
Abstract
The fruit fly Drosophila melanogaster has long served as a model system for circadian rhythm research. Various aspects of its genetic, molecular, and circuit-level properties are the subject of investigation, based on which several circadian behaviors and their neuronal controls have been unraveled. In an attempt to address the question of functional significance of circadian organization using a comparative approach, we studied activity/rest rhythm of wild-caught D. melanogaster (DM) and its close relative, Drosophila ananassae (DA). We compared features of the rhythm such as the ability to anticipate morning and evening transitions, presence or absence of morning-associated or evening-associated activity peaks, and phase of these peaks in both species. We found that these 2 sympatric species are different from each other in several aspects of activity/rest rhythm. Unlike DM, which showed a distinct bimodal activity pattern with both morning and evening peaks and a midday interval of relative inactivity under a 12:12-h light/dark regime, DA exhibited unimodal activity with a predominant morning peak, restricting most of its activity to the light phase with no apparent “siesta” during midday. While daytime sleep levels were not different between the 2 species, DA exhibited significantly lesser nighttime activity and higher, more consolidated sleep. This predominant morning activity of DA was also reflected in persistence and phasing of the morning peak under a range of photoperiods. Both under long and short days, the morning peak was the most dominant and persistent peak of DA, whereas the evening peak was more dominant in DM. In addition, DA had a significantly faster circadian clock and more consolidated activity compared with DM. Hence, we hypothesize that these recently diverged sympatric species of fruit flies occupy distinct temporal niches due to differences in their underlying circadian clocks and speculate that they occupy different spatial microenvironments in the wild.
Circadian clocks are believed to enable organisms to time their physiology and behavior in a manner that is most adaptively advantageous to them. It is plausible therefore to assume that timing of various behaviors is phased to enable organisms to gain maximum fitness benefit with minimum risk. For example, in Drosophila melanogaster (DM), the timing of emergence of adults from pupae under natural conditions coincides with dawn, which is also the time of highest humidity and lowest temperature (De et al., 2012). This is believed to enable flies to expand their wings upon emergence (Saunders, 2002). While emergence is predominantly unimodal in DM, activity/rest rhythm shows distinct bimodality with a large proportion of activity occurring within peaks coinciding with lights-on and lights-off under symmetric 12:12-h laboratory light/dark (LD) cycles (Saunders, 2002). Such a preponderance of activity during twilight hours is thought to enable flies to escape harsh conditions during the middle of the day, although there is no empirical evidence for the same (Pittendrigh, 1993). Another line of thought has been that bimodality in behavior is simply a reflection of the nature of circadian timing systems, which are composed of 2 oscillators, one of which is coupled to dawn and the other to dusk, and show differential sensitivity to light and temperature (Helfrich-Förster, 2009). In recent years, many studies on DM have tried to examine the neuronal correlates of the dual-oscillator organization and its so-called “morning” and “evening” oscillator cells (Grima et al., 2004; Stoleru et al., 2004; Rieger et al., 2006). However, it is increasingly becoming evident that such bimodality in behavior is governed by a plastic network comprising a large number of neurons (reviewed in Sheeba et al., 2008) and is largely dependent on photoperiod, light intensity, and temperature (Rieger et al., 2003; Miyasako et al., 2007; Yoshii et al., 2009; Zhang et al., 2010). An even more recent study revealed that 2 strains of DM from northern and southern latitudes show differences in their ability to entrain to long photoperiods and that this behavior is dependent on temperature and twilight conditions (Rieger et al., 2012). Furthermore, under natural conditions, there appear to be 3 rather than 2 peaks of activity in DM (Vanin et al., 2012).
Even though several insect species have been used to study circadian rhythms (reviewed in Helfrich-Förster et al., 1998), DM have provided the highest traction on several aspects such as the genetic, molecular, and cellular bases of circadian behavior due to their genetic amenability. We reason that we can gain greater insight into the functional significance of rhythmic behaviors if we compared DM with another closely related sympatric species, Drosophila ananassae (DA). Both DM and DA are cosmopolitan species and belong to the species group melanogaster (subgenus Sophophora) and show ~83% similarity in their genomes (Drosophila 12 Genome Consortium, 2007). DA is thought to have originated in Southeast Asia (Tobari, 1993), later having invaded more temperate regions (Dobzhansky and Dreyfus, 1943; Das et al., 2004), and now both species are sympatrically distributed in several tropical regions. DA are rarely found in more temperate latitudes, whereas DM are also distributed widely across temperate regions.
A recent study comparing these 2 species has demonstrated divergence in courtship behavior and its neuronal basis that may have contributed to their reproductive isolation (Riabinina et al., 2011). Yet another study reveals that synaptic structure at the neuromuscular junction of DM and DA has significantly diverged from each other to extents that are not explained by their phylogenetic relationships (Campbell and Ganetzky, 2012). Although circadian behaviors of both DA and DM have been previously studied, no systematic comparisons between the two, providing insights into their circadian organization, have been carried out until very recently (Hermann et al., 2012). This report, which appeared while our manuscript was under review, found a high degree of similarity in the neuroanatomy of circadian neurons in the brains of DA and DM in terms of cell number and expression of core circadian proteins such as VRILLE (VRI) and Par Domain Protein 1 (PDP1) and an important circadian neuropeptide, Pigment Dispersing Factor (PDF). The protein sequence identity of circadian photoreceptor CRYPTOCHROME and neuropeptide Ion Transport Peptide (ITP) was also found to be as high as 86.9% and 98.7%, respectively (Hermann et al., 2012), while there was no difference in PDF sequence between the 2 species.
Our studies were focused on overt activity/rest rhythmic behavior, which is the outcome of the cellular and molecular machinery of the circadian clock organization, under a wide range of photoperiods, in the 2 species DM and DA. We found that while DM displayed the expected bimodal activity pattern with peaks around dawn and dusk, DA were predominantly day-active, with maximum activity after lights-on and weak evening activity peak, suggesting that these 2 species show significant differences in their preference for timing of activity/rest behavior.
Materials and Methods
Fly strains
Both DM and DA were wild-caught from within Bangalore, India (12°58’N, 77°38’E), in the summer of 2005 using fruit-traps as bait and net sweeps. We tried to avoid some of the usual drawbacks of laboratory strains of flies, which are often highly inbred, by starting with founder populations that were moderately sized (DM, n = 70; DA, n = 100) and subsequently avoided populational bottlenecks and therefore genetic drift by maintaining them as large random mating populations of ~1200 individual flies (with roughly 1:1 sex ratio) on a discrete-generation stock maintenance cycle of 21 days. These flies were maintained on standard cornmeal medium under LD 12:12 (~1.5 W/m2) conditions at ~25 °C and ~70% humidity.
Activity recording
Two- to 3-day-old virgin males (except when specified) of each species were placed individually into glass tubes (3 or 5 mm diameter and 65 mm long) and recorded using Drosophila activity monitors (TriKinetics, Waltham, MA) at light intensity ~0.28 W/m2 and temperature 25 ± 0.5 °C. To estimate free-running period (τ), flies were first exposed to 2 days of LD 12:12, after which they were released into constant darkness (DD) in an incubator (MIR-154; Sanyo, Tokyo, Japan). Activity in constant light (LL) was assayed after 4 days of LD 12:12, when lights remained ON throughout. DD experiments were repeated at least 4 times with similar results (Table 1). Photoperiods LD 20:4, 18:6, 16:8, 14:10, 12:12, 10:14, 8:16, 6:18, and 4:20 were created in an incubator (DR-36VLC8; Percival Scientific, Perry, IA).
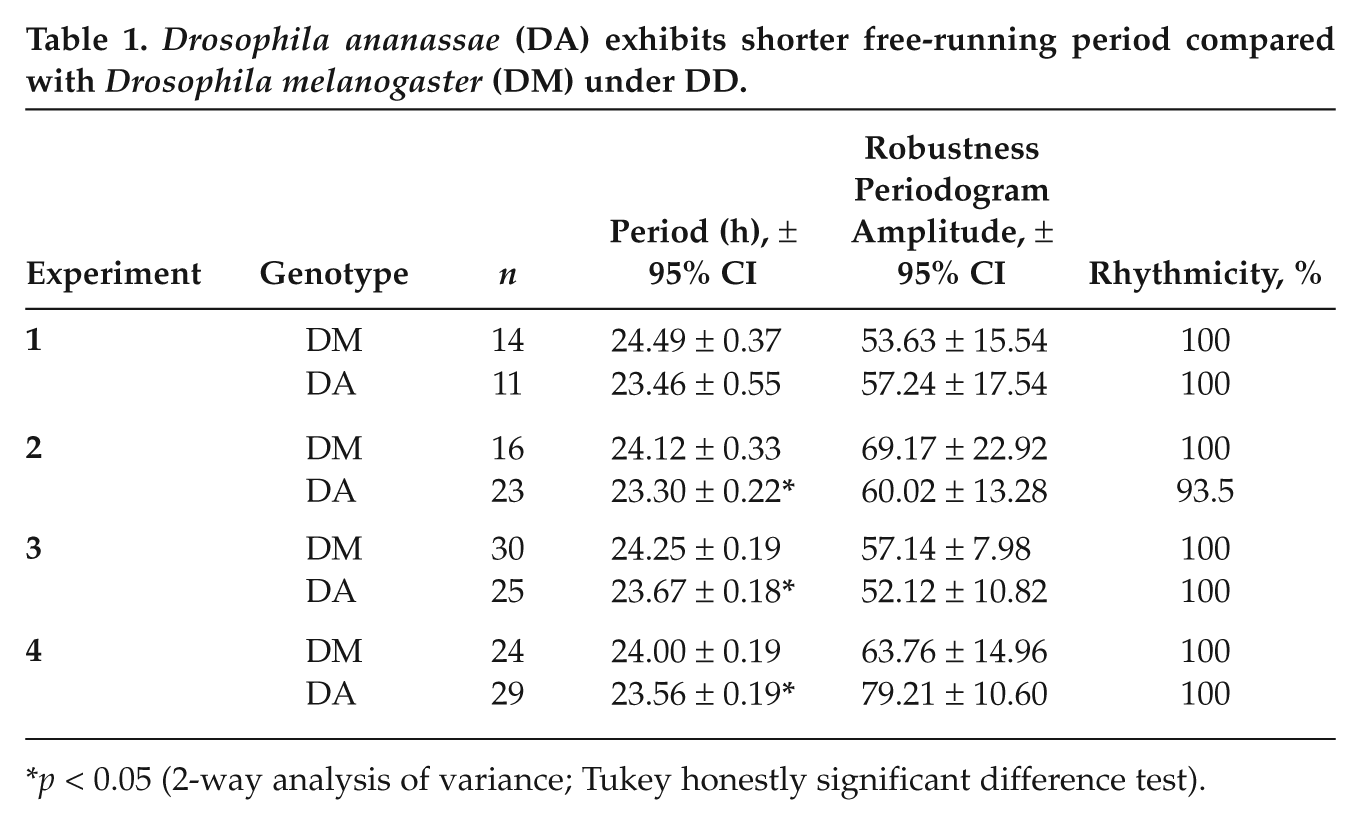
Drosophila ananassae (DA) exhibits shorter free-running period compared with Drosophila melanogaster (DM) under DD.
p < 0.05 (2-way analysis of variance; Tukey honestly significant difference test).
Analysis of activity
Activity was recorded in 5-min bins. Raw time-series data from individual flies further binned into 15 min were used to obtain average actograms. For Figure 1, activity profiles were obtained by averaging raw activity counts across 7 days for each fly and averaging across flies. Morning and evening anticipation indices were estimated only for LD 12:12 for each species since “true” peaks coincided with D/L and L/D transitions only in this regime. Data of individual flies were averaged across 7 days, and ratios of activity in the final 3 h prior to lights-on and lights-off, to that which occurs in the 6 h preceding the transitions (Harrisingh et al., 2007), were used as indices of anticipation and compared across species using the Student t test.
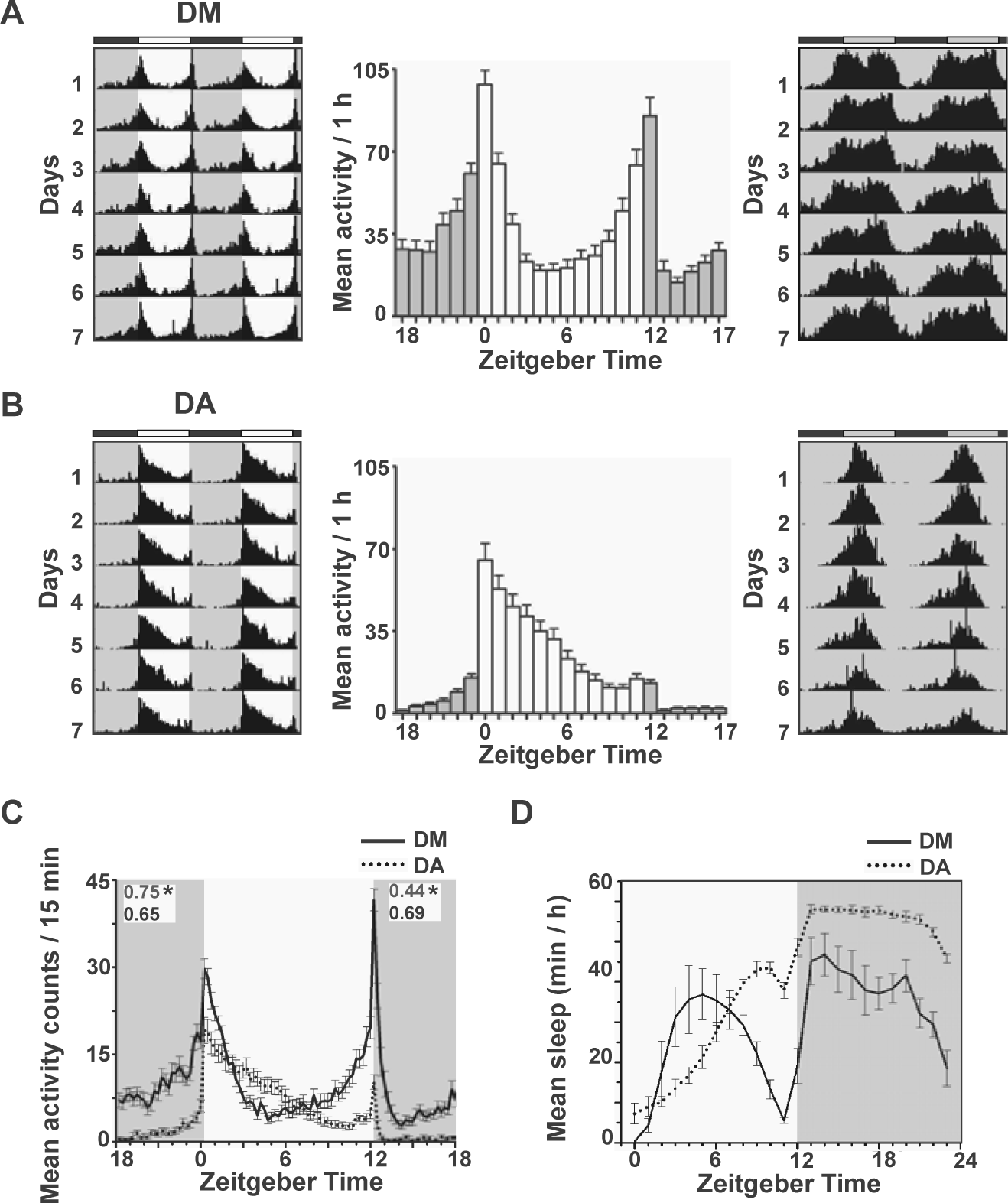
Drosophila ananassae (DA) showed preference for morning activity compared with Drosophila melanogaster (DM) under LD 12:12. (A) Average double-plotted actograms of male DM (n = 28) under LD 12:12 at 25 °C (left) and DD (right, n = 25). The x-axis represents time of day from 0 to 48 h; consecutive days are plotted along the y-axis. Histograms show average activity in 1-h bins across 7 days under LD 12:12 (mean ± SEM). Gray-shaded areas in actograms represent darkness while gray bars in the histogram (middle panel) denote activity in the dark phase under LD 12:12. (B) Actograms and activity profiles of male DA (n = 28) in LD and DD (n = 30). All other details are the same as in panel A. (C) Raw activity counts in 15-min bins under LD 12:12 averaged across 7 days for both DA and DM. Morning anticipation (top left values) of DA is significantly higher than that of DM (*p < 0.05). The evening peak of DA is mostly a response to lights-off with very little anticipation for the light/dark transition (top right values), which is significantly lower than that of DM (*p < 0.05). Error bars are SEM. (D) Sleep profiles under LD 12:12 show midday peak in sleep for DM (n = 28), while DA show a gradual increase in sleep throughout the day phase with highest daytime sleep just before dusk (n = 28). Error bars are 95% confidence interval. *p < 0.05.
Sleep analysis
Based on previous studies on DM, sleep was defined as any duration of uninterrupted immobility (0 counts/min) lasting ≥ 5 min (Andretic and Shaw, 2005). Sleep profiles for individual flies were plotted by averaging minutes of sleep per hour across 7 days. These profiles were then averaged across individual flies to obtain average sleep profiles. The interval of time between sleep bouts was considered as a bout of waking. Average number of sleep bouts per 30 min and average duration of sleep-bouts and wake-bouts were estimated for individual flies for each day using a macro in Excel (MS Office; Microsoft, Redmond, WA) written by Paul Shaw and modified by the Cirelli lab. Activity and sleep levels were analyzed using 2-way analysis of variance (ANOVA) with species and time interval as fixed factors followed by the post hoc Tukey honestly significant difference (HSD) test.
Free-running period
Raw time-series data obtained under DD were analyzed using Lomb-Scargle (LS) periodogram method in ClockLab (Actimetrics, Wilmette, IL), with p = 0.05 as a threshold for rhythmicity. To avoid transients, only data from the last 7 days were used for analysis. The τ and robustness (amplitude of LS periodogram) (Table 1) were compared using 2-way ANOVA followed by the Tukey HSD test.
Photoperiod analysis
Average activity profiles (mean ± SEM) were plotted using 5-min bin data for both species by first averaging across days for individual flies and then averaging across flies. To avoid transients, data from the last 7 days of entrainment were used for the analysis (except in the case of LD 4:20, where only the last 6 days were used because of the larger number of transients). We first manually scanned activity profiles of individual flies obtained after averaging across 7 days and identified 15-min time windows that corresponded to “true” (having anticipation and/or gradual decline in activity) peaks and also startle responses associated with morning and evening transitions (Table 2). In cases where startle peak was indistinguishable from true peak, they were considered as the true peak. The true peak phase values thus obtained for individual flies were averaged across flies to obtain mean phases of the peaks for each species in each photoperiod.
Number of flies showing true peaks (T) or startle peaks (S) for morning and evening transitions.
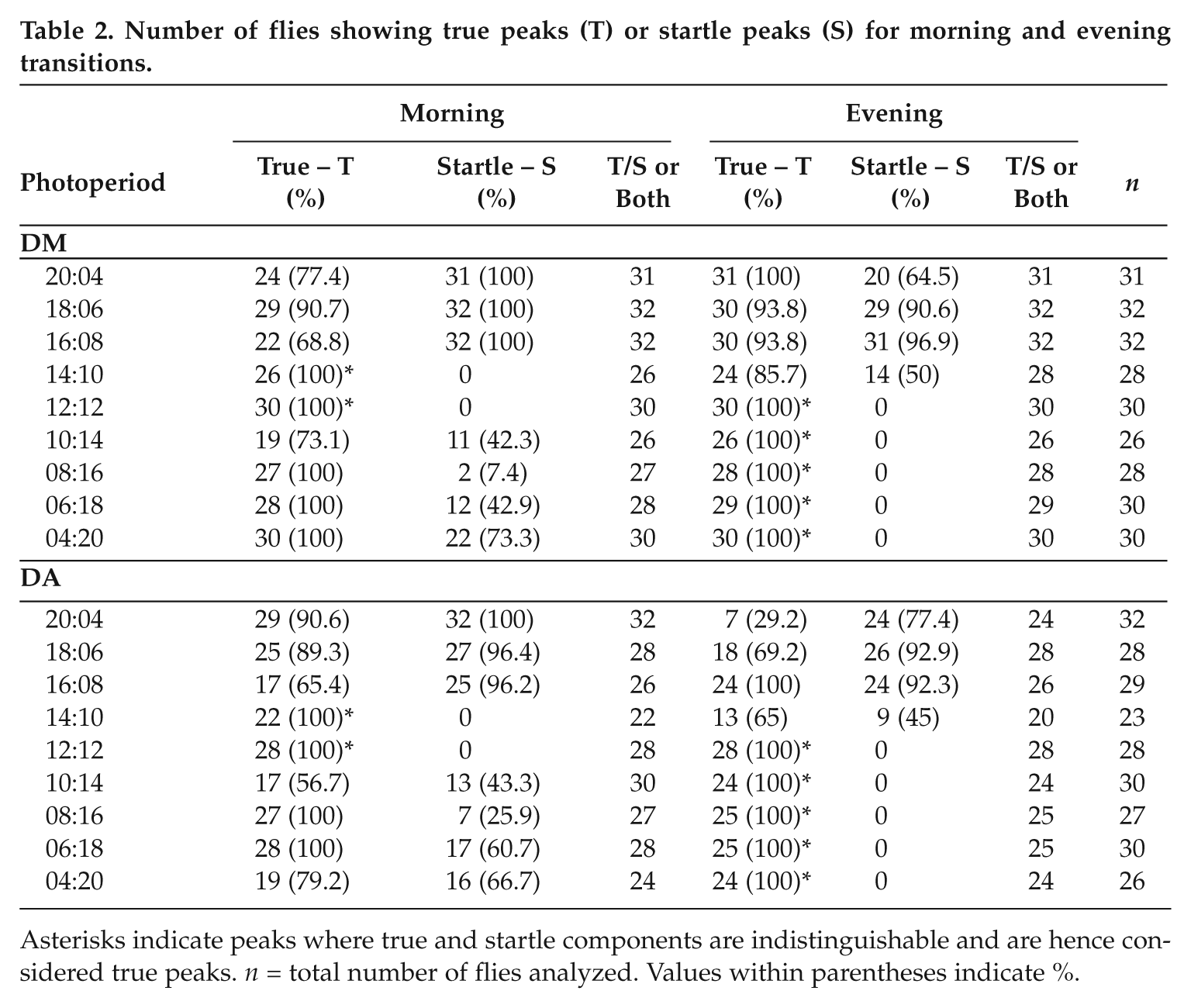
Asterisks indicate peaks where true and startle components are indistinguishable and are hence considered true peaks. n = total number of flies analyzed. Values within parentheses indicate %.
To quantify morning and evening preference for activity in 2 species under different photoperiods, we compared levels of activity during different parts of the day or night (Figs. 2 and 3). For long photoperiods, we estimated forenoon (FN) and afternoon (AN) indices during the light part of the day when flies were most active. We divided the light duration into 4 equal quarters, excluding 30 min after lights-on to avoid startle activity. The FN index was calculated as the ratio of activity during the first quarter to that during the first half of the light phase. The AN index was calculated as the ratio of activity during the last quarter of the light phase to that during the second half of the light phase.
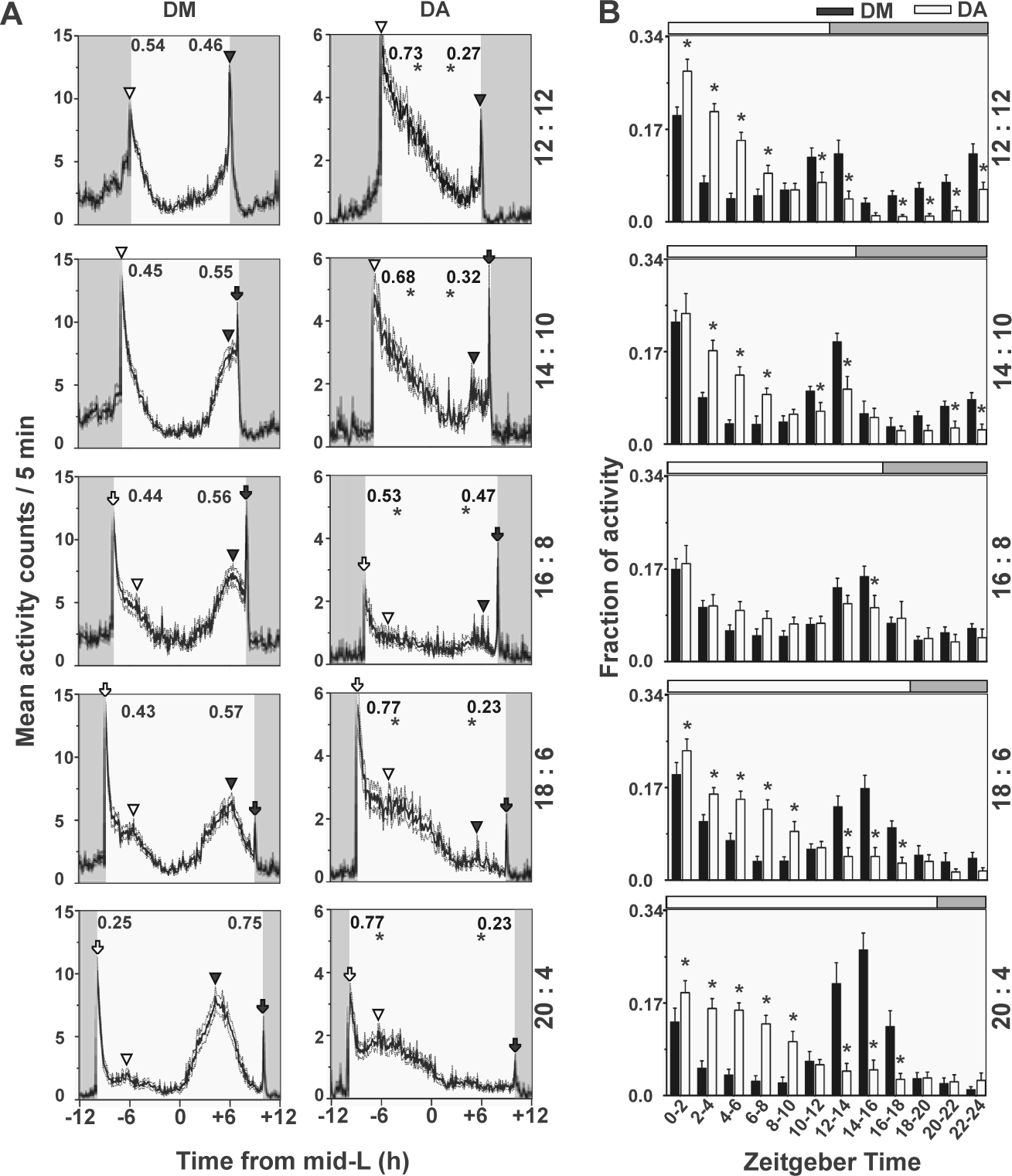
Drosophila ananassae (DA) exhibited higher forenoon activity while Drosophila melanogaster (DM) exhibited higher afternoon activity under long photoperiods. (A) Activity profiles with mean activity counts in 5-min bins (±SEM) averaged across 7 days. In each profile, morning and evening preference estimated by forenoon (FN; top left) and afternoon (AN; top right) activity indices are indicated for both species. Under all long photoperiods, DA show significantly higher morning preference compared with DM. DM show higher evening preference compared with DA. Arrowheads indicate presence of true peaks, while arrows indicate startle peaks when shown by more than 50% flies (Table 2). Unfilled (white) symbols denote morning peaks and black symbols, evening peaks. (B) Quantification of mean activity (2-h bin) normalized to total activity (±95% confidence interval). Horizontal white and gray bar above each graph denotes duration of light and dark, respectively, in all photoperiods. All other details are in Figure 1.
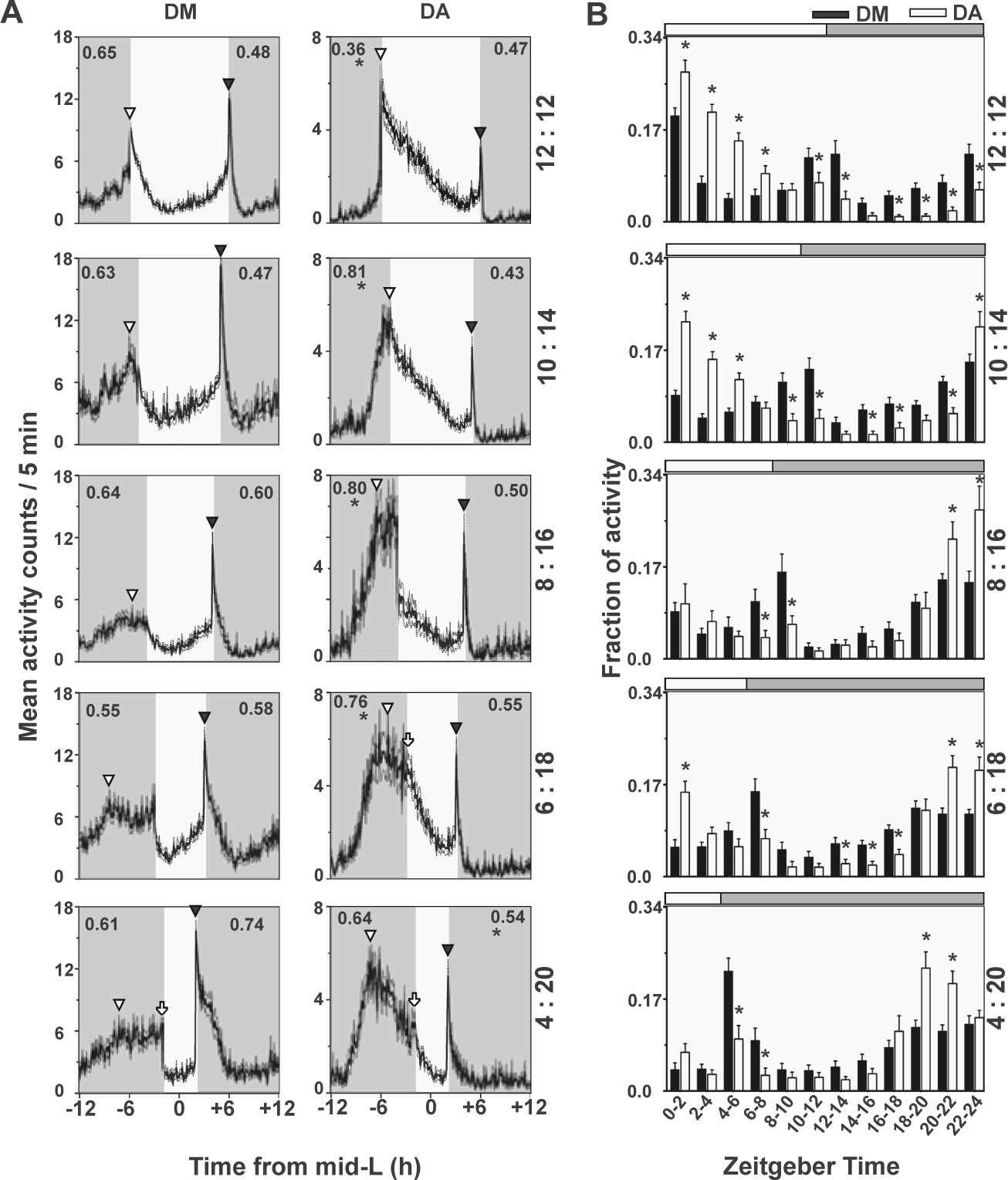
Under short photoperiods, Drosophila ananassae (DA) exhibited clear predawn activity peak compared with Drosophila melanogaster (DM). (A) Predawn (Pr-DN, top left of each profile) and postdusk (Po-DK, top right) activity indices show that DA exhibit greater morning preference compared with DM except for LD 4:20. Po-Dk is not significantly lower than DM except under LD 4:20. (B) Quantification of mean activity (2-h bin) normalized to total activity (±95% confidence interval). All other details are similar to Figure 2.
Under short photoperiods, since flies were mostly active at night, we first excluded 30-min data just after lights-off to eliminate the startle response. We then divided the entire night into 4 equal quarters. We defined an index of activity during the predawn segment (Pr-DN) as the ratio of activity in the last quarter of the night to that during the second half of the night. To examine activity patterns after the L/D transition, we defined the postdusk (Po-DK) index as the ratio of activity in the first quarter of the night to that in the first half of the night.
To compare distribution of activity of the 2 species across both light and dark durations within each photoperiod, the 24-h day was divided into 2-h intervals, and lights-on was considered as ZT0 in all photoperiods. Normalized mean activity in each 2-h interval was estimated, and 2-way ANOVA with species and time as fixed factors, followed by Tukey HSD, was performed to evaluate differences. Pr-DN, Po-DK, FN, and AN indices and phase of morning and evening peaks were derived from 15-min binned activity profiles and compared using the Student t test.
All statistical tests were done using STATISTICA-7 (StatSoft, Inc., Tulsa, OK) with level of significance set to p < 0.05.
Results
DA exhibited unimodal activity pattern compared with bimodal activity of DM under LD 12:12
Comparison of activity profiles of the 2 species under LD 12:12 revealed that unlike DM, DA showed a distinct unimodal morning preference in activity (Fig. 1A-C). As expected, virgin male DM showed 2 peaks of activity that coincided with lights-on and lights-off, with anticipation to both lights-on and lights-off and reduced activity both during midday and midnight (Fig. 1A, left and middle panels; Fig. 1C). In contrast, DA males showed a single peak of activity coinciding with lights-on, following which activity counts gradually tapered off (Fig. 1B, left and middle panels; Fig. 1C). Although both species showed anticipation to lights-on, the index was significantly higher for DA compared with DM (0.75 ± 0.09 vs 0.65 ± 0.03, mean ± 95% confidence interval [CI]; Fig. 1C). This was due to a steeper increase in activity in DA immediately before lights-on and very little nighttime activity compared with DM (Fig. 1A,B, left and middle panels; Fig. 1C). On the other hand, anticipation to the L/D transition in DA was very low and could be visualized only at higher resolution (15-min bins; Fig. 1C), which became undetectable when data were binned in 1-h intervals (Fig. 1B, middle panel). Compared with DM, it was significantly lower, as estimated by evening anticipation indices (0.44 ± 0.07 vs 0.69 ± 0.05, mean ± 95% CI; Fig. 1C). While daytime activity levels did not differ between the 2 species (DM = 499.4 ± 40 and DA = 397.1 ± 41 counts/12 h), DA showed significantly lower nighttime activity (65.68 ± 7.6 counts/12 h vs 432.8 ± 38, mean ± 95% CI; 2-way ANOVA, Tukey HSD). Such morning preference was also seen in females (Suppl. Fig. S1A,B). When male flies were allowed to free-run under DD after 2 to 4 days of LD, we found that DA activity in DD followed from the morning peak in LD, whereas the DD activity of DM predominantly followed from the evening peak (Suppl. Fig. S1C). Thus, we find that these 2 species exhibit different temporal preferences for activity under LD cycles.
Next, we examined whether the altered phasing of activity in these 2 species is achieved because of differences in τ. DA showed significantly shorter τ compared with DM (F1, 164 = 69.32, p < 0.05; Fig. 1A,B, right panels; Table 1) with no significant difference in robustness or percentage rhythmicity. Moreover, even under DD, DA showed a single narrow band of activity, whereas DM activity was much broader and appeared to be composed of elements from both morning and evening components (Fig. 1A,B, right panels). The average wake-bout duration of DA under DD was only 3.1 ± 0.2 h, whereas that of DM was 7.6 ± 0.4 h. Thus, these 2 species with modest differences in τ adopt distinct activity patterns under LD 12:12 and DD. To examine whether light promotes and/or darkness inhibits activity of DA and also to verify whether circadian clocks of this species are also susceptible to constant light (LL), we assayed activity of males of both species under LL and found that, like DM, DA were also arrhythmic in this regime (Suppl. Fig. S1D).
We expected that the difference in activity patterns under LD would also be reflected in their patterns of sleep. Since previous studies in DM have demonstrated that 5 min of immobility can be considered as sleep, we reasoned that the same criterion is likely to provide a reasonable estimate of a sleep-like state in DA also, although we have not performed experiments to validate the same. Indeed, DA showed distinctly altered sleep patterns under LD 12:12 compared with DM with very little sleep in the early half of the day and lack of midday siesta (Fig. 1D). Daytime sleep of DA peaked around ZT10 with a minor dip at ZT11 (which corresponds with a low level of evening anticipation; Fig. 1C), whereas DM sleep levels fell between ZT8 and ZT11 (Fig. 1D). Moreover, DA slept more at night, remaining asleep for most part of the dark phase and only beginning to wake within about 1 h prior to dawn (Fig. 1B,D). Analysis of sleep levels showed significantly higher nighttime sleep in DA, although no significant difference in overall daytime sleep was detectable (Suppl. Fig. S2A). Comparison of sleep-bout numbers between the 2 species revealed that DA had a significantly higher number of daytime sleep-bouts and a significantly lower number of nighttime sleep-bouts (Suppl. Fig. S2B, 2-way ANOVA, Tukey HSD). These differences are expectedly accompanied by longer average nighttime sleep-bout duration in DA, although mean daytime sleep-bout duration did not differ from DM (Suppl. Fig. S2C, 2-way ANOVA, Tukey HSD). Taken together, it appears that unlike DM, which are considered crepuscular, DA clearly preferred to be active during daytime and exhibited more consolidated sleep at night.
Unimodal activity of DA persisted under short and long photoperiods
We asked if such preference for daytime activity in DA persists under different photoperiods and examined if oscillators that regulate activity/rest rhythm in these 2 species have differential sensitivity to light duration. We subjected flies to a range of photoperiods from extremely short (LD 4:20) to extremely long (LD 20:4; Suppl. Fig. S3A). We found that morning preference for activity in DA was further accentuated under short and long photoperiods. With increasing day length, DM gradually shifted most of their activity toward lights-off, while DA strongly preferred morning (Suppl. Fig. S3A). This was most evident in LD 20:4 (Fig. 2 and Suppl. Fig. S3A), where evening activity peak of DM occurred about 6 h ahead of lights-off, whereas the DA activity peak, while being damped, still remained mostly close to lights-on with very little activity toward the end of the light phase. Interestingly, at intermediately long photoperiods (LD 14:10 and LD 16:8), DA showed a small increase in evening activity just before lights-off, which disappeared when day length increased beyond 18 h (Fig. 2 and Suppl. Fig. S3A).
With decreasing day length, the activity peak of DM occurred under darkness, and invariably a blunted predawn peak occurred, although they continued to show a significant evening peak coinciding with lights-off and acquired more prominence in contrast to morning activity (Fig. 3 and Suppl. Fig. S3B). In contrast, DA continued to show a preference for the D/L transition even when day length was reduced to 4 h. DA timed most of their activity to predawn duration with a prominent morning peak, up to 4 to 6 h ahead of lights-on in the most extreme short photoperiod (LD 4:20; Fig. 3 and Suppl. Fig. S3B). Thus, over a range of photoperiods, DA show a preference for morning activity, suggesting that oscillators controlling this behavior are tightly coupled to dawn. This also suggests that circadian organization of DA is quite different from that of DM, thus providing a novel model system for comparative analysis with the canonical model on which most of our current understanding of invertebrate circadian circuit organization is based.
DA distributed most of its activity around the morning transition under both long and short photoperiods
To quantify morning and evening preference, we carried out finer analyses of activity in the 2 species by examining average activity profiles of individual flies under each photoperiod and assigning true or startle tags to the peaks (Table 2, Figs. 2 and 3). Next, we compared activity levels between the 2 species during various parts of the day/night under long (Fig. 2) and short photoperiods (Fig. 3). Under long photoperiods, most of the activity and differences between species were seen during daytime, and hence we estimated forenoon and afternoon activity indices (FN and AN; see Materials and Methods). Under LD 12:12, although both species exhibited high activity during the early half of the day (Fig. 2A), DA showed significantly higher FN and lower AN compared with DM. Comparison of activity in 2-h bins showed that DA exhibited significantly greater activity during the light phase for the first 8 h, whereas both in the last 2 h of the day and for most of the night, they displayed significantly lower activity compared with DM (Fig. 2B). Upon increasing day length to 14 h, a similar pattern of differences between species persisted (Fig. 2B). Comparison of 2-h bins showed that the differences in activity are significant for most of the day except the first 2-h bin, which likely included the startle response in both species (Fig. 2B). As seen in LD 12:12, DA became significantly more active compared with DM during the forenoon and less active compared with DM during the afternoon (Fig. 2B). In the intermediately long photoperiod LD 16:8, both species appeared to show similar patterns of activity, with DA showing a hint of evening activity (Fig. 2A and Table 2), while DM showed bimodal but blunted morning and evening peaks. Comparison of 2-h bin activity showed no difference between species during most of the day and night in this photoperiod except in the bin immediately preceding lights-off, when DA were less active (Fig. 2B). With increasing day length (LD 18:6 and LD 20:4), the evening activity peak became predominant in DM, whereas DA continued to phase most of their activity to the early part of the day. Under LD 18:6, neither DM nor DA appeared to anticipate lights-on, but a phase-delayed, true morning peak was detectable (Fig. 2A) in ~90% flies in both species (Table 2). While the true evening peak of both species was shifted well into the day and advanced compared with the L/D transition, the DA evening peak was of very low amplitude (Fig. 2A). Comparison of 2-h bins showed that DM exhibited significantly lower activity in the forenoon, while in the last 6 h of the day, the pattern was reversed—DM became significantly more evening-active, as evidenced by the presence of the prominent evening peak that reappeared in this photoperiod (Fig. 2B). Under LD 20:4, DM showed a prominent evening peak that was as high as the morning startle (Fig. 2A). Comparisons of 2-h activity bins showed that DA had significantly higher activity up to 10 h after lights-on, whereas this trend was reversed between ZT12 and ZT18. Under all long photoperiods, FN and AN indices showed completely opposite trends in the 2 species, with DA exhibiting significantly high FN and low AN activity compared with DM (Fig. 2A). Thus, under increasing photoperiods, evening activity became predominant in DM, whereas in DA, the morning component continued to persist while any signs of evening activity became undetectable under the extremely long photoperiod, suggesting that the morning oscillator is the dominant entity in DA.
We quantified the difference in activity profiles under short photoperiods using another set of indices: predawn (Pr-DN) and postdusk (Po-DK; see Materials and Methods). Under LD 12:12 (Fig. 3A), Pr-DN activity of DA was significantly lower than that of DM, whereas Po-DK activity did not differ. When day length was made less than 12 h (LD 10:14, 8:16, 6:18), DA shifted most of their activity to the interval preceding dawn such that Pr-DN became significantly higher than that of DM (Fig. 3A). Under LD 8:16 and 6:18, the activity of DA preceding dawn lasted for even longer durations than in LD 10:14 (Fig. 3). However, no difference in Po-DK activity was detectable between species, although DA appeared to show very little activity during that interval (Fig. 3A). Comparing 2-h data in LD 10:14 revealed that during early morning (i.e., up to 6 h after lights-on), DA were more active than DM (Fig. 3B). This trend was reversed toward the end of the day and continued through most of the night when DM were more active compared with DA except for the bin immediately preceding lights-on. Under LD 8:16 and 6:18, DM switched from a clear bimodal pattern of activity to showing a prominent evening peak, with Pr-DN activity becoming flattened and spread out over a period of 4 to 6 h (Fig. 3A). Here, although all flies showed a low-amplitude true morning peak, less than 43% showed morning startle (Table 2) with large interindividual variance in the peak phase (Fig. 4). While some degree of flattening of the morning peak occurred even in DA, 100% of the flies exhibited a high-amplitude true morning peak (Fig. 3A and Table 2). In all short photoperiods, it was not possible to distinguish the evening startle activity from a potential true peak in either species, since they all appeared to have some degree of anticipatory activity except under LD 4:20. Under an extremely short photoperiod (LD 4:20), no difference in Pr-DN was detectable between species, although DA showed a distinct peak compared to flattened morning activity of DM (Fig. 3A). In contrast, Po-DK activity of DM was significantly higher than that of DA (Fig. 3A). The sharper predawn peak of DA was detected as significantly higher values during ZT18 to ZT22 (Fig. 3B). Thus, our analyses point toward morning preference for activity in DA compared with DM even under extremely short photoperiods and the possibility of an oscillator controlling the diurnal activity of DA being strongly coupled to dawn.
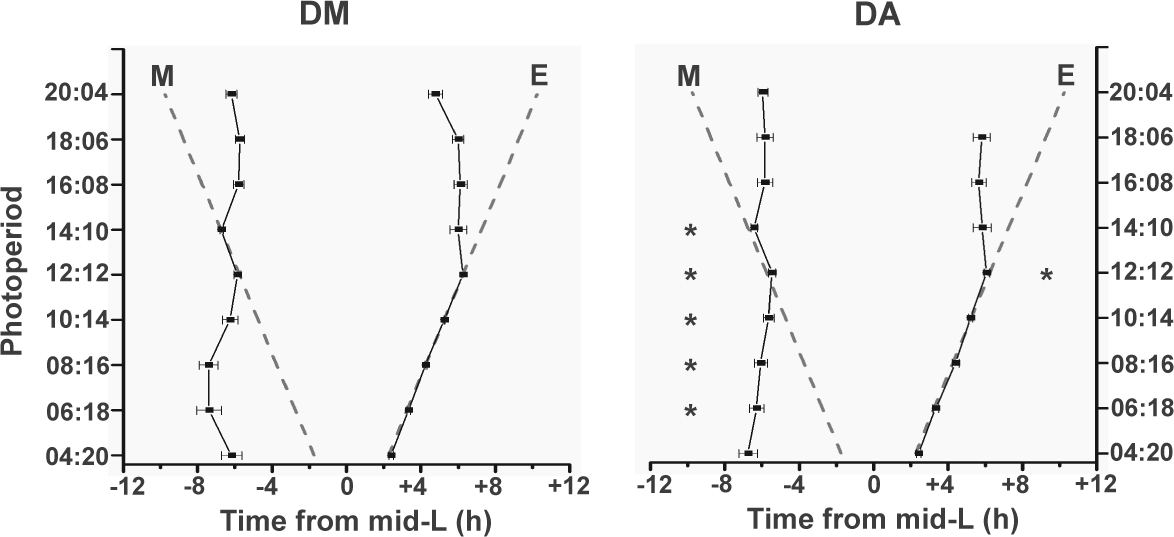
Phasing of true peaks of Drosophila melanogaster (DM) and Drosophila ananassae (DA) under different photoperiods. Phase plots of true morning and evening peaks under all photoperiods for both species (mean ± 95% confidence interval). Dashed gray lines indicate phases of lights-on and lights-off in each photoperiod. M = morning; E = evening. *p < 0.05.
We compared the phasing of true peaks of the 2 species under both increasing and decreasing photoperiods (irrespective of amplitude) to examine whether there may be differences among species in coupling strengths of the circadian clocks with either morning or evening transitions (Fig. 4). Under LD 12:12, DM showed tight coupling of both morning and evening peaks with D/L and L/D transitions, respectively. The morning peak of DA was significantly delayed, while the evening peak was advanced (Fig. 4). Under LD 14:10, the morning peak of DA was phase-delayed compared to DM, while evening peaks of both species were similarly phased. Under day lengths longer than 14 h, both species showed similar phase relationships with both transitions, except in LD 20:4, where the true evening peak was shown only by <30% of DA flies (Table 2). Under short days (LD 10:14, 8:16, and 6:18), DA were consistently less phase-advanced compared with DM (by 0.7 h, 1.4 h, and 1.1 h, respectively) and showed lesser interindividual variation (Fig. 4). Interestingly, under extremely short days (LD 4:20), both species have similarly advanced morning peaks (Fig. 4). True evening peaks of both species were indistinguishable from startle under short days or LD 12:12.
Thus, DA are clearly a morning-preferring species compared with DM, whose activity, although symmetrically distributed around both morning and evening transitions under LD 12:12 conditions, reveal themselves to be a predominantly evening-preferring species when examined under either long or short photoperiods. The above results underscore separation of the 2 species on a temporal axis and suggest that these 2 sympatric species probably adopt different physiological measures to deal with the environmental challenges under different photoperiods.
Discussion
Our studies suggest that 2 sympatric species of Drosophila may occupy distinct temporal niches under identical environmental conditions. We show that DA prefer to remain active during the day and limit their activity at night to extremely low levels. Interestingly, the time at which DA were most active, under LD 12:12 (ZT1-6), also coincided with the time of day when DM rapidly reduced their activity (Fig. 1 and Suppl. Fig. S1). It is likely that DM avoid activity during midday when minimal humidity, maximum temperature, and maximum light levels occur to minimize chances of desiccation (Low et al., 2008). Therefore, it is puzzling that DA, a close relative of DM, in fact prefer to be most active at this time (Fig. 1 and Suppl. Fig. S1). Moreover, DM showed a distinct bimodality similar to many crepuscular animals that are thought to use the relatively milder conditions of twilight to minimize the effects of environments at other harsher times of the day. Such preference for twilight was clearly lacking in DA. It is possible that on a local scale, the 2 species occupy different habitats even though they occur in the same geographical areas. Anecdotal evidence suggests that DA are more often found in areas around human habitation such as inside homes, kitchens, garbage dumps, markets, and grocery stores, whereas DM are more likely to be caught around orchards and open fields. We speculate that systematic net sweep studies done in a time-logged manner may reveal that the 2 species experience very different micro-climates while living in the same area.
Compared with DM, DA exhibit much more consolidated sleep at night. During the day, even though mean sleep-bout duration of DA does not differ from DM, the former exhibits a greater number of daytime sleep-bouts, suggesting that these flies do not have sustained activity during their preferred active phase and that activity is interspersed with short sleep-bouts. In contrast, their nighttime sleep is more consolidated compared with DM as they exhibit significantly fewer sleep-bouts and also significantly longer mean sleep-bout duration. Such differences also persist under DD, where only 1 narrow band of activity is exhibited by DA compared with more spread-out activity of DM (Fig. 1 and Suppl. Fig. S1C). DM may have the required physiology and sensory apparatus that allow activity even in the absence of light. Nights are accompanied by cooler temperatures, and hence it is possible that DA avoid activity at night and that may partly be the reason for their distribution being restricted to warmer, more tropical clines compared with DM (Das et al., 2004). Future studies on other physiological and behavioral aspects of sleep in DA such as the arousal threshold, sensitivity to temperature, and light intensity may help in a better understanding of DA sleep characteristics. One of the limitations of our studies is that they were all done under laboratory conditions, and thus, future studies that examine rhythmic behaviors under cycling temperature cycles and/or seminatural conditions may provide further clues toward the ecological significance of this temporal preference.
Our studies show that phasing of the morning peak with the D/L transition is modulated by photoperiod in both species (Fig. 4). Under LD 12:12, it is tightly coupled with lights-on, especially in DM, and both species show a significant advancement of phase under short-day conditions, with DM being more advanced compared with DA. This suggests that if indeed dual oscillators that are coupled to dawn and dusk regulate the 2 peaks in these 2 species, then the morning oscillator of DA has a stronger coupling to lights-on compared with DM. On the other hand, in DM, the persistence of the evening peak, especially under long photoperiods, suggests the presence of a dominant evening oscillator. The coupling of evening oscillator to dusk in DM is further confirmed upon release into DD (from LD 12:12), by the persistence of a major evening activity component and its phasing (Suppl. Fig. S1C). This suggests that strong morning and evening oscillators regulate DM activity behavior (Grima et al., 2004; Stoleru et al., 2004, reviewed by Helfrich-Förster, 2009), whereas DA probably have only a dominant morning oscillator and a much weaker evening oscillator. Our results do not rule out the existence of an evening oscillator in DA since these flies show small bouts of evening anticipatory activity under several regimes, especially becoming evident under intermediately long photoperiods (LD 16:8 and 18:6; Fig. 2A). On the basis of the coupled-oscillator model of Pittendrigh and Daan (1976), previous studies have hypothesized that in DM, the morning oscillator is dominant under short photoperiods while the evening oscillator has a greater effect under long photoperiods (Stoleru et al., 2007). Under long photoperiods, DA show high variation in the evening peak phase both among individual flies and across days (Figs. 2A, 4), suggesting that under a long photoperiod, even in DA, the otherwise dominant morning oscillator coexists with a strengthened evening oscillator. In comparison with the neuronal substrates of the morning and evening oscillators that have been unraveled in DM, DA would be expected to have either a greater number of so-called morning cells (which was not seen in a recent study by Hermann et al., 2012) or that “morning” cells exert a stronger influence on the motor circuits controlling activity rhythm. Alternatively, a lesser number or weaker coupling of evening cells with the circuit can result in such a dominance of the morning oscillator in the organization of circadian pacemakers of DA.
It is also possible that the cellular identity of the morning oscillator in DA is different from that of DM, and thus, our studies point toward potential differences in the nature of the underlying circadian pacemaker circuit and/or its outputs in DA compared with DM. Interestingly, the morning preference in DA is also accompanied by a faster ticking clock, suggesting that the molecular clockwork may also differ from that of DM. Hermann and colleagues (2012) find no difference between DM and DA in terms of cell number among PDF-, VRI-, and PDP1-expressing cells, but minor differences were seen in the projection patterns of ventral lateral neurons. They speculate that in contrast to DM, where only 1 subset— the small ventral lateral neurons, which modulate morning activity—sends projections toward the dorsal protocerebrum, in DA, the second subset, the large ventral lateral neurons, also project dorsally. Another significant anatomical difference between the 2 species was seen in the number of neuropeptide ITP-expressing cells, ipc-2 and ipc-4, although the role of these neurons in regulating diurnal activity/rest behavior is as yet unknown. In DA, compared with DM, fewer neurons of the ipc-2 subset were seen, while the ipc-4 subtype was undetectable. Taken together with our own results, this suggests that in these 2 closely related species with high homology in the sequences of the genes and proteins involved in circadian clocks and also similar neuroanatomical features of circadian neuronal network, the striking divergence in temporal preference for activity/rest behavior probably arises from some subtle differences in the coupling among those neurons. We propose that DA may serve as a useful model to conduct comparative studies of the neuronal circuitry and underlying genetic basis of circadian clocks, given that the DA genome has been fully sequenced (Drosophila 12 Genome Consortium, 2007). In the future, modern tools of transgenesis that have proven to be invaluable for DM could also be modified to be used in DA. Furthermore, downregulating or enhancing neuropeptide (or its receptor) expression or manipulating electrical properties by ectopic expression of ion channels within the circadian clock neurons may reveal features of circadian circuit organization that have not been discovered through studies on DM. Our studies suggest an interesting temporal separation of behavior that may allow better understanding of the functional significance of timing of activity. Functionally, this preferential timing could imply better competitive advantage, foraging, and mating-related movements and/or exploratory behavior. We propose that comparison of closely related sympatric species under natural conditions in parallel with laboratory studies will allow additional insights into mechanistic details and functional significance of circadian clocks and the behaviors that they control.
Footnotes
Acknowledgements
The authors thank 3 anonymous reviewers for critical comments on a previous version of the manuscript, as well as Vijay Kumar Sharma, Sheetal Potdar, and other lab members for comments and suggestions. Sudeshna Das is greatly acknowledged for the collection of both species. The authors also thank Rajanna and Muniraju for assistance in experiments. This work was supported by a Ramanujan Fellowship (Department of Science and Technology, India) to V.S. and Council of Scientific and Industrial Research (India) fellowship to P.M.P.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.