
Abstract
Regenerative medicine and stem cell therapy may represent the solution for the treatment of non-curable human diseases such as type 1 diabetes. In this context of growing demand for functional and safe stem cells, human amniotic epithelial cells (hAECs) from term placenta have attracted increasing interest for their wide availability, stem cell properties, and differentiation plasticity, which make them a promising tool for stem cell-based therapeutic applications. We initially assayed the stemness characteristics of hAECs in serum-free conditions. Subsequently we developed a culture procedure on extracellular matrix for the formation of three-dimensional (3D) spheroids. Finally, we tested the immunomodulation and differentiation potential of hAEC spheroids: the presence of pancreatic endocrine hormones was revealed with transmission electron microscopy and immunofluorescence analyses; the release of C-peptide in hyperglycemic conditions was assayed with ELISA.
The serum-free culture conditions we applied proved to maintain the basic stemness characteristics of hAECs. We also demonstrated that 3D spheroids formed by hAECs in extracellular matrix can be induced to differentiate into insulin-producing cells. Finally, we proved that control and induced cells equally inhibit the proliferation of activated mononuclear cells.
The results of this study highlight the properties of amnion derived epithelial cells as promising and abundant source for cell-based therapies. In particular we are the first group to show the in vitro pancreatic induction of hAECs cultured on extracellular matrix in a 3D fashion. We accordingly propose the outcomes of this study as a novel contribution to the development of future cell replacement therapies involving placenta-derived cells.
Introduction
Regenerative medicine and stem cell therapy represent promising tools for the treatment of non-curable human diseases such as type 1 diabetes (T1D), which is characterized by the autoimmune destruction of pancreatic beta cells, resulting in the lack of insulin. The impairment of glucose homeostasis functions causes hyperglycemia and serious long-term complications. The shortage of transplantable pancreatic islets and the side effects of prolonged insulin injection to treat T1D, 1 have stimulated research into expandable and renewable sources of insulin-producing cells (IPCs). Stem cells hold great promise for supplying an adequate amount of donor cells for replacement therapies. In this connection, human embryonic stem cells (hESCs) and induced pluripotent stem (iPS) cells represent a virtually unlimited source of transplantable cells. Many studies have reported the derivation of IPCs from hESCs 2 and iPS cells, 3 but their future clinical application is hampered by the handling complexity, high cost of procedures, safety issues, and ethical concerns. 4 Stem cells isolated from other tissues may surmount the issues related to the use of hESCs and iPS cells. Among them, placenta-derived stem cells are a promising source for cell transplantation and regenerative medicine. Human term placenta and fetal membranes are discarded after birth, so they can be easily obtained in large amounts without ethical concerns or invasive isolation procedures. Human amniotic epithelial cells (hAECs) are derived from human amniotic membrane and originate from epiblast at days 8–9 during embryological development, before gastrulation. 5 hAECs express markers associated with hESCs and pluripotent stem cells, 6 but unlike hESCs, hAECs are considered a safe alternative for cell therapies since they do not cause teratomas upon transplantation nor activation of the immune system. 7 Moreover, hAECs have been successfully induced to differentiate towards mesodermic lineage, 8 neural-ectodermic lineage, 9 hepatic-endodermic lineage, 10 and pancreatic-endodermic lineage. 11 For all these reasons hAECs are considered to retain most of the stem cell peculiarities of hESCs. In addition, the immune privilege of placental tissue grants hAECs the unique characteristic of being associated with poor antigenicity and immunomodulatory properties: in particular they inhibit mixed lymphocyte reactions as well as mitogen stimulated T cell proliferation in vitro12,13 and they do not induce immune reaction when transplanted in vivo. 7
Previous studies successfully described the isolation and differentiation of hAECs into IPCs using classic or stepwise differentiation protocols, 11 in the presence of media enriched with components of animal origin. The use of animal sera as growth supplement in human cell cultures might compromise the reproducibility and reliability of downstream assays and, therefore, could hamper the chance of clinical translation.14,15
Along with the requirements related to culture media composition, it is likewise worthy of consideration the adoption of culture methods that mimic the in vivo environment, such as three-dimensional (3D) culture systems. Several studies showed that stem cells cultured on 3D matrices enhance their ability to form higher-order structures like embryoid bodies, spheroids, and glandular-like acini.16,17 Moreover, 3D matrices enhance the stem cell potential and viability, while providing the suitable microenvironment for cell attachment, proliferation, and differentiation. 18 These effects are a direct consequence of the synergic cell-to-cell and cell-to-ECM interactions that cannot be provided in classical monolayer cultures.19,20 In particular, recent cell therapies studies for diabetes showed that 3D cultures could play a crucial role in the induction of hESCs into mature IPCs. 21
In this perspective, in the present study we validated the stemness and immunomodulatory properties of hAECs in defined serum free conditions. We successfully developed a serum-free 3D culture procedure, based on extracellular matrix (ECM) hydrogel, for multicellular hAEC spheroid formation. Finally, for the first time in the field of placental stem cell research, we demonstrated that hAECs cultured in 3D serum-free conditions, functionally produce newly synthesized insulin in response to glucose stimulation.
Materials and methods
Ethics
The study was approved by the Local Research Ethics Committee (Azienda Ospedaliero-Universitaria di Modena, prot. n 1171; 12/04/2010) and written informed consent was obtained from each healthy donor before specimen collection.
Isolation and expansion of human amniotic epithelial cells (hAECs) in basal growth media (BM)
Amnion membranes were retrieved from term pregnancies (37–40 weeks of gestation) delivered by Cesarean section. hAECs were isolated using a modified procedure of the protocol previously reported by Ilancheran et al.: 22 the amnion layer was mechanically peeled off the chorion layer and immediately washed in Hank’s Balanced Wash Solution without calcium and magnesium (HBSS, PAA Laboratories GmbH, Pasching, Austria) until blood clots were completely removed. The amnion was minced into small pieces (4 cm2 approximately) and incubated for 10 min at 37°C with 1X trypsin-EDTA (PAA Laboratories) to exclude debris. Membranes were then incubated for a second and third 40 min enzymatic digestion to release the amniotic epithelial cells. Single cell suspension was washed with Phosphate Buffered Saline (PBS, PAA Laboratories) and tested for viability with trypan blue dye (Biochrom AG, Berlin, Germany). Only samples with >90% viability were used for further assays.
Cell pellets were resuspended in serum-free medium (SFM), which consisted of Quantum286 (PAA Laboratories) serum-free medium for epithelial cells with 1% Penicillin-Streptomycin solution (PAA Laboratories) and epidermal growth factor (EGF) (10 ng/mL; Sigma-Aldrich, St. Louis, MO, USA), or serum-rich medium (SRM) made of DMEM (Invitrogen, Carlsbad, CA, USA) supplemented with heat-inactivated 10% fetal bovine serum (FBS) (PAA Laboratories), 1% Penicillin-Streptomycin solution (PAA Laboratories), and EGF (10 ng/mL; Sigma-Aldrich). The plating concentration in cell culture flasks was 80,000 cells/cm2. Before further assays, the cells were allowed to attach to the substrate and grow in culture medium until 80–90% confluency.
Cell proliferation assay
In order to evaluate cell proliferation potential, hAECs were plated at 60,000 cells/cm2 in 24-well plates and cell growth was assessed using Alamar Blue assay. Alamar Blue 10% (Invitrogen) was added to basal medium (BM) for 4 h and then fluorescence was measured using Victor Multilabel Counter (Perkin Elmer, Boston, MA, USA). The same procedure was repeated on days 3, 5, 8, 10, 12, and 15. Data were plotted as Fluorescence Arbitrary Unit (AU fluorescence) analyzed at specific time points.
Flow cytometric characterization of hAECs
Flow cytometric analysis was performed on primary cultures of hAECs. Cells were detached with Trypsin-EDTA and incubated with antibodies (1 µg/mL for 30 min at 4°C). The antibodies used were: anti-CD105-FITC (Beckman Coulter, Miami, FL, USA), anti-panCytokeratin-PE (Santa Cruz Biotechnology, Santa Cruz, CA, USA), anti-SSEA-4-APC (R&D Systems, Minneapolis, MN, USA), anti-OCT4 primary antibody (Santa Cruz Biotechnology), and secondary anti-mouse IgG-FITC (Beckman Coulter). Cells were then washed and analyzed on a flow cytometer (Navios FC, Beckman Coulter); acquired data were elaborated with Kaluza FC Analysis software.
RNA isolation, reverse transcription and real-time PCR (qPCR)
Total RNA was extracted from hAECs grown in culture flasks using the PureLink RNA Mini Kit (Invitrogen) according to the manufacturer’s instructions. cDNA was synthesized from 1–2 µg of total RNA using RETROscript First Strand Synthesis Kit and oligo dT (Invitrogen). Oligonucleotide pairs were designed using qPCR SciTools software (Integrated DNA Technologies). Primer sequences and gene accession numbers for stemness gene markers (Ki67, OCT4a, NANOG, SOX2) are listed in Table 1. qPCR assays were performed in a StepOne System (Applied Biosystems, Foster City, CA, USA) with KAPA SYBR FAST Master Mix (KAPA BIOSYSTEMS, Woburn, MA, USA). Data were normalized against the internal control beta-actin (ACTB).
PCR primers and gene accession number.

Immunomodulatory properties of hAECs
In order to evaluate the immunomodulatory potential of hAECs on activated peripheral blood mononuclear cells (PBMCs), hAECs were plated in 6-well plates (60,000 cells/cm2) in SFM or SRM, and were allowed to stabilize in culture for 7 days. Then, PBMCs were isolated from healthy donors by density gradient centrifugation (Ficoll-Paque 1077, Sigma-Aldrich) and plated on hAEC monolayer at a density of 2 × 106 cells/well in RPMI 1640 (Lonza, Walkersville, MD, USA). PBMCs were activated by addition of Phytohemagglutinin (PHA, 5 µg/mL, Sigma-Aldrich) and incubated for 72 h at 37°C, 5% CO2. Finally PBMCs were resuspended, recovered, and fixed with 70% ethanol at 4°C, stained with Propidium Iodide (Beckman Coulter) at room temperature for 10 min. The negative control consisted of PBMCs without PHA stimulation, while positive control was PBMCs with PHA (5 µg/mL) in the absence of hAECs. Cell cycle phase distribution of PBMCs was analyzed by flow cytometer.
Three-dimensional (3D) culture
3D culture of hAECs was performed according to the overlay method previously described by Debnath et al. 16 Briefly, isolated hAECs were first resuspended in SFM added with 2.5% reduced growth factor basement membrane extract (BME) from Engelbreth-Holm-Swarm (EHS) mouse sarcoma (Trevigen, Gaithersburg, MD, USA). Then, resuspended cells were seeded (60,000 cells/cm2) on a thick layer of 100% reduced growth factor BME measuring approximately 1 mm in thickness. Multicellular spheroids formed after 5–6 days, partially embedded in the gelled layer of basement membrane. Cell culture medium was changed every 3 days. hAEC spheroids were harvested and collected for further assays using Cultrex 3D culture cell harvesting kit (Trevigen Inc.).
In vitro pancreatic stepwise differentiation
Prior to differentiation, hAECs were allowed to grow for 5 days in SFM until the sufficient spheroid size. Pancreatic induction of 3D cultured cells was performed using a stepwise protocol divided into three stages: Stage 1: hAEC spheroids were pre-differentiated for 4 days in SFM supplemented with 100 ng/mL Activin A; Stage 2: SFM was supplemented with 100 ng/mL Activin A and Nicotinamide (NAM, 10mM) for 4 additional days; and Stage 3: terminal differentiation was performed switching the medium to SFM with NAM (10 mM) and Retinoic Acid (RA) (2 µM) for 10 days. Glucose concentration of SFM was ⩾15 mM. This hyperglycemic culture condition is suitable for epithelial cells growth and was maintained for pancreatic differentiation assay to enhance the induction of hAECs into IPCs. 23 Media supplements were all obtained from Sigma-Aldrich.
Immunofluorescence analysis and confocal microscopy
3D hAEC spheroids were fixed with 2% Paraformaldehyde (PFA, Sigma-Aldrich) for 20 min at room temperature. After washing 2 × 10 min in PBS, spheroids were permeabilized in 0.5% Triton X-100 (Sigma-Aldrich) in PBS for 10 min and then rinsed again 2 × 10 min in PBS. Antigen blocking was performed for 1 h at room temperature in blocking solution: 5% normal goat serum (Vector Laboratories, Burlingame, CA, USA), 0.2% Triton x-100, 0.1% Bovine Serum Albumin (BSA) (Sigma-Aldrich). Primary and secondary antibodies were diluted in freshly prepared blocking solution. Primary antibodies were incubated at 8°C overnight and secondary antibodies for 40–50 min at room temperature. The following primary antibodies and dilutions were used: mouse anti-insulin 1:100 (Abcam ab46707); rabbit anti-glucagon (Abcam ab8055); rabbit anti-C-peptide 1:100 (Cell Signalling #4593); rabbit anti-CK19 1:100 (Thermo Scientific PA5-16726); mouse anti-laminin 1:100 (Thermo Scientific MA1-21194). Secondary antibodies were: DyLight 594 goat anti-mouse 1:200 (Jackson Immuno Research Labs.); Alexa Fluor 488 goat anti-rabbit 1:400 (Molecular Probes); CF 488 goat anti-mouse 1:400 (Biotium); DyLight 594 goat anti-rabbit 1:200 (Thermo Scientific). Nuclei counterstaining was performed with Fluoroshield+DAPI (Sigma-Aldrich). Imaging was performed by confocal microscopy (Leica TCS SP2, Leica Microsystems, Mannheim, Germany) and images were processed using Leica Confocal Software.
Transmission electron microscopy (TEM)
Pancreatic induced hAECs cultured in 3D matrices and undifferentiated control cells were gently washed with Tyrode’s buffer, pH 7.3, and centrifuged at 10,000 x g for 5 min at 10°C. The resulting pellets were fixed overnight with 2.5% Glutaraldehyde (Electron Microscopy Sciences, Hatfield, PA, USA) in Tyrode’s buffer, postfixed for 2 h in 1% Osmium Tetroxide (Electron Microscopy Sciences), dehydrated, and embedded in Spurr resin (Electron Microscopy Sciences). Since cells were not homogeneously distributed within matrices, semi-thin sections were routinely stained with Toluidine Blue and observed with a Zeiss Axiophot light microscope (Carl Zeiss, Jena, Germany) to locate cells in each cut layer. Ultra-thin sections were performed on areas selected for the presence of spheroids and observed with a Jeol 2010 electron microscope (Jeol, Tokyo, Japan) at 200 kV after staining with Uranyl Acetate and Lead Citrate (Electron Microscopy Sciences).
Glucose-stimulated release and expression of C-peptide
To confirm the synthesis of insulin, C-peptide release from differentiated and undifferentiated hAEC spheroids was measured in hypoglycemic and hyperglycemic conditions. First, cells were incubated in Krebs-Ringer solution with bicarbonate and HEPES (KRBH; 129 mM NaCl, 4.8 mM KCl, 2.5 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, 5 mM NaHCO3, 10 mM HEPES, 0.1% (wt/vol) BSA) (Sigma-Aldrich) for 70 min as a preliminary washing step. Next, spheroids were incubated in KRBH with 2mM D-Glucose (Sigma-Aldrich) for 1 h and then switched to hyperglycemic stimulation condition consisting of KRBH with 20 mM D-Glucose and 30 mM KCl for 1 h. C-peptide levels in supernatant from basal (2 mM D-Glucose) and stimulation condition (20 mMD-Glucose + 30 mM KCl) were measured using the Human Ultrasensitive C-peptide enzyme-linked immunosorbent assay (ELISA) kit (Mercodia, Uppsala, Sweden) and normalized to total amount of cellular DNA using CyQUANT Cell Proliferation Assays Kit (Invitrogen). The fold stimulation was calculated as the ratio of the C-peptide concentration in supernatant from hyperglycemic condition to the C-peptide concentration from hypoglycemic condition. Glucose-stimulated spheroids were immediately assayed for the presence of C-peptide with immunofluorescence analysis.
Immunomodulatory activity of hAEC spheroids
The immunomodulatory activity of hAECs was analyzed after stepwise pancreatic-endocrine differentiation in 3D culture. PBMCs were isolated as previously described and plated on hAEC spheroids at the end of pancreatic differentiation protocol (hAECs/PBMCs ratio 1:2). PBMCs were activated by addition of Phytohemagglutinin (PHA) (5 µg/mL; Sigma-Aldrich) and hAEC spheroids/PBMCs co-cultures were incubated for 72 h at 37°C, 5% CO2. The negative control consisted of PBMCs without PHA stimulation and the positive control was PBMCs with PHA (5 μg/mL) in the absence of hAEC spheroids. Finally PBMCs were resuspended, recovered, and plated in 96-wells plate for BrdU incorporation analysis. BrdU incorporation was evaluated using a colorimetric immunoassay, according to the manufacturer’s instructions (Cell Proliferation ELISA, BrdU – colorimetric kit, Roche, Basel, Switzerland).
Statistical analysis
The assays were repeated a minimum of three times independently and the results were expressed as the mean ± SD. Statistical analysis was performed using two-tailed, paired T-test, or one-way ANOVA test. P <0.05 was considered to be statistically significant.
Results
hAEC epithelial morphology and proliferation rate
hAECs were isolated from term placenta and grew as adherent cells when seeded in classical serum-rich medium (SRM) or in serum-free medium (SFM). In both culture conditions hAECs regularly proliferated and formed a confluent monolayer of cells with epithelial cobblestone-shaped morphology (Figure 1a). In order to evaluate the culture medium influence on hAEC growth kinetics and proliferation ability, we performed a cell growth assay for 15 days. The resulting growth curve showed a similar proliferation rate for hAECs in both SRM and SFM (Figure 1b).
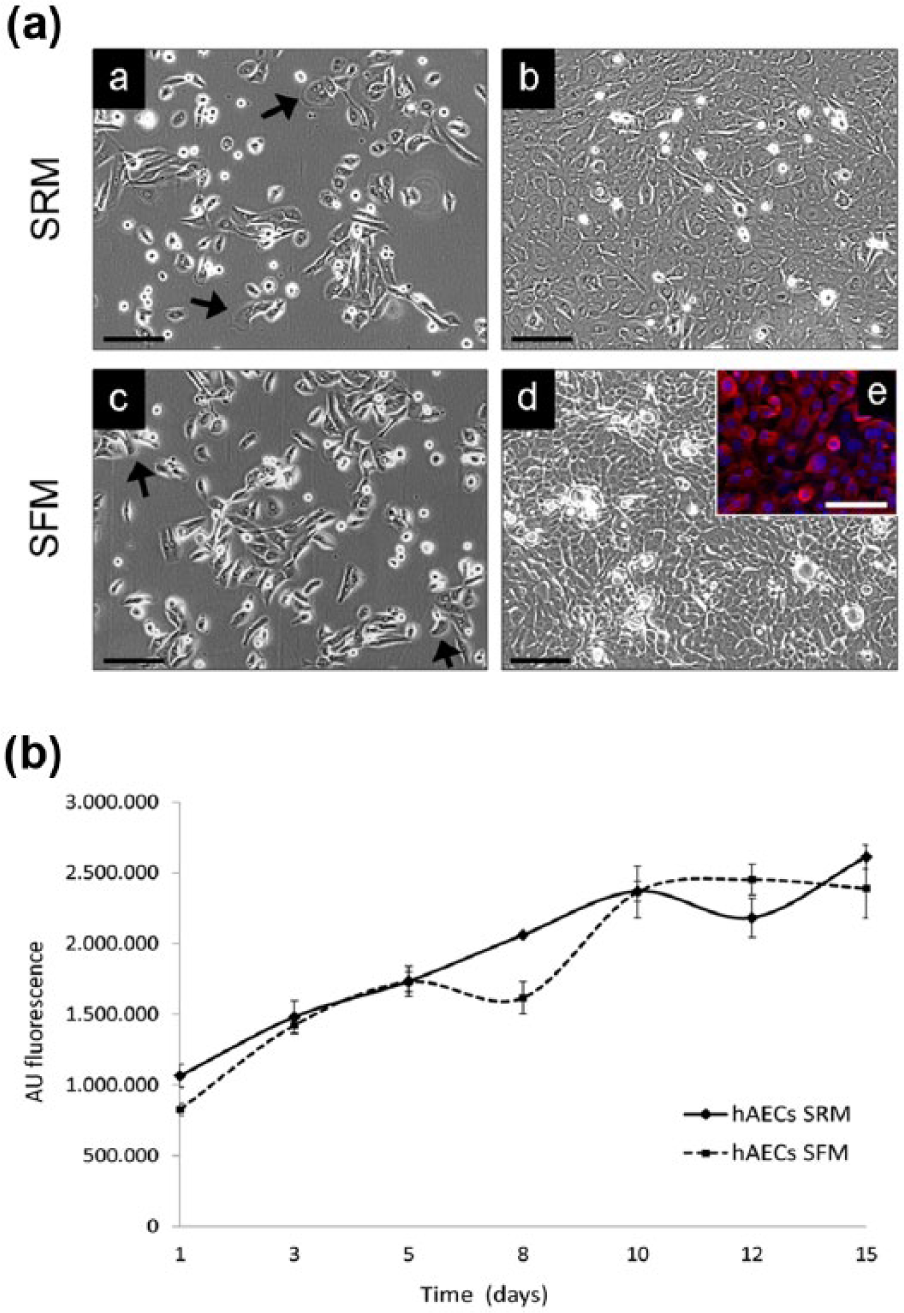
Essential characterization of SFM-cultured hAECs
To evaluate the effects of culture media, we tested whether serum deprivation could influence hAEC stemness features and immunomodulatory properties. First, we examined the expression of a selected antigenic surface pattern (Figure 2a). Flow cytometric analysis displayed a similar hAECs immunophenotype for both cell culture media: protein expression levels of stemness markers, OCT4 and SSEA-4, were clearly positive (OCT4: 99.1 ± 0.2% in SRM and 99.3 ± 0.3% in SFM; SSEA-4: 97.3 ± 0.4% in SRM and 98.9 ± 0.2% in SFM); characteristic mesenchymal antigen CD105 was expressed (94.5 ± 0.5% in SRM and 93.7 ± 0.6% in SFM) as previously reported; 8 distinctive epithelial marker PanCK was uniformly expressed in the hAECs population (99.9 ± 0.1% in SRM and 99.9 ± 0.1% in SFM). These data proved that SFM culture does not affect the immunophenotype of hAECs.

Moreover, qPCR was carried out to test for serum-induced changes in the expression of proliferative gene Ki67 and stemness markers NANOG, SOX2, and OCT4a. Ki67 mRNA decrease in serum-free medium is not significant, while gene expression levels for stemness markers were clearly and significantly higher in serum-free conditions than in serum-rich media, namely 70-fold for NANOG, 36-fold for SOX2, and 12-fold increase for OCT4a mRNA (Figure 2b).
Finally, to assess hAEC immunomodulatory ability in serum-free culture conditions, PHA-stimulated PBMCs were co-cultured with hAECs in SRM and SFM, and their proliferation was assayed to test hAEC immunomodulatory properties. The distribution of PBMCs in the cell cycle phases was evaluated (Figure 2c). Non-activated PBMCs were all in G1-G0 phase (96.4 ± 0.3%), while activated PBMCs in the absence of hAECs significantly decreased in the G1-G0 phase (68.7 ± 3.3%) and increased in the S (18.6 ± 0.5%) and G2-M (12.1 ± 2.0%) phases (P <0.05). After co-culture with hAECs in SRM or SFM, we observed a significant increase of PBMCs in G1-G0 (83.0 ± 4.1% in SRM and 82.7 ± 4.9 in SFM) and an equally sizable reduction of PBMCs in the S (11.5 ± 1.7 in SRM and 12.0 ± 0.6% in SFM) and G2-M (4.4 ± 2.6% in SRM and 3.7 ± 2.6% in SFM) phases (P <0.05). We may assume from these observations that hAECs cultured in either SRM or SFM, similarly reduce the proliferation of stimulated PBMCs.
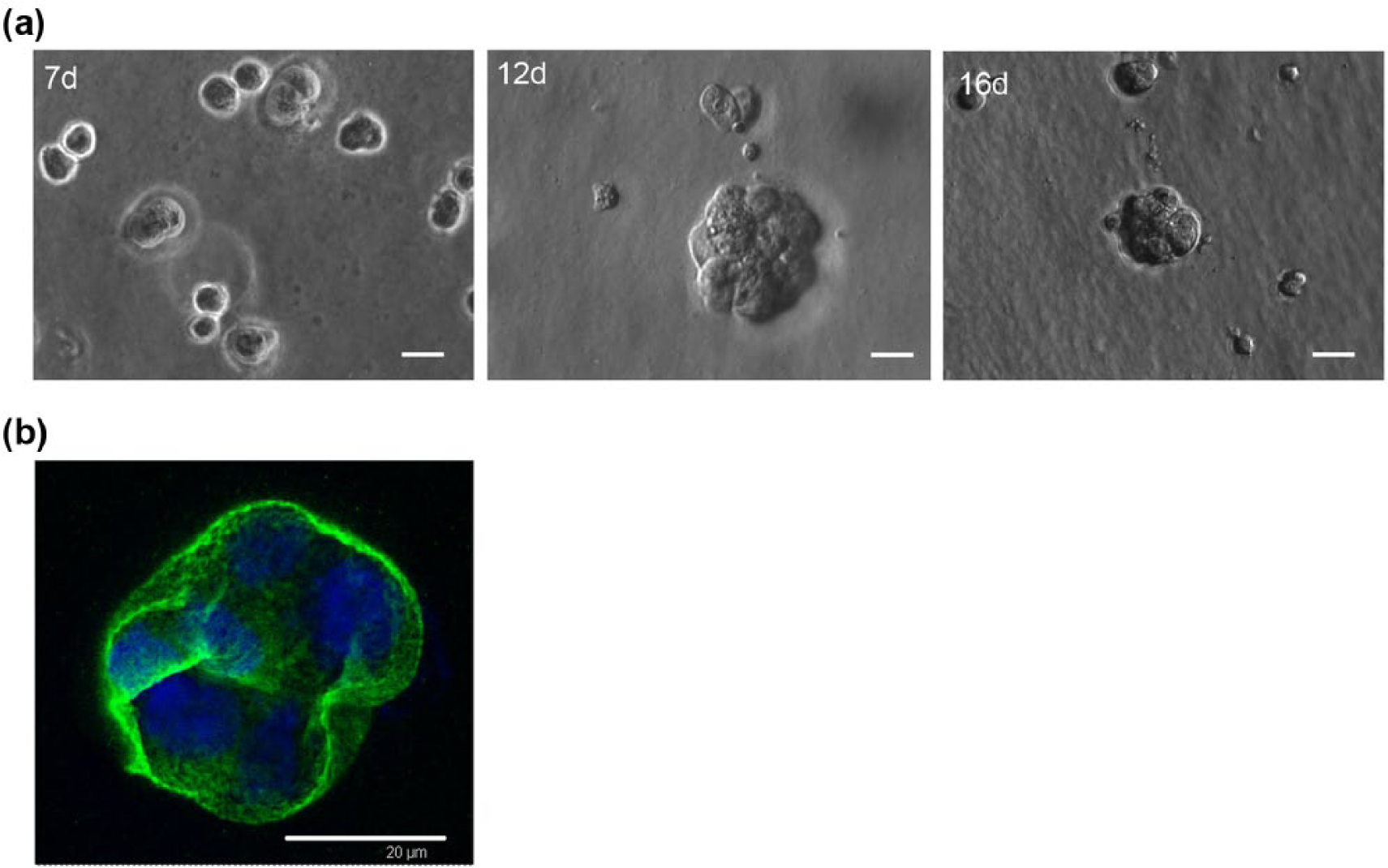
Morphogenesis of hAEC spheroids grown in 3D culture
When cultured on a thick bed consisting of 100% gelled basement membrane extract (BME), hAECs adopted a spheroid, 3D morphology (Figure 3a). After 1 week of culture, we observed the formation of multicellular structures which increased in complexity, size, and cellular density over time, reaching a complete and definitive spheroid architecture (> 40 µm) around days 10–12. The immunofluorescence analysis (Figure 3b) highlights the peculiar laminin localization at the cell mass outer perimeter, separating the spheroid from the surrounding extra cellular matrix. Laminin spatial arrangement, suggests the acquisition of a correct apico-basal polarization by hAEC spheroids. In addition, SFM was more effective in maintaining a stable spheroid morphology after 2 weeks of culture, while the serum-rich conditions caused the development of irregular and larger cell clusters (data not shown).
hAEC spheroids inhibit proliferation of mitogen-activated PBMCs
In order to investigate if hAEC immunomodulatory ability was maintained even after pancreatic differentiation in 3D culture system, we co-cultured hAEC differentiated spheroids with PHA-stimulated PBMCs and measured BrdU incorporation levels in PBMCs (Figure 4). Non-activated PBMCs do not incorporate BrdU (2.8 ± 1.6%), while PHA-activated PBMCs regularly incorporate high levels of BrdU (98.6 ± 1.4%). BrdU incorporation levels significantly decreased after co-culture with induced (8.1 ± 5.7%) or non-induced (8.5 ± 4.4%) hAEC spheroids (P <0.05). To conclude, no significant differences in inhibition activity on PBMC proliferation were observed between differentiated and non-differentiated hAEC spheroids.

Differentiated hAEC spheroids express pancreatic hormones and display islet-like ultrastructure
To determine whether our stepwise differentiation protocol efficiently induced hAEC spheroids into hormone-producing cells, we performed immunofluorescence analysis and checked for the presence of pancreatic hormones: glucagon and insulin (Figure 5a). Basal expression of glucagon was present in undifferentiated control cells (data not shown), but glucagon immunoreactivity became more intense upon differentiation. Basal expression of insulin was absent from undifferentiated hAECs. After 20 days of pancreatic induction in serum-free conditions, spheroids became insulin-positive. Confocal micrographs revealed the external localization of insulin-positive cells: merged images of immunofluorescence and phase-contrast micrographs showed the membrane-related position of insulin, while glucagon was exclusively expressed in hAECs localized in the central area of the spheroids. The peripheral arrangement of insulin is consistent with the exocytosis mechanism for the release of insulin granules in response to high glucose levels.

To assess the endogenous production of insulin and verify whether induced-hAECs formed mature secretory granules we used transmission electron microscopy (TEM) (Figure 5b). TEM ultrastructural analysis of induced hAECs (versus undifferentiated control cells) revealed the high complexity of the cytoplasm and the presence of several vesicles, consistent with intense protein synthesis. In particular, cells contained various secretory granules with a clear halo surrounding an electron-dense core, which is peculiar to mature insulin-containing granules.
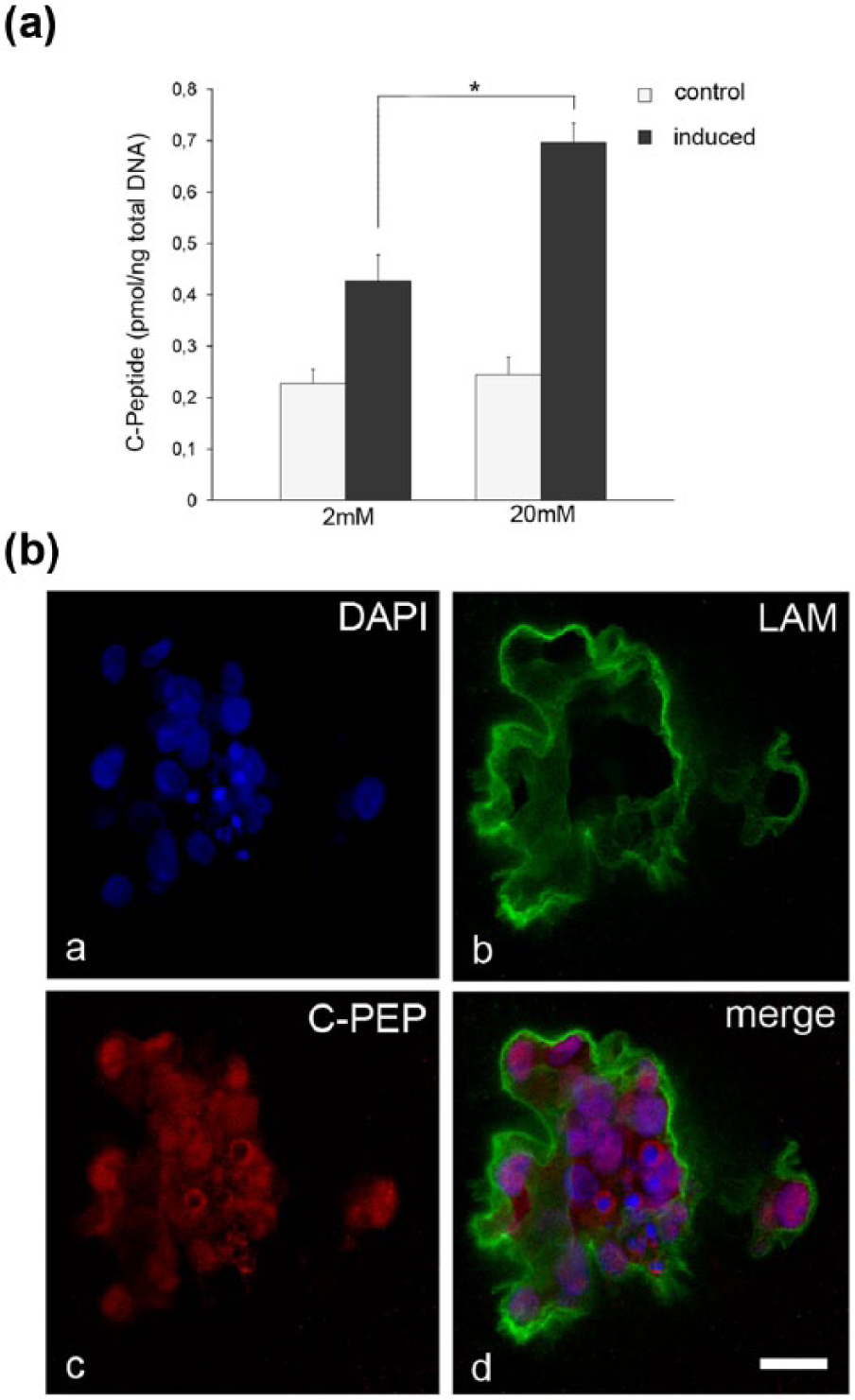
C-peptide release and expression in hAEC spheroids
To further confirm the endogenous nature of the detected insulin, C-peptide secretion assay was performed in differentiated and non-differentiated spheroids (control group) (Figure 6a). Basal secretion of C-peptide was measured in control group at 2 mM D-glucose and 20 mM D-glucose concentrations (0.227 ± 0.027 pmol/ng DNA and 0.244 ± 0.033 pmol/ng DNA, respectively). Pancreatic-endocrine differentiated spheroids secreted higher levels (P <0.02) of C-peptide in 20 mM D-Glucose condition (0.697 ± 0.036 pmol/ng DNA), than differentiated spheroids in 2 mM D-glucose condition (0.427 ± 0.050 pmol/ng DNA). These data further support the success of the differentiation protocol and show the acquisition of a glucose-stimulated insulin release mechanism from hAEC spheroids. Subsequently, immunofluorescence analysis performed on differentiated hAEC spheroids, revealed a clear and widespread presence of C-peptide-positive cells (Figure 6b).
Discussion
In this study, we propose a novel set of procedures to induce placental-derived hAECs into insulin-producing cells (IPCs), combining different cell culture approaches in order to overcome issues such as the presence of animal serum in the media composition and cell differentiation inefficiency.
Effective and safe cell therapies require strictly standardized and GMP operating methodologies, including optimal clinically compliant culture procedures.14,15 Most of the current in vitro data obtained from studies on stem cells were generated under artificial conditions using standard culture media containing FBS, which may impair cell culture assay reproducibility, alter any experimental outcome, and delay the clinical translation.
In this regard, we first validated serum free culture condition from cell isolation to cell differentiation: we concisely confirmed the outcomes from previous studies6,8 and extended the observations to our serum-free conditions. The unaltered epithelial morphology, the positive immunophenotype profile of key stemness markers (OCT4, SSEA-4), the growth kinetics, and the over-expression of stemness genes (NANOG, SOX2, OCT4a), all confirmed that hAEC features and stemness potential are preserved in serum-free conditions. In particular, we proved to maintain a strong proliferation rate in serum-free conditions without affecting the gene expression profile of proliferation marker Ki67.
In the second phase of our study, we focused on the generation of 3D spheroids from hAECs cultured on ECM. The vast majority of cell culture assays are still performed on synthetic substrates which force cells to adapt to a flat and rigid surface. Monolayer cultures compared to 3D cultures present several limitations concerning: viability, morphology, proliferation, response to stimuli, gene and protein expression, and differentiation efficiency.24–26 Hence, two-dimensional (2D) culture systems scarcely reproduce the in vivo environment and may, thus, have limited predictive value for clinical applications. A more physiological context is afforded by 3D culture methods which are more suitable for creating an adequate number of intercellular contacts and maintaining the in vivo phenotype and functionality.27,28 In the last decade, several reports described the generation of higher-order structures (embryoid bodies, glandular-like acini) from stem cells grown in matrices or other 3D supports.16,27,29 These studies have consistently increased our knowledge of several biological issues such as organ/tissue morphogenesis, cell growth, cancer development, and drug resistance. 30 Therefore, despite the complex handling of 3D culture procedures, assays performed on a 3D system hold higher consistency than any 2D approach. 19 In the present study we successfully show, for the first time, the generation of 3D spheroids obtained from placenta-derived primary epithelial stem cells. hAEC spheroids micrographs showed the typical round-shaped cell aggregates. Laminin expression revealed the encircling distribution of the basement membrane around spheroids, which suggests the correct acquisition of epithelial apico-basal polarity. The correct establishment and maintenance of cell polarity are crucial for normal cell physiology.
At this stage, we tested whether in vitro 3D culture was a valid approach to expand and efficiently differentiate hAECs into IPCs. Stepwise differentiation protocols applied to monolayer culture systems still remain the most adopted solutions to obtain IPCs from hESCs, iPS cells3,31,32 and mesenchymal stem cells (MSCs). 33 Recently, Takeuchi et al., 21 showed that pancreatic-endocrine differentiation of hESCs cells cultured in monolayer, produced insulin-positive cells unable to secrete insulin in response to glucose stimulation, while 3D culture cell aggregates became functional IPCs. Although previous studies reported considerable evidence about successful insulin production from different stem cells, 34 setting up an in vitro procedure to differentiate hAECs into IPCs has been a substantial task. We initially had performed direct differentiation with Nicotinamide as others reported,6,35 which only led to sporadic expression of insulin (data not shown) and no insulin secretion in response to glucose. Therefore, to emulate the in vivo pancreatic development pathway, we adapted, for our study on hAECs, the stepwise differentiation protocols from previous reports.11,31,32 SFM was used throughout the entire differentiation process and only three induction molecules were employed: Activin A, in the pre-induction phase, to drive the endoderm formation; 36 Nicotinamide (NAM), with Activin A in the second stage, to induce pancreatic-endocrine differentiation; 37 and finally RA with NAM to promote IPC formation. 38 Differentiated hAEC spheroids displayed insulin in the outer part of the spheroid and no consistent trace in the central core, where cells displayed an alpha cell phenotype (glucagon expression). Confocal micrographs revealed a defined cell membrane-associated localization of insulin. Here, as shown by TEM analysis, mature insulin-containing granules are gathered and eventually release their hormone content. C-peptide analysis undoubtedly confirmed the synthesis of insulin and revealed the glucose-dependent insulin secretion. The data showed the presence and production of insulin from a relatively limited number of hAECs (60,000 cells plated for each condition) and the fold increase of hormone produced by differentiated hAEC spheroids is comparable to analogous in vitro studies performed with adult stem cells. 39
One of the most important characteristics required from candidate stem cells for regenerative medicine, is the ability to not elicit any immune response when transplanted. Culture procedures (passages, differentiation, 3D cultures) might compromise the immunomodulatory activity of cells that become no longer suitable for cell therapy approaches. Thus, besides the evaluations that led us to the derivation of insulin-producing cells, we simultaneously tested hAECs for their immunomodulatory properties in vitro. During serum-free validation we showed that FBS deprivation does not alter hAEC ability to inhibit PHA-activated PBMC proliferation in monolayer co-cultures. The same scenario was detected in hAEC spheroids: comparison between non-differentiated hAEC spheroids and differentiated hAEC spheroids (Figure 4) showed the same robust immunomodulation. Hence, hAEC spheroids maintained the immunomodulatory properties observed in monolayer cultures. Collectively, our results suggested that neither the cell transcriptome variations from 2D to 3D culture, nor the secretome modifications 40 upon differentiation altered the immunogenicity of hAECs, which might, therefore, be suitable for transplantation.
In sum, our findings show the first attempt of hAEC pancreatic-endocrine differentiation, performed with the combination of serum-free culture condition, 3D multicellular spheroids creation, and stepwise differentiation protocol. The encouraging outcomes presented here usefully contribute to the implementation of similar procedures for in-depth in vitro and in vivo experiments, and highlight the role of hAECs as a possible cell therapy source for incurable diseases like diabetes.
Footnotes
Acknowledgements
We thank Cinzia Restani and Mariangela Ferrari for their valuable technical assistance.
Author contributions
BO and LI conceived, designed the study, and supervised the experiments. BO, RC, DQ, and FR performed the experiments. BO, FA, RC, and LI interpreted data and performed statistical analysis. BO, FA, and RC wrote the manuscript. PP, LI, MD, DQ, and LB provided materials, reagents, and analysis tools. All authors have seen and approved the final version of the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.