Their mean tau was 180.3 ± 1.4 days (range, 170.5-189.1 days) and did not differ from tau of controls (p = 0.751, multifactorial ANOVA, followed by LSD test) (Fig. 4). The remaining 8 animals initially regressed gonads later than the controls and were then out of phase with the photoperiodic cycle. These animals seemed to skip the next reproductive phase to become reproductive at about the same time as the controls start their third reproductive phase. Their mean tau was 328.2 ± 12.2 days and ranged from 305.5 to 358.8 days. The latter is close to the period length of 2 zeitgeber cycles. One animal that initially could not follow the 6-month photoperiodic cycle clearly re-entrained to the accelerated PP.
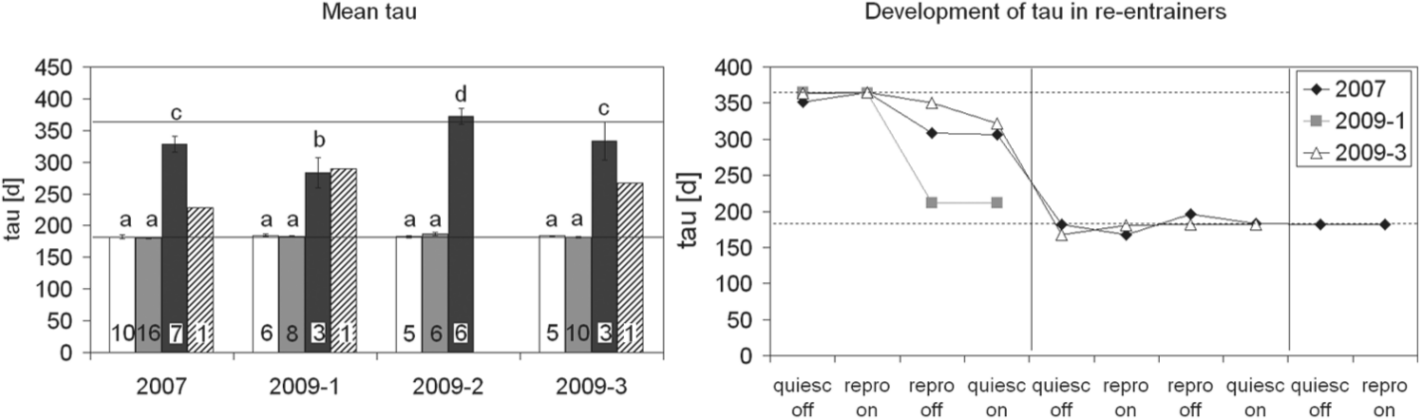
(Left) Mean circannual tau of the subgroups of each experiment: white: control animals; gray: entrained pinealectomized animals; dark gray: nonentrained pinealectomized animals; hatched: re-entrained pinealectomized animals. Letters indicate a significant difference to values labeled with other letters (multifactorial ANOVA, followed by LSD test, p < 0.001). Numbers at the bottom of the columns give the sample size. (Right) The time course for tau of the reproductive cycle in the 3 re-entrainers. They appear in the left panel with an intermediate value. Note that for each cycle (separated by vertical lines), 4 different values for tau are available, namely between onsets and offsets of the reproductive phase and sexual quiescence. Horizontal lines in both panels indicate 182 days or 364 days, that is, 1 or 2 zeitgeber periods.
Does the Timing of Pinealectomy Have an Effect?
In all 3 groups of the 2009 experiment, control animals entrained to the accelerated PP in all measured parameters (Fig. 5). The entrainment rate of animals that were pinealectomized around the LDmax was similar, independent of whether surgery was performed at an age of 3 or 9 months (Table 2).
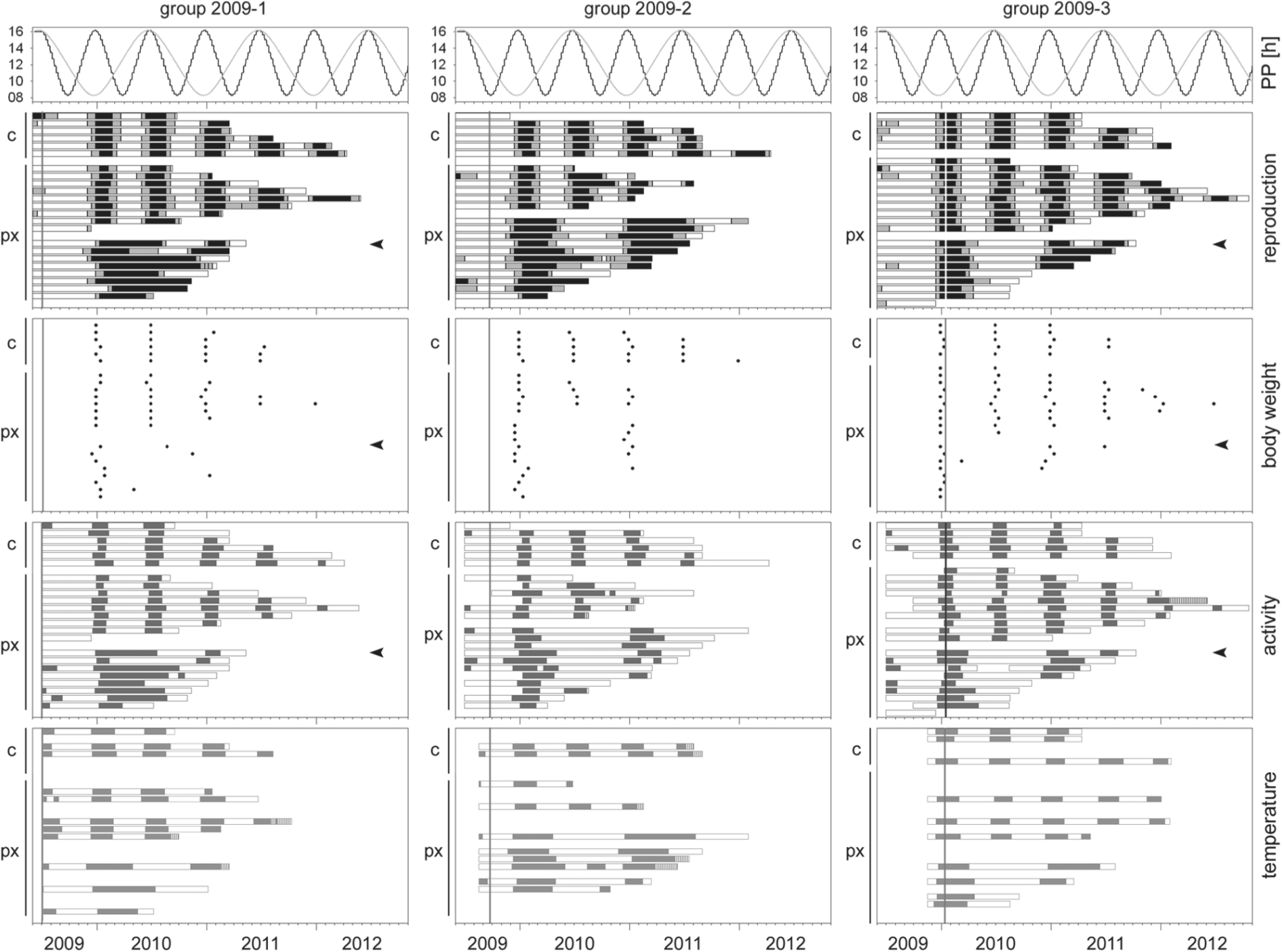
Seasonal changes in physiological parameters of male European hamsters pinealectomized (px) or sham operated (c) at an age of 3 (left column), 6 (middle column), or 9 months (right column) around the summer solstice or winter solstice of the accelerated PP (upper panels). Vertical lines indicate the day of surgery. Hatched ends of the bars indicate uncertainties at the end of the animals’ life span. See Figures 2 and 3 for further details.
Percentage of pinealectomized entrainers or re-entrainers to the accelerated photoperiodic regime
Pinealectomies were also performed at different seasons. Clear entrainment was observed in 56% to 72% of the animals when they were pinealectomized around the LDmax. In contrast, only 35% of the animals pinealectomized around the LDmin could follow the accelerated PP (Table 2). The majority of nonentrainers of this group (2009-2) was clearly phase advanced with the onset of their first physiological summer patterns after pinealectomy (Fig. 5), compared with the controls or the pinealectomized entrainers. It occurred a mean 12 days earlier in nonentrainers than in controls, which corresponds to 24 angular degrees in the complete cycle of 182 days.
In all groups, tau of the control animals and of the entrained pinealectomized animals matched the 182-day period length of the experimental cycle (Fig. 4). The mean tau of the nonentrained pinealectomized animals varied strongly between groups. It was always clearly longer than 182 days but mostly shorter than 364 days. It should be pointed out that all nonentrainers that lived long enough increased tau between cycles towards 364 days. All re-entrainers initially showed a tau close to 364 days and switched then to a 182-day cycle. The multifactorial ANOVA revealed a significant effect of subgroups (controls, entrainers, and nonentrainers; F2,70 = 512.02; p < 0.0001) due to the different ratio of entrainers (Table 2) in the start year (F3,70 = 9.7841; p < 0.0001) and the interaction of both (F6,70 = 8.0021; p < 0.0001). While there were, in no cases, any differences in tau between controls and entrainers (p > 0.5, LSD test) (Fig. 4), nonentrainers had a significantly higher tau than entrainers or controls of all groups (p < 0.0001, LSD test).
Discussion
Main Results
The present work clearly demonstrates in repeated experiments that European hamsters are able to entrain to changes in the PP even in the absence of the pineal hormone melatonin. Among the animals that were pinealectomized around the LDmax, the percentage of animals that entrained to the 6-month cycle was at least 56% to 72%. The histological and post mortem verification of pinealectomy as well as the final control of plasma melatonin excluded the possibility that our results could be a consequence of failed pinealectomy. Moreover, control animals and “entrainers” showed a more negative seasonal phase angle to the photoperiodic cycle than animals in a natural PP. While the former became reproductive around the LDmax, the latter become reproductive already at an intermediate PP in early April (Monecke and Wollnik, 2005). This altered phase angle shows that in the present study, we observe true entrainment and not masking, since the phase angle is dependent on the relationship of the periods of the endogenous rhythm and the zeitgeber (Pittendrigh and Daan, 1976). This has also been shown for the European hamsters in different square wave T-cycles (Monecke et al., 2009). The present work thus demonstrates the existence of a melatonin-independent pathway that entrains the circannual clock.
However, since a RIA allows only a quantification of melatonin levels above the detection limit, we cannot exclude the possibility that minimal melatonin values beneath that limit are still present in the plasma after pinealectomy. Whether such potential minimal melatonin values show any daily or seasonal variations and whether they are of any relevance for the animal in terms of seasonal synchronization have never been described and might be doubted.
Quite a few of the “nonentrainers” seem not to free run but to entrain to a 1-year cycle, since the onset of their second reproductive phase under an accelerated PP is similar to that of the third reproductive phase in controls or entrainers. Furthermore, the mean taus of nonentrainers of the 2007, 2009-2, and 2009-3 groups came very close to 364 days (Fig. 4), which is the period length of 2 accelerated cycles. That a shorter mean tau was observed in the nonentrainers of the 2009-1 group might be due to their shorter life span, since tau of nonentrainers often increased with time towards 364 days. The less than 364-day values seen in the first cycle after pinealectomy may thus represent a transient value towards a new steady state of entrainment to 364 days.
That they nevertheless entrain to 2 photoperiodic cycles instead of one is explainable by phase-dependent photoperiodic resetting in circannual species (Randall et al., 1998; Miyazaki et al., 2005; Monecke et al., 2009). The animals concerned were out of phase with the environmental cycle after surgery. Subsequent changes in the PP probably matched an endogenous phase that did not permit large phase shifts to resynchronize to the 6-month regime but were sufficient to keep the animal entrained to 1 year. It is thus possible that we would have observed a higher percentage of entrainers if we had chosen an accelerated PP with a period length longer than 6 months.
That some animals were phase shifted after pinealectomy (the nonentrainers) and not others (the entrainers) might be explained by individual variations of the underlying circannual cycle and phase response curve. The lack in melatonin might have been interpreted as a long day signal, and depending on which endogenous phase was matched, the circannual cycle might have phase shifted the cycle.
Our results provide strong evidence that the majority of pinealectomized European hamsters entrain to a 6-month cycle, and they also suggest that most nonentrainers presumably entrain to a 12-month cycle. However, we could not find evidence that the remaining animals in an accelerated PP are unable to entrain. Most of them were phase shifted and needed to re-entrain to the photoperiodic cycle. This might take longer than is possible given the animal’s life span.
Differences between Circannual and Photoperiodic Species
Clearly, in the European hamster, a melatonin-independent pathway exists to communicate the photoperiodic message to the circannual clock or at least to the circannual functions studied. Considering these new insights, we reviewed the existing literature on the effect of pinealectomy on the entrainment capacities of mammalian species. Many authors have reported that pinealectomy prevents an appropriate photoperiodic response or entrainment. Nevertheless, closer examination of these works with regard to the present results reveals some of them in a different light. Indeed, we found strong support for our results in other circannual species.
A representative recording of a pinealectomized golden-mantled ground squirrel (Spermophilus lateralis) showed that body weight and reproductive state stayed entrained to a simulated natural PP over 4.5 years (Hiebert et al., 2000). Moreover, in superior cervical ganglionectomized minks (Mustela lutreola) kept in natural conditions, the timing of the progesterone peak and the prolactin increase barely differed from control animals (Martinet and Allain, 1985). When superior cervical ganglionectomy was performed in June, dormice (Glis glis) show entrainment in thyroid-stimulating hormone (TSH), testosterone, and luteinizing hormone (LH) cycles to a natural PP in the same way as controls (Jallageas et al., 1993). Furthermore, the annual progesterone peak in pinealectomized female wolves (Canis lupus) was entrained to the natural year and occurred each year in January/February exactly as in sham-operated controls (Asa et al., 1987). Finally, pinealectomized or superior ganglionectomized Soay rams also entrained well to natural conditions in the parameters of testis diameter, testosterone, and prolactin and less robustly in sexual skin flush and follicle stimulating hormone (FSH) (Lincoln et al., 1989). In short, these circannual species were clearly able to process seasonal information without melatonin. Since the animals were kept outdoors, the authors attributed the entrainment to zeitgebers other than the PP. Temperature (Canguilhem and Petrovic, 1974) or social cues (Wayne et al., 1989) might have played a role.
In the present work, neither social nor nonphotic zeitgebers were present. Entrainment by olfactory cues can also be excluded, since nonentrainers that were surrounded by entrainers hibernated when the latter were reproductive. Thus, our study delivers unequivocal proof that in pinealectomized European hamsters, entrainment is clearly achieved by photoperiodic cues. It is likely that this applies also to the above-mentioned circannual species, since none of the known potential nonphotoperiodic zeitgebers has sufficient year-to-year precision, which would explain the precise entrainment of pinealectomized wolves (Asa et al., 1987) or rams (Lincoln et al., 1989).
In contrast to circannual species, PP-induced seasonal adaptation is severely impaired in most photoperiodic species after pinealectomy. Even so, a thorough literature analysis revealed that they are not completely unresponsive to the PP, but the small reactions that are observed after a change in the PP seem to be physiologically irrelevant. Examples in this category include Syrian hamsters (Mesocricetus auratus) (Reiter, 1974, 1975) and white-footed mice (Peromyscus leucopus) (Petterborg et al., 1981; Johnston et al., 1982).
Although the present results demonstrate that the photoperiodic signal can be integrated independently of melatonin to entrain the circannual cycle, this does not mean that in circannual species, the melatonin-dependent signal is not effective. For example, pinealectomized sheep can be entrained by season-specific melatonin infusions (Woodfill et al., 1994). In European hamsters kept in an LP, constant melatonin infusions induced gonadal regression (Masson-Pévet et al., 1987). In fact, both mechanisms exist, but to which extent melatonin acts either on the central circannual clock mechanism or directly on the hypothalamic-pituitary-gonadal axis as in photoperiodic species is unclear at the present stage.
Our findings suggest a fundamental difference between photoperiodic and circannual entrainment. Photoperiodic response is predominantly dependent on the melatonin pathway, while the resetting of circannual clocks is largely melatonin independent. As Boissin and Canguilhem (1988) suggested, seasonal species generally use both photoperiodic and circannual components for entrainment, but the proportion in which they occur is species specific.
This is supported by findings in the Siberian hamster, which is well known as a photoperiodic species (Hoffmann, 1982), but in a small number, a circannual reproductive rhythm could be demonstrated (Anchordoquy and Lynch, 2000). Their photoperiodic responsiveness after pinealectomy was intermediate between typical photoperiodic and typical circannual species: in an SP, the testis size of pinealectomized animals was significantly lower than in an LP but still significantly larger than in controls (Brackmann and Hoffmann, 1977). Furthermore, breaking refractoriness of Siberian hamsters requires the detection of a PP; still, it can be broken in pinealectomized animals (Kauffman et al., 2003; Butler et al., 2010).
Nature of the Melatonin-Independent Pathway
This result raises the question of the nature of the melatonin-independent photoperiodic signal and of the identification of the nervous or neuroendocrine pathways involved. The perfect entrainment in 4 different physiological parameters showed that this melatonin-independent signal can directly entrain the central circannual clock. In sheep, its location is assumed to be the pars tuberalis/pituitary gland (Lincoln et al., 2006; Dupre and Loudon, 2007; Lincoln and Hazlerigg, 2010; Hazlerigg and Lincoln, 2011). In European hamsters, it is not yet identified, but we know that it is neither located in the pineal gland (Masson-Pévet et al., 1994) nor in the olfactory bulb or suprachiasmatic nucleus (SCN) (unpublished results).
Notwithstanding, strong evidence supports the idea that the SCN is part of this melatonin-independent pathway, leading towards the circannual clock. For both the melatonin-dependent and -independent pathway, the SCN is the site where the photoperiodic message is generated. Moreover, during the sensitive phase to an SP around the summer solstice, the SCN-driven activity pattern changes profoundly in the European hamster and becomes very precise (Monecke and Wollnik, 2005). When the PP then decreases below the critical value of LD15.5:8.5 (Canguilhem et al., 1988), gonadal regression is initiated on the day on which the activity pattern reverts to the arrhythmic winter pattern but only after an advance in sunset/lights-off and not after a delay in sunrise/lights-on (Monecke et al., 2006). This shows a tight interaction of the circadian and circannual system for the melatonin-independent pathway.
After the SCN, there should be a specific pathway to transduce the melatonin-independent photoperiodic seasonal signal, analogous to the melatonin-dependent pathway. Research is in progress to identify it.
Conclusions
This work hence breaks down 2 dogmas: first, contrary to what was previously considered, we show that the melatonin pathway is not the only means by which photoperiodic information can be conveyed; and second, circannual and photoperiodic species show differences in the way that they process photoperiodic information for seasonal entrainment, which was not thought to be the case. While photoperiodic species rely predominantly (but not exclusively) on the melatonin-dependent pathway, circannual species rely predominantly (but also not exclusively) on a currently uncharacterized melatonin-independent pathway. Circannual resetting is thus more complex than was previously thought.
Footnotes
Acknowledgements
This study was supported by grants from the following organizations: German Research Foundation (DFG Wo 345/12-1 and Mo 1742/1-1), the Region of Alsace, the DREAL Alsace, the German Wildlife Foundation, and the CNRS. The authors thank Harald Feuchter, Daniel Bonn, and Olivier Arnaud for expert animal care; Michel Saboureau for his help in pinealectomies; Berthe Vivien-Roels, Beatrice Bothorel, and Christiane Calgary for the analysis of blood samples; Agnès Dégardin and Rim Bousalham for help in blood sampling; and Angeline Votteler for technical assistance. David Hicks corrected the English. The study was conducted at the Chronobiotron (CNRS-UMS 3415).
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.