
Abstract
Human adolescence is arguably one of the most challenging periods of development. The young adult is exposed to a variety of stressors and environmental stimuli on a backdrop of significant physiological change and development, which is especially apparent in the brain. It is therefore unsurprising that many psychiatric disorders are first observable during this time. The human intestine is inhabited by trillions of microorganisms, and evidence from both preclinical and clinical research focusing on the established microbiota-gut-brain axis suggests that the etiology and pathophysiology of psychiatric disorders may be influenced by intestinal dysbiosis. Provocatively, many if not all of the challenges faced by the developing teen have a documented impact on these intestinal commensal microbiota. In this review, we briefly summarize what is known about the developing adolescent brain and intestinal microbiota, discuss recent research investigating the microbiota-gut-brain axis during puberty, and propose that pre- and probiotics may prove useful in both the prevention and treatment of psychiatric disorders specifically benefitting the young adult.
Highlights
The young adult is exposed to numerous challenges and stressors during adolescence, a period in which the brain is still developing and many psychiatric disorders first manifest. The composition and diversity of the gut microbiota are also changing during adolescence, and the trajectory of these changes is affected by environmental challenges. Alterations to the gut microbiota during adolescence can influence the development of the brain and may contribute to the manifestation of various psychiatric illnesses associated with puberty. Microbial-based treatments may provide safe and effective methods for improving teenage psychiatric health and resiliency to the ill effects of experienced adverse events.
Introduction
Human adolescence is known to be a challenging period of development; it is a time when the young adult is frequently exposed to a variety of psychological stressors, often less than optimal nutrition and sleep, and increased opportunity for exposure to known neurodevelopmental teratogens such as alcohol and drugs. These challenges and exposures are occurring on a background of critical wiring for brain development—in fact, together with the neonatal period, adolescence is one of the most dynamic and important periods for growth and development in the brain. 1 –7
Perhaps unsurprisingly, adolescence is also a period of development when psychiatric illnesses frequently first manifest. Symptoms of anxiety, depression, eating disorders, psychosis and substance abuse are often first observed during this critical window. 2,8 We are beginning to increasingly understand that the pathophysiology of these disorders can be traced to changes that are unfolding in the adolescent brain, 8,9 alterations that can easily be influenced by the challenges and exposures frequently faced during the adolescent period (see Figure 1). 1,8,10
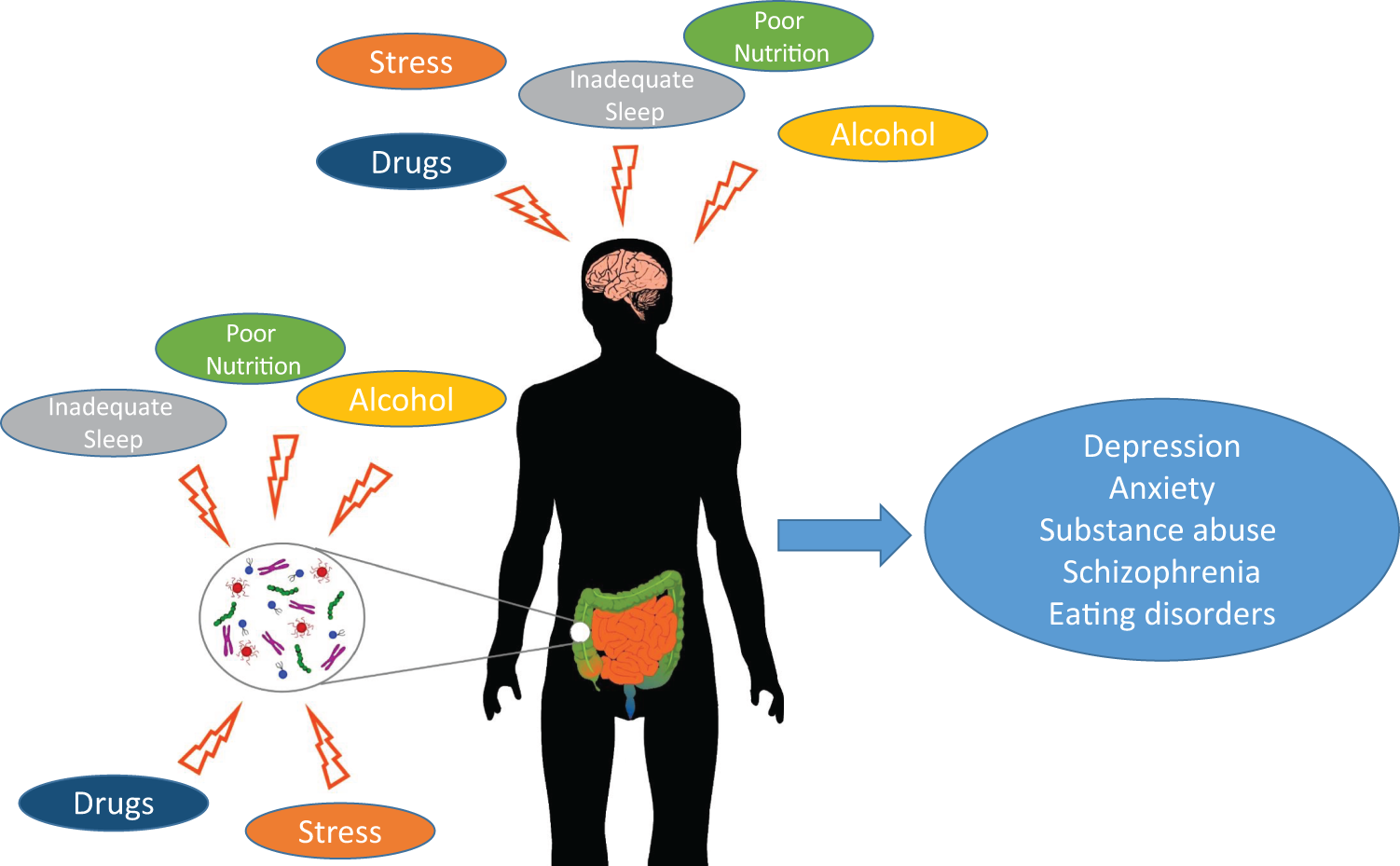
Adolescent exposures to stress, poor nutrition, inadequate sleep, and potentially alcohol and drugs affect both gut microbiota and brain developmental trajectories, which may have deleterious effects on mental health outcomes.
There has been a recent surge in evidence that the trajectory of postnatal nervous system development is influenced by the intestinal microbial status. 11 –14 Work in preclinical models of intestinal microbial dysbiosis, such as those using germ-free (GF) rodents or antibiotic-treated animals, has shown alterations to behaviour in addition to changes in associated brain neurochemistry. 13,15 –19 Clinical studies examining intestinal microbial status in relation to a number of gastrointestinal and psychiatric disorders indicate that the bacteria housed in the gut likely play a significant role in disease presentation and/or pathogenesis. 20 –27 In fact, the well-established gut-brain axis, or the bidirectional highway of communication connecting the gut and brain, has been recently extended to include the bacterial components of the intestinal lumen, leading to the now extended concept of a microbiota-gut-brain axis. 28 –31 Mechanisms by which communication occurs in this axis is the focus of robust scientific efforts and likely involves a combination of activity from immune, endocrine, metabolic, and neural pathways. 32
In this review, we consider the rapidly expanding evidence of the microbiota-gut-brain axis, particularly with respect to the developing adolescent brain. We briefly summarize what is known about neural development during this critical window and the increasing information we have about the intestinal microbial status at this time point. We hypothesize that alterations to the bacterial load of the gut during this time may be influencing the development of the brain and may play a mechanistic role in the manifestation of various psychiatric illnesses associated with puberty. Last, we propose that therapeutics aimed at the intestinal microbiota will provide alternatives for both the prevention and treatment of psychiatric illnesses associated with adolescence.
Brain Development during Adolescence
The physiological changes that occur in the adolescent are thought to reflect the evolution of the young adult readying to leave the safety of home and to lead to the ultimate achievement of independence from caregivers. Similarly, significant social and cognitive changes are associated with puberty in the transition to adulthood that allow for that same independence. 3,33 Historically, the brain was considered to have been largely finished in development by puberty, but we now know that this period of profound transition is also reflected in neuroanatomical change and maturation.
Postmortem Evidence for Structural Brain Development in Adolescence
Research on postmortem human brains in the late 1960s and early 1970s first revealed that the frontal lobes of the brain continue to develop well past adolescence and into early adulthood. 34,35 The frontal lobes are key structures involved in executive functioning, cognition, and working memory, 36,37 and 2 main structural alterations were found to occur there before and after puberty: changes in synaptic density and axonal myelination.
The developmental focus in early life is on synaptogenesis, but this is soon followed by a period of synaptic elimination and pruning. 33,38 Synaptic pruning is thought to be critical for the fine-tuning of neural networks in the brain, rendering the communication between the remaining synaptic circuits more efficient, 39 as it involves the elimination of many of the synaptic connections formed early in development. While the exact mechanisms by which pruning occurs are unknown, we do know that resident microglia play a role in the eradication of synapses and the strengthening and maintenance of others. 40,41 In sensory regions, synaptogenesis occurs during the first few months following birth, after which time pruning begins, with synaptic density reaching adult levels before or around adolescence. 35,42,43 The time course of synaptic development in the frontal lobes, however, is strikingly different. In the prefrontal cortex, there is a proliferation of synapses during childhood and again in puberty, followed by a plateau in synaptic density and a subsequent postpubertal elimination of synaptic connections. 34,44,45 These data suggest that synaptic pruning occurs primarily during adolescence in the frontal lobes and results in a reduction in the total amount of synaptic connections in adulthood.
The second main difference between brain structure in the pre- and postpubertal periods pertains to changes in myelin. As neurons develop, glial cells produce myelin, which wraps around axons to increase the speed of neural communication. In the first few years of life, sensory and motor areas are already fully myelinated, while the axons in the frontal cortex continue to undergo myelination throughout adolescence. 46 The implication of these findings is that the speed of neural communication in the frontal lobes is increasing from childhood to adolescence.
Imaging the Adolescent Brain
The advent of noninvasive brain imaging such as magnetic resonance imaging (MRI) has provided further evidence of the ongoing maturation of the frontal cortex in adolescence. During childhood, the frontal and parietal lobes both show general increases in grey matter cortical volume, which peak in puberty and then begin to decrease. 47 –53 Concurrent with this, volumes of white matter show a relatively constant increase. 47,54 These changes have widely been attributed to alterations in synaptic pruning and myelination. 8,33,38 It is likely that axonal myelination during adolescence results in an increase in white matter and a simultaneous decrease in grey matter as observed with MRI. Another possible interpretation is that changes to grey matter volume reflect synaptic remodeling; the increase and decrease of grey matter volume at the onset and after puberty have been attributed to synaptogenesis and synaptic pruning, respectively. 33,38
Pubertal Changes in Stress Reactivity
Adolescence and puberty are marked by significant changes in the endocrine system, resulting in altered hormonal signaling. Although most studies have focused on gonadal hormones, recent attention has been drawn to understanding the development of the hypothalamic-pituitary-adrenal (HPA) axis and stress responsivity during adolescence (for reviews, Eiland and Romeo, 55 McCormick and Mathews, 56 Romeo and McEwen, 57 and Romeo 58 ). The majority of evidence for development of stress responsivity in adolescence comes from research in rodent models. During puberty, gonadal hormones increase dramatically, whereas basal levels of stress hormones remain relatively constant. 59 In contrast, the increase in corticosterone to both physical and psychological stressors is prolonged in prepubertal compared to adult rats. 60 –62 Interestingly, this elongation of stress hormone signaling is not mediated by the different gonadal hormonal milieus of these 2 developmental stages. 61 Repeated exposure to the same stressor induces habituation of the hormonal stress response in adult rats; however, prepubertal males, but not females, display a higher peak level of corticosterone but a faster return to baseline. 63 In line with these findings, studies have found increased anxiety- and depressive-like behaviours in prepubertal rats previously exposed to a stressor. 64 –66 Taken together, the literature clearly shows that HPA axis signaling and stress responsivity undergo development during adolescence, but the mechanisms underlying these changes remain to be elucidated.
Dynamics of Gut Microbiota during the Adolescent Period
Recent improvements in next-generation microbial sequencing technology have allowed us to more reliably examine the constituent bacteria in the human gut and to look for patterns in diversity over the course of the life span. 67 We now know that while the intestinal microbiota of an adult is abundant, highly varied, and relatively stable, the infant microbiota is much simpler and less stable. 9,68 Colonization of the infant is considered to occur primarily during birth and the immediate postnatal period, and historically, the infant’s bacterial load was thought to be determined primarily by that which is resident in the mother. It is now becoming clear that this may only be true if the infant is vaginally born, as mode of delivery (i.e., vaginal or caesarean) has a distinct impact on bacterial colonization patterns. 69 –71 Various additional factors have been identified as influencing the continued development of microbiota during early life, including if the infant was hospital or home born, 72,73 formula or breastfed, 74 –78 and if there was any antibiotic use during this period. 79 –81
During childhood, we generally observe less diversity in microbial species compared to that which is seen in adults, 82 but during development and growth into the adolescent period, the intestinal microbiota similarly develops and slowly changes, with decreases occurring in numbers of aerobes and facultative anaerobes, as well as concurrent increases in anaerobic species. 83 A recent high-throughput analysis of distal gut microbiota that compared adults to adolescents found a significantly higher abundance of both Bifidobacterium and Clostridium genera in the adolescents, while the number of species was similar between the 2 groups. 84
While we are still far from an exhaustive list of changes that occur over the life span in terms of colonizing intestinal microbiota, what is abundantly clear is that the gut houses a community of diverse bacteria that are changing throughout life in response to physiological and environmental cues. Intriguingly, many if not all of the stressors that teenagers are exposed to are documented to affect the bacterial constituents of the intestinal lumen. Changes to diet, sleep patterns, stress, alcohol and drug use, and prescription antibiotic use all result in alterations to the intestinal microbiota. 15,85 –88 Given that adolescents are exposed to such a vast array of external challenges and stressors, all with known effects on the intestinal microbiota, we are then left with pertinent questions regarding the potential role of these bacteria in the psychiatric disorders that often emerge during the teenage years.
Adolescent Behaviour and the Microbiota-Gut-Brain Axis
A large-scale National Comorbidity Study conducted in the United States from 2001 to 2003 indicated that the peak age of onset for psychiatric disease as a whole is 14 years. 89 The mood disorders, including anxiety and depression, schizophrenia, and eating disorders, 8 often first manifest at this time. In healthy adolescents without clinical psychiatric illness, alterations in behaviour corresponding to increased social behaviours, risk taking, 7,90 sensation seeking, 91 impulsivity, 92 and emotional instability 7 are frequently observed.
In rodent models, the emergence of maladaptive behaviours during adolescence can be directly linked to the occurrence of early life challenges, with many of the preclinical studies examining the emergence of behavioural alterations during adolescence found in the social behaviour and autism literature. Injecting pregnant dams with valproic acid (VPA) is one of the most robust and well-characterized models for studying autistic-like symptoms in rodent offspring. It has been established that not only does this prenatal challenge result in altered social behaviours in adolescent offspring, but it also induces changes to the commensal intestinal microbiota. 93 Similarly, a recent study by Foley et al. 94 demonstrated that injecting rat dams with either lipopolysaccharide or propionic acid (a fatty acid product of many enteric bacteria) also resulted in long-term alterations to social behaviours in offspring, with the aberrations particularly apparent during the adolescent period. Both of these treatments also showed marked effects on the composition of the intestinal microbiota. Intriguingly, a recent study using maternal immune activation to induce autism-like symptoms in young adult offspring demonstrated that feeding the pups commensal bacteria Bacteroides fragilis ameliorated the otherwise observed behavioural deficits. 95
Given our growing understanding of the relationship between the microbiota and behaviour, there are still remarkably few studies examining the role of the microbiota-gut-brain axis in the development and function of the adolescent brain. To investigate the effects of manipulating the bacterial contents of the gut during this critical window of development, researchers have examined the impact of adolescent bacterial colonization of GF mice (mice bred and raised in the total absence of bacteria), while others have examined the impact of antibiotic treatment in otherwise normal juvenile animals.
GF mice show distinct abnormalities in HPA axis activity. Specifically, these animals show lifelong stress hyperresponsivity, with increased plasma adrenocorticotropin hormone (ACTH) and corticosterone following exposure to a stressor. 14 These alterations are accompanied by an abnormal behavioural phenotype, such as anxiolytic-like behaviour, 13,16 social behaviour deficits, 17 and cognitive dysfunction. 19 Intriguingly, if these mice are introduced to colonizing bacteria by the adolescent time period (in mice roughly 4 weeks of age), the dysfunctions in stress responsivity can be reversed, but not if the bacteria are introduced to GF mice in adulthood (i.e., past 6 weeks of age). This finding is important because it indicates that there may be a brief window in life, up to and encompassing the adolescent period, whereby commensal intestinal microbiota may have a lasting effect on central stress circuitry. Thus, given that the dysregulation of brain circuitry responsible for stress responsivity is implicated in all of the psychiatric illnesses that often first appear in adolescence, the fact that these circuits are responsive to changes in gut microbiota during this same time point is highly provocative.
A recent study examining the effects of antibiotics administered at weaning in mice (generally between 3 and 4 weeks of age and again considered roughly equivalent to an adolescent period) found significant differences in adult intestinal microbial content, behaviour, and brain neurochemistry posttreatment compared to controls. 15 Specifically, mice were given a combination of antibiotics administered in drinking water for 4 weeks, after which adult behaviours were assessed. Mice receiving antibiotic showed reduced anxiety-like behaviour in the light-dark test and altered cognitive functioning and social behaviours, as assessed in a novel object recognition test and social transmission of food preference test, respectively. In addition, mice treated with antibiotics in young adulthood showed increased serum tryptophan and decreased kynurenine levels relative to controls and significantly reduced hippocampal brain-derived neurotrophic factor (BDNF) messenger RNA (mRNA). Alterations in brain monoamines and monoamine metabolites as well as hypothalamic oxytocin and vasopressin mRNA expression were also observed following antibiotic treatment. 15 This study demonstrates that altering the intestinal microbial population through antibiotic treatment in the adolescent time period can have long-term effects on brain function and gene expression. Intriguingly, the anxiolytic-like behaviour observed in these mice mirrors those observed in GF mice, which not only demonstrate reduced anxiety-like behaviour but also associated changes in hippocampal BDNF expression. 13,16
Future Directions
Given our expanding understanding of microbiota-gut-brain communication, we are now at an exciting time for the design and development of potential therapeutics targeting psychiatric disease. There is currently a high level of interest in the use of pre- and/or probiotics as a novel avenue of therapeutic possibilities in the often refractory psychiatric illnesses. Indeed, the therapeutic potential of bacteria in the treatment of psychiatric illnesses has led to the development of the term psychobiotic and has been initially examined in a few recent studies. 96 Most clinical studies examining the potential impact of commensal bacteria on behavioural symptoms have been done on patients with irritable bowel syndrome (IBS). Patients with IBS frequently present with comorbid anxiety and depressive illnesses, and several different strains of bacteria have proven to be useful in combatting the psychological symptoms associated with the disorder. 97 –100 Similarly, a randomized, double-blind, placebo-controlled study of patients with chronic fatigue syndrome also found commensal bacteria to be beneficial in the treatment of anxiety symptoms. 101 Studies in healthy volunteers have also shown psychological benefits of certain bacterial strains, with probiotic treatment associated with reduced levels of psychological distress and improved overall mood. 101,102
Concluding Remarks
While there are many examples of preventative measures directed at adolescents for navigating through the various challenges of this tumultuous time in life, we would be wise to direct our consideration to potential therapeutics aimed at the intestinal microbiota. Possible preventative and/or ameliorative treatment effects of pre- and probiotics have been heretofore unexamined for adolescents and would provide an exciting, safe, and potentially highly effective method for improving teenage resiliency to the multiple and varied challenges they face. The vulnerability of the adolescent brain due to its higher degree of plasticity can be exploited to our advantage. The malleability of the nervous system at this critical period can be harnessed, allowing us to intervene in high-risk individuals. Specifically, we could target the intestinal bacteria and thus the microbiota-gut-brain axis to both incur protection to the brain from challenges and to ameliorate the ill effects of experienced adverse events. Improving teenage mental resiliency through microbial-based interventions is an exciting and heretofore unexamined possibility, one with highly anticipated opportunities for better psychiatric health.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.