
Abstract
Intertidal species have both circadian and circatidal clocks. Although the behavioral evidence for these oscillators is more than 5 decades old, virtually nothing is known about their molecular clockwork. Pigment-dispersing hormones (PDHs) were originally described in crustaceans. Their insect homologs, pigment-dispersing factors (PDFs), have a prominent role as clock output and synchronizing signals released from clock neurons. We show that gene duplication in crabs has led to two PDH genes (β-pdh-I and β-pdh-II). Phylogenetically, β-pdh-I is more closely related to insect pdf than to β-pdh-II, and we hypothesized that β-PDH-I may represent a canonical clock output signal. Accordingly, β-PDH-I expression in the brain of the intertidal crab Cancer productus is similar to that of PDF in Drosophila melanogaster, and neurons that express PDH-I also show CYCLE-like immunoreactivity. Using D. melanogaster pdf-null mutants (pdf01) as a heterologous system, we show that β-pdh-I is indistinguishable from pdf in its ability to rescue the mutant arrhythmic phenotype, but β-pdh-II fails to restore the wild-type phenotype. Application of the three peptides to explanted brains shows that PDF and β-PDH-I are equally effective in inducing the signal transduction cascade of the PDF receptor, but β-PDH-II fails to induce a normal cascade. Our results represent the first functional characterization of a putative molecular clock output in an intertidal species and may provide a critical step towards the characterization of molecular components of biological clocks in intertidal organisms.
Virtually all living organisms are exposed to cyclic changes in their environment like the alternation between day and night or the ebb and flow of tidal waters. The ability to anticipate these changes has obvious adaptive value as indicated by the widespread distribution of biological clocks. These endogenous pacemakers oscillate with a period close to that of the environmental cycle they are synchronized to and regulate the timing of physiological and behavioral processes. Thus, circadian clocks oscillate with approximately 24-hour periods and are usually entrained by the 24-hour light-dark (LD) cycle, while circatidal clocks oscillate with periods close to the tidal period and are synchronized by environmental changes associated with high and low tides (de la Iglesia and Hsu, 2010; Palmer, 2000; Wilcockson and Zhang, 2008).
The evolutionary conservation of the mechanisms underlying circadian rhythmicity is well established and is underscored by the fact that gene homologs, whose products constitute the molecular circadian clockwork, are shared by all animals studied so far (Bell-Pedersen et al., 2005). Interestingly, animals living in the intertidal zone are exposed to both the solar day and the tidal cycle, and behavioral studies have established the existence of both circadian and circatidal pacemakers within the same individual. Nevertheless, the molecular and neural mechanisms constituting these two biological clocks and whether they share common clockwork components remain unknown (de la Iglesia and Hsu, 2010; Naylor, 1996; Palmer, 2000; Wilcockson and Zhang, 2008).
Pigment-dispersing hormones (PDHs) are a broadly conserved neuropeptide family in both crustaceans and insects (Rao and Riehm, 1993). Several PDH genes have been described in crustaceans, leading to the subdivision of α-PDH and β-PDH forms (Rao, 2001). These peptides induce pigment dispersion in the retina and melanophores of several crustacean species (Fernlund, 1976; Rao et al., 1987; Rao et al., 1985). Multiple isoforms of β-PDH have been documented within a single species by sequencing purified peptides, molecular cloning, or mass spectrum analysis (Desmoucelles-Carette et al., 1996; Fu et al., 2005b; Klein et al., 1994; Rao and Riehm, 1989; Yang et al., 1999); yet, the differential role of these peptides, if any, remains to be determined. Pigment-dispersing factor (PDF) constitutes the β-PDH ortholog in insects (Helfrich-Forster and Homberg, 1993). In Drosophila melanogaster (Drosophila), PDF has been extensively characterized as a critical output molecule of circadian pacemaker neurons and as a key coupling signal among them (Frenkel and Ceriani, 2011; Lin et al., 2004; Peng et al., 2003; Renn et al., 1999). Whether some of the β-PDH orthologs could serve a similar circadian role in crustaceans remains unknown.
We have previously described two β-PDHs in the intertidal crab Cancer productus (Hsu et al., 2008). These two β-PDHs produce mature peptides differing in 6 of 18 amino acids (the Canpr-β-PDH-I sequence is NSELINSILGLPKVMNDA, and the Canpr-β-PDH-II is NSELINSLLGLSRLMNEA); Drosophila PDF (NSELINSLLSLPKNMNDA) differs in 3 and 5 amino acids with Canpr-β-PDH-I and II, respectively. β-PDH-II is expressed in the neurosecretory sinus gland, the main neurosecretory organ in decapods, whereas β-PDH-I is expressed in several neuronal groups within the eye stalk but also expressed in neurons within the supraesophageal ganglion (the brain) (Hsu et al., 2008). This latter discrete expression pattern suggests a nonpigment-dispersing function for β-PDH-I.
Based on a phylogenetic tree constructed with most known β-PDHs and PDFs (Fig. 1) and the spatial distribution of C. productus β-PDH-I (Hsu et al., 2008) and Drosophila PDF, we hypothesize that, although β-PDH-I may also retain the ability to disperse pigments, it may represent a functional ortholog of Drosophila PDF, acting as a neurotransmitter/modulator neuropeptide signaling within a putative clock-neuron network. Determining if this is the case would represent an important step towards identifying molecular and neural clock components in an intertidal organism. Because functional analysis is not possible in the crab, we studied the ability of each C. productus β-PDH as a modulator of circadian behavior using Drosophila as a heterologous system. By expressing either β-PDH-I or β-PDH-II in the PDF circuit of Drosophila pdf01-null mutants, we demonstrate that only β-PDH-I can act as a bona fide PDF functional ortholog and lead to a full rescue of the pdf01 mutation, suggesting β-PDH-I as a putative molecular clock output in intertidal crustaceans.
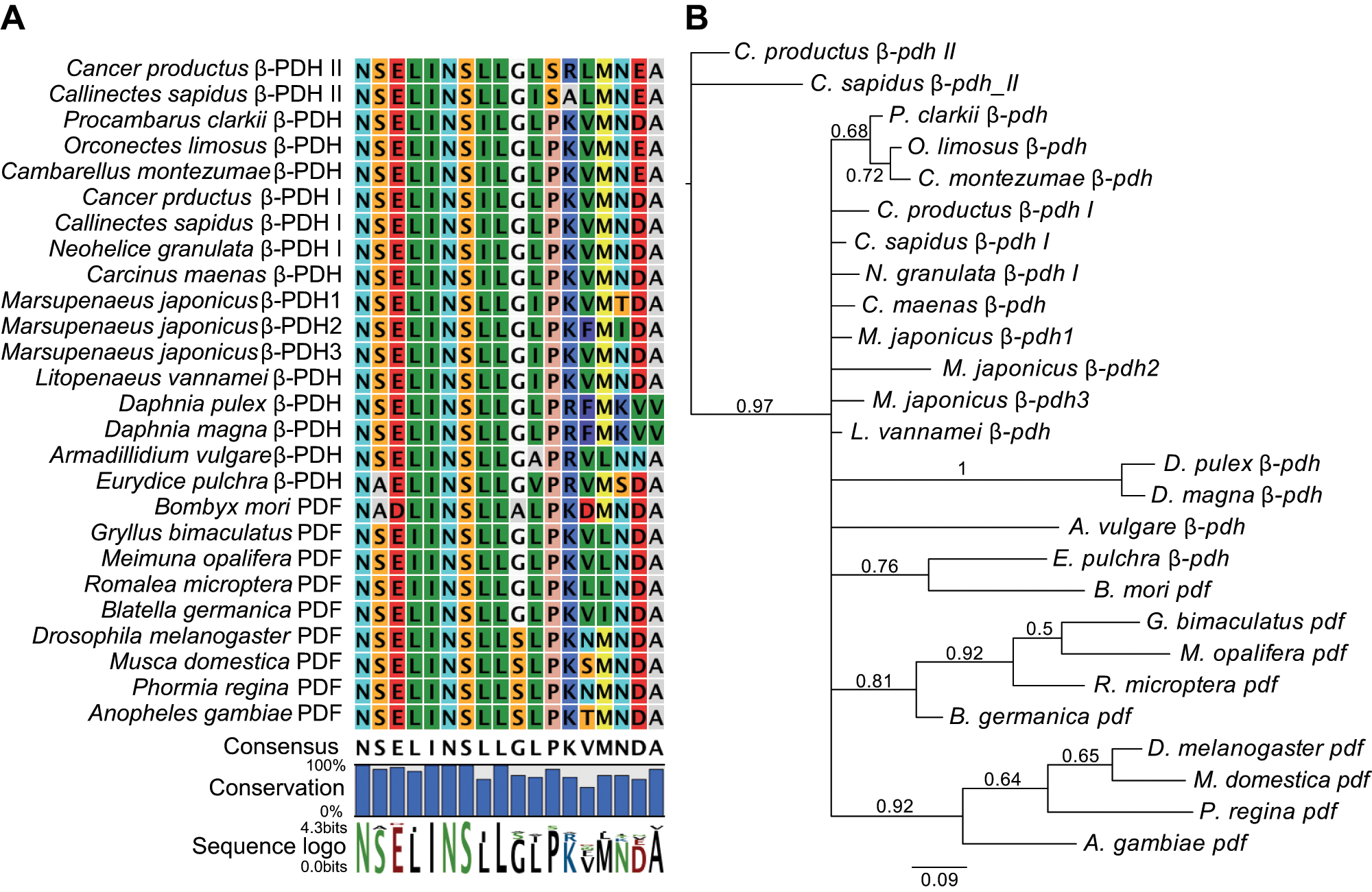
Crustacean β-pdh-Is form a monophyletic group with insect pdfs. (A) Alignment of all β-PDH and PDF peptides identified so far. Pro12 is conserved in all family members except in the crustacean β-PDH-IIs so far identified, in which this hydrophobic residue is replaced by a hydrophilic Ser. Consensus sequence and conservation are shown below. Lettering size in the sequence logo represents the most consensual amino acid. The gene accession numbers are detailed in Supplementary Table S1. (B) Genealogy for the coding sequences of all peptides shown in A, based on a Bayesian phylogenetic analysis. Posterior probabilities ≥0.50 are shown on branches. Scale bar represents the number of nucleotide changes expected per site.
Materials and Methods
Phylogenetic Analysis
Sequences for β-PDHs and PDFs were searched in the “Nucleotide collection (nr/nt)” database within NCBI BLAST (National Institutes of Health, Bethesda, MD). The β-PDH coding region of Canpr-β-pdh-I, Canpr-β-pdh-II, and the PDF coding region of Drosophila pdf were used as query sequences. The corresponding β-PDH or PDF peptide sequences were aligned using CLC DNA Workbench version 6.0.2 (CLC Bio, Germantown, MD) to generate Figure 1A. Nucleotide sequences for the peptides’ coding region were aligned with MacVector version 11.1.2 (MacVector, Cary, NC).
A Bayesian phylogenetic tree was generated using MrBayes version 3.1.2 (http://mrbayes.sourceforge.net/index.php) (Huelsenbeck and Ronquist, 2001; Ronquist and Huelsenbeck, 2003). The Akaike information criterion in jModeltest v2.2 (Dr. David Posada, Universidad de Vigo, Vigo, Spain; Posada, 2008) was used to determine the best-fit nucleotide substitution (HKY+I+Γ). The Bayesian analysis was run with default heating values for 1 million generations (sampling every 100 generations). The first 1000 samples were discarded as burn-in, and the remaining 9000 samples were combined to produce a 50% majority rule consensus tree. This tree was imported into FigTree version 1.3.1 (Dr. Andrew Rambaut, University of Edinburgh, Edingurgh, UK) to generate the tree shown in Figure 1B.
C. productus Whole-Brain Immunohistochemistry
Adult C. productus crabs were collected at various locations throughout the San Juan Islands archipelago of Washington state and housed under a 12:12 LD cycle. Brains were dissected and postfixed overnight in 4% paraformaldehyde in sodium phosphate buffer, and whole-brain immunohistochemistry was performed as previously described (Hsu et al., 2008). The anti-PDH rabbit polyclonal antibody was generated against synthetic β-PDH (NSELINSILGLPKVMNDAamide) conjugated to bovine thyroglobulin via glutaraldehyde by Dr. K. Rao (University of West Florida, Pensacola, FL) (Dircksen et al., 1987). The anti-CYC guinea pig antibody (a kind gift of Dr. Isaac Edery) was raised against the whole protein synthesized in bacterial expression vectors (Bae et al., 2000). Preadsorption controls could not be done because the expressed full Drosophila CYC protein is not available. Cross-reactivity between the anti-PDH and the anti-CYC antibodies, or their secondary antibodies, was discarded by single-labeling immunohistochemistry with each antibody (not shown).
Fly Stocks
The control strain used during the generation of novel transgenic lines as well as in behavioral analysis was w1118. The original y–w–, pdf01 stock (Renn et al., 1999) was backcrossed 5 times to the control (w1118). pdf-GAL4, UAS-pdf (Renn et al., 1999), and the newly generated UAS-pdh transgenic lines were crossed into a pdf01 background. Male flies of the genotype w;Clock[856 (8.2/2)]-Gal4/UAS-Epac-1-camps(50A) were used in all live imaging sessions (Gummadova et al., 2009; Shafer et al., 2008). To test for the necessity of the PDF receptor, we used the PDF receptor mutant han5304 (Hyun et al., 2005).
Generation of UAS-pdh Transgenic Lines
The open reading frames of β-PDH-I and IIa were amplified from constructs described in Hsu et al. (2008) with the following primers: β-PDH-IEcoRI: GAATTCATGCGTAGCGCCGTGGTCG, β-PDH-IKpnI: GGTACCttatctcctgccggcgtcgTTC, β-PDH-IIaEcoRI: GAATTCATGCGCACCGGTATGGTTATGACCGTGGTG, and β-PDH-IIaKpnI: GGTACCtcatctcctgccggcctcg. PCR products were cloned into the pUAS vector and injected into w1118 embryos with pUChpsD2-3 helper plasmid (Misra and Rio, 1990). Several independent transformant lines were obtained on the second and third chromosomes. To minimize GAL4 toxicity (Rezaval et al., 2007), only one copy of the pdf-GAL4 construct was employed.
Drosophila Whole-Brain Immunohistochemistry
Adult heads were cut, while flies were pinned down to a Sylgard dish (Corning, Lowel, MA) in phosphate-buffered saline, and whole-brain immunohistochemistry was performed as previously described (Fernandez et al., 2008). The anti-PDH rabbit polyclonal antibody (Dircksen et al., 1987) was diluted 1/1000, and the secondary antibody used was Cy3-conjugated donkey anti-rabbit (Jackson ImmunoResearch, West Grove, PA) diluted 1/250. Images were acquired with a LSM 510 Meta Zeiss laser scanning confocal microscope (Zeiss, Jena, Germany). The images were adjusted for levels, brightness, and contrast using ImageJ (National Institutes of Health) software.
Quantitative Real-Time PCR
Total RNA isolation from fly head extracts was performed using Trizol (Invitrogen, Carlsbad, CA), and SuperScript II (Invitrogen) was used for reverse transcription following the manufacturer’s instructions. The real-time assays were conducted in the Stratagene Mx3000P QPCR System (La Jolla, CA) using SYBR green as the detection system and Taq platinum (Invitrogen). The primers were designed using Primer3 (available online at http://frodo.wi.mit.edu/primer3/), β-PDH-I forward primer: GGGCAGGATCTCAAGTACCA, β-PDH-I reverse primer: GTTCATCACCTTGGGCAGTC, β-PDH-II forward primer: ATGGTTATGACCGTGGTGGT, and β-PDH-II reverse primer: AGTCTGGAGAGTCCGAGCAG. mRNA levels were assessed from three independent RNA extractions; three technical replicates were performed on each sample. The ΔΔ Ct method was used to calculate the results, and only primer pairs with efficiency between 90% and 110% were used. Two independent reference genes (tubulin and RpL49) were used for normalization, both showing similar results.
Behavioral Analyses
Newly eclosed male flies were placed in glass tubes and monitored for activity with commercially available activity monitors (TriKinetics, Waltham, MA). Fly activity was recorded under LD conditions for 4 days, and then flies were released into constant DD for 10 days. Rhythmicity and period were estimated using the ClockLab software (Actimetrics, Evanston, IL) during the DD phase; data from the first 2 days were not taken into account for period and rhythmicity analysis. Flies with a single peak over the significance line in a χ2 analysis (p < 0.05) were scored as rhythmic, which was confirmed by visual inspection of the actograms. All flies used for behavioral analysis were kept at 25 °C and 12-hour:12-hour light-dark regimen throughout development; behavioral assays were also carried out at 25 °C. The experiment was conducted 3 times, with more than 20 flies per genotype in each experiment.
To construct the average activity plots, each 30-minute activity bin was normalized to the total activity of the corresponding animal per day, the mean value for each time point was obtained averaging data from 4 consecutive days for each fly, and the average for all the flies of a given genotype was obtained. The anticipatory phase scores (APSs) for both morning and evening activity were calculated as previously described (Harrisingh et al., 2007).
Ex Vivo PDFR Activity Imaging
Flies were anesthetized on ice, and brains were dissected in cold Ringer’s solution and mounted under HL3 saline (Stewart et al., 1994), contained by a Petri dish insert (Bioscience Tools, San Diego, CA), and placed within a 35-mm Falcon dish (Becton Dickson Labware, Franklin Lakes, NJ). The sLNv and LNd were identified by their basal Epac-camps fluorescence and distinguished by their locations and projection patterns using an epifluorescent illumination for YFP. FRET imaging was performed with an Olympus FV 1000 laser scanning confocal microscope (Olympus, Tokyo, Japan), through a 20x (UMPlanFL N) N.A. 0.5 water immersion objective. Using the FluoView (Olympus) software, regions of interest were selected over the soma of single neurons, and frames were scanned at a frequency of 1 Hz for 5 minutes.
Frames were scanned with a 440-nm laser using a DM405-440/515 dichroic mirror (Olympus). CFP and YFP emission was separated by means of a SDM510 dichroic mirror (Olympus). For each neuron of interest, a spillover-corrected inverse FRET ratio was determined for each time point. Inverting the FRET ratio makes the ratio directly proportional to relative cAMP levels (Tomchik and Davis, 2009). The proportion of CFP fluorescence spillover in to the YFP channel was determined to be 0.444 on our imaging system. Inverse FRET was calculated as follows:
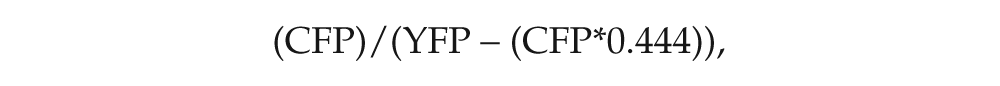
where CFP and YFP are the CFP and YPF emission intensities, and 0.444 is the proportion of CFP spillover into the YFP channel. Inverse FRET traces were filtered with a 10-point moving average and normalized to the value of the first time point (=1.0). Average traces were made by taking the average ratio and standard error for each time point for each treatment, neuronal type, and genotype.
β-PDH-I and II were synthesized by GenScript (Piscataway, NJ). Drosophila PDF was synthesized by PolyPeptide Group (San Diego, CA). The peptides were dissolved directly in DMSO and brought to volume with HL3 saline as needed. Final concentration was 10–4 M in 0.1% DMSO; 0.1% DMSO served as vehicle control. For the vehicle, β-PDH-I and β-PDH-II treatments, we sampled from 5 to 6 brains totaling 15 to 22 neurons per treatment and neuron type. For Drosophila PDF, whose response is well characterized (Shafer et al., 2008), we sampled from 2 to 3 brains totaling 8 to 10 neurons per neuron type.
Results
β-PDH-I and II Are the Results of Crab-Specific Gene Duplication
Until now, β-PDH-II homologs have only been identified by cloning in two crab (Brachyura) species. Although Ohira et al. (2002) named one of the β-PDHs identified in the shrimp Marsupenaeus japonicus “β-PDH-II,” this peptide lacks the characteristic β-PDH-II Ser12 and instead has Pro12, which is shared by all the remaining members of the family (Fig. 1A). This hydrophobic (Pro) to hydrophilic (Ser) amino acid change is unique to the β-PDH-IIs identified in Callinectes sapidus (Klein et al., 1994) and C. productus (Hsu et al., 2008) as well as to the Cancer borealis β-PDH-II, which was identified only by mass spectrometry but not by cloning (Fu et al., 2005a). We therefore renamed the three M. japonicus β-PDHs with Arabic instead of Roman numerals. That β-PDH-IIs from crabs represent a unique set of peptides is clearly shown by the Bayesian phylogenetic analysis of the coding nucleotide sequence for the different β-PDHs and PDFs (Fig. 1B). Using the Canpr-β-pdh-II sequence as the outgroup, the only sequence that was unresolved from it was C. sapidus β-pdh-II. This β-pdh-II cluster constituted a single clade with a posterior probability of 1.0 in every Bayesian phylogenetic analysis we ran, whether analyzed with or without Canpr-β-pdh-II as the outgroup. Importantly, all the remaining family members, including the insect pdfs and all crustacean β-pdhs, form a single clade that is well supported (posterior probability = 0.96). This clade also includes Canpr-β-pdh-I and C. sapidus-β-pdh-I, clearly indicating that β-PDH-I, in the two species in which β-PDH-II was also identified by cloning, is phylogenetically more closely related to all PDFs, including Drosophila PDF.
β-PDH-I Is Expressed in Neurons Expressing a Putative Clock Protein Homolog
We hypothesized that the differential distribution of the β-PDHs in C. productus (Hsu et al., 2008) may underlie different functions: while β-PDH-II may act as a pigment-dispersing neurohemal peptide, β-PDH-I could serve a neurotransmitter/modulator function in the brain similar to the function of PDF in Drosophila. In an attempt to identify putative clock neurons in the crab brain, we used an antibody developed against Drosophila CYCLE (dCYC), a fly core clock transcription factor (Rutila et al., 1998). We identified a cluster of 2 to 3 CYC-like–expressing neurons within neuronal cluster 6 of the brain of C. productus. Double immunolabeling with anti-dCYC and anti-β-PDH determined that a subset of these CYC-like–immunopositive neurons expressed the β-PDH-I peptide (Fig. 2) because previous in situ hybridization studies have determined that only β-pdh-I is expressed in the brain (Hsu et al., 2008). CYC-like staining appeared to be cytoplasmic, but staining in the crab Carcinus maenas at other zeitgeber times (ZTs) showed more intense nuclear staining (A. Christie, personal communication). Although the possibility exists that the antibody against dCYC may recognize a nonclock component antigen, these results suggest that these cells may indeed be part of a clock-neuron network, similar to the PDF-expressing small lateral neurons ventral (sLNvs) in Drosophila (Helfrich-Forster and Homberg, 1993).
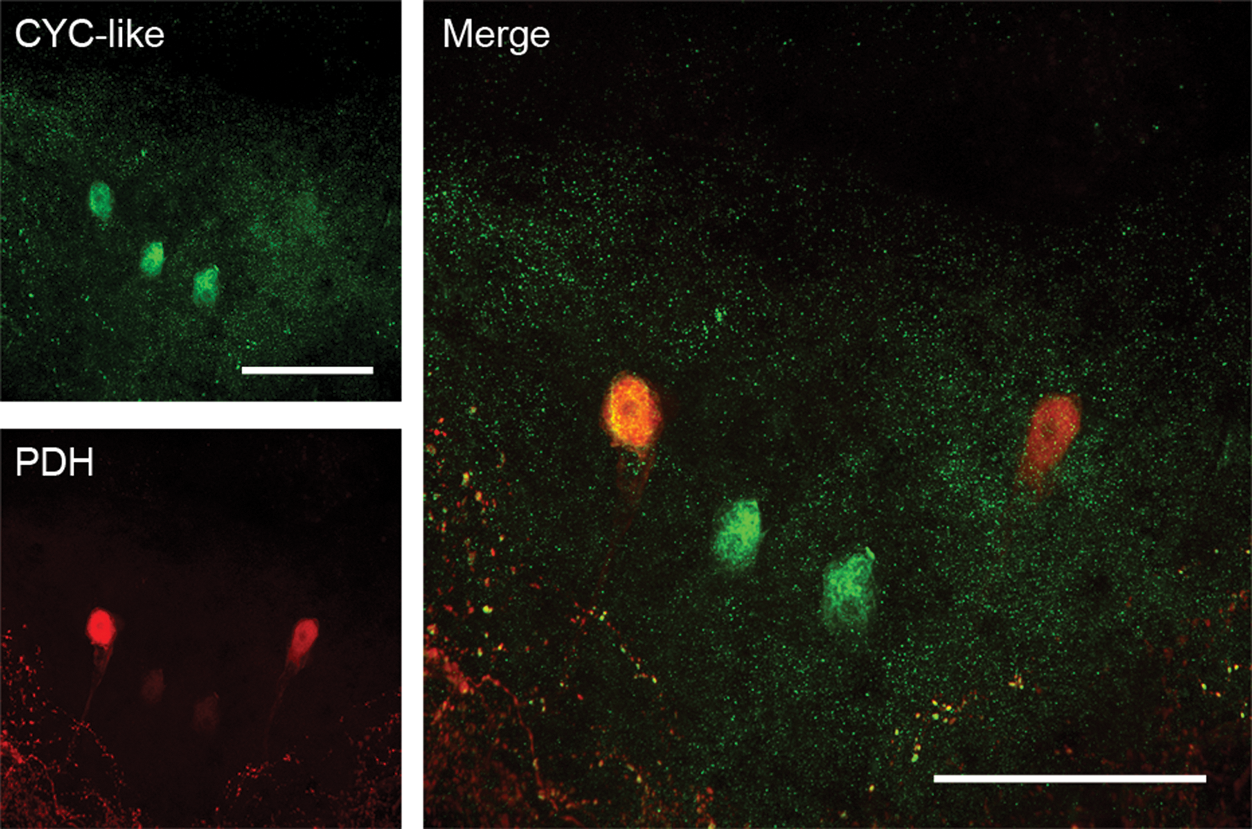
β-PDH-I is expressed in CYC-like–immunopositive neurons. Cells within the supraesophageal ganglion of C. productus that show CYC-like immunoreactivity are also immunoreactive for β-PDH-I. Red: PDH immunofluorescence. Green: CYC-like immunoreactivity. Images represent a 47-µm-thick projection of optical images taken every 1.8 µm from a whole-mount brain from a crab dissected at ZT12. Scale bar = 200 µm.
Heterologous Expression of β-PDH-I or II in Drosophila Re-creates the PDF Subcellular Localization in the PDF Circuit
Both the phylogenetic proximity between β-PDH-I and PDF and the expression of β-PDH-I within CYCLE-like–immunoreactive cells suggest that β-PDH-I could represent a functional ortholog of PDF. Because functional analysis of candidate genes is not a possibility in C. productus, we capitalized on the Drosophila GAL4/UAS system (Brand and Perrimon, 1993) and generated transgenic fly strains that express the full pre-prohormone genes for either β-PDH-I or β-PDH-II under the control of the UAS sequence. We directed the constructs to the PDF circuit using the pdfGAL4 driver line. All these strains were introduced in the pdf01-null mutant background in order to test the ability of each β-PDH to rescue the pdf01 behavioral phenotype. To corroborate proper expression of the transgenes in the PDF circuit, we performed immunohistochemistry using the anti-PDH antibody in brains from β-pdh-I-, β-pdh-II-, or pdf-expressing flies dissected at ZT2 (2 hours after the dark-light transition). Figure 3 shows similar patterns of expression in all three strains. The somas of the small and large LNvs show intense and homogeneous staining, while the dorsal projections of the sLNvs, a critical set of fibers in the circadian circuit of the fruit fly (Fernandez et al., 2008), depict a punctate signal typical of the PDF circuit. Because the antibody against PDH could potentially recognize the peptides with different affinity, we also measured mRNA levels of the β-pdh-I and β-pdh-II transgenes by real-time reverse-transcription PCR at ZT2, using tubulin as a reference gene. The relative abundance of β-pdh-I and β-pdh-II mRNAs was not significantly different (0.065 ± 0.003 and 0.062 ± 0.016 arbitrary units, respectively; Student t test, p = 0.85). These experiments indicate that both crab transgenes are similarly recognized by the fly’s transcription, translation, and potentially, posttranslation machinery, validating the comparison between flies expressing β-PDH-I and II within the PDF circuit.
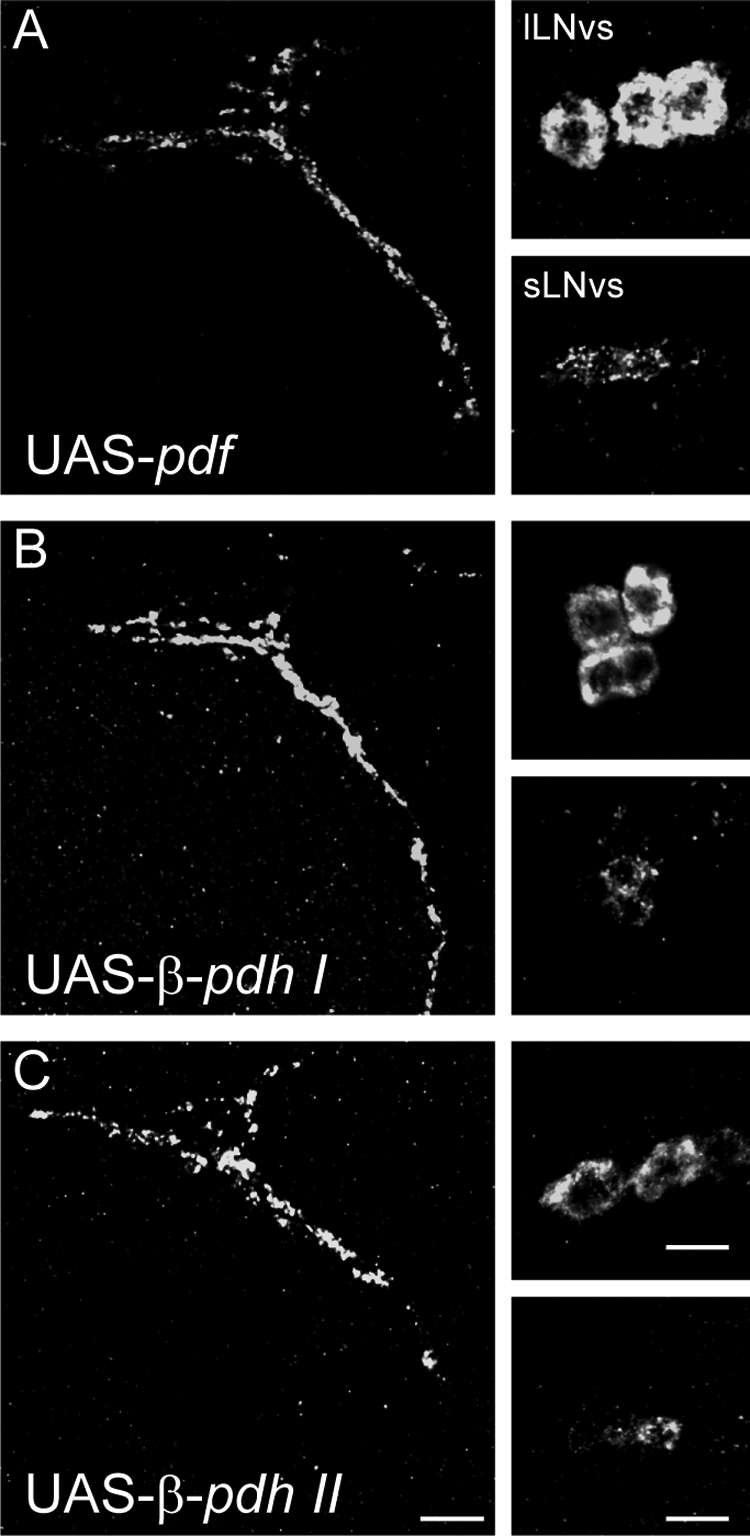
Heterologous expression of crustacean β-PDHs in the brain of Drosophila leads to proper peptide distribution in transgenic flies. Representative β-PDH immunostaining of the PDF circuit of adult transgenic fly brains. (A) pdfGAL4/UAS-pdf;pdf01/pdf01, (B) pdfGAL4/UAS-β-pdh-I;pdf01/pdf01, and (C) pdfGAL4/UAS-β-pdh-II;pdf01/pdf01. For each genotype, the left panel illustrates the dorsal projection of the sLNvs, while the right panels show the lLNvs somatas (upper) and the sLNvs somatas (bottom). Scale bar = 10 mm.
β-PDH-I but Not β-PDH-II Rescues the pdf01 Mutation, Mimicking Native PDF
The circadian clock of wild-type flies maintained under LD cycles consolidates locomotor activity during dawn and dusk, with a prominent siesta during the middle of the day (Fig. 4A, 4B, and 4G and Table 1). The morning and evening activity peaks consist of a gradual increase in the activity that starts in the hours before each transition. Upon release of the animals into constant darkness (DD), the morning peak is reduced, and the activity is consolidated throughout the subjective day, with a burst of activity around the subjective evening (Hamblen et al., 1986). In contrast, pdf01-null flies under LD conditions show no increased morning activity before lights-on, and their evening anticipatory activity is advanced and reduced in amplitude. Furthermore, they show decreased rhythmicity and a shortened circadian period under DD (Lin et al., 2004; Peng et al., 2003; Renn et al., 1999) (Fig. 4C and 4G).
β-PDH-I and PDF show no differences in their ability to rescue the behavioral phenotype of pdf01 flies.
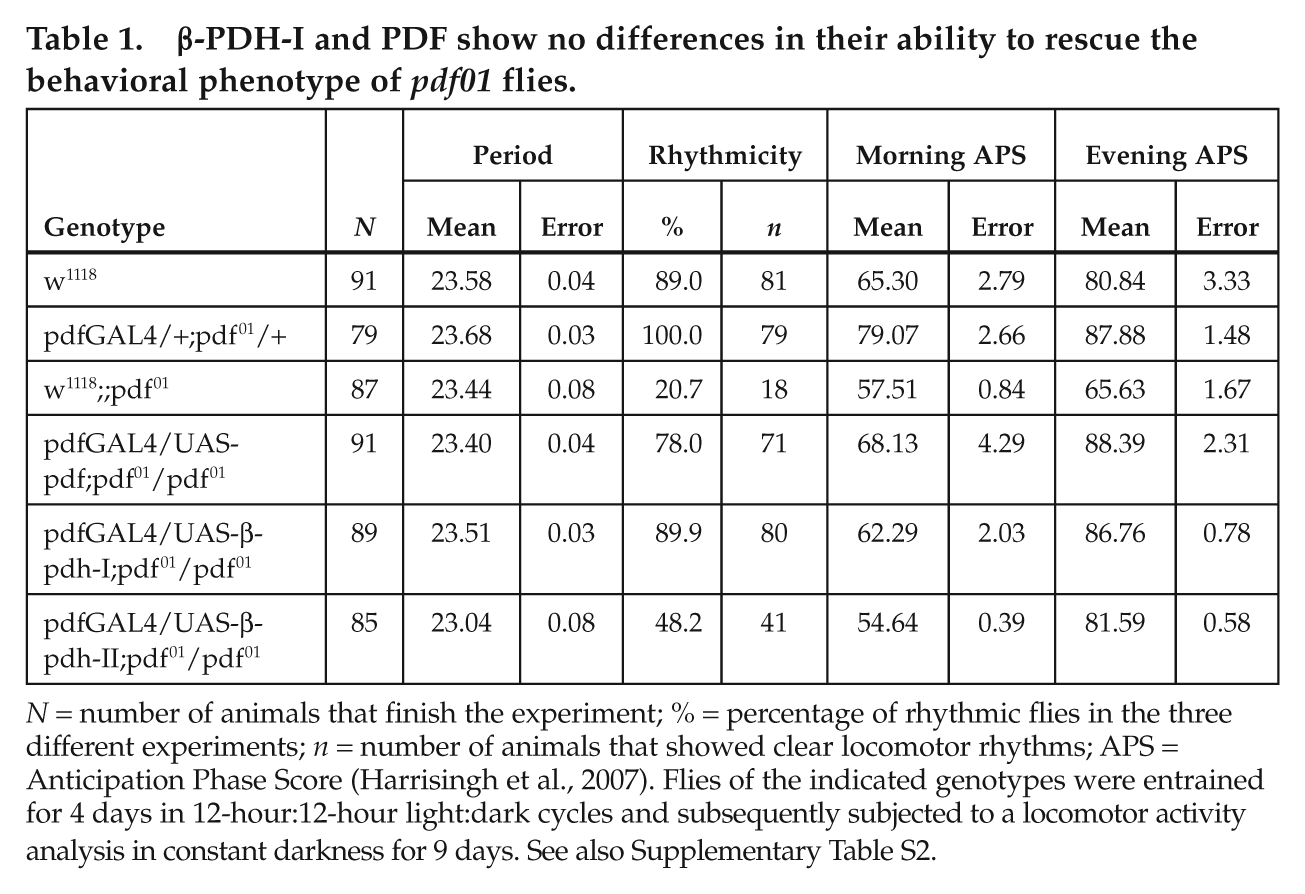
N = number of animals that finish the experiment; % = percentage of rhythmic flies in the three different experiments; n = number of animals that showed clear locomotor rhythms; APS = Anticipation Phase Score (Harrisingh et al., 2007). Flies of the indicated genotypes were entrained for 4 days in 12-hour:12-hour light:dark cycles and subsequently subjected to a locomotor activity analysis in constant darkness for 9 days. See also Supplementary Table S2.
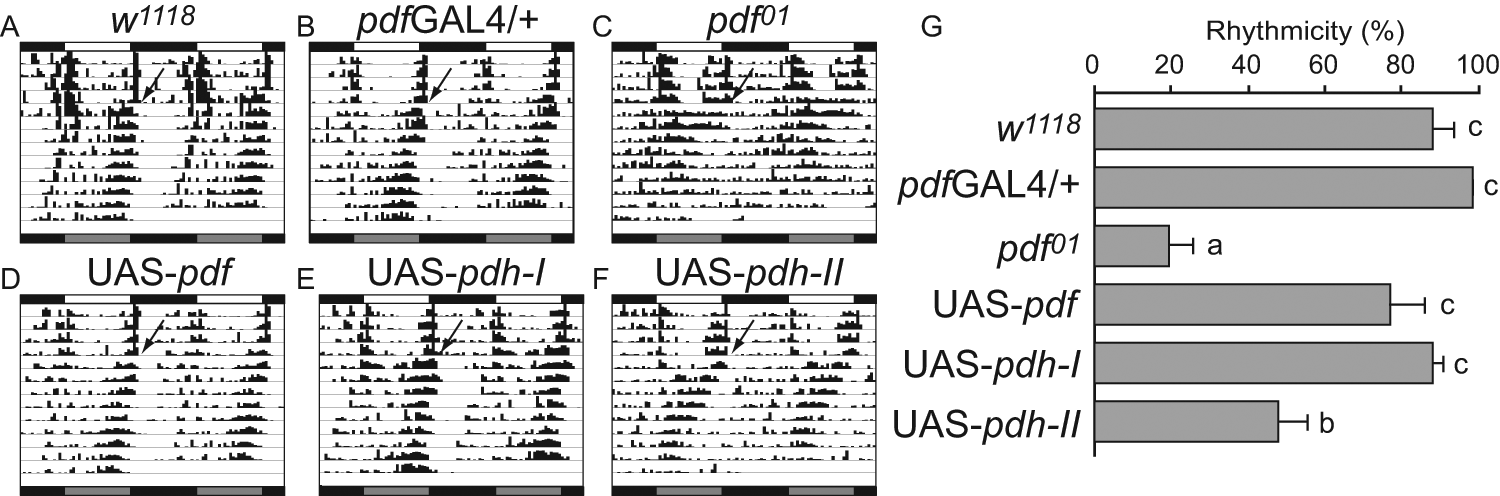
β-PDH-I but not β-PDH-II rescues the pdf01 mutation, mimicking native PDF. (A-F) Representative double-plotted actograms of flies of the following genotypes. (A) Control w1118, (B) pdfGAL4 driver alone in the w1118;;pdf01 genetic background, and (C) w1118;;pdf01 mutant, and rescues by overexpression of pdf (D), β-pdh-I (E), and β-pdh-II (F) in the w1118;pdfGAL4;pdf01 background. Each panel depicts the activity of a single fly throughout the experiment. Open horizontal bars = day; black horizontal bars = night; gray boxes = extrapolated subjective day after release into DD. Black arrows indicate transfer to constant darkness. (G) Percentage of rhythmic flies for the same genotypes shown in A to F. The mean value of three independent experiments ± SEM is plotted. Same letter indicates nonstatistical differences in a Tukey comparison (p = 0.05).
To assess the ability of each peptide to rescue the pdf01 phenotype, we evaluated locomotor activity rhythms of transgenic flies bearing the pdfGAL4/UAS-β-PDH-I or II construct in a pdf01 background and their appropriate controls (Suppl. Table S2). As previously reported (Renn et al., 1999), driving UAS-pdf expression with pdfGAL4 induces a full rescue of the pdf01 behavioral phenotype (Fig. 4D and 4G). Remarkably, expression of UAS-β-PDH-I in the PDF circuit also achieves this complete phenotypic rescue (Fig. 4E and 4G). In contrast, the expression of UAS-β-PDH-II modestly improves the rhythmicity, and flies display an unconsolidated activity profile. In addition, β-PDH-II–rescued flies show a short circadian period, a landmark of pdf01 flies (Fig. 4F and 4G and Table 1). The lack of restoration of the mutant phenotype is independent of the transposon landing site because an independent β-PDH-II transgenic strain tested in parallel shows similar results (Suppl. Table S2).
Interestingly, both β-PDH-I and PDF fully restored the characteristic wild-type phenotype under LD conditions (Fig. 5A, 5B, 5D, and 5E, black arrowheads). In contrast, β-pdh-II expression induced a modest morning peak and an evening peak that resemble those of pdf01 (Fig. 5C and 5F, gray and white arrowheads, and Table 1 for APS).
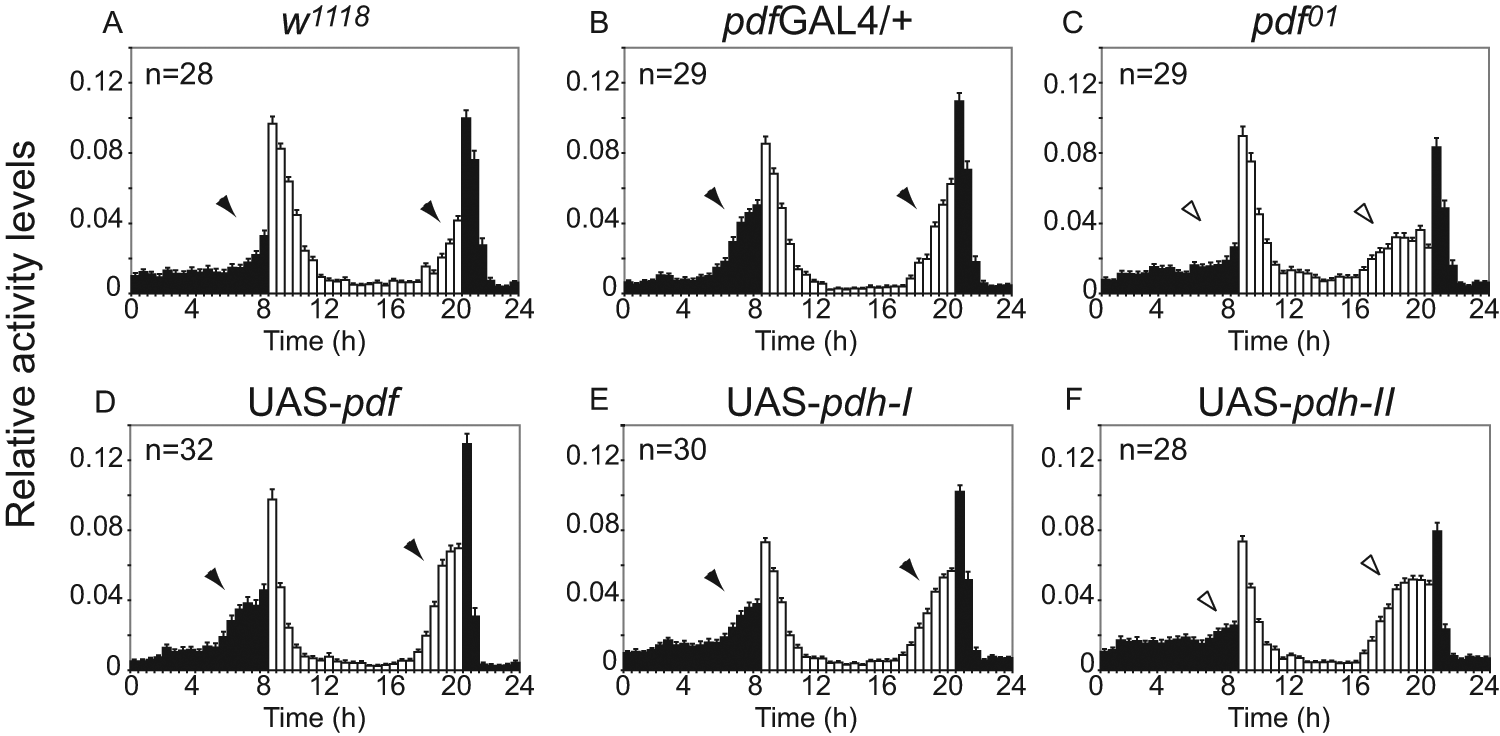
β-PDH-I rescues morning and evening anticipatory activity, while β-PDH-II only rescues evening anticipatory activity. (A-F) Representative 24-hour waveforms of locomotor activity of flies for the same genotypes shown in Figure 4. Each bar represents the normalized mean locomotor activity in a 30-minute interval (±SEM). The total number of individuals in each experiment is shown in the upper left corner. Black arrowheads indicate clear anticipatory activity, and white arrowheads indicate absence of a wild-type phenotype. See Table 1 for APS. White bars = day; black bars = night.
In summary, the restitution of rhythmic behavior achieved through β-pdh-I expression, but not by β-pdh-II, is indistinguishable from that accomplished with the native Drosophila pdf gene, supporting functional conservation between PDF and β-PDH-I.
β-PDH-I but Not β-PDH-II Recognizes the PDF Receptor, Mimicking Native PDF
While β-PDH-I–rescued flies are indistinguishable from PDF-rescued ones, β-PDH-II rescue induces an intermediate phenotype. This inability to fully rescue behavioral rhythmicity could represent an impairment of the fly biochemical machinery to process the prepro-β-PDH-II peptide. Alternatively, properly cleaved β-PDH-II may fail to recognize the fly PDF receptor. To address this possibility, we used the Epac-based cAMP sensor to measure cAMP levels, which increase after PDF receptor (PDFR) activation induced by PDF in vitro and ex vivo (Mertens et al., 2005; Shafer et al., 2008). While PDF and β-PDH-I induced similar increases in cAMP levels in the sLNvs, the induction by β-PDH-II was significantly lower (Fig. 6A), demonstrating a functional divergence between both PDH peptides in their ability to interact with the PDFR and activate the signaling cascade. Two-way ANOVA of the cAMP responses binned every 30 seconds, with peptide treatment and time as factors, yielded a significant effect of treatment, time, and interaction (p < 0.0001 for each of the three factors). Post hoc Tukey contrasts (p = 0.05) showed that the induction by each of the three peptides was significantly different from the vehicle treatment, that the induction by PDF and β-PDH-I did not differ, and that the induction by β-PDH-II was significantly lower than that of PDF or β-PDH-I.
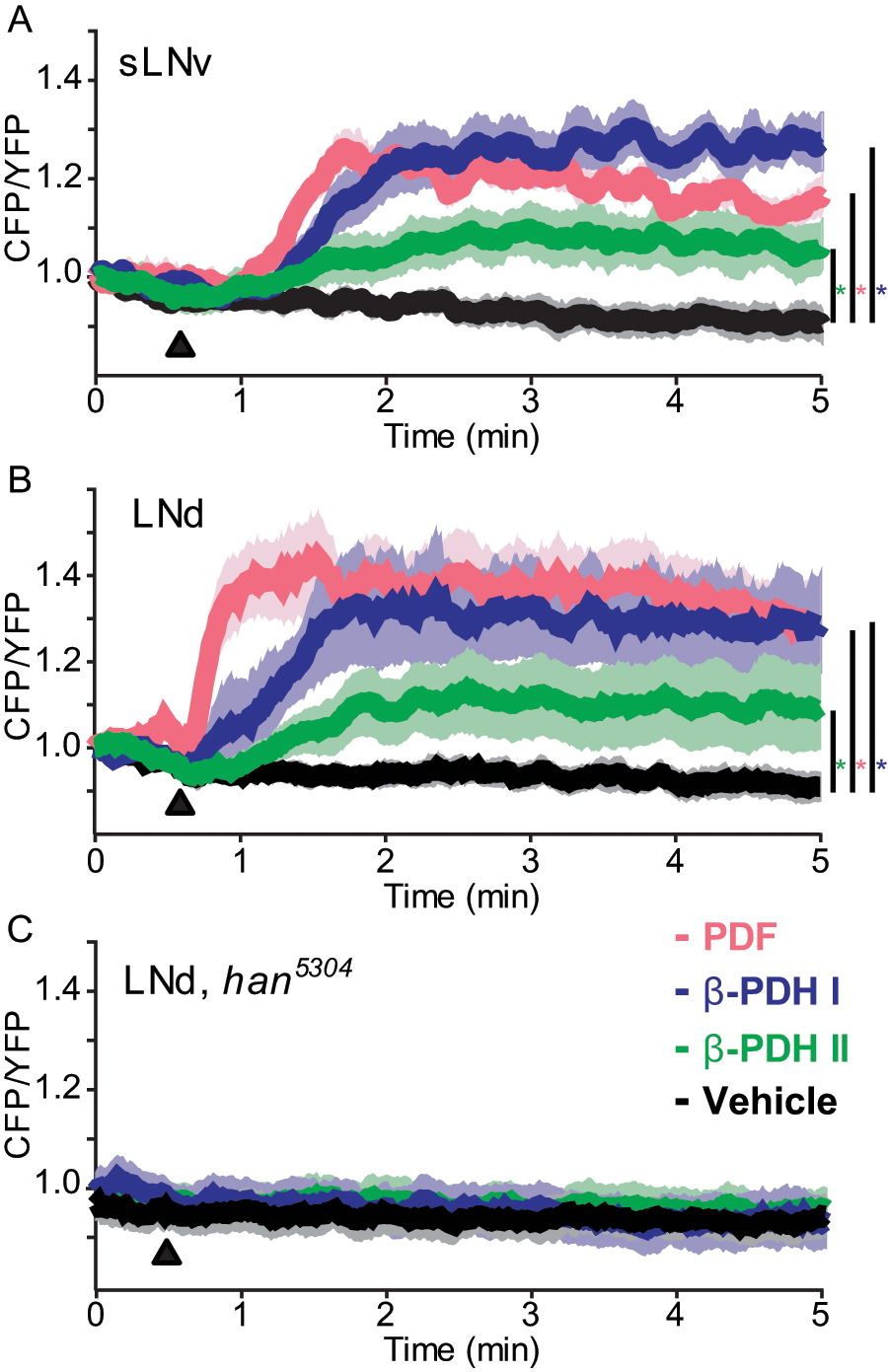
cAMP responses from Drosophila’s sLNv and LNd clock neurons following bath application of crustacean β-PDH neuropeptides. Responses from the small ventrolateral neurons (sLNv) and the dorsolateral neurons (LNd) were recorded using UAS-Epac1-camps driven by the Clock[856(8.2/2)]-gal4 element in male flies. (A) Average CFP/YFP (inverse FRET) ratio traces from the Epac1-camps sensor in the sLNv in response to bath application of PDF (magenta), β-PDH-I (blue), β-PDH-II (green), and the vehicle control (black). All peptides were applied at a final concentration of 10–4 M, in 0.1% DMSO in HL3 saline. (B) Average Epac1-camps inverse FRET traces from the LNd in response to bath-applied neuropeptides and vehicle control as in A. (C) Average traces from the LNd of han5304 mutant flies in response to bath-applied β-PDH-I (blue) and β-PDH-II (green), and the vehicle control, 0.1% DMSO (black). Black triangles in A, B, and C represent the introduction of the stimulus. Color-coded asterisks represent statistically significant differences between the indicated peptide and the vehicle; see the text for statistical analysis details.
Previous work has dissected the Drosophila crepuscular behavior under LD and argued that morning and evening activity peaks rely on different neuronal clusters. Under this scenario, sLNv neurons control the morning peak and the phase of the evening peak, while the LNds govern the rise of the evening activity burst (Grima et al., 2004; Stoleru et al., 2004; Stoleru et al., 2005). Because overexpression of β-PDH-II failed to rescue the main features of rhythmic locomotor activity but was effective in improving the evening anticipatory activity (Table 1), we evaluated the response of the LNd cluster to the three peptides (Fig. 6B). A 2-way ANOVA revealed a significant effect of treatment (p < 0.0001), time (p < 0.0001), and interaction (p = 0.0025). Post hoc Tukey contrasts (p = 0.05) showed that the induction by each or the three peptides was significantly different from the vehicle treatment and that all three peptides differ from each other in their induction of cAMP increases (PDF > β-PDH-I > β-PDH-II) (Fig. 6B). Post hoc Tukey contrasts for individual 30-second intervals showed no differences between PDF and β-PDH-I at any interval but differences between PDF and β-PDH-II for the 1- to 1.5-minute interval. Thus, similarly to the response in sLNvs, β-PDH-I in the LNds mimicked better the effect of PDF than β-PDH-II.
To determine if the response to each peptide in the different cell clusters specifically requires PDFR, we evaluated the response to their ex vivo application in a PDFR-null background using the han5304 strain (Hyun et al., 2005). Neither the sLNvs (not shown) nor the LNds showed any responses to either β-PDH-I or II, ruling out a nonspecific effect of the peptides through different receptors (2-way ANOVA: no effect of treatment [p = 0.19], effect of time [p = 0.01], and no effect of the interaction [p = 0.99]) (Fig. 6C, response for LNds).
In summary, the differential ability of β-PDH-I and II to recognize PDFR and activate its signal transduction cascade mirrors the differential ability of the peptides to rescue the circadian rhythmic behavior of pdf01 flies.
Discussion
Based on the presence of β-pdh-I– but not β-pdh-II–expressing neurons in the brain of C. productus, we hypothesized that β-pdh-I likely represented a functional ortholog of pdf in Drosophila (Hsu et al., 2008). Accordingly, here we demonstrate that all β-pdh-Is and pdfs identified to date cluster within a monophyletic group that does not include β-pdh-II. Furthermore, CYC-like–immunoreactive neurons within the brain of C. productus express β-PDH-I. As predicted, β-PDH-I, but not β-PDH-II, is able to mimic the effects of native PDF within the Drosophila clock-neuron network in its ability both to restore rhythmic behavior in pdf01 flies and to recognize PDFR.
Members of the β-PDH Family as Putative Clock Output Signals
Our phylogenetic analysis clearly places β-pdh-I and all other crustacean β-pdhs, except for β-pdh-II, within a clade that includes all known pdfs. This analysis, together with the fact that β-pdh-II genes have only been identified in brachyurans, suggests that the duplication that gave rise to the two β-pdh genes is unique to crabs. We postulate that this duplication has led to a neofunctionalization, with β-PDH-I serving as a clock output signal and β-PDH-II as the pigment-dispersing hormone.
The expression of β-pdh-I but not of β-pdh-II in the brain of C. productus favors this view (Hsu et al., 2008). In Drosophila, the circadian clock neuronal network is defined by cells with or without PDF expression. Although future studies will determine whether cells immunostained with the dCYC antibody truly express a functional homolog of the dcyc gene, the same seems to be true in the brain of C. productus, where a CYC-like peptide is coexpressed with β-PDH-I in some neurons but not in others. β-PDH-I–expressing neurons are located in cell cluster 6 within the anterior median protocerebrum of the supraesophageal ganglion of C. productus and project densely to the rest of the median protocerebrum (Hsu et al., 2008). A recent report in two species of crayfish showed that similarly located β-PDH–immunopositive neurons are colocalized with fibers that express the circadian photoreceptor CRIPTOCHROME and are part of the brain photoreceptor neuropils (Sullivan et al., 2009). Interestingly, lesion experiments by these authors suggest that this neuronal network contains both a circadian oscillator driving locomotor activity rhythms and its photoreceptors. Finally, in agreement with the expression of PDF in the Drosophila optic lobe (Helfrich-Forster, 1997), β-PDH-I is also expressed in neurons of the optic ganglia of the eyestalk of C. productus (Hsu et al., 2008).
Although future studies may discover β-PDH-II and its coding gene in other crustaceans, exhaustive searches in crustaceans such as isopods (Wilcockson et al., 2011) and Daphnia (Strauss et al., 2011) suggest the existence of only one β-PDH gene. Interestingly, the pattern of expression of β-PDH in Daphnia, as in the crayfish (Sullivan et al., 2009), suggests that the peptide may also be part of a circadian clock neuronal network. On the other hand, the role of β-PDH in the dispersion of pigments in crayfish has also been characterized (McCallum et al., 1991), suggesting that the same peptide could accomplish divergent functions—clock output and pigment dispersion—within the same species. Our phylogenetic analysis, together with the functional roles proposed for β-PDHs and PDFs in arthropods, suggests a functional spectrum across phylogeny. In insects, PDF likely plays a role as a clock output and coupling signal but has no role in pigment dispersion. In some crustaceans, a single β-PDH could act both as a clock-signaling molecule and a pigment-dispersing neuroendocrine signal. Finally, in a subgroup of crustaceans, gene duplication has led to a subfunctionalization of two orthologs that exert these respective functions.
β-PDH-I Is Equally Efficient as PDF in the Drosophila Circadian Network
The lack of genetic and transgenic tools in crabs led us to develop a heterologous assay to evaluate the putative circadian function of crustacean β-PDHs in Drosophila. This approach has been successfully used within the circadian system of Drosophila to assess the function of clock components (Levine et al., 1995; Nishinokubi et al., 2006; Shigeyoshi et al., 2002). In the case of β-PDHs, the approach offers a unique opportunity to test the prediction that the paralog that is phylogenetically closely related (β-pdh-I) should be more efficient than the phylogenetically divergent paralog (β-pdh-II) in rescuing the function of the Drosophila ortholog. Importantly, we were able to recapitulate the PDF expression pattern in pdf01 flies by expression of either β-pdh-I or β-pdh-II, which suggests that the fly’s transcriptional and translational machinery recognizes both foreign genes. Although previous studies have employed PDF expression to rescue the pdf01 circadian phenotype (Renn et al., 1999) and to study the properties of the PDF peptide (Yoshii et al., 2009), a full recovery of rhythmic locomotor activity with absolute penetrance using a crab peptide is remarkable. This is particularly so when we consider the poor ability of β-pdh-II to rescue the circadian activity profile.
Functional Conservation in Ligand-Receptor Interaction between β-PDH-I and PDF
Because the gene products of the β-pdh/pdf family are synthesized as pre-prohormones, the inability of β-PDH-II to rescue circadian locomotor activity could reflect the inability of fly neurons to posttranslationally process the peptide. The functional assessment of the ligand-receptor interaction allowed us to evaluate the ability of each mature peptide to interact with the native PDFR. β-PDH-I but not β-PDH-II mirrored the response induced by PDF, strongly suggesting that the poor behavioral rescue induced by overexpression of β-pdh-II is a reflection of the impaired ability of β-PDH-II to recognize PDFR. Interestingly, β-PDH-II was not able to restore the wild-type endogenous period of 24 hours in DD, which is controlled by the sLNvs, but it achieved considerable improvement in the evening anticipation peak under LD conditions, which relies on non–PDF-expressing clock neurons, including the LNds (Grima et al., 2004; Stoleru et al., 2004; Stoleru et al., 2005). These results suggest that partial activation of PDFR signal transduction pathway in the LNds may be sufficient to rescue the role of these cells within the circadian clock neuronal network.
The present study constitutes the first functional characterization of a clock output signal in an intertidal organism. Our results clearly demonstrate that β-PDH-I is able to fully recapitulate PDF activity in Drosophila, consistent with a conserved function between these peptides. Although it remains to be determined whether β-PDH-I signaling in C. productus sustains circadian or circatidal rhythmicity, our results represent a critical step towards the identification of the molecular and neural bases of circatidal rhythmicity.
Footnotes
Acknowledgements
We thank P. H. Taghert for the UAS-PDF and the pdf01 flies, N. Glossop for the dClk-Gal4 lines, K. Rao for the anti-PDH antibody, I. Edery for the anti-CYC antibody, J. Acevedo for advice with the generation of transgenic flies, and A. Leaché and V. Di Stilio for help with phylogenetic analysis. E.J.B. is supported by a graduate fellowship from the Argentine Research Council (CONICET). M.F.C. is a member of CONICET. This work was supported by NSF grant number IOB-0618029 to H.O.D., NIH grant NINDS R00NS62953 to O.S., and ANPCyT (Argentina) PICT2006-1249 and FIRCA-NIH grant 1R03TW008342 to M.F.C.
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.