
Abstract
Background: Differences in brain energy metabolism have been found between migraine patients and controls in previous phosphorus magnetic resonance spectroscopy (31P-MRS) studies, most of them emphasizing migraine with aura (MwA). The aim of this study was to verify potential changes in resting-state brain energy metabolism in patients with migraine without aura (MwoA) compared to control subjects by 31P-MRS at 3 tesla.
Methods: Quantification was performed using the phantom replacement technique. MRS measurements were performed interictally and in the medial occipital lobe of 19 MwoA patients and 26 age-matched controls.
Results: A significantly decreased phosphocreatine concentration ([PCr]) was found as in previous studies. While adenosine triphosphate concentration ([ATP]) was considered to be constant in previously published work, this study found a significant decrease in the measured [ATP] in MwoA patients. The inorganic phosphate ([Pi]) and magnesium ([Mg2+]) concentrations were not significantly different between MwoA patients and controls.
Conclusions: The altered metabolic concentrations indicate that the energy metabolism in MwoA patients is impaired, certainly in a subgroup of patients. The actual decrease in [ATP] adds further strength to the theory of the presence of a mitochondrial component in the pathophysiology of migraine.
Introduction
Migraine is a common, disabling, primary headache disorder, with episodic manifestations, affecting women three times more than men (1). Migraine is subdivided into two major subtypes: migraine without aura (MwoA) and migraine with aura (MwA), previously known as common and classic migraine, respectively (2). The aura is characterized by a gradual development of transient and reversible focal neurological symptoms, most often visual, and may be related to cortical spreading depression (3). The headache attack suggests a significant role of the activation of the trigeminovascular system (4). Migraine attacks are often triggered by external factors, with psychological stress, hormones and fasting being the most common (5).
Despite the high prevalence of migraine in the general population, its pathophysiology is still largely unknown. The current assumption is that subcortical structures, probably including the brain stem, hypothalamus and thalamus, are involved in the generation of migraine attacks (6). Even more puzzling are the mechanisms at the basis of the interictal brain disorder that predisposes migraine patients to develop an attack. Until now no integrative model has been formulated that accounts for all the factors that may play a role in migraine neurobiology. Some of these factors include genetic background, nitric oxide hypersensitivity (7), lack of cortical habituation (8,9) and a disturbed energy metabolism. Genetic background and disturbed energy metabolism are discussed below.
Twin studies and familial aggregation studies strongly suggest that migraine is genetically determined (10). The mode of inheritance is most likely multifactorial in both MwA and MwoA (11). No genetic mutations have been found in the common forms of migraine, but a variety of gene polymorphisms, most often irreproducible, have been described (12). Recently though, the first genetic risk factor for migraine (MwA and MwoA) has been described (13). Molecular genetic studies have not detected specific mitochondrial DNA (mtDNA) mutations in patients with migraine, but other studies suggest that particular genetic markers (i.e. neutral polymorphisms or secondary mtDNA mutations) might be present in some migraineurs (10,14). For instance, in migraineurs with occipital stroke (15) as well as in children with MwoA or cyclic vomiting, which can be a migraine equivalent (16), an increased number of mutations was detected in the noncoding control regions of mtDNA. The noncoding region of mtDNA has an extremely high mutation rate and is therefore highly polymorphic. Collections of mtDNA mutations derived from the same ancestor (i.e. haplogroups) can influence oxidative phosphorylation performance and could thus play a more subtle role in migraine pathogenesis, predisposing subjects to the disorder (14). A monogenic subtype of migraine is familial hemiplegic migraine (FHM), a rare form of MwA, in which three known different missense mutations have been found, all causing ionopathies, affecting ion homeostasis and eventually leading to cortical hyperexcitability by increasing synaptic glutamate levels, involved in the generation of an aura (17). No convincing evidence, however, has been obtained that these same genes play a major role in the common forms of migraine (12).
In vivo energy metabolism can be studied by phosphorus magnetic resonance spectroscopy (31P-MRS). This technique allows for the noninvasive quantification of phosphorylated compounds, including high-energy phosphates such as adenosine triphosphate (ATP) and phosphocreatine (PCr) and low-energy phosphates such as inorganic phosphate (Pi). 31P-MRS can also determine intracellular pH (pHi) and intracellular magnesium (Mg2+). In addition, adenosine diphosphate (ADP) and the phosphorylation potential (PP) can be calculated based on the creatine kinase equilibrium. Figure 1 shows resting-state brain energy metabolism and the corresponding metabolites that can be detected by 31P-MRS (18). In the past 20 years, several 31P-MRS studies suggested an abnormal cerebral energy metabolism in migraine patients during ictal and interictal periods (see Table 1 for a review) (19–29). These alterations concern energy metabolism and are not limited to the brain but have also been observed in muscle (22–24,27,30). The reduced energy potential was interpreted as being indicative of a reduced mitochondrial reserve and was hypothesized to be the biochemical substrate of the susceptibility to migraine attacks (28,31). As summarized in Table 1, studies have been performed in a wide variety of migraine subtypes, during either the ictal or interictal period, mostly localized in the occipital lobe and mostly in patients who did not undergo prophylactic treatment. In several of these studies, the migraine patient group was heterogeneous and information about the attack frequency was scarce. These studies were performed at different field strengths and provided little information, if any, about the procedure for quantification of the metabolites. Most importantly, the ATP concentration ([ATP]) was always assumed constant and equal to that of normal controls, being 3 mM (32), or was not mentioned at all. However, cortical ATP levels can be decreased, as was demonstrated with 31P-MRS in other pathologies, such as systemic lupus erythematosus (33), episodic ataxia type 2 (34), progressive supranuclear palsy (35) and Parkinson’s disease (36).
Resting-state brain energy metabolism, adapted from Kemp et al., 2000 (18). In the resting state, intracellular adenosine triphosphate (ATP) results from the balance between ATP use and ATP synthesis. ATP is derived almost exclusively from mitochondrial oxidative phosphorylation, depending on glucose and oxygen supply. Changes in phosphocreatine (PCr) reflect the time integral of the mismatch between ATP usage and supply, as adenosine diphosphate (ADP) can be rephosphorylated through the creatine kinase reaction, with conversion from PCr to creatine (Cr). Magnesium (Mg2+) is bound to ATP in order for ATP to be biologically active. The metabolites in boxes can be detected by 31P-MRS. Literature survey of 31P-MRS studies performed in the brain of migraine patients MwpA = migraine with prolonged aura. MS = migrainous stroke. N = number of included patients. PCr = phosphocreatine. Pi = inorganic phosphate. PP = phosphorylation potential. ADP = adenosine diphosphate. ATP = adenosine triphosphate. Mg2+ = magnesium. PDE = phosphodiesters.[ ]are molar concentrations. PCr/Pi and Pi/ATP are ratios. aOther = MS or MwpA, b— = information not mentioned in the study.

In this study, the aim was to revisit quantitative 31P-MRS at 3 tesla in the medial occipital lobe in patients with migraine without aura in the interictal phase, using quantification based on the phantom replacement technique.
Materials and methods
Patients and control subjects
Participant characteristics

SD = standard deviation.
The study was approved by the local ethics committee and all subjects gave written informed consent. The migraine patients were diagnosed with MwoA according to the criteria of the International Headache Society (2). Patients experienced two to eight attacks per month, were not using any prophylactic medication and were attack-free for at least 48 hours. None of the 19 patients experienced a migraine attack within 24 hours after the spectroscopy study.
31P-MRS and region of interest
All measurements were performed on a 3 tesla Siemens TrioTim whole-body scanner (Erlangen, Germany), using a 26.5-cm-diameter quadrature dual tuned (31P-1H) transmit/receive birdcage head coil (Rapid Biomedical, Würzburg-Rimpar, Germany). Spectra were acquired using a two-dimensional chemical shift imaging (CSI) phase-encoding scheme applying a pulse-free induction decay sequence. Manual shimming of the B0 magnetic field and manual optimization of the transmitter pulse power were used.
The field of view (FOV) was placed occipitally, covering the visual cortex (Figures 2a and 2b), localized on T1-weighted gradient-echo images in three orthogonal planes with a slice thickness of 1 mm, a repetition time (TR) of 1550 ms and an echo time (TE) of 2.37 ms.
(a) Axial T1-weighted image and (b) sagittal T1-weighted image with the field of view and the voxel in the medial occipital lobe. (c) Spectrum acquired in the highlighted voxel. (d) The same spectrum with curve fitting. A CSI (chemical shift imaging) slice of (240 × 240 × 30) mm3 was placed and the nominal voxel volume was 30 ml.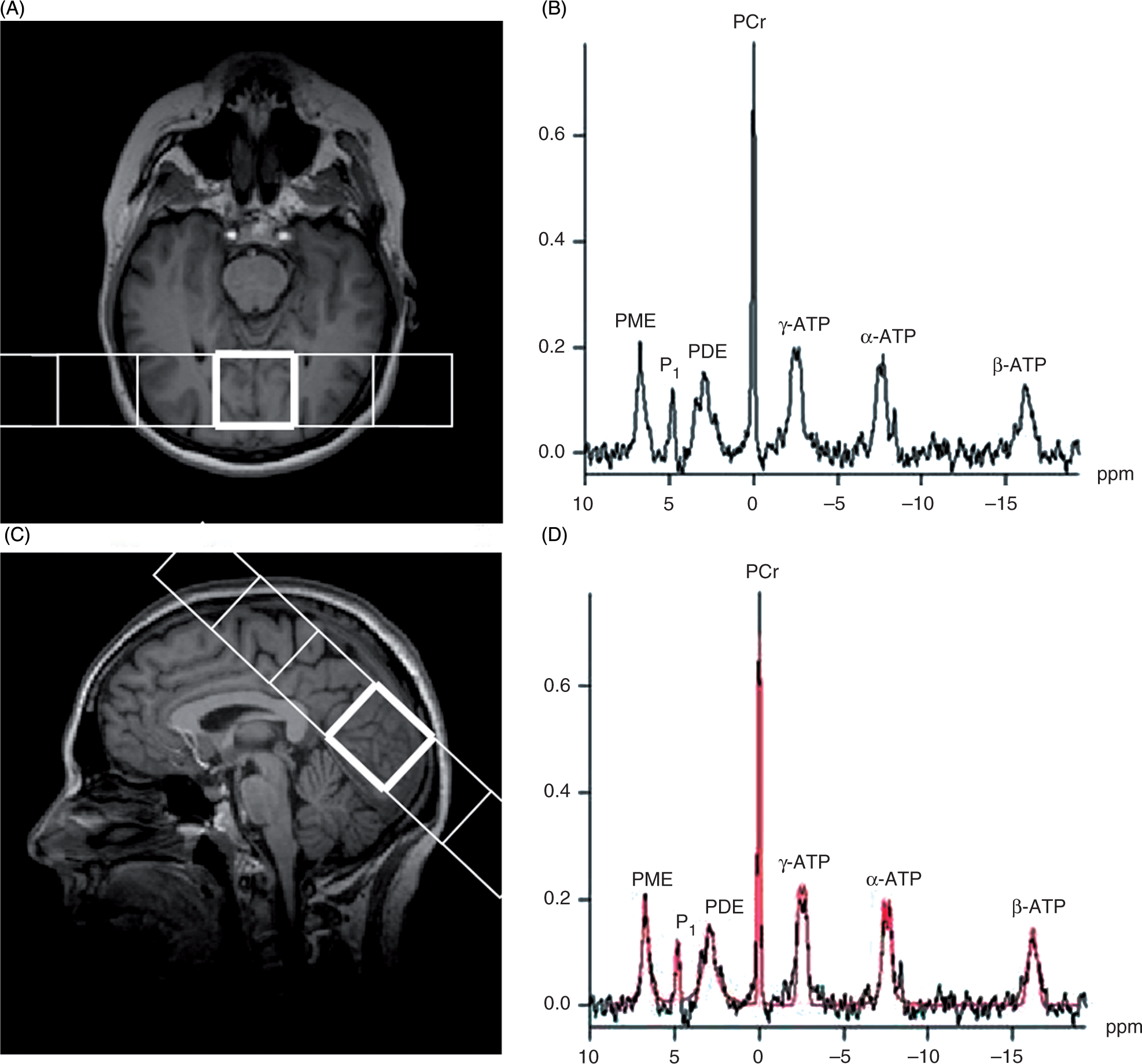
A 240 ×240 × 30 mm3 two-dimensional CSI slice was recorded. Phase encoding was used with a weighted acquisition scheme, resulting in an axial slice with a nominal thickness of 30 mm and 30 × 30 mm2 in plane resolution (four averages, flip angle of 90°, TR of 4000 ms and TE of 2.3 ms). The raw data of each acquisition consisted of 1024 complex-valued data points, at a sampling period of 0.4 ms. The corresponding bandwidth was 2500 Hz. The total duration of the measurement was approximately 10 minutes.
External calibration
Signal intensities were quantified in terms of arbitrary units (a.u.) by using the phantom replacement technique. The reference phantom contained an aqueous solution (pH 7) of 10 mM PCr (Sigma Aldrich). Sodium chloride (NaCl) and sodium azide (NaN3) were added to change the conductivity and to avoid mycotic growth, respectively. The phantom was made of plastic, was spherical and had a diameter of 10.4 cm.
The complete equation for calculating the in vivo concentrations is given by:
Spectral analysis
Figure 2c shows a typical 31P-spectrum. The 31P resonances can be allocated to ATP, PCr, phosphodiesters (PDE), phosphomonoesters (PME) and Pi. The 31P-MRS spectra for ATP contain three signals corresponding to the three phosphorus nuclei of the molecule: the α-ATP resonance contains contributions from both NADH and α-ADP, the γ-ATP resonance contains contributions from β-ADP, and β-ATP is proportional to the total cellular ATP content. We therefore used the β-ATP resonance to quantify [ATP] (37). Following apodization (exponential filter, width = 110 ms) and zero-filling, the Fourier transformed free induction decays were phase and baseline corrected. Peak areas were obtained by the classical Levenberg-Marquardt frequency domain-fitting method, using the software on the scanner (syngoMR B15, Numaris 4 [Siemens]) (Figure 2d).
Creatine kinase reaction
pHi was calculated from the chemical shift of Pi in relation to PCr (38). Brain cytosolic free [Mg2+] was assessed by a semi-empirical equation that correlates the chemical shift of the β-ATP signal from PCr to the free [Mg2+] (39).
The biochemical equation for the creatine kinase reaction is as follows:
Based on this equilibrium, the cytosolic ADP concentration ([ADP]) was calculated as follows:

Statistical analysis
Statistical analysis was performed using the SPSS software (SPSS 15.0 for Windows; Chicago, IL, USA). Descriptive statistics was applied for age, Vtra, metabolite concentrations and calculated values. Wilcoxon’s rank-sum test was applied to compare Vtra, metabolite concentrations and calculated values between MwoA patients and controls. Results were considered to be significant at p < .05.
Results
Concentration values in a.u. and calculated values (mean ± SD)

a.u. = arbitrary units. SD = standard deviation. *Level of significance, p < .05.
First, no significant differences were observed for any of the variables between males and females in the control group (data not shown).
A significantly low PCr content was found in MwoA patients (p = .001). We also fitted the β-ATP signal to calculate [ATP]. On average, the ATP content was found to be significantly lower in MwoA patients compared to controls (p = .023). This corresponds with an average reduction of [ATP] by approximately 15%. A subgroup of the MwoA patients (i.e. 20 %) demonstrates [ATP] values at least 2 standard deviations (SD) beneath the average [ATP] of controls, as illustrated in Figure 3. This subgroup corresponds with those patients who had the highest attack frequency.
Distribution of the concentrations of the high-energy phosphates adenosine triphosphate (ATP) and phosphocreatine (PCr). Mean concentrations are illustrated with a horizontal line. The mean minus 2x the standard deviation is also shown. Note that in every plot, four patients have concentrations beneath this threshold. These patients are not the same for the two plots. In the case of [ATP], these values correspond with the patients who had the highest attack frequency.
Mean [Pi] showed no significant difference between the MwoA patient group and the controls (p = .129). Calculated values of pHi and [ADP] showed no differences between MwoA patients and controls (p = .702 and p = .735, respectively). The PP was significantly decreased in the MwoA patient group (p = 0.001). Finally, the Mg2+ content did not demonstrate a significant difference between MwoA patients and controls (p = .254).
Discussion
31P-MRS provides a reliable noninvasive tool for the in vivo assessment of mitochondrial functionality by measuring cytosolic [ATP], [PCr], and [Pi] and by calculating [ADP], PP, pH and [Mg2+], all playing crucial roles in the creatine kinase equilibrium (Figure 1).
In migraine, several magnetic resonance spectroscopy studies, in particular 31P-MRS, have been performed. These studies were the first to document intrinsic biochemical abnormalities in migraine. In most of these studies, data was obtained in patients with migraine with prolonged aura (MwpA) and MwA patients, including patients with FHM (Table 1). Only two studies emphasized MwoA; in these, measurements were performed interictally (24,28).
To assess the resting-state brain energy metabolism, we performed 31P-MRS on a 3 tesla high-field scanner in the medial occipital lobe of MwoA patients, who were attack-free and were not using any prophylactic medication, and compared the results with those of previous 31P-MRS studies. It is worth underlining that the aim was to study a very homogeneous group of migraine patients who experienced a well-defined number of attacks (two to eight per month). This is in contrast to several other studies, in which a heterogeneous group of migraine patients was examined and in which information about the attack frequency is not always available (21,25). By focusing on MwoA, the potential influence of the predisposition to aura, which is most often visual and thus related to the occipital cortex too, was avoided in this study. The chance of migraine attack–related brain disturbances was minimized by examining the patients interictally, by assuring patients were at pain-free for at least 48 hours before the procedure and pain-free for at least 24 hours after the procedure.
We calibrated the in vivo spectra to an external standard to quantify metabolite concentrations rather than using raw signal intensities.
Because we were looking for metabolic disturbances, measurements were performed in the occipital lobe, as it has been found that the regional cerebral metabolic oxygen rate (CMRO2) is significantly higher in this brain area compared to other cortical regions (42). Additionally, the regional cerebral metabolic glucose rate (CMRgl) has been found to be the highest in occipital white matter and the visual cortex (43). An additional advantage is that the visual cortex remains metabolically unchanged with advancing age (44).
High-energy phosphate metabolism was found to be altered in MwoA patients in the medial occipital lobe. The average PCr concentrations were decreased significantly, which is comparable with previous data (24). However, others could not confirm this (28). In contrast to all other 31P-MRS studies in migraine, in this study [ATP] was calculated from the spectrum. All other 31P-MRS studies in migraine assumed a constant [ATP] of 3 mM (22–28). This assumption was based on a 31P-MRS study in healthy subjects (32). We observed a significant decrease in the average [ATP] of approximately 15% in MwoA patients as compared to controls.
The PP is an index of mitochondrial functionality and of the energy status of the cell. The higher the PP, the more free energy is available in the cell. The PP was significantly decreased in MwoA patients compared to controls. This is in concordance with the findings of previous studies (23,24). ADP is the major driving force for mitochondrial energy production. The concentration of free cytosolic ADP is in the micromolar range and is below the sensitivity of MRS in vivo. However, [ADP] can be calculated from the creatine kinase equilibrium. In contrast to most other studies, we did not find a difference between MwoA patients and controls. In all other studies, [ADP] was derived assuming an ATP concentration of 3 mM in both MwoA patients and controls. When calculating [ADP] assuming a constant [ATP] in our subjects, there was still no significant difference in [ADP] between MwoA patients and controls.
No cytosolic pH difference was found between MwoA patients and controls, as was also the case in previous studies (24,28).
Mg2+ is an important enzymatic cofactor and can influence the equilibrium constant of several biochemical reactions, including the creatine kinase reaction. Brain cytosolic free [Mg2+] was not significantly different in MwoA patients compared to controls. In a previous study, a significant decrease in interictal [Mg2+] was found in MwA patients (27). In another study, whereas a significantly reduced brain [Mg2+] was observed ictally in both MwA and MwoA patients, this was not the case for interictal brain [Mg2+] in some of the patients (45). The absence of a significant interictal reduction of [Mg2+] may be attributed to the large heterogeneity of the patient group, the variability of the examined brain area examined, or the semi-empirical method to calculate [Mg2+].
The decrease in high-energy phosphates suggests a mitochondrial component in the neurobiology of migraine. The brain is one of the most energy-expensive tissues and although it comprises only 2–3 % of the total body weight (46), it utilizes approximately 25% of the total glucose. The baseline metabolic rate of the brain is very high and most energy is thought to support glutamatergic neurotransmission, at least in the cortical grey matter (47). The brain at rest relies almost entirely on aerobic metabolism with glucose as the principal fuel (48). Glycogen can also be used but is only found in small amounts in the astrocytes (49). Lactate, an indicator of anaerobic glycolysis, accumulating in case of mitochondrial dysfunction, can be detected by 1H-MRS (50). A recent resting state 1H-MRS study did not show any quantifiable lactate in the visual cortex of MwoA patients (41). The observed decrease in [ATP], often called the ‘molecular unit of currency’ of intracellular energy transfer, might thus be explained by a decrease in ATP production through aerobic glycolysis and oxidative phosphorylation (Figure 1). We emphasize that the decrease of high-energy phosphates in MwoA was detected at rest (interictally), implying the constant nature of this energy disturbance rather than being a transient phenomenon. This cannot be explained by hypermetabolism, as a recent [18]fluorodeoxyglucose–positron emission tomography (PET) study in migraine patients failed to show any hypermetabolic brain regions, including in the occipital lobe (51).
Is the reduction of interictal [ATP] related to a decrease in the number of mitochondria or to a decrease in mitochondrial efficiency? It is very difficult to draw conclusions in this regard since the brain is not readily accessible for histological and biochemical studies. There is, however, additional evidence for a mitochondrial component in migraine pathophysiology.
With 31P-MRS, a decreased postexercise recovery of high-energy phosphates was found in the gastrocnemius muscle of MwoA patients compared to controls (24,52). Platelet mitochondrial enzyme activities were found significantly lower in MwoA patients than in controls (53). Plasma lactate and pyruvate levels were found significantly increased in migraine patients compared to controls (54). Alterations comparable to those in migraine have been found in mitochondrial encephalomyopathies, and could conceivably result either from errors in energetic, oxidative pathways limiting the energy supply of cells, or to defects in ionic conductances or some specific neurotransmitters, responsible for neuronal excitability, whose failure increases energy expenditure in excitable cells (55). Stroke-like episodes and migraine are the predominant symptoms of mitochondrial encephalomyopathy with lactic acidosis and stroke-like episodes (MELAS), but the mtDNA point mutations at bp 3243 and 3271, generally associated with this syndrome, were not found in migraine (56). 31P-MRS studies in mitochondrial diseases such as MELAS show similar results as in migraine both in brain (57) and in muscle (58). In both aforementioned studies, no data concerning [ATP] is shown or [ATP] is, surprisingly, assumed constant, respectively. An occasional mtDNA mutation has been found in one study (59); however, this was not the case in systematic studies (60). Additionally, another study showed mitochondrial abnormalities in muscle biopsies of some migraine patients compared to controls (30). Mitochondrial metabolic enhancers such as riboflavin (61) and coenzyme Q10 (62) have a prophylactic effect in a subgroup of migraine patients. Response to riboflavin seems to be related to a specific mitochondrial haplotype (14).
Stress, female hormones and fasting are the most common trigger factors for migraine attacks (5). Normobaric hypoxia is able to trigger a migraine attack (63), as well as hypoglycemia (5) in a subgroup of patients. It is intuitive to think that such specific triggers, having a direct effect on oxygen and glucose metabolism, respectively, would put a significant strain on the mitochondria. The hormones progesterone and estrogen regulate oxidative metabolism in brain mitochondria (64). It has also been shown, albeit in cardiac myocytes, that noradrenaline, a typical stress mediator, causes calcium overload and results in a decreased mitochondrial respiration (65). Stress activates the noradrenergic locus coeruleus (66), which projects widely to the cortex, including the occipital visual cortex (67).
A mitochondrial defect may reduce the threshold for migraine. The hypothesis of migraine being a “biobehavioural” (68) or a threshold disorder (69) states that it is a disorder in which an intrinsic metabolic defect renders the brain more susceptible to various factors that trigger an attack. It is hypothesized that trigger factors would act by increasing the metabolic energy demand or decreasing the metabolic energy supply. When a certain metabolic threshold is reached in the brain, which is already near to its maximum energetic capabilities, a metabolic crisis could be induced which is responsible for the headache attack. Our data show that [ATP] is profoundly reduced in a subgroup of MwoA patients. Rather than being a generic component of migraine neurobiology, we hypothesize that a reduced mitochondrial energy reserve may be one of the many factors determining the migraine threshold. This hypothesis is in line with the observation that a subgroup of migraine patients respond to mitochondrial enhancers, such as riboflavin (61) and coenzyme Q10 (62), and that this response may be related to a specific mitochondrial haplotype (14).
In conclusion, a significant depletion of high-energy phosphates, both ATP and PCr, was found at rest in the medial occipital lobe of MwoA patients, compared to controls. This suggests a decrease in oxidative phosphorylation and implies a mitochondrial component in the pathophysiology of migraine without aura. A decreased mitochondrial energy reserve is pivotal in lowering the threshold for a migraine attack, at least in a subgroup of patients. It has to be emphasized that also other factors such as a lack of cortical habituation, NO hypersensitivity and genetic aspects play crucial roles in migraine pathophysiology. 31P-MRS is sensitive enough to reveal defects of cell energy production of MwoA patients even in the absence of any symptoms and signs.
Footnotes
Acknowledgements
This research is funded by the Special Research Fund PhD grant B/07768/02 and performed at the Ghent Institute for Functional and Metabolic Imaging. The Ghent University Department of Radiation Oncology and Experimental Cancer Research is also greatly acknowledged for the use of the radiophysics laboratory