
Abstract
The study aim was to investigate the effect of dichloroacetate (DCA) on thymus and the thymocyte cycle in rats. Wistar male gonad-intact and castrated rats (4–5 weeks) were investigated in the following groups: (1) control; (2) treated with DCA; and (3) treated with the DCA and sodium valproate (NaVP) combination. Rats were treated for 4 weeks with DCA 200 mg/kg/day alone and 300 mg/kg/day of NaVP plus 200 mg/kg/day of DCA (every second week, beginning with NaVP). After the experiment, the thymus was weighted, and the thymus lobe was taken for thymocyte flow cytometry. In gonad-intact rats, the thymus weight of the control was higher than in rats treated with DCA (P <0.001) or with the NaVP–DCA combination (P <0.04); a comparison of thymus weight between DCA- and NaVP–DCA-treated groups revealed a higher thymus weight loss in the DCA-treated group (P <0.03). Flow cytometry shows that DCA treatment increased the percentage of cells in the G2–M phase (P <0.03) and reduced in G1–G0 (P <0.02). The DCA treatment effect was determined only in gonad-intact but not in castrated rats. The authors discuss the possible DCA and NaVP interaction mechanisms.
Sodium dichloroacetate (DCA) is a pyruvate dehydrogenase kinase (PDHK) inhibitor. DCA is a small molecule of 150 Da, rapidly absorbed from the gastrointestinal tract. DCA is transported across cell membranes by the monocarboxylate transporter system and gains entry into the mitochondrial matrix via the mitochondrial pyruvate transporter. 1 The DCA principal site of action is the pyruvate dehydrogenase (PDH) complex inhibiting PDHK, keeping PDH in the unphosphorylated catalytically active form and facilitating the oxidative removal of pyruvate.1,2 PDH is present in all tissues. PDHK has been suggested as a specific target in proliferating cells such as cancer cells. 3 DCA has been employed as an investigational medicine for indication in chronic treatment seeking to decrease the blood lactate acid level in congenital lactic acidosis, 4 to inhibit anaerobic glycolysis which renders most cancer cells resistant to apoptosis induction in cancer research. 3
The thymus of gonad-intact and castrated rats could be a valuable model for investigating the impact of medicinal products on thymocyte proliferation. Castration of rats induced a hyperplasia of thymus (proliferation of thymocytes) and NaVP has been shown to reduce thymus weight.5,6 We tested the influence of DCA as well as of the sodium valproate (NaVP) and DCA combination on thymus weight and thymocyte cycle as both medications are known to have an effect on cell proliferation inhibition.3,5 The present study showed a significant thymus weight loss in gonad-intact DCA- and NaVP–DCA-treated male rats and a significant decrease of thymocytes in the G0–G1 phase, as well as thymocyte accumulation in the G2–M phase, but such changes are not characteristic of castrated males treated with a DCA or a NaVP and DCA combination.
Materials and methods
Experimental design
Male Wistar rats (gonad-intact; n = 18; 4–5 weeks old) and castrated age-matched rats (n = 18) were investigated. Gonad-intact male rats were randomly divided into three groups (n = 6 in a group): (1) control; (2) treated with DCA; and (3) treated with NaVP and DCA. In the same three groups (n = 6 per group), castrated male rats were investigated. Rat castration was performed by removing the testes at the age of 4 weeks (in the peripubertal period − 28 ± 2 days). The effect of castration and treatment on thymus in rats was assessed. The permission of the State Food and Veterinary Service of Lithuania to use experimental animals for research was obtained (2015-05-18 No. G2-28). The animals were purchased from the Animal Facility of the Lithuanian University of Health Sciences, Veterinary Academy (Kaunas, Lithuania). The animals were housed in standard colony cages with free access to food; they were housed under conditions of constant temperature (21 ± 1°C), humidity, and a light/dark cycle (12/12 h). A commercial pellet diet was provided ad libitum. The experiments were performed in compliance with the relevant laws. The animals were housed in the described conditions and acclimated for at least 5 days before the experiment.
At 5 weeks of age, rats were treated with DCA 200 mg/kg/day in drinking water and 300 mg/kg/day of NaVP plus 200 mg/kg/day of DCA (every second week, beginning with NaVP and ending with DCA) aqueous solutions for 4 weeks. The DCA and NaVP solutions were offered to animals ad libitum as their only source of drinking (in aluminum foil-wrapped bottles to avoid light decomposition). Control groups were given fresh drinking water provided ad libitum. The treatment of castrated animals was started 1 week after castration (at 5 weeks of age). After 4 weeks of treatment, animals were killed in a 70% CO2 camera. The thymus was separated and weighted, and its one lobe (right) was taken for the thymocyte flow cytometry analysis.
NaVP and DCA were purchased from Sigma-Aldrich GmbH, Germany.
Thymocyte preparation
To minimize contamination with red cells, the carotid arteries were cut and the animals exsanguinated. Thymocytes were isolated from the thymus: the thymus was removed with forceps and cleared of blood vessels. Contaminating blood was removed by rinsing with RPMI-1640 (Biological Industries, Israel). The thymus was then minced and pipetted vigorously with a syringe several times. Large cell aggregates and connective tissue were removed by passage through two layers of surgical gauze. The resulting suspension of lymphocytes was washed free of red blood cell contamination by centrifugation two times in RPMI-1640 at 400 × g for 8 min each, and the supernatant was decanted. Thymocytes thus isolated were resuspended in RPMI-1640 supplemented with 10% (v/v) fetal calf serum (CE-Immundiagnostika GmbH, Germany) 100 µg/mL penicillin and 100 U/mL streptomycin (Sigma-Aldrich, Italy). Total counting of thymocytes was made in Fast-Read 102 chambers (Biosigma, Italy) stained with eosin (Reagena, Finland).
Flow cytometry analysis
The cell cycle analysis of control and DCA, NaVP–DCA-treated rat thymocytes was performed using an APC-BrdU flow kit (#552598) from BD Bioscience Pharmingen, US, according to the manufacturer’s instructions: thymocyte suspensions were labeled in vitro with BrdU (10 µM) added directly to the RPMI-1640 medium supplemented with fetal calf serum, penicillin, and streptomycin, for 24 h, at 37°C in a humidified atmosphere with 5% of CO2. The cells were then fixed, permeabilized, and treated with DNase (300 µg/mL, 1 h, 37°C) before staining with anti-BrdU-APC monoclonal antibodies and the 7-AAD reagent as recommended by the manufacturer’s protocol. The cells were analyzed on a FACSCanto (Becton Dickinson, USA) for APC and 7-AAD fluorescent dye, respectively. In total, 10,000 events were counted for each sample.
Statistical analysis
Statistical analysis was performed by using the Statistical Package for the Social Sciences, version 22.0 for Windows (IBM SPSS Statistics V22.0, USA). The normality assumption of data was verified with the Kolmogorov–Smirnov test. The results are presented as the mean ± standard deviation (SD). When the normality assumptions are not met, data are expressed as a median and a range (minimum and maximum values). Differences between two independent groups were evaluated using the non-parametric Mann–Whitney U-test. The one-way ANOVA analysis was used to determine significance among the groups and post-hoc tests with Fisher’s least significant difference (LSD) were used for comparison between the individual groups. Spearman’s rank correlation coefficient (r) was used to assess relationships between thymus weight and rat body weight. Differences at the value of P <0.05 were considered significant.
Results
The effect of castration on thymus
The mean age of rats at the end of the experiment was 60 ± 3 days and did not differ significantly among the groups. Statistically significant difference was found in the thymus weight between the intact male control and castrated male control groups (Figure 1): the thymus weight in castrated males significantly increased, i.e. castrated males showed a progressive thymus hyperplasia. No significant difference was found when comparing the rat weight of gonad-intact and castrated male rat groups (255.89 ± 48.30 and 245.12 ± 17.53; P >0.05). The weight of DCA-treated castrated rats was significantly lower than of their control (208.17 ± 22.77 and 245.12 ± 17.53; P <0.02). No statistically significant correlation was found among gonad-intact and castrated control rats’ weight and their thymus weight in all treated groups.

Investigation data of DCA and NaVP–DCA treatment effect on gonad-intact and castrated male rat thymus.
The effect of treatment on thymus
Comparing the thymus weight of the gonad-intact DCA-treated rat group with their control, the thymus weight was found statistically different—the treatment with DCA caused a significant decrease of thymus (Figure 1). A comparison of thymus weight of gonad-intact control and NaVP–DCA-treated rats showed a significant thymus weight loss after the NaVP–DCA treatment. A comparison of thymus weight between gonad-intact DCA-treated and NaVP–DCA-treated groups revealed a significantly higher thymus weight loss in the DCA-treated rat group. A moderate and statistically non-significant correlation between rat body weight and thymus weight was found in rat groups treated with DCA and in those treated with DCA and NaVP combination (P >0.05); also, statistically non-significant differences were found comparing rat weight among the gonad-intact control, DCA-treated and NaVP–DCA-treated rat groups (P >0.05).
No significant effect of DCA as well as of NaVP–DCA treatment combination was found on thymus weight of castrated rats (P >0.05). A comparison of thymus weight among castrated rats treated with DCA, treated with a DCA and NaVP combination, and respective groups of gonad-intact treated rats shows a significantly lower thymus weight in gonad-intact DCA- and NaVP–DCA-treated groups (Figure 1).
Thymocyte cycle analysis
Flow cytometry analysis was performed, and the baseline percentages of cells in G1–G0, S, pre-G, and G2–M phases are shown in Table 1. No statistically significant difference was found between the gonad-intact male control and castrated male control groups. A more pronounced effect was observed in the gonad-intact DCA-treated male group. The DCA induced a significant increase in the percentage of cells in the G2–M phase (P <0.03) and reduced the percentage of the G1–G0 (P <0.02), respectively, compared with the control group. The cell percentage in the S phase was in the range of 5.5–7.15, and there was no significant fluctuation among the groups (Table 1). No significant changes in the cell cycle distribution were observed in the castrated DCA- and NaVP–DCA-treated groups when compared with the castrated control.
Thymocyte cycle of the thymus of investigated rat groups.
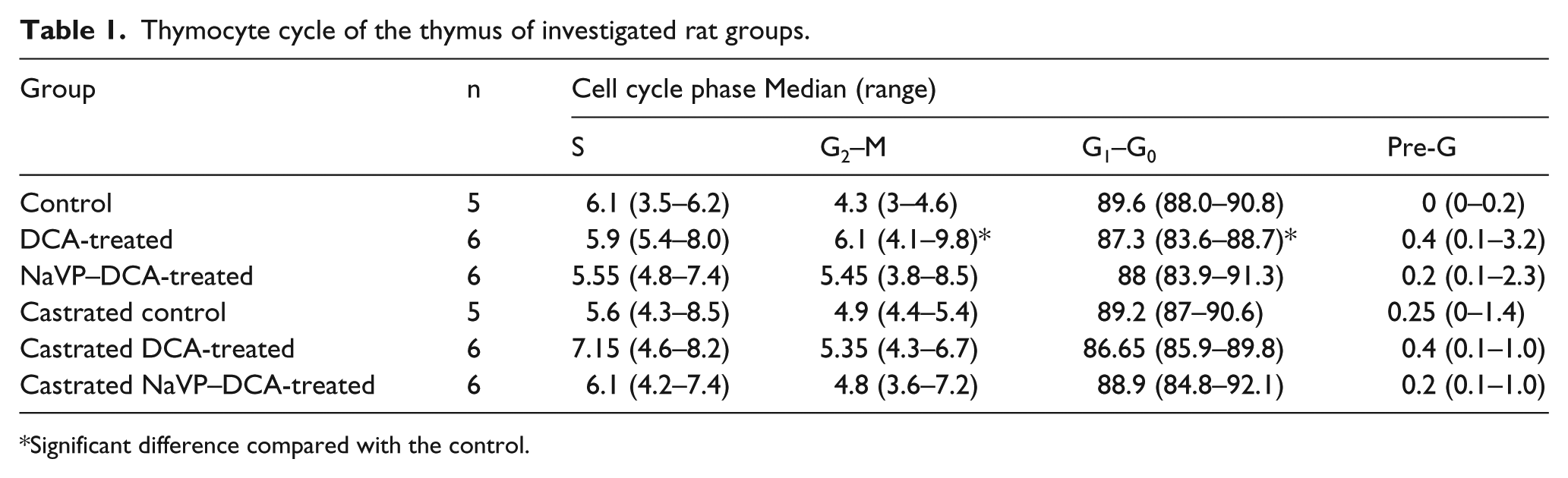
Significant difference compared with the control.
Discussion
The DCA as well as the NaVP–DCA treatment caused a significant reduction of thymus weight in the gonad-intact rat groups. The described thymus weight loss could be related to the effect of DCA on the thymocyte cycle: the DCA induced a significant increase of thymocyte percentage in the G2–M phase and reduced the percentage of the G0–G1 phase. A similar effect of DCA, causing the analogous G2–M phase arrest in multiple myeloma cell lines, was reported. 7 The possible explanation of this phenomenon might be the medication toxicity related to the oxidative stress. The DCA has been described to increase the reactive oxygen species generation with a concomitant cellular shift from glycolysis to oxidative metabolism, resulting in an increased apoptosis. 8 DCA leads to p53 accumulation, and the p53-dependent pathway regulates the G2–M transition in response to genotoxic stress. 9
A comparison of gonad-intact thymus weight between DCA- and NaVP–DCA-treated groups revealed a possible DCA and NaVP interaction. The lower or no effect of the combined NaVP–DCA therapy in gonad-intact and castrated male rats, respectively, could be related also to the pharmacokinetic interaction of DCA with NaVP. The DCA metabolism occurs primarily through glutathione (GSH) conjugation catalyzed by GSH S-transferases (GSTs) in the liver. 10 The NaVP treatment of rats increased α-GST levels in both serum and liver. 11 The elimination of DCA depends on the activity of glutathione transferase (GSTZ1), which catalyses the GSH-dependent dechlorination of DCA, and DCA is the main inactivator of GSTZ1. 12 So, in the study, NaVP treatment followed by DCA treatment could induce changes in DCA metabolism. The DCA and NaVP interaction mechanism could also be related to their effect on gonadal hormones. NaVP was associated with an increase in the testosterone level and with a reduction of estrogen serum levels. 13 Also, the intracellular chloride concentration could be an important factor for DCA metabolism. Chloride protects GSTZ1 from the irreversible inactivation by DCA. 14 NaVP increasing GABA-A receptor activity and thus decreasing the intracellular chloride level could have an impact on cell proliferation. 15 Also, the fluctuations in intracellular chloride could change the intracellular activity of DCA.
Conclusions
Gonadal hormones could be important for the effect of DCA on the thymus as well as on the thymocyte cycle regulation in male rats. An important finding of the study could be the possible pharmacokinetic interaction of DCA and NaVP.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This research was funded by the Research Council of Lithuania.