
Abstract
When food is available during a restricted and predictable time of the day, mammals exhibit food-anticipatory activity (FAA), an increase in locomotor activity preceding the presentation of food. Although many studies have attempted to locate the food-entrainable circadian oscillator in the central nervous system, the pathways that mediate food entrainment are a matter of controversy. The present study was designed to determine the role of dopaminergic and histaminergic systems on FAA. Mice were given access to food for 2 h (ZT12-ZT14), and FAA was defined as the locomotor activity that occurred 2 h before the availability of food. Dopamine D1 receptor (R), D2R, and histamine H1R-specific antagonists were used to clarify the role of dopamine and histamine receptors in FAA induced by food restriction (FR). FAA was monitored by infrared locomotor activity sensors. Mice were sacrificed at ZT12 on the 14th day of FR, and monoamine concentrations were determined by high-performance liquid chromatography coupled to electrochemical detection (HPLC-ECD). The results showed that pretreatment with the D1R antagonist SCH23390 at 1, 3, or 10 µg/kg significantly reduced FAA by 19% (p < 0.05), 26% (p < 0.05), or 19% (p < 0.01), respectively, and the D2R antagonist raclopride at 22, 67, or 200 µg/kg significantly reduced FAA by 16% (p < 0.05), 36% (p < 0.01), or 41% (p < 0.01), respectively, as compared with vehicle control. Moreover, coadministration of SCH23390 (10 µg/kg) and raclopride (200 µg/kg) synergistically inhibited FAA by 57% (p < 0.01) as compared with vehicle control. Consistently, the levels of dopamine and its metabolites in the striatum and midbrain were significantly increased during FAA, even with the pretreatment of D1R and D2R antagonists. However, pretreatment with pyrilamine at 2.5, 5, or 10 mg/kg did not significantly reduce FAA, although it reduced the locomotor activity during the dark period in ad libitum mice. These results strongly indicate that the dopaminergic system plays an essential role in the FAA in mice.
When food is available during a restricted and predictable time of the day, mammals quickly develop increased locomotor activity preceding food access, and this has been termed food-anticipatory activity (FAA) (Mistlberger, 1994). This food entrainment has been linked to the expression of a circadian oscillator because FAA is only entrained when food is provided in a circadian range, with free runs for several cycles during constant-fasting conditions, and it exhibits transients in response to phase shifts of food access (Stephan, 1992).
The suprachiasmatic nucleus (SCN) in the hypothalamus has been identified as the light-entrained oscillator, whereas the mechanisms underlying the food-entrained oscillator (FEO) remain elusive. FAA is robust after a complete bilateral lesion of the SCN (Stephan et al., 1979). Mutant mice lacking known circadian clock function in all tissues exhibit normal FAA both in a light-dark cycle and in constant darkness (Storch and Weitz, 2009), indicating that FAA does not require the known circadian clock. Effects to localize oscillators driving food-anticipatory behavioral rhythms have for many years relied on lesion experiments. In rats, cell-specific lesions of the dorsomedial hypothalamic nucleus (DMH) were reported to significantly, but only partially, attenuate or eliminate FAA (Gooley et al., 2006). However, other groups working with rats and mice showed different results for the role of DMH in the expression of circadian FAA (Landry et al., 2007; Moriya et al., 2009). Likewise, lesions of the ventromedial hypothalamic nucleus (VMH), the lateral hypothalamic area (LH), the paraventricular hypothalamic nucleus (PVH), and the olfactory bulb did not prevent FAA (Davidson, 2009). Although these studies attempted to locate the food-entrainable circadian oscillator in the central nervous system, the pathways that mediate food entrainment are not yet fully understood.
Several studies have showed that both histaminergic and dopaminergic systems were involved in the regulation of arousal, locomotor activity, reward, and feeding circadian rhythm (Doi et al., 1994; Hood et al., 2010; Inoue et al., 1996; Qu et al., 2010). In restricted feeding condition, histamine-containing neurons in the tuberomammillary nucleus (TMN) increased Fos-ir expression before mealtime (Inzunza et al., 2000), and the increased locomotor activity that anticipates mealtime coincides with the activation of the TMN (Meynard et al., 2005). Carr et al. (2001) reported that intracerebroventricular injections of the D1 and D2 receptor (R) agonists produced greater reward and locomotor-activation effects in food-restricted than in ad libitum–fed rats, and the activation of ERK 1/2 MAP kinase, CREB, and Fos-immunostaining in caudate putamen and nucleus accumbens increased in food-restricted rats (Carr et al., 2003; Haberny et al., 2004). Moreover, food restriction strongly increased mRNA levels of tyrosine hydroxylase and the dopamine transporter in the ventral tegmental area (Lindblom et al., 2006). Based on these studies, we hypothesized that histaminergic and dopaminergic systems may play an important role in the mechanisms of FAA underlying food restriction.
D1R and D2R are the most widely and abundantly expressed receptors for dopamine in the brain (Kobayashi et al., 2004), and the histaminergic system regulates several functions such as arousal, circadian rhythm of locomotor activity, and exploratory behavior via the histamine H1 receptor (H1R) (Huang et al., 2001; Inoue et al., 1996). In the present study, we used a pharmacological and neurochemical approach to investigate the roles of dopaminergic D1R and D2R and histaminergic H1R in the FAA caused by FR. Our findings strongly indicate that dopaminergic system plays an essential role in the FAA in mice.
Materials and Methods
Animals and Housing Condition
Male ICR mice, weighing 25-27 g (8 weeks old), were obtained from the Laboratory Animal Center, Chinese Academy of Sciences (Shanghai, China). Animals were housed individually in an ambient temperature of 22 ± 0.5 °C with a relative humidity of 60% ± 2% and an automatically controlled 12 h light/12 h dark cycle (lights on at 0700 h, illumination intensity ≈ 100 lux), and mice had free access to food and water, unless otherwise stated. Experimental protocols were approved by the Medical Experimental Animal Administrative Committee of Shanghai. Every effort was made to minimize the number of animals used and any pain and discomfort experienced by the subjects.
Food Restriction Protocol
Mice were kept on a 12-h light/dark cycle (ZT0 defined as lights-on time [0700 h] and ZT12 as lights-off [1900 h]). After having free access to food and water for 4 days, experimental mice were subjected to 2 kinds of feeding conditions for 14 days. The FR mice were maintained with 2 h of food access per day and free access water. The group fed ad libitum (ad libitum mice) had free access to food and water. To ensure that the manipulation of removing and adding food did not interfere with the light-dark cycle during the light periods, we chose the restricted feeding regime: the food was given at ZT 12 (lights off) and withdrawn at ZT 14 (Fig. 1A).
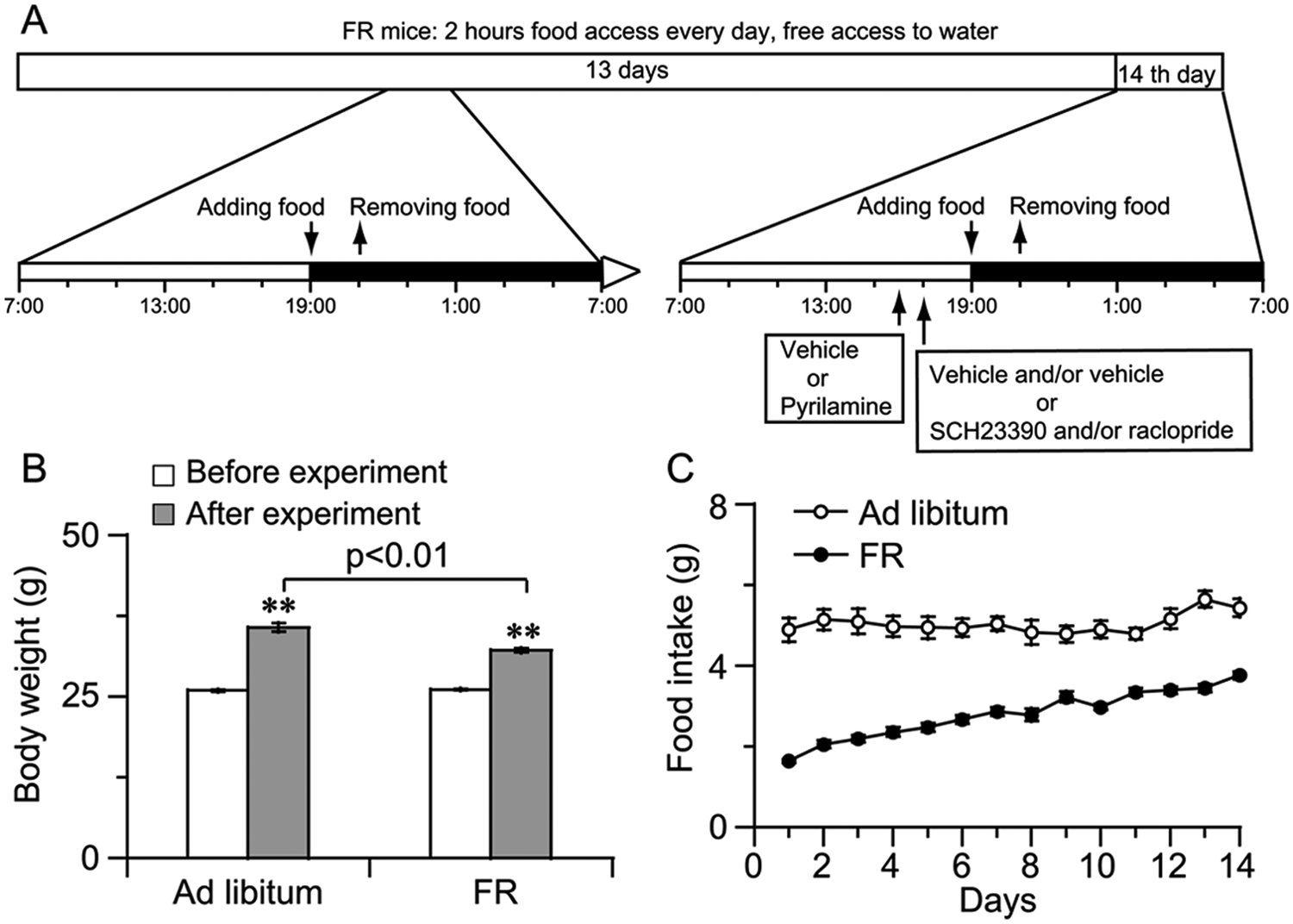
Schematic for FR and the health status of FR mice. (A) FR mice were kept on a 12 h light/dark cycle and maintained with 2 h of food access every day and free access water. Pyrilamine was given at 1600 h, and SCH23390 and/or raclopride was administered at 1700 h. Horizontal open and closed bars indicate light and dark, respectively. (B) Changes in body weight of FR and ad libitum–fed mice before and after the experiment. (C) Time-course changes in food intake of FR mice and ad libitum mice for 14 days. **p < 0.01 vs. before the experiment, n = 12-14.
Body Weight and Food Intake
To track the health status of FR mice, the body weight of all mice was registered before and after the experiment. In addition, food intake for 2 h of food access every day for FR mice and for the 24-h interval for ad libitum mice was monitored for 14 days.
Locomotor Activity Recordings
Locomotor activity for an individual mouse was detected with a passive infrared sensor (Biotex, Kyoto, Japan) placed 17.5 cm above the floor of the recording cage (28 cm × 16.5 cm × 13 cm) as previously reported (Nakamura et al., 2008). FAA was defined as the total locomotor activity for 2 h (ZT10-ZT12) before food access (Poulin and Timofeeva, 2008).
Pharmacological Treatments
Pyrilamine (Sigma-Aldrich), a selective H1R antagonist; SCH 23390 (Sigma-Aldrich), a selective D1R antagonist; or raclopride (Sigma-Aldrich), a selective D2R antagonist, was dissolved in sterile saline immediately before intraperitoneal administration to the mice on the 14th day of FR. For exploring contribution of H1R, D1R, or D2R alone, and D1R/D2R together to FAA, an H1R antagonist pyrilamine alone was given at 1600 h (ZT9), whereas D1R antagonist SCH23390 or D2R antagonist raclopride alone, or combination of SCH23390 and raclopride was administered at 1700 h (ZT10) (Fig. 1A). To investigate the role of coadministration of D1R and D2R antagonists or H1R antagonists on the spontaneous locomotor activity during the dark period in ad libitum mice, SCH23390 and raclopride together or pyrilamine alone was administered intraperitoneally at 1900 h (ZT12).
The dosages for H1R antagonist pyrilamine (Huang et al., 2006) and D1R/D2R antagonists SCH23390/raclopride were selected based on previous reports and our preliminary data. In previous studies, SCH23390 at 15 and 50 µg/kg and raclopride at 0.5, 0.25, and 0.125 mg/kg did not influence spontaneous locomotor activity (Benturquia et al., 2008; Rodriguez-Arias et al., 2000). Our preliminary results showed that SCH23390 at 10 µg/kg and raclopride at 0.2 mg/kg did not decrease spontaneous locomotor activity in mice either, respectively. Therefore, we chose SCH23390 10 µg/kg and raclopride 0.2 mg/kg as the largest doses that did not affect spontaneous locomotor activity and selected 1 and 3 µg/kg for SCH23390 and 22 and 67 µg/kg for raclopride as small and middle dosages, respectively.
Sampling Procedure and Brain Dissection
Mice used for brain dissection were sacrificed at 1900 h on the 14th day of FR or ad libitum access, respectively. Following decapitation, the brains were rapidly removed and the cortex, hippocampus, striatum, midbrain, thalamus, hypothalamus, and medulla oblongata were dissected on ice according to previous report (Glowinski and Iversen, 1966). The tissues were weighed and then stored at −80 °C until analysis by HPLC-electrochemical detector (ECD).
Determination of Catecholamines and 5-HT Levels in Brain Regions
To determine dopaminergic neurotransmission in the FR-induced FAA, the levels of catecholamines and their metabolites together with 5-HT were determined using HPLC with electrochemical detection as reported previous (Wang et al., 2012). Briefly, frozen brain tissues were homogenized in ice-cold 0.2 M perchloric acid containing 100 µM Na2-EDTA and 1 ng/mL isoproterenol as an internal standard. The homogenates were centrifuged at 15,000 g at 4 °C for 20 min. The supernatant was mixed with 1 M sodium acetate. The volume of sample injected was 20 µL. The levels of norepinephrine (NA), dopamine (DA), and 5-hydroxytryptamine (5-HT) and their respective metabolites 3,4-hydroxyphenylacetic acid (DOPAC), homovanillic acid (HVA), and 5-hydroxyindoleacetic acid (5-HIAA) were determined by reverse-phase HPLC-ECD. A C18 reverse-phase column (EICOMPARK SC-5ODS 3.0 × 150 mm, 5 µm, Eicom, Kyoto, Japan), a precolumn (DIKMA 3.0 × 15.0 mm, 5 µm), and an electrochemical detector (Waters 2465, USA) were used. The mobile phase was 0.1 M acetic acid/citric acid buffer (pH 3.5) containing Na2-EDTA (10 mg/L), octanesulfonic acid (200 mg/L), and methanol (17%). The flow rate was 0.5 mL/min. The peak areas of the external standards were used to quantify the sample peaks.
Data Analysis
All results were expressed as mean ± SEM. Time-course changes in the amounts of the locomotor activity were compared between drug and vehicle treatment by using 2-way repeated-measures analysis of variance (ANOVA) followed by the post hoc Fisher probable least-squares difference (PLSD) test or 2-tailed Student t-tests. Multiple comparisons between the 2 groups were performed by using a 1-way ANOVA followed by the Student-Newman-Keuls test. Differences were considered to be statistically significant at p < 0.05.
Results
Body Weight and Food Intake
Compared to ad libitum mice, FR mice looked healthy. Both body weight and food intake in FR and ad libitum mice progressively increased for 14 days (Figs. 1B and 1C). However, the increase of body weight in FR mice was lower than that in ad libitum mice (Fig. 1B).
D1R Antagonist Inhibited FAA in FR Mice
To investigate the contribution of D1R and D2R to the FAA, we administered D1R antagonist SCH23390 or D2R antagonist raclopride at 1700 h (ZT10) on the 14th day of FR. As shown in Fig. 2A, the change in time courses of locomotor activity revealed that SCH23390 at 10 µg/kg decreased locomotor activity in FR mice by 19% compared with vehicle control. Fig. 2B summarizes the locomotor activity during the 2 h before dark in the mice treated with vehicle or 3 different doses of SCH23390 with FR or ad libitum access, respectively. Compared with vehicle control, pretreatment with the D1R antagonist SCH23390 at 1, 3, or 10 µg/kg significantly decreased FAA by 19% (p < 0.05), 26% (p < 0.05), or 19% (p < 0.01) in FR mice, respectively, whereas pretreatment with the D1R antagonist SCH23390 at 1, 3, or 10 µg/kg did not inhibit the locomotor activity for 2 h (ZT10-ZT12) in the ad libitum mice (Fig. 2B). These findings indicated that the D1R antagonist SCH23390 could attenuate FAA in FR mice.
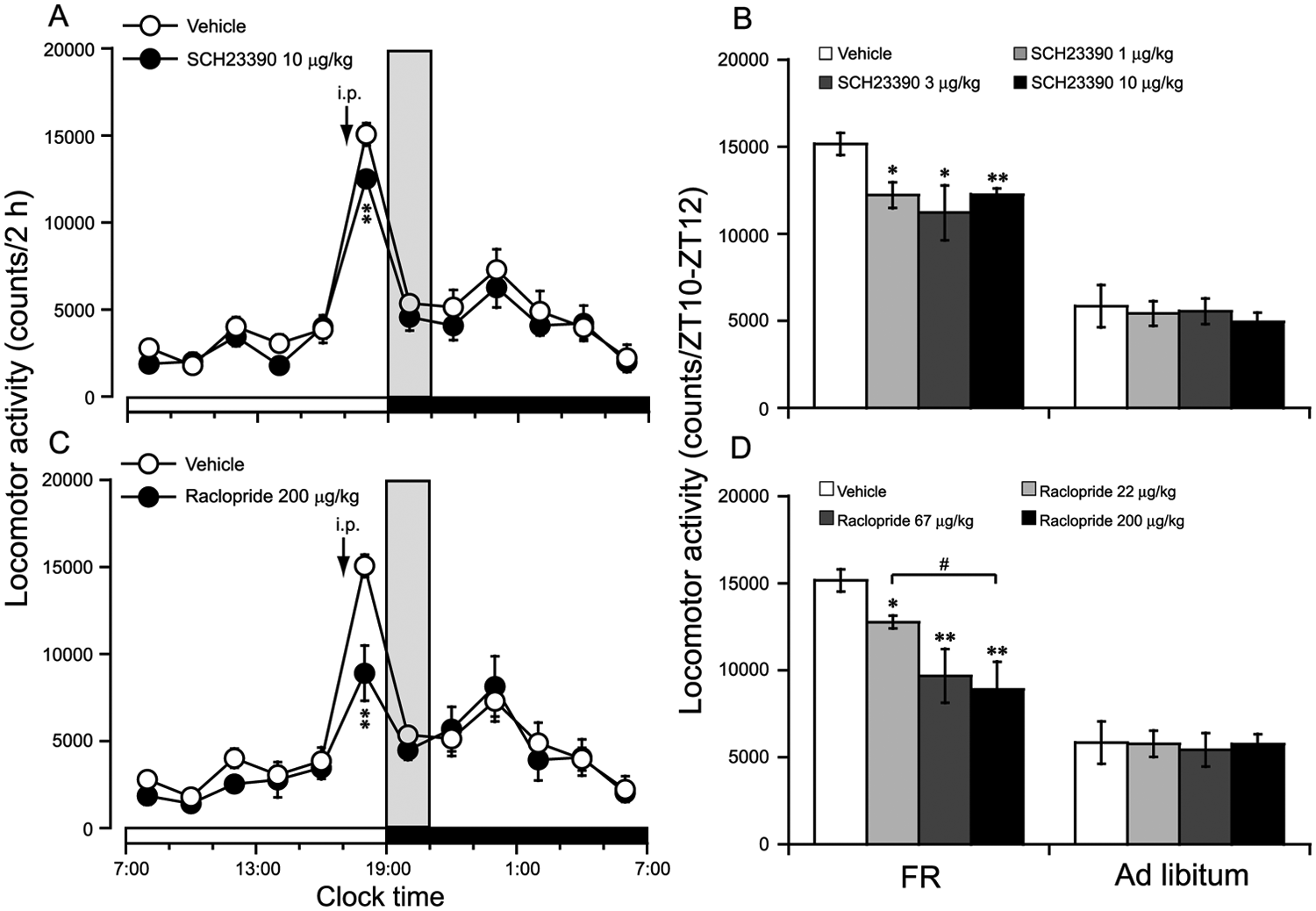
D1R antagonist SCH23390 and D2R antagonist raclopride inhibited FAA in FR mice. FAA was defined as the total locomotor activity for 2 h (ZT10-ZT12) before food access. Time-course changes in the locomotor activity in FR mice treated with 10 µg/kg SCH23390 (A) or 200 µg/kg raclopride (C) at 1700 h (ZT10) on the 14th day of FR, respectively. The dose-effect relationship in the locomotor activity for 2 h (ZT10-ZT12) in the food-restricted and ad libitum–fed mice treated with 1, 3, or 10 µg/kg SCH23390 (B) or 22, 67, or 200 µg/kg raclopride (D), respectively. The time when food was available is indicated by gray shading in the FR mice. *p < 0.05; **p < 0.01 vs. vehicle, n = 6-8.
D2R Antagonist Significantly Inhibited FAA in FR Mice
As shown in Fig. 2C, the change in time courses of locomotor activity revealed that raclopride at 200 µg/kg decreased locomotor activity in FR mice by 41% compared with vehicle control. Fig. 2D summarizes the locomotor activity during the 2 h before dark in the mice treated with vehicle or 3 different doses of raclopride with FR or ad libitum access, respectively. Pretreatment with the D2R antagonist raclopride at 22, 67, or 200 µg/kg did not inhibit the locomotor activity for 2 h (ZT10-ZT12) in the ad libitum mice, whereas pretreatment with raclopride at 22, 67, or 200 µg/kg significantly decreased FAA by 16% (p < 0.05), 36% (p < 0.01), or 41% (p < 0.01) in FR mice, respectively, as compared with vehicle control (Fig. 2D). The inhibitory effect on FAA of raclopride at 200 µg/kg was much stronger than that of raclopride at 22 µg/kg. These findings indicated that the D2R antagonist raclopride significantly inhibited FAA in a dose-dependent manner.
Coadministration of D1R and D2R Antagonist Significantly Inhibited FAA in FR Mice in a Synergistic Manner
To investigate the cooperative or synergistic functional interactions between D1R and D2R in FAA, the D1R antagonists SCH23390 at 10 µg/kg and the D2R antagonists raclopride at 200 µg/kg were coadministered at 1700 h (ZT10) on the 14th day of FR. Figs. 3A and 3B are single-plotted actograms of daily locomotor activity under FR and ad libitum access. Compared with vehicle control, coadministration of SCH23390 and raclopride significantly inhibited FAA in FR mice (Fig. 3A); however, copretreatment with SCH23390 and raclopride did not inhibit locomotor activity for 2 h (ZT10-ZT12) in the ad libitum mice (Fig. 3B). As shown in Fig. 3C, the change in time courses of locomotor activity revealed that coadministration of SCH23390 and raclopride significantly reduced locomotor activity in FR mice by 57% (p < 0.01) compared with vehicle control. Fig. 3D summarizes the locomotor activity for 2 h (ZT10-ZT12) before food access in the mice treated with vehicle or coadministered SCH23390 10 µg/kg and raclopride 200 µg/kg under FR and ad libitum conditions. Copretreatment with SCH23390 and raclopride did not inhibit locomotor activity for 2 h (ZT10-ZT12) in the ad libitum mice, whereas copretreatment with SCH23390 (10 µg/kg) and raclopride (200 µg/kg) significantly reduced FAA in FR mice as compared with vehicle control. These results clearly indicated that D1R and D2R inhibited FAA in a cooperative or synergistic manner.
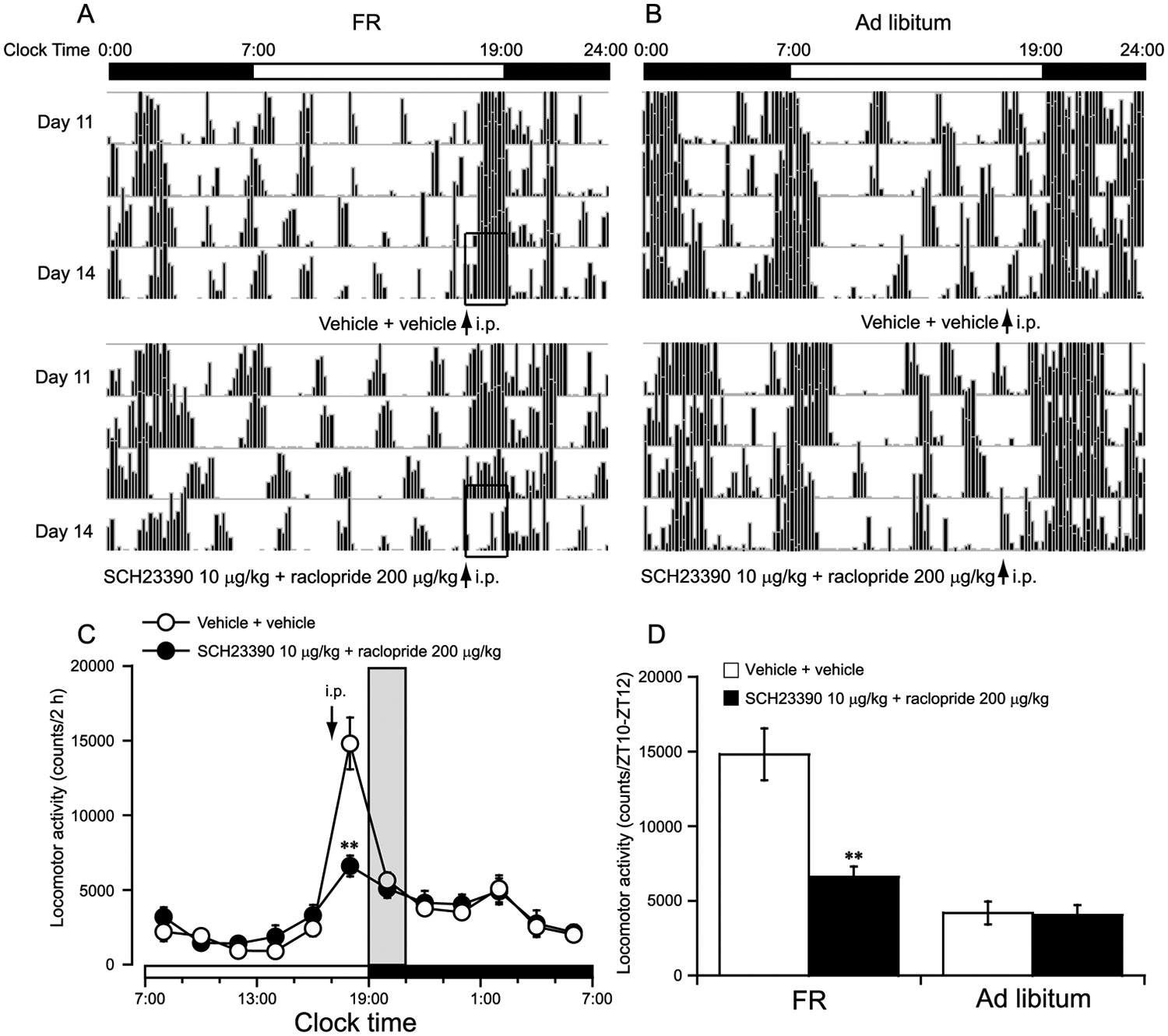
Coadministration of D1R and D2R antagonist significantly inhibited FAA in FR mice in a synergistic manner. FAA was defined as the total locomotor activity for 2 h (ZT10-ZT12) before food access. Representative single-plotted actograms of daily locomotor activity under FR (A) and ad libitum access (B). The open bars indicate the changes of FAA after coadministration of D1R and D2R antagonist or vehicle in FR mice. Time-course changes in the locomotor activity in FR mice coadministered SCH23390 10 µg/kg and raclopride 200 µg/kg (C). The synergistic effect of coadministration of 10 µg/kg SCH23390 and 200 µg/kg raclopride on FAA in FR mice (D). The time when food was available is indicated by gray shading in FR mice. **p < 0.01 vs. vehicle + vehicle,
H1R Antagonist Did Not Inhibit FAA in FR Mice
To investigate the contribution of H1R to FR-induced FAA, we administered the H1R antagonist pyrilamine at 1600 h (ZT9) on the 14th day of FR. Figs. 4A and 4B are single-plotted actograms of daily locomotor activity under FR and ad libitum access. Compared with vehicle control, pretreatment with the H1R antagonist pyrilamine at 10 mg/kg did not significantly inhibit locomotor activity in FR mice (Fig. 4A) and ad libitum mice (Fig. 4B). As shown in Fig. 4C, the change in time courses of locomotor activity revealed that pretreatment with the H1R antagonist pyrilamine at 10 mg/kg did not significantly inhibit locomotor activity in FR mice compared with vehicle control. Fig. 4D summarizes the locomotor activity for 2 h (ZT10-ZT12) before food access in the mice treated with vehicle or 3 different doses of pyrilamine under FR and ad libitum conditions, respectively. Pretreatment with the H1R antagonist pyrilamine at 2.5, 5, or 10 mg/kg did not significantly inhibit FAA in FR mice (Fig. 4D). Similarly, pretreatment with the H1R antagonist pyrilamine at 2.5, 5, or 10 mg/kg did not significantly inhibit the locomotor activity for 2 h (ZT10-ZT12) before food access in the ad libitum mice (Fig. 4D), showing that the H1R antagonist pyrilamine could not attenuate FAA.
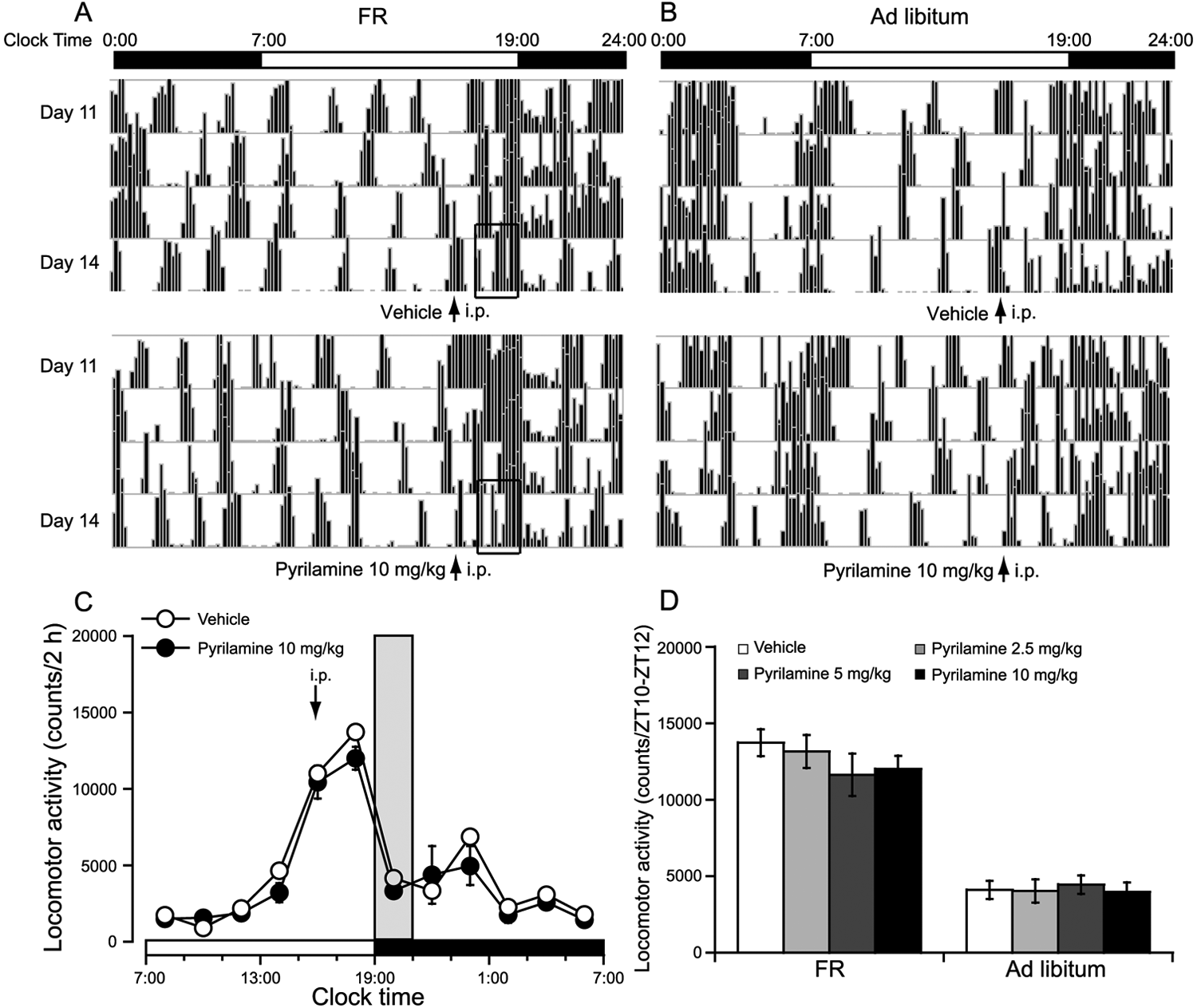
H1R antagonist pyrilamine did not inhibit the FAA in FR mice. FAA was defined as the total locomotor activity for 2 h (ZT10-ZT12) before food access. Representative single-plotted actograms of daily locomotor activity under FR (A) and ad libitum access (B); the open bars indicate changes in FAA after administration of pyrilamine or vehicle in FR mice. Time-course changes (C) and dose-effect relationship (D) in the locomotor activity in FR and ad libitum–fed mice. Pyrilamine 2.5, 5, or 10 mg/kg was administered intraperitoneally at 1600 h (ZT9) on the 14th day of FR, respectively. The time when food was available is indicated by gray shading in FR mice; n = 6-8.
H1R Antagonist Inhibited Spontaneous Locomotor Activity in the Dark Period in Mice Given Ad Libitum Access
To investigate the role of the D1R and D2R antagonists and the H1R antagonists on spontaneous locomotor activity during the dark period in the ad libitum mice, SCH23390 and raclopride together or pyrilamine alone was administered intraperitoneally at 1900 h (ZT12). Figs. 5A and 5B represent single-plotted actograms of daily locomotor activity under FR and ad libitum access. Compared with vehicle control, copretreatment with SCH23390 (10 µg/kg) and raclopride (200 µg/kg) did not significantly inhibit spontaneous locomotor activity in the dark period in the ad libitum mice (Fig. 5A), similar to previous reports in which SCH23390 at 10 µg/kg or raclopride at 200 µg/kg did not alter spontaneous locomotor activity (Benturquia et al., 2008; Rodriguez-Arias et al., 2000). However, when administered at 1900 h (ZT12) in the ad libitum mice, this H1R antagonist at 10 mg/kg significantly inhibited spontaneous locomotor activity in the dark period (Fig. 5B) as compared with vehicle control. As shown in Figs. 5C and 5D, the change in time courses of locomotor activity revealed that copretreatment with SCH23390 (10 µg/kg) and raclopride (200 µg/kg) at 1900 h (ZT12) did not significantly inhibit spontaneous locomotor activity in the dark period in the ad libitum mice (Fig. 5C); however, pretreatment with the H1R antagonist pyrilamine at 10 mg/kg at 1900 h (ZT12) significantly inhibited spontaneous locomotor activity in FR mice as compared with vehicle control, respectively (Fig. 5D). These results showed that SCH23390 at 10 µg/kg and raclopride at 200 µg/kg significantly inhibited FAA without significantly altering spontaneous locomotor activity, whereas the H1R antagonist pyrilamine significantly inhibited spontaneous locomotor activity without significantly altering FAA.
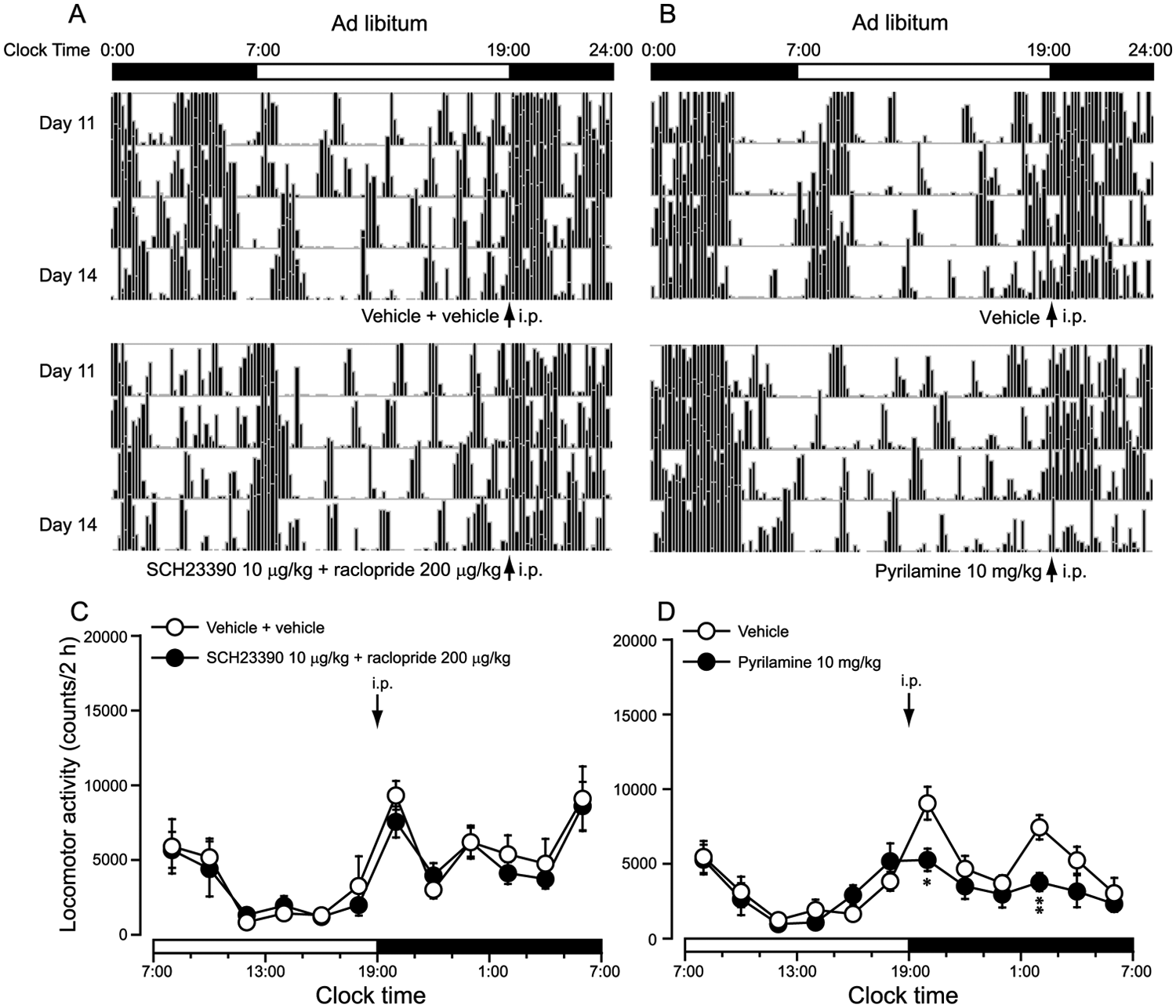
The spontaneous locomotor activity during the dark period in ad libitum–fed mice was inhibited by H1R antagonist pyrilamine but not by coadministration of SCH23390 and raclopride. Representative single-plotted actograms of daily locomotor activity in mice coadministered SCH23390 at 10 µg/kg and raclopride at 200 µg/kg (A) or 10 mg/kg pyrilamine (B) under ad libitum conditions. Time-course changes in the locomotor activity in mice coadministered 10 µg/kg SCH23390 and 200 µg/kg raclopride (C) or 10 mg/kg pyrilamine (D) at 1900 h (ZT12). *p < 0.05; **p < 0.01 vs. vehicle, n = 6-8.
Dopamine and Its Metabolites Increased in the Striatum and/or Midbrain in FR Mice
Pharmacological studies have showed that both D1R and D2R are involved in FAA. Therefore, we further determined the levels of catecholamine, 5-HT, and their metabolites in various brain regions of mice. As shown in Fig. 6A, the level of dopamine in the striatum of FR mice pretreated with vehicle was significantly increased by 32% and 33%, respectively, as compared with the ad libitum mice pretreated with vehicle or SCH23390 and raclopride. The dopamine metabolite DOPAC in the striatum also significantly increased in FR mice as compared with the ad libitum mice pretreated with vehicle. In the midbrain (Fig. 6B), the levels of DOPAC and HVA were significantly elevated in FR mice as compared with the ad libitum mice, whereas there were no differences observed in dopamine concentrations among groups. With respect to dopamine turnover, an index of neuronal activity calculated as a ratio of (DOPAC+HVA)/DA in the midbrain was significantly increased in FR mice as compared with the ad libitum mice pretreated with vehicle. However, levels of norepinephrine, DA, DOPAC, HVA, 5-HT, and 5-HIAA were not altered in the cortex, hippocampus, thalamus, hypothalamus, or medulla oblongata (data not shown). Taken together, these findings suggest that FR increased dopaminergic transmission, which might be involved in FR-induced FAA.
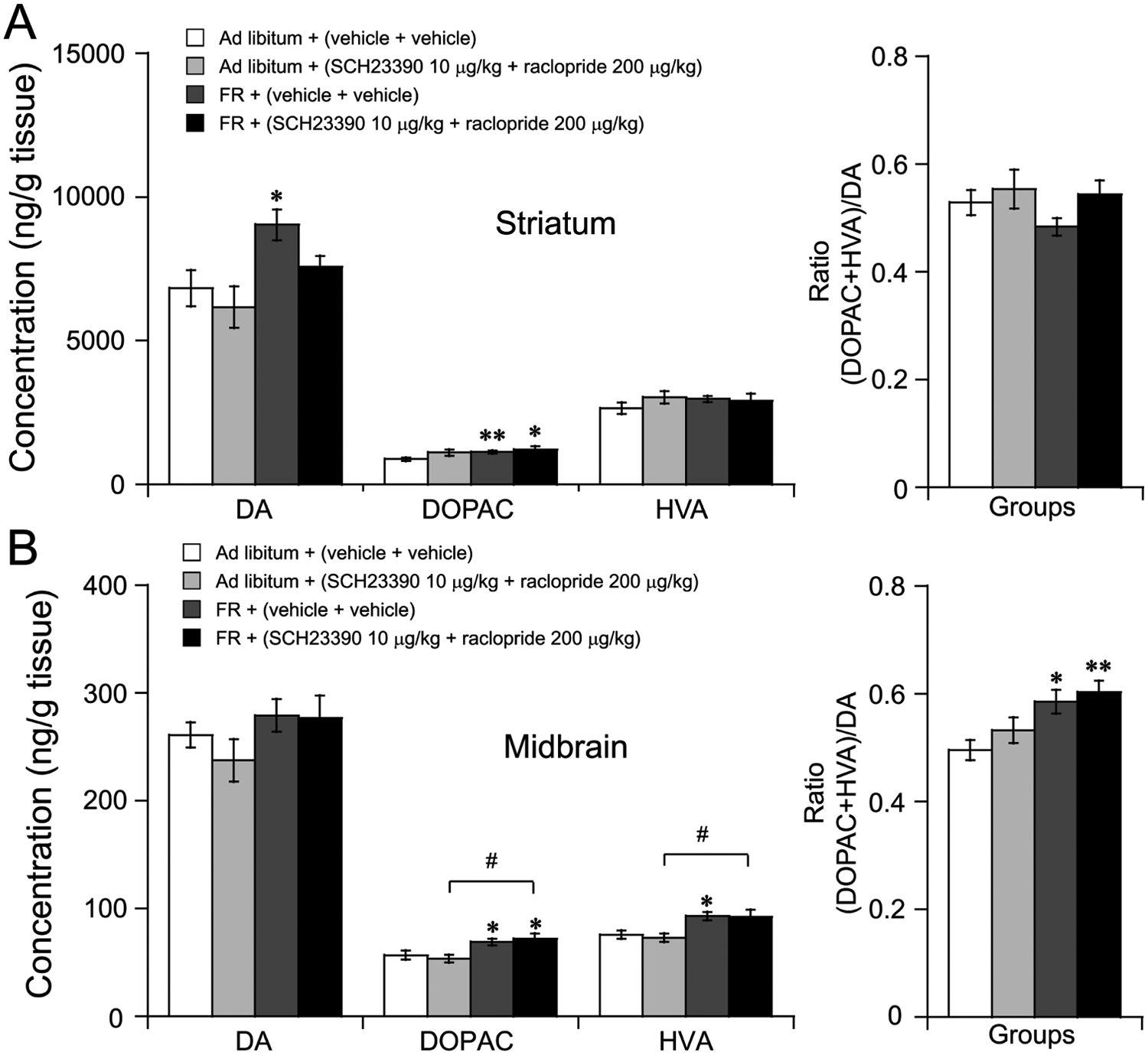
Dopamine and its metabolites’ contents were increased in striatum and midbrain in FR mice. Animals were killed 2 h after administration of vehicle or SCH23390 and raclopride. Brains were removed and frozen immediately. The levels of dopamine and its metabolites were measured by HPLC-ECD. Changes in concentrations of dopamine (DA), DOPAC, HVA, and the ratio of (DOPAC+HVA)/DA in the striatum (A) and midbrain (B). *p < 0.05; **p < 0.01 vs. ad libitum + (vehicle+vehicle), n = 6-8.
Discussion
The present results showed that the dopaminergic system including dopamine, D1R, and D2R, but not histaminergic H1R, mediated FAA induced by FR.
D1R and D2R are the most widely and abundantly expressed receptors for DA in the brain (Monti and Monti, 2007). D1R is a postsynaptic receptor coupled to adenylate cyclase, and its stimulation facilitates the activity of the enzyme. D2R is expressed both as presynaptic autoreceptors on dopamine neurons and terminals and as postsynaptic receptors on target cells, and the D2R subfamily is coupled to the inhibition of adenylate cyclase (Monti and Monti, 2007; Usiello et al., 2000). D1R and D2R play an important role in control of locomotor and motivated behavior (Kobayashi et al., 2004; Tomiyama et al., 2001). Activation of postsynaptic D1R or D2R or blockade of presynaptic D2R increased locomotor activity, whereas activation of D2 autoreceptors or blockade of postsynaptic D1R or D2R inhibited spontaneous locomotor activity (Costall et al., 1981; Rouillon et al., 2007; Skirboll et al., 1979). Selective D1R antagonist SCH23390 and D2R antagonist raclopride blocked morphine- or nerve growth factor-induced hyperactivity, and the effect of D2R antagonist appears to be stronger than that of D1R antagonist (Kobayashi et al., 1997; Rodriguez-Arias et al., 2000). These results support our present findings that D1R and D2R are involved in FAA, with D2R being more important than D1R. In addition, activation of D2R has been demonstrated to regulate expression of clock genes in the striatum (Hood et al., 2010; Imbesi et al., 2009), a region important for motor control, food feeding, and reward, suggesting that the dopaminergic system in the striatum might be involved in circadian food anticipation. Although previous studies showed that mutant mice lacking clock genes exhibited normal FAA (Pendergast et al., 2009; Storch and Weitz, 2009), the existence of compensatory mechanism in these mice may address this reason. These mice lack clock genes throughout the brain and body, making it impossible to localize the actions of clock genes or to determine whether the effects are due to disruptions of circadian rhythms or simply to the lack of clock genes (Amir and Stewart, 2009). Contrary to our results, Mistlberger and Mumby (1992) found no effects of dopamine receptor inhibition and large dopamine lesions of nucleus accumbens on FAA in rats. In their studies, dopamine receptor antagonist was given 30 min prior to feeding time; therefore, it did not inhibit total locomotor activity for 2-3 hours (FAA) before food access. In the present study, dopamine receptor antagonist was given 2 hours prior to feeding time and increased total locomotor activity for 2 hours before food access was significantly inhibited. In the studies by Mistlberger and Mumby, large nucleus accumbens lesions did not inhibit FAA; however, we determined the levels of dopamine and its metabolite DOPAC in the striatum and the midbrain but not lesioned nucleus accumbens. Our results showed that the levels of dopamine and its metabolite DOPAC in the striatum and dopamine metabolites in the midbrain were significantly increased during FAA. These results are similar to previous reports that food deprivation or restriction increases locomotor activity and levels of dopamine and its metabolite in the nucleus accumbens (Heffner et al., 1980; McCullough and Salamone, 1992). Another study showed that lesions of the core of nucleus accumbens attenuated FAA, whereas lesions of the nucleus accumbens shell had no effects (Mendoza et al., 2005). Therefore, the difference in administration time of dopamine receptor antagonists and lesion regions might result in the discrepancy between our findings and those of Mistlberger and Mumby (1992). Taken together, our results indicated that FAA could be regulated by the dopaminergic system, including dopamine, D1R, and D2R.
Concomitant activation of D1R and D2R is crucial in the control of diverse physiological function (Aizman et al., 2000). Mice that are double-KO for D1R and D2R are severely growth retarded, with altered feeding behavior and a dysfunctional gastrointestinal system, suggesting an interrelationship between D1 and D2 receptor–mediated regulation of motor activity, food intake, and gastrointestinal functions (Kobayashi et al., 2004). Pretreatment of D2R KO mice with a D1R antagonist completely abolished the arousal effects of modafinil (Qu et al., 2008). In the present study, coadministration of SCH23390 and raclopride was more potent in inhibiting FAA induced by FR as compared with using either alone. However, the spontaneous locomotor activity in the dark period in the ad libitum mice was not inhibited by copretreatment with SCH23390 and raclopride. These results indicated the presence of cooperative or synergistic functional interactions between D1R and D2R in regulation of FAA.
The histaminergic system is thought to modulate many functions of the hypothalamus that exhibit circadian and other rhythms, such as arousal state, locomotor activity, and feeding (Huang et al., 2001; Inoue et al., 1996). Mutant mice lacking H1R demonstrated decreased spontaneous locomotor activity and exploratory behavior (Inoue et al., 1996). The H1R antagonist pyrilamine significantly inhibited spontaneous locomotor activity (Lozeva et al., 2000). These results suggested that histamine increases spontaneous locomotor activity through H1R in mice given ad libitum access. Consistent with these results, our study also showed that the H1R antagonist pyrilamine at 10 mg/kg decreased the spontaneous locomotor activity during the dark period in the ad libitum mice. However, during the restricted feeding schedule, pretreatment with the H1R antagonist pyrilamine did not significantly inhibit FAA, indicating that the role for the histaminergic H1R in FR-induced FAA is different from that for spontaneous locomotor activity. Several studies have shown that the food anticipatory–induced signals from the food-entrainable oscillator are transmitted downstream to the arousal related areas, such as orexin neurons in the perifornical area (PeF) (Akiyama et al., 2004; Kaur et al., 2008). The TMN receives direct projections from the orexinergic neurons and may help regulate arousal (Huang et al., 2001). Angeles-Castellanos et al. (2004) reported that the TMN might be involved in the arousal state during food anticipation. Furthermore, the activation of the TMN was associated primarily with arousal during feeding-related motivated behavior (Valdes et al., 2005). Although histaminergic neurons of the TMN show activation of c-Fos protein during the food anticipatory phase in FR rats (Inzunza et al., 2000; Meynard et al., 2005), these data were based on the sampling of a single horizontal section of the TMN; therefore, their interpretation must be addressed with limitations, especially for the TMN, which contains many neuronal phenotypes. Collectively, our results suggested that the activation of the TMN might mediate only the nonspecific arousal state but not increased locomotor activity in the FAA-related activity.
Several lines of evidence indicate that FAA might not rely on a single nucleus but rather might be a distributed system consisting of interacting nuclei (Poulin and Timofeeva, 2008; Storch and Weitz, 2009). This distributed network may ensure the maintenance of vital physiological functions and behavior directed toward food seeking, feeding, and adequate preservation of energy resources. Therefore, it is important that such essential functions be maintained by redundant neurotransmission in a distributed network that functions harmoniously as a whole but also has a capacity for compensation and plasticity. D1R and D2R play an important role in the maintenance of many physiological functions and behavior, such as feeding and food- or drug-seeking behaviors (Ball et al., 2011; Sun and Rebec, 2005). Double-KO mice for D1R and D2R do not survive to the second or third week after birth due to severely growth-retarded with altered feeding behavior (Kobayashi et al., 2004). Adequate preservation of energy resources is closely related with feeding (Currie et al., 2011). Several studies suggest the influence of gastrointestinal tract-elicited signals as a driving force for FAA (LeSauter et al., 2009; Morgado et al., 2011). D1R and D2R have been reported to mediate regulation of gastrointestinal functions (Cooper and Barber, 1990; Kobayashi et al., 2004), suggesting that gastrointestinal D1 and D2 receptors signal might be one of factors responsible for modulating FAA. Because D1R and D2R are the most abundant and widely expressed DA receptor in the brain (Monti and Monti, 2007), these receptor antagonists might decrease FAA mainly through the central nervous system. Although our findings do not address the whole distributed network that drives FAA, they provide information on the role of dopamine and D1R and D2R as components of the distributed system for the food-entrainable system.
In other studies, food is given in the middle of the light phase in order to observe FAA (Moriya et al., 2009; Storch and Weitz, 2009). However, the manipulation of removing and adding food during the light phase interferes with the activity of the light-dark cycle because during the light phase, rodents spend most of the time resting. In the present study, in order to prevent the manipulation of removing and adding food from interfering with the light-dark cycle during the light periods, we chose the restricted feeding regime: The food was given at ZT 12 (lights off) and withdrawn at ZT 14. When food is provided during the day, the increased nocturnal activity is not altered by FAA (Moriya et al., 2009; Storch and Weitz, 2009). In present studies, food was provided during the first 2 hours of the dark phase; once food was accessed, the nocturnal activity counts would temporarily decreased. However, the total nocturnal activity counts were more than the total diurnal activity counts, suggesting that the increased nocturnal activity still exists in the FR mice.
Herein we demonstrated that the levels of dopamine in the striatum and dopamine metabolites in the midbrain were significantly increased during FR-induced FAA. Pretreatment with the dopaminergic D1R or D2R antagonist significantly inhibited FAA, and there appear to be cooperative or synergistic effects between D1R and D2R. However, pretreatment with histaminergic H1R antagonist did not significantly inhibit FAA in FR mice, although it decreased the spontaneous locomotor activity during the dark period in the ad libitum mice. In conclusion, these findings indicated that the dopaminergic system, but not histaminergic H1R, is involved in FAA induced by FR.
Footnotes
Acknowledgements
This study was supported in part by grants-in-aid for scientific research from the National Basic Research Program of China Grants (2009CB5220004, 2011CB711000), National Natural Science Foundation of China (30970955, 30901797, 31070957, 31171010, 31121061), Shanghai Committee of Science and Technology (09JC1402500, 10XD1400400, 10441901600), Shanghai Leading Academic Discipline Project (B119), Ph.D. Programs Foundation of Ministry of Education of China (20110071110033), and China National Science and Technology Major Project for Drug Discovery (2009ZX09303-006).
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.