
Abstract
Pterygium is a common ocular surface disease observed in humans. Chronic ultraviolet (UV) exposure is extensively recognized as an aetiological factor in the pathogenesis of this disease. This hypothesis is sustained by epidemiological and histopathological data in relation to UV injured skin. Although some findings have indicated that genetic factors, anti-apoptotic and immunological mechanisms are involved in the pathogenesis of pterygium, the mechanism by which it develops remains poorly understood. In this study, we analysed the in vivo production of IL-17A, IL-6, IL-10 and nitric oxide (NO) in the tears and sera from Algerian patients. Interestingly, we observed that IL-6, IL-17A and NO production in the tears and sera of all patients was strongly associated with inflammatory infiltration, NOS2, NF-κB and Bcl2 expression in pterygia biopsies. Collectively, our results indicate a relationship between local inflammation and anti-apoptotic processes in pterygium disease, leading to both tissue damage and enhanced cellular proliferation.
Introduction
Pterygium is a proliferative, inflammatory and invasive ocular surface disease associated with excessive exposure to ultraviolet (UV) light1,2 and has long been believed to be an environmental pathology. UV radiation has been found to play a main role in pterygium development. 2 In addition, pterygium occurs more frequently in Australia, 2 Africa2,3 and particularly, Algeria. Pterygium is a wing-shaped ocular surface lesion traditionally described as an encroachment of bulbar conjunctiva onto the cornea.1,2
Moreover, pterygium has historically been considered a degenerative disease, characterized by the degradation of Bowman’s layer and elastosis. 2 Clinical advances in surgical treatment involve the use of conjunctival autografts and amniotic membranes in combination with adjuvant therapies, including mitomycin C, β-radiation and 5-fluoroacil, to reduce recurrence. 4 In addition, this disease is characterized by cellular proliferation, inflammatory infiltration, fibrosis, angiogenesis and the breakdown of extracellular matrix. 1
The mechanism of pterygium pathogenesis and its progression remain an ophthalmic question. Interestingly, pterygium exhibits a preference for the nasal limbus and affects only humans, indicative of an unique ocular morphology. 2
While the pathogenesis of pterygium is not completely understood, important progress has been made towards understanding the mechanisms involved in its pathogenesis. Some researches indicate that signalling pathways activated by UV light result in the induction of mediators responsible for the growth of pterygium.1,2 In the same context, genetic studies have reported the implication of hereditary factors in pterygium formation. 1 In rural Australia, a strong positive correlation between climatic UV radiation and pterygium prevalence was found, providing further evidence of a causal relationship. 5 However, the mechanism by which UV light induces the development of pterygium remains elusive. 6 To this end, several investigations have been conducted to identify the signalling pathways activated by UV light in pterygium epithelial cells (PECs) that mediate the production of cytokines and growth factors. 7 In one report, it was postulated that heparin-binding epidermal growth factor-like growth factor (HB-EGF) in the same cultured cells is involved in the development of pterygia in relation to UV irradiation. 6
Following the discovery of Th17 cells, many queries regarding the physiopathology of certain chronic inflammatory diseases can now be clarified.8–11 Th17 cells are primarily characterized by the transcription factor RORγt, the surface markers IL-23R and CCR6, as well as the elevated production of the pro-inflammatory cytokines IL-17A, IL-17F, IL-21, IL-22, tumour necrosis factor α (TNF-α) and chemokine (C-C motif) ligand 20 (CCL20).12,13 In addition, these pro-inflammatory cytokines are capable of promoting local tissue damage.13,14 It has been reported that Th17 responses play pivotal roles in the pathology of chronic inflammatory diseases.11,15 Th17 cells are activated by the combination of IL-6 and transforming growth factor (TGF-β), stimulating the transcription factor RORγt.13,16
The generation and secretion of high nitric oxide (NO) concentration by infiltrating cells may lead to perpetuation of local tissue damage.11,17,18 We have previously reported that pro-inflammatory cytokines induce inducible nitric oxide synthase (NOS2) expression and activity in chronic inflammatory diseases such as inflammatory bowel diseases and Behçet disease.10,19 In this study, we focused our attention on understanding the mechanism of inflammation involved in pterygium. In this context, we examined the potential association of IL-17 and NO pathways with pterygium development by investigating the production of IL-17A, IL-6, IL-10 and NO in the sera and tears of Algerian patients with pterygium (n = 43). This study was conducted concomitantly with histological analyses, examination of inflammatory infiltrates and the expression of NOS2, NF-κB and Bcl2 expression in pterygium specimens.
Patients and methods
Patients
A total of 43 Algerian patients with Pterygia (32 men and 11 women; age range: 35−62 years) were enrolled in this study. Patients were diagnosed by standard examination in the ophthalmological department (CHU Annaba, Est of Algeria). Most patients exhibited pterygium with a unilateral location. Healthy controls samples (n = 43) were obtained from adult volunteer donors who were operated for strabismus. Positive controls samples (n = 30) were obtained from patients with inflammatory ocular diseases (conjunctivitis, n = 20) or another inflammatory affection (Sjögren-Gougerot syndrome, n = 10). All participants provided informed consent for this study, which was carried out according to the World Medical Association Declaration of Helsinki adopted by the ethical committee of Algerian Agency of Research Development in Health (ATRSS). None of the subjects had received pharmacological treatments. The negative controls did not exhibit any inflammatory or infectious diseases.
Pterygia specimens and controls
A total of 43 pterygia and 63 other conjunctiva (positive and negative controls) were surgically removed. The pterygia excisions were made under topical anaesthesia. The surgery was performed behind the limbus, by first removing the heads and extending the incisions into the bodies until the semi-lunar folds (we eliminated pterygia which have invaded Bowman’s layer in the cornea from our study). The other conjunctiva samples (controls) were collected from the temporal bulbar conjunctival region of the eye affected by strabismus or conjunctivitis disease. In the case of primary Sjögren syndrome, labial salivary glands biopsies were removed from the lower lips. For proper orientation, the specimens were sutured onto a sterile cardboard, immediately fixed in 10% neutral buffered formalin and subsequently embedded in paraffin.
Serum collection
Blood samples, collected from healthy donors and patients, were centrifuged at 600g for 10 min to obtain the sera. All serum samples were stored at −45°C until the assessment of IL-6, IL-10, IL-17A and NO at a later time.
Tear collection
Reflex tears were collected using glass capillary micropipettes that had been flame polished before processing (taking care not to touch the conjunctiva and the cornea). Following the stimulation of tears by exposing the diseased and control eyes to a slit lamp, tears were collected in sterile vials and then stored at −70°C until further processing. 20
Histological analysis
The histological sections were stained with haematoxylin-eosin. Samples were fixed for 24 h in 10% buffered formalin at room temperature. The specimens were dehydrated by graded ethanol and embedded in paraffin blocks. Tissue sections (3-μm thick) were deparaffinized with toluene and rehydrated through a graded series of ethanol, then stained with haematoxylin and eosin. Digital images were captured with a digital camera integrated with a LEICA light microscope at different magnifications.
Immunohistochemistry procedures
The immunohistochemical study of CD3, CD68, Bcl2, NOS2 and NF-κB expression was performed on formalin-fixed, paraffin-embedded samples of pterygia and control conjunctiva. Primary Sjögren syndrome samples served as positive controls for NOS2 immunostaining. These paraffin blocks were cut into 3-μm thick sections, deparaffinized in xylene and rehydrated through a graded series of ethanol. For antigen retrieval, the tissue sections were boiled in a target retrieval solution (Dako) for 40 min in a 96°C water bath. For immunohistochemical detection, anti-CD3 (monoclonal antibody clone, dilution of 1:25–1:50, Dako), anti-CD68 (monoclonal antibody clone, dilution of 1:50–1:100, Dako) and anti-Bcl2 (monoclonal antibodies, dilution of 1:50–1:100, Dako) were applied for 1 h at room temperature. To detect NOS2 and NF-κB expression, the sections were incubated with anti-NOS2 (rabbit polyclonal antibodies clone, dilution of 1:500, Santa Cruz Biotechnology) and anti-NF-κB p65 subunit (rabbit monoclonal antibodies, dilution 1:100, Santa Cruz Biotechnology), respectively, overnight in a humidified chamber at 4°C. These steps were followed by the application of a secondary antibody (horseradish-peroxidase-conjugated anti-mouse IgG Ab) and then the DAB (3,3′,4,4′ diaminobenzidine). The DAB chromogen solution was freshly prepared, counterstained with Mayer’s haematoxylin and mounted with Faramount (both from Dako). The slides were observed using a standard light microscope (LEICA), and pictures were taken using a digital camera at different magnifications.
Enzyme-linked immunosorbent assay for IL-17A, IL-6 and IL-10
The levels of IL-17A, IL-6 and IL-10 in the serum and tears (sensitivity: <2, <1 and <1 pg/mL, respectively) were determined using an enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer’s instructions (Life Technologies). The absorbance was read on an ELISA reader at 450 nm (LABSYSTEM). These assays detected only human cytokines. The results are expressed as pg/mL in relation to a standardized dose curve of the relevant recombinant cytokine (IL-17A, IL-6 or IL-10).
NO assay
NO production was assessed by the determination of the end products of NO oxidation in the sera and tears. The modified Griess method as previously described by our team
21
was used to determine the nitrite levels
Statistical analysis
The results are expressed as the mean ± standard deviation. The data were analysed using Student’s t-test for the comparison of two means. We considered the difference to be statistically significant with P < 0.05. Statistical analyses were performed using Minitab 16 software.
Results
Pterygia histopathology
In this study, the examination of the histological features revealed that the pterygium exhibits a conjunctivo-epithelial structure (Figure 1(b)–(d)). In most patients, the epithelial coverage appears as a stratified squamous-type epithelium without keratinization, covered with vacuolated goblet cells (GCs) constituting hyperplasia (Figure 1(c)). We observed that there was no evidence of GCs in the normal conjunctiva sections (Figure 1(a)). In comparison with these control biopsies, the pterygia connective tissue (Substantia Propria) comprised a dense and locally invasive follicular inflammatory infiltrate. This infiltrate is tightly connected to the inferior edge of the epithelium (Figure 1(c)). Moreover, the pterygia is often characterized by fibrosis and neovascularization and a substantial network of blood vessels surrounds the inflammatory infiltrates, likely related to angiogenesis (Figure 1(d)).
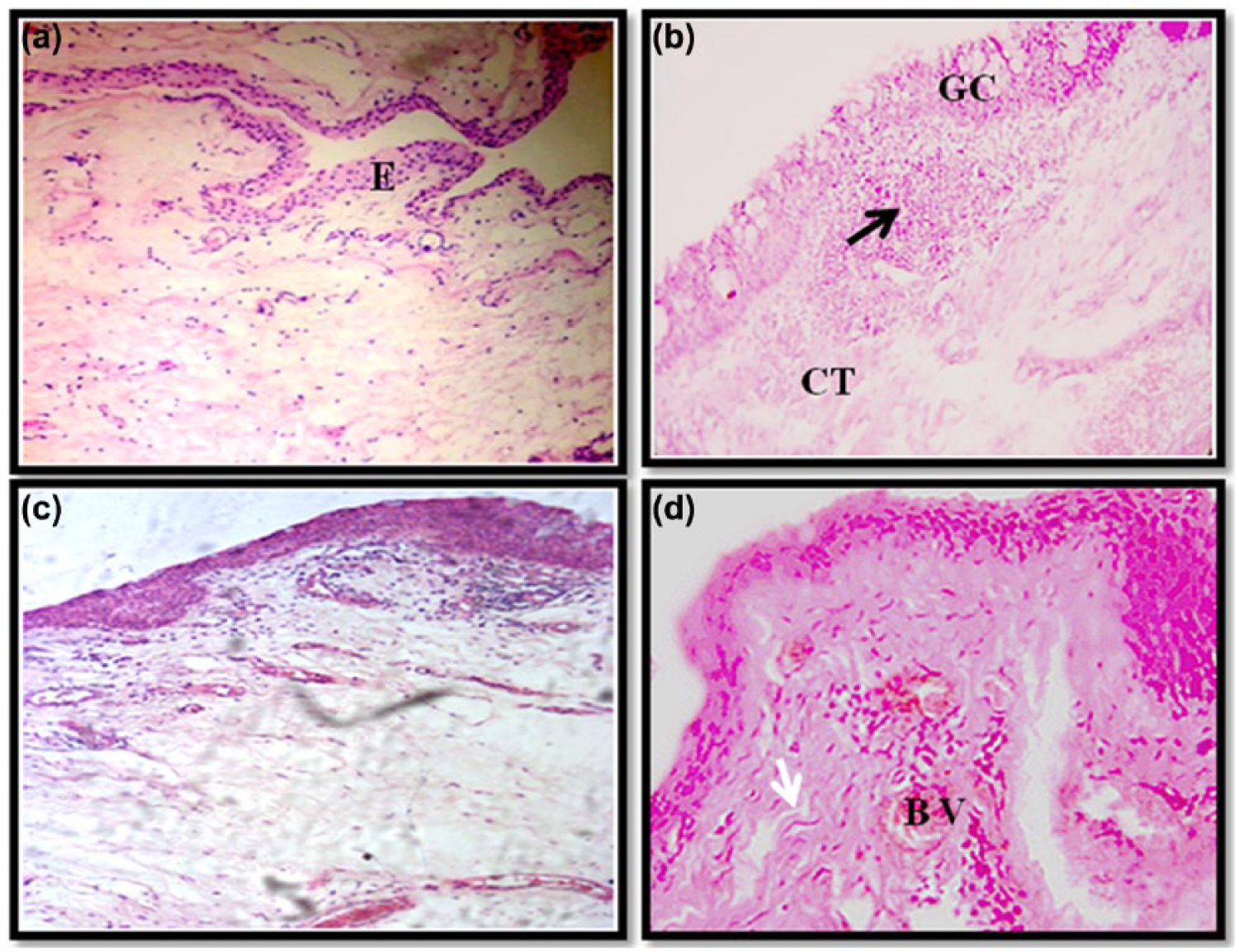
Histopathology of normal conjunctiva tissues (control) (a) and pterygium (b, c and d) biopsies. BV: blood vessels, CT: connective tissue, E: epithelium, GC: goblet cells. Black arrows denote the inflammatory infiltrate and white arrows indicate fibrosis (a and b: original magnification ×100 and c and d: original magnification ×250).
Detection of CD3, CD68, Bcl2, NOS2 and NF-κB in pterygium biopsies
Our results have shown that the number of T-lymphocytes and macrophages were significantly higher in the pterygium sections (CD3+ and CD68+ cells) than the negative controls (normal conjunctivas) (Figure 2(a) and (d) vs Figure 2(b) and (e), respectively). Cells were detected in the epithelium and in the connective tissue. In positive controls, the marked cells were predominantly present in the connective tissue (Figure 2(c) and (f)). Interestingly, in comparison to macrophages, T-lymphocytes represent the majority of the immune cells in the pterygium inflammatory infiltrate. In positive controls, the two populations showed a higher staining (Figure 2(a), (c), (d) and (f)). Interestingly, we noted that most T cells and macrophages were found around blood vessels and cover unevenly parts of the pterygium connective tissue (Figure 2(a) and (d)).

Immunohistochemical staining for CD3 (a, b and c) and CD68 (d, e and f), in pterygium, normal conjunctiva (negative controls) and conjunctivitis (positive controls) biopsies. All slides were counterstained with Mayer’s haematoxylin. The arrows denote the marked regions. Original magnification ×400.
Microscopic examination revealed an overexpression of NOS2 (inducible NO synthase) within the epithelium and the connective tissue of the pterygium (Figure 3(d)). In contrast, this was not observed in the normal conjunctival sections (Figure 3(e)). Moreover, in the same pterygium region, we noted an overexpression of NF-κB which was not noted in the control sections (Figure 3(a) and (b)). In contrast, the epithelium and the connective tissue in the labial salivary glands of the patients with primary Sjögren-Gougerot were also positively stained for both markers (Figure 3(c) and (f)).

Immunohistochemical staining for NF-κB (a, b and c) and NOS2 (e, f and g), in pterygium, in normal conjunctiva (negative control) and in labial salivary glands of primary Sjögren’s syndrome patients biopsies (positive control). All slides were counterstained with Mayer’s haematoxylin. The arrows express the marked regions. Original magnification ×400.
In comparison with normal conjunctival specimens, we remarked with interest that, although significant levels of Bcl2 staining were predominantly found in the basal layer of the PECs, intense positivity was observed throughout the thickness of the epithelium layer (Figure 4(a) and (b)). Furthermore, our results have demonstrated an overexpression of Bcl2 surrounding the blood vessels and nearby regions area of the pterygium connective tissue (Figure 4(a)). In contrast, positive controls showed a strong expression of Bcl2 in the connective tissue in association with the basal layer of the epithelial cells (Figure 4(c)).

Immunohistochemical staining forBcl2 in pterygium (a), normal conjunctiva (b, negative controls) and conjunctivitis (c, positive controls) biopsies. All slides were counterstained with Mayer’s haematoxylin. The arrows denote the marked regions. Original magnification ×400.
IL-17A, IL-6, IL-10 and NO production in the tears and sera of patients with pterygium
As illustrated in Figures 5–8, our results indicate that in all cases, the pro-inflammatory cytokines (IL-6 and IL-17A) and NO levels are considerably higher in patients with pterygia compared with the healthy controls. Interestingly, cytokines and NO levels are more pronounced in the tears of the patients compared to their sera (IL-6 tears vs IL-6 sera: 69.90 ± 5.59 pg/mL vs 7.66 ± 0.71 pg/mL, respectively; IL-17A tears vs IL-17A sera: 242.20 ± 24.24 pg/mL vs 18.04 ± 2.15 pg/mL, respectively; NO tears vs NO sera: 106.7 ± 9.02 µM vs 58.28 ± 7.40 µM, respectively). Therefore, these findings indicate that inflammatory mediators were abundantly present in the tears. Additionally, we observed that the concentrations of IL-10 (an immunoregulatory cytokine) were lower in the patients’ tears and sera; however, this difference was not statistically significant (Figure 6). Therefore, the concentration of IL-10 appears to be insufficient to regulate the enhanced inflammatory process.

Concentration of IL-6 in the tears (a) and sera (b) from patients with pterygium.
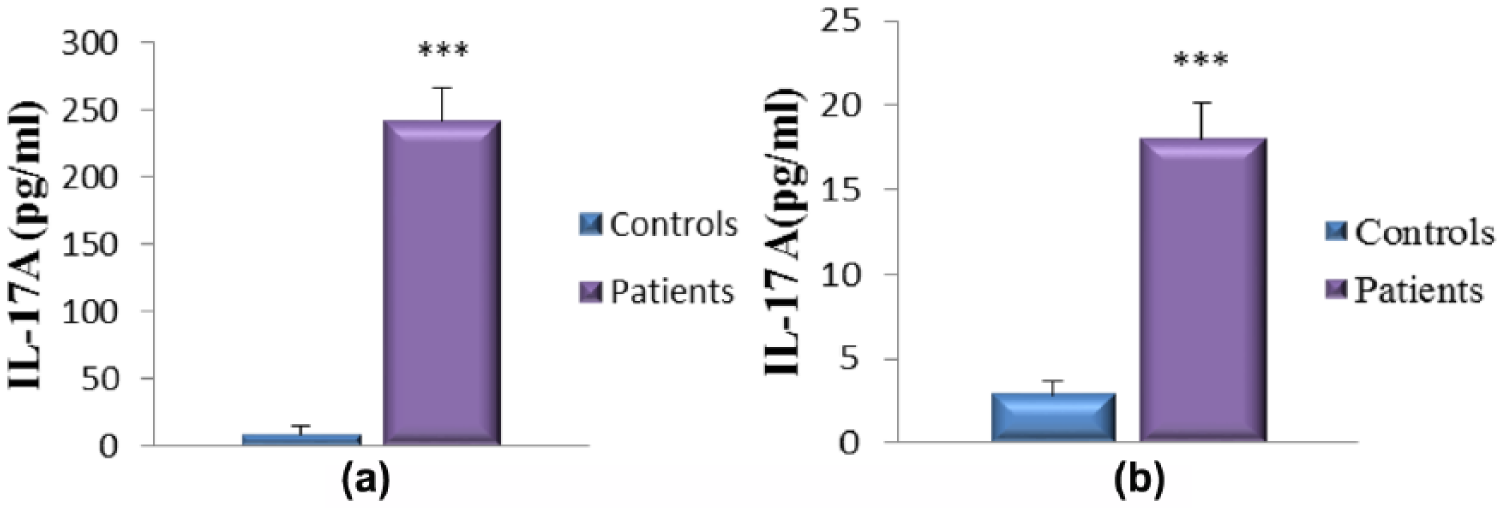
Concentration of IL-17A in the tears (a) and sera (b) from patients with pterygium.
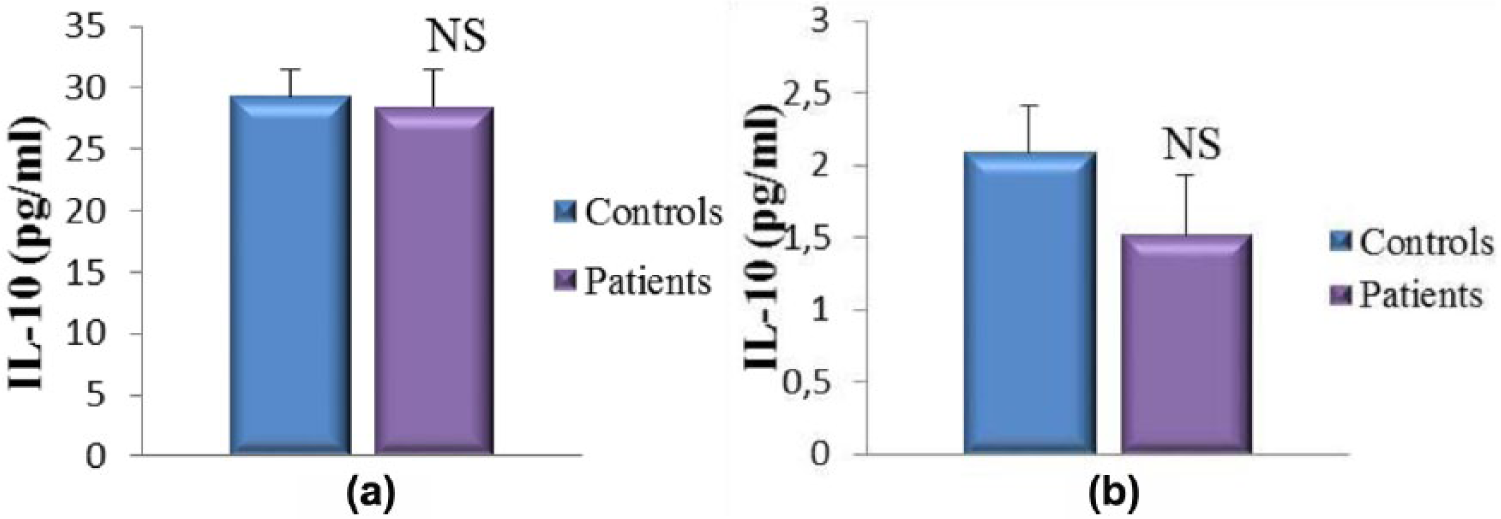
Concentration of IL-10 in the tears (a) and sera (b) from patients with pterygium.

Total nitrite concentration in the tears (a) and sera (b) from patients with pterygium.
Discussion
Pterygium can be described as a benign uncontrolled growth of the conjunctive tissue that covers the sclera. However, pterygium can severely alter visual function in advanced cases and become inflamed, leading to redness and irritation of the affected area. 22 It appears that pterygium is due to an inflammatory and degenerative process that stems from uncontrolled cellular proliferation of the subconjunctival and fibrovascular linking tissue on the cornea. 2
The main aetiological factor incriminated in pterygium apparition is UV radiation exposure. The molecular mechanism by which tissue proliferation is induced remains unclear. Indeed, it has been reported that a mutagenesis caused by the effects of reactive oxygen species generated by UV light could constitute the principal trigger of non-cancer ocular diseases, including pterygium. 2 Chronic exposure to UV light has been shown to trigger excessive production of NO through a photochemical reaction. 23 Previous studies showed controversial results concerning the presence of this radical in pterygium. Ozdemir et al. 24 in 2005 found reduced NO levels in patients biopsies while the groups of Balci et al. 25 in 2011 and Kormanovski et al. 23 in 2014 showed increased tissue content. Our study, although realized in both sera and tears, is in accordance with the last study. Moreover, this result is comforted by the local expression of NOS2, suggesting a local production of NO in tissue. This presence was reported previously concomitantly with vascular endothelial growth factor (VEGF), one of the main actors in pterygium pathgenesis.26,47,48 These data indicate that NO might be an important factor in the development of pterygium. This radical is likely to cause changes in the oxidant/antioxidant status of the human cornea. 23
NOS2 expression is triggered by several immunological factors as pro-inflammatory cytokines. The main transcriptional factor responsible for this induction is NF-κB. 27 In our study, we showed an important expression and activation of this factor in situ. NF-κB expression was broader than NOS2 expression since it affected all tissue parts. Our results comfort previous reports of NF-κB activation both in vivo and in vitro in cultured epithelial cells showing increased Iκ-Bα phosphorylation.28,29 Thus, the NF-κB pathway appears to play a key role in pterygium inflammation, which enhances pterygia pathogenesis and its progression.
NF-κB regulates gene expression implicated in different processes, including cellular proliferation, inflammation and other immune responses and activating genes associated with pro-inflammatory cytokines. 30 The activation of NF-κB in pterygia could be one of the mechanisms involved in the inflammatory responses that enhance pterygia pathogenesis and its progression. 31 The concomitant increase in the local NO and IL-6 secretion with NF-κB expression suggests a relationship between IL-6, NOS2 expression and NF-κB activation in pterygia tissue. In fact, it has been reported that several inflammatory markers can be induced following in vitro activation of NF-κB in primary pterygium fibroblasts.7,28
In the same line, several in vitro studies showed the induction of pro-inflammatory cytokines by corneal limbus epithelial cells. 32 This response seems to be triggered in vivo since the same profile can be observed in pterygium biopsies. 33 The high levels of IL-17A, IL-6 and NO assessed in the tears of patients with NOS2 and NF-κB expression in pterygium sections suggest a probable association of the local inflammatory response with pterygium disease. This hypothesis is supported by the high levels of inflammatory infiltrates observed in pterygium samples. In addition, we observed a high expression of CD3 and CD68 associated with the elevated levels of IL-17A, IL-6 and NO in tears of patients with pterygium compared to the healthy controls. Our data suggest that IL-17A and IL-6 production may contribute to the up-regulation of NOS2 expression in patients, thereby inducing the inflammatory process. It appears that the inflammatory response is likely regulated via the Th17 pathway. Our findings are supported by recent studies demonstrating IL-17A expression in the inflammatory regions of pterygium sections.34,35 There is increasing evidence that IL-17A is a strong mediator of the inflammatory response in various tissues.9,36,37 In particular, IL-17A promotes the pro-inflammatory responses induced by IL-1β and TNF-α.36,37 Moreover, IL-17A is primarily produced by Th17 populations differentiated upon exposure to combinations of IL-1β, IL-6 and TGF-β.9,38–40
We postulate that UV light exposure may have provided the initial trigger that stimulates epithelial and immune cells to produce IL-17A, IL-6 and NO. Indeed, a previous study reported the possible implication of ultraviolet B (UVB) radiation in the synthesis of two inflammatory cytokines (IL-6 and IL-8).7,41 Therefore, the production of such mediators may initiate a cascade of events that includes inflammation, proliferation, angiogenesis and antiapoptosis. 42 Furthermore, cytokines have been shown to stimulate the expression of matrix metalloproteinases (MMPs) and their tissue inhibitors, tissue inhibitors of metalloproteinases (TIMPs), implicating the involvement of these pathways in tissue remodelling, which could contribute to pterygium formation and/or progression.43,48
The maintenance of cellular homeostasis is essentially regulated by two processes: (1) cellular proliferation and (2) cellular apoptosis. 44 To assess whether enhancement of anti-apoptotic pathway is involved in the development of pterygia, we evaluated the expression of the anti-apoptotic protein, Bcl2, in pterygia and normal bulbar conjunctival biopsies (negative controls). Our findings revealed that Bcl2 is overexpressed in pterygia sections compared to the negative controls. These results are corroborated by previous research conducted by Tan et al. 4 and Liang et al. 46 Bcl2 positivity in the pterygium biopsies may play an important role in the disease. It is likely that pterygium is related to tumours which form in response to inflammatory processes.45,46 Based on the findings of this study, in addition to NOS2 up-regulation with pro-inflammatory cytokines, we suggest the potential role of IL-17A and IL-6 in the induction of inflammation and anti-apoptotic events. The concomitant expression of Bcl2 in pterygia biopsies observed in our study sustains this hypothesis and indicates a relationship between inflammation and anti-apoptotic processes in pterygium pathogenesis, thereby enhancing both tissue damage and dysregulated cellular proliferation.
Footnotes
Acknowledgements
The authors thank the patients, controls and the surgical staff of the Ophthalmological Department of the Ibn Rochd University Hospital, Annaba, Algeria. The authors express gratitude to the Department of Anatomopathology of the Ibn Rochd University Hospital, Annaba, Algeria, and Dr Sarah Benchabane for her technical help in particular for immunohistological analysis of positive controls.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
This work was supported by the Algerian Agency of Research Development in Health (ATRSS).