
Abstract
Circular depressions are concave, shallow depressions found on planar landscape surfaces in the southern Namib Desert. They occur on gravelly substrates with nearly level to very slightly inclined surfaces. The depressions range from 6 to 10 m in diameter with centers typically depressed 10–20 cm below the level of the surrounding terrain. Locations of individual circular depressions were mapped at one site using ground-based measurements and at three additional sites using Google Earth imagery. At all sites, circular depressions are highly overdispersed with densities ranging from approximately 10–20/ha and corresponding nearest neighbor distances of 17–24 m. Large fragments of weathered calcrete and stones occur on soil surfaces surrounding circular depressions, but not within the depressions. Circular depressions at one site contained active burrow systems of Brants’ whistling rat (Paratomys brantsii). Bioturbation by these rodents contributes to the non-cohesive nature of the sandy substrate, which promotes aeolian deflation and formation of the depressions. Excavations of the burrow systems by the honey badger (Mellivora capensis) in search of rodent prey can transfer large stones and calcrete fragments from the subsurface to the surface and subsequently move those materials about the surface. Even if such sequential, horizontal displacements are in random directions, such movements can eventually yield a central, clast-free area surrounded by a peripheral zone where the clasts accumulate once they have been displaced beyond the margin of the area to which the predator is drawn in search of rodent prey. A conceptual model consisting of a two-dimensional random walk of large clasts about the surface until they are displaced from the focal “arena” of rodent occupation provides a novel explanation for origin of a spatially organized pattern that is initiated through the random displacement of those materials. Comparable microtopographic patterning associated with bioturbation in other arid and semi-arid environments may have similar origins.
Introduction
Within the last few decades, biogeomorphology has developed into a major area of study focused on how organisms interact with and alter the physical environment (Viles, 1988, 2020; Coombes, 2016). Two important conceptual/theoretical issues directing current biogeomorphic research are (1) how do ecological and geomorphic systems interact and (2) how important are such interactions in shaping the landscape (Viles, 2020). In terrestrial environments, such interaction and their consequences are diverse, and include modification of topographic relief and altered soil conditions. In terrestrial environments, many kinds of animals, through their earth-moving activities, are agents of geomorphic change (Butler, 1995). Major biogenic processes due to excavation activities by animals include bioturbation, which has a variety of cascading consequences, as well as bioconstruction, where the excavation activities directly create distinct geomorphic features (Corenblit et al., 2011).
Displacement of excavated soil by animals can contribute to formation of both topographic convexities (mounds) and concavities (depressions). The Mima mounds of the western United States are one particularly well-studied example, with strong evidence that soil movement by burrowing pocket gophers is a major contributor to their formation (Burnham and Johnson, 2012). Topographic depressions have received considerably less research attention, but include wallows and pits created by large ungulates, elephants, large marine pinnipeds, and other animals (summarized in Butler, 1995). In addition to microtopographic alteration, altered soil conditions, including permeability and soil nutrient content, can have substantial ecological effects (Holtmeier, 2015). In arid and semi-arid environments, even small alterations of local relief and soil texture can greatly influence surface hydrology, thereby contributing spatial variability in soil moisture availability, and ultimately, vegetation responses and ecosystem function (Eldridge and James, 2009). Bioturbation by burrowing animals can also alter soil characteristics, including the susceptibility to erosion, either by water or wind (Jouquet et al., 2021; Yair, 1995).
Louw and Seely (1982, p. 143, Fig. 9.2) briefly mentioned and photographically illustrated widely separated, circular patches of vegetation in the central Namib Desert, attributing the patches to creation of more favorable soil conditions by burrowing activities of gerbils and ground squirrels. Cox (1987) investigated the same kind of circular patches of ephemeral grasses in the central Namib Desert, showing that the patches are regularly spaced, and averaging less than 6 m in diameter. The patches contained sandy soil, and were frequently surrounded by a ring of coarser gravels. Half of all the patches were occupied by heaps of soil materials displaced by gerbil burrow systems, and some of the patches contained a depression 15 cm or less in depth. He hypothesized that creation of burrow systems by the rodents displaced soil materials outwards towards the periphery of occupied areas. Subsequent removal of fine soil materials from those accumulations by the wind was suggested as the process that concentrated a lag of coarser gravels on the surface surrounding the patch. Additionally, accumulation of aeolian sand within the collapsed central areas of the zones originally containing rodent burrow systems contributed to deeper, sandy soils. The modified soil conditions within the patch contributed to greater infiltration and moisture storage capacity, and hence, greater vegetation productivity. Shaanika (2020) also investigated these kinds of vegetation patches associated with burrowing of Gerbillurus setzeri in the central Namib Desert. Those patches, approximately 5 m diameter and spaced 10–15 m apart, exhibited significantly greater vegetation productivity, attributed to modifications of soil conditions by the burrowing rodents.
In this paper, we investigate the distribution and factors involved in formation of regularly spaced, microtopographic features consisting of circular-shaped, slight depressions that occur widely in the southern portion of the Namib Desert. In some places, the features entirely lack vegetation, and consequently, these structures are better described on the basis of their physical morphology and are referred to as circular depressions. Although these structures possess some of the same characteristics of the circular vegetation patches described above, they differ in other features. In particular, the depressions are often surrounded by a peripheral concentration of cobble-sized clasts on the surface. The clasts are typically much coarser than the gravelly materials that surround vegetation circles described by Cox (1987) and are unlikely to have been displaced over a scale of several meters simply through the burrowing activity of small rodents. We provide evidence for the role of excavations made by larger mammals in search of rodent prey in moving these coarser materials. Additionally, we present a conceptual model of how the repeated, sequential displacement of such materials, even in random directions, eventually can generate central areas within circular depressions that are free of large clasts, with those materials concentrated about the margins of the structure. Comparable geomorphic features described from other arid regions of the world may have similar origins. The observations and conceptual model presented in this paper provide a new perspective regarding processes that can contribute to the origin of spatially ordered distribution of surface materials through the excavating activities of animals.
Study area and methods
Study area and access requirements
The study took place in the Tsau ǁKhaeb (Sperrgebiet) National Park, which extends along the entire Atlantic coast from approximately 70 km north of Lüderitz, south to the Orange River and border with South Africa (Figure 1). Soon after the local discovery of diamonds, the area was designated as the Sperrgebiet (prohibited zone) by the German government in 1908 in order to control access and mining operations. Since then, the area has been largely inaccessible to the public. As a consequence, this exclusion has greatly restricted research and understanding of many environmental features and processes of the region (Burke, 2001). The national park was designated in 2008, yet entry to the area remains restricted and tightly controlled due to continued diamond mining operations. Access and permission to conduct our field research, including collection of soil samples, required permits from the Namibian Ministry of Mines and Energy, the Ministry of Environment and Tourism, as well as entry and exit permission and inspections by officials of Namdeb, the diamond mining corporation owned jointly by the Namibian government and private enterprise. Location of study in Tsau ǁKhaeb (Sperrgebiet) National Park, Namibia. Dotted lines indicate routes of travel during fieldwork of 2021 and 2022. Numbered triangles are locations where observations of circular depressions were made; numbers correspond with site numbers listed in Tables 1 and 3 and discussed in text.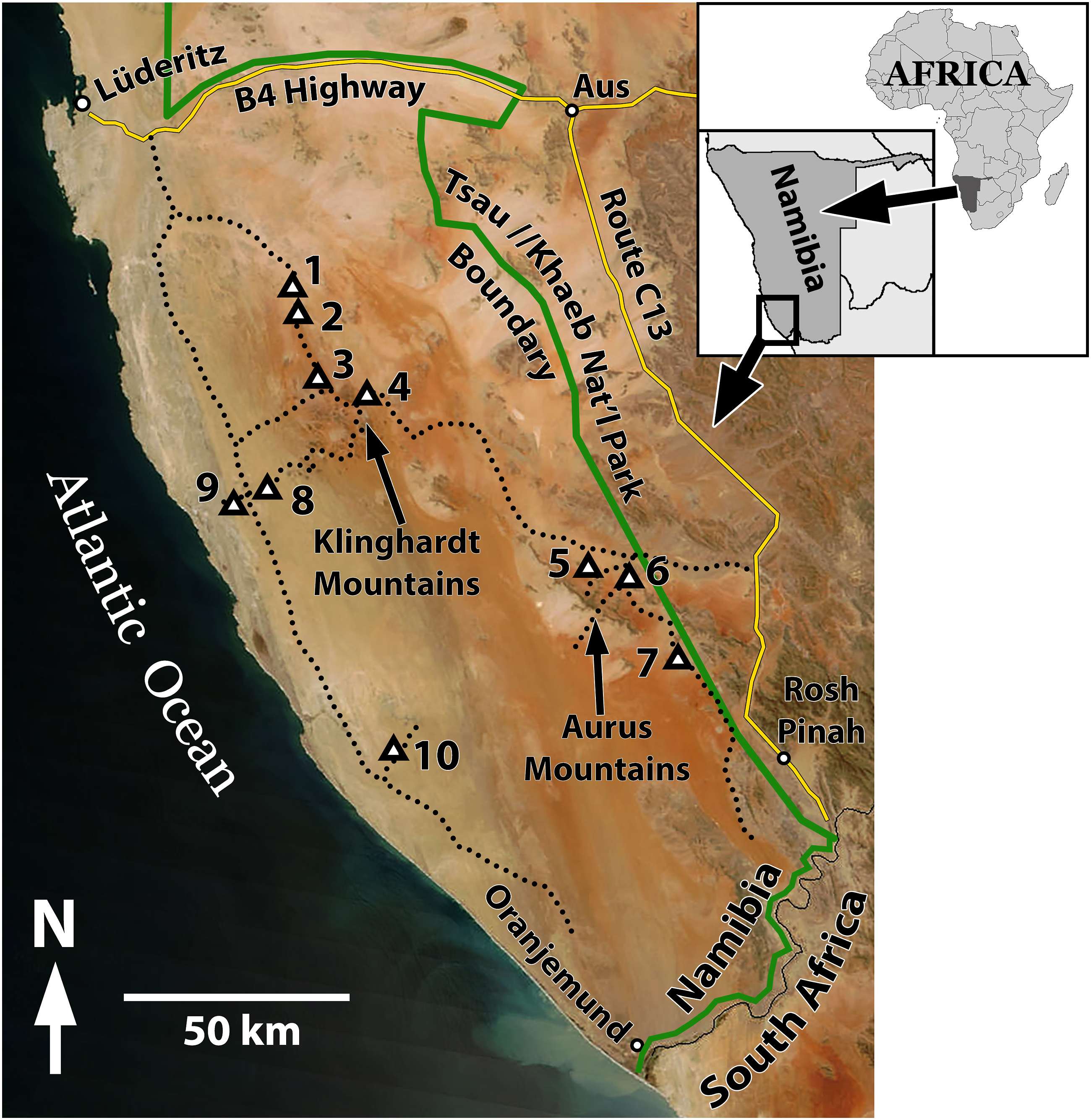
Climate and vegetation
Annual precipitation at Lüderitz on the Atlantic coast averages 16.7 mm (Muller et al., 2008). Annual precipitation increases inland to approximately 50 mm, and there is a strong southwest to northeast gradient of decreasing amounts of winter precipitation (Burke, 2001). Orographic uplift by isolated, low mountains locally increases precipitation, as indicated by increased vegetation density in those elevated areas (Burke, 2006). Coastal fog provides significant moisture in the coastal area. Strong, nearly perpetual winds throughout much of the year characterize the entire area (Burke, 2001; Corbett, 1993; Lancaster, 1985).
The area represents the northern extension of the Succulent Karoo Biome (Jürgens, 1991), and a major component of the vegetation consists of small, succulent-leaved shrubs in the family Aizoaceae (Mesembryanthemaceae), which are dominant in the southwestern part of the study area, the coastal region, and in inselbergs and mountains. Ephemeral grassland becomes more prevalent towards the northeast portion of the national park. Vegetation composition and cover also varies substantially as a function of soil characteristics and orographic effects on precipitation (Burke, 2006). Due to the long history of restricted access and use, this is the only large, contiguous area of the Succulent Karoo Biome that has never been subjected to grazing by domestic livestock, except for some localized portions in the eastern-most part of the park adjoining private farmland.
Methods
Initial field reconnaissance was conducted during 3 days from 17 to 19 October 2021, and additional, more detailed fieldwork was conducted over a 12-day period 23 September–3 October 2022, in areas of the coastal plain and further inland around low mountain ranges (Klinghardt and Aurus mountains) (Figure 1). Locations of detailed observations and photographs were recorded on a hand-held GPS unit, enabling subsequent location of observed features using Google Earth imagery. Slope inclinations of areas containing circular depressions were estimated in Google Earth, along linear paths through areas of ground observation and using the Path measurement tool, which provided path length and elevations of start and end points. Dominant plant species present within circular depressions as well as those present in the surrounding area were recorded. One of the sites (Site 7) was within an area for which high-resolution aerial imagery was present in eight separate years over a 21-year period from 2003 to 2023. This time series was used to examine short-term changes in vegetation cover during that period within circular depressions in relation to a documented, major precipitation event.
Nearest neighbor analyses of circular depressions.
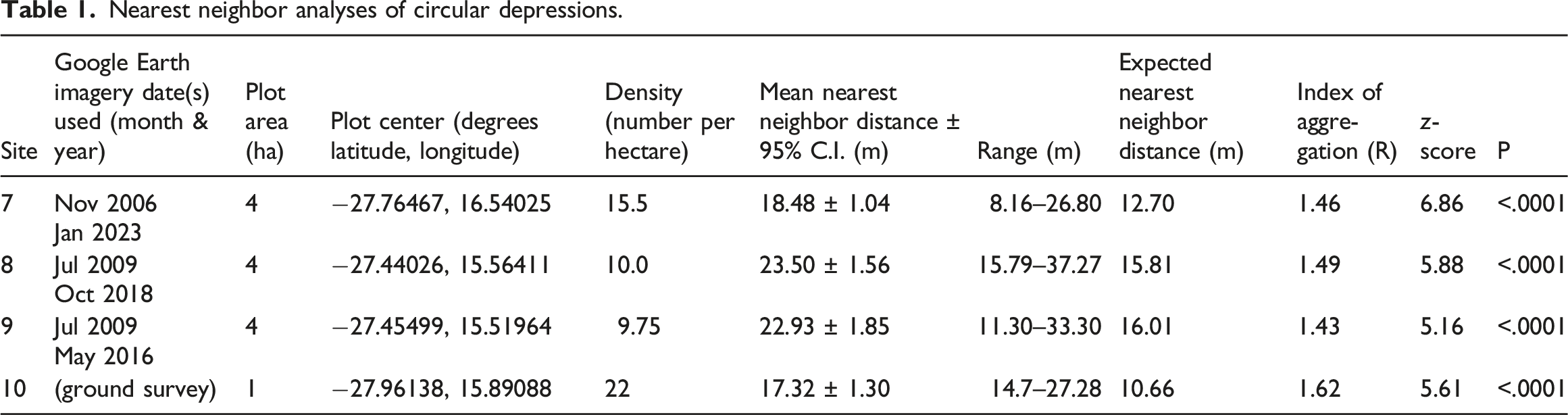
For each of the four sites described above (Sites 7–10), where locations of circular depressions were mapped in Google Earth, the distance between the center of each circular depression and the center of its nearest neighboring circular depression was measured with the Google Earth Line measuring tool and recorded to the nearest 0.1 m. In cases where the nearest neighbor circular depression was located outside of the plot boundary, that measurement was taken to permit an unbiased estimate of mean nearest neighbor distance (Krebs, 1989). For each of the four sites, mean expected nearest neighbor distance, the index of spatial aggregation (R), and associated z statistic for testing of statistical significance were computed following Clark and Evans (1954) as presented in Krebs (1989). Values of R are computed as the ratio of the mean of measured mean neighbor distances and mean expected nearest neighbor distance. For a randomly dispersed set of objects, R = 1; R with statistically significant values less than 1 represent spatial aggregation (clumping), and those with statistically significant values greater than 1 represent overdispersion (tending toward regular spacing).
At Site 10, where locations of circular depressions were mapped on the ground, diameters of a representative subset of seven depressions were measured to the nearest 0.1 m with a graduated measuring scale. Depths of five depressions were measured to the nearest centimeter by holding a vertical scale at the center of the depression and moving the vertical height of a horizontal sighting line made with a hand-held level (Suunto clinometer) to the height on the vertical scale level with the peripheral rim of the depression. The number of rodent burrow entry holes per depressions was recorded from five depressions, and within each of those depressions, the diameters of five entrance holes were measured to the nearest millimeter for a total of 25 measurements.
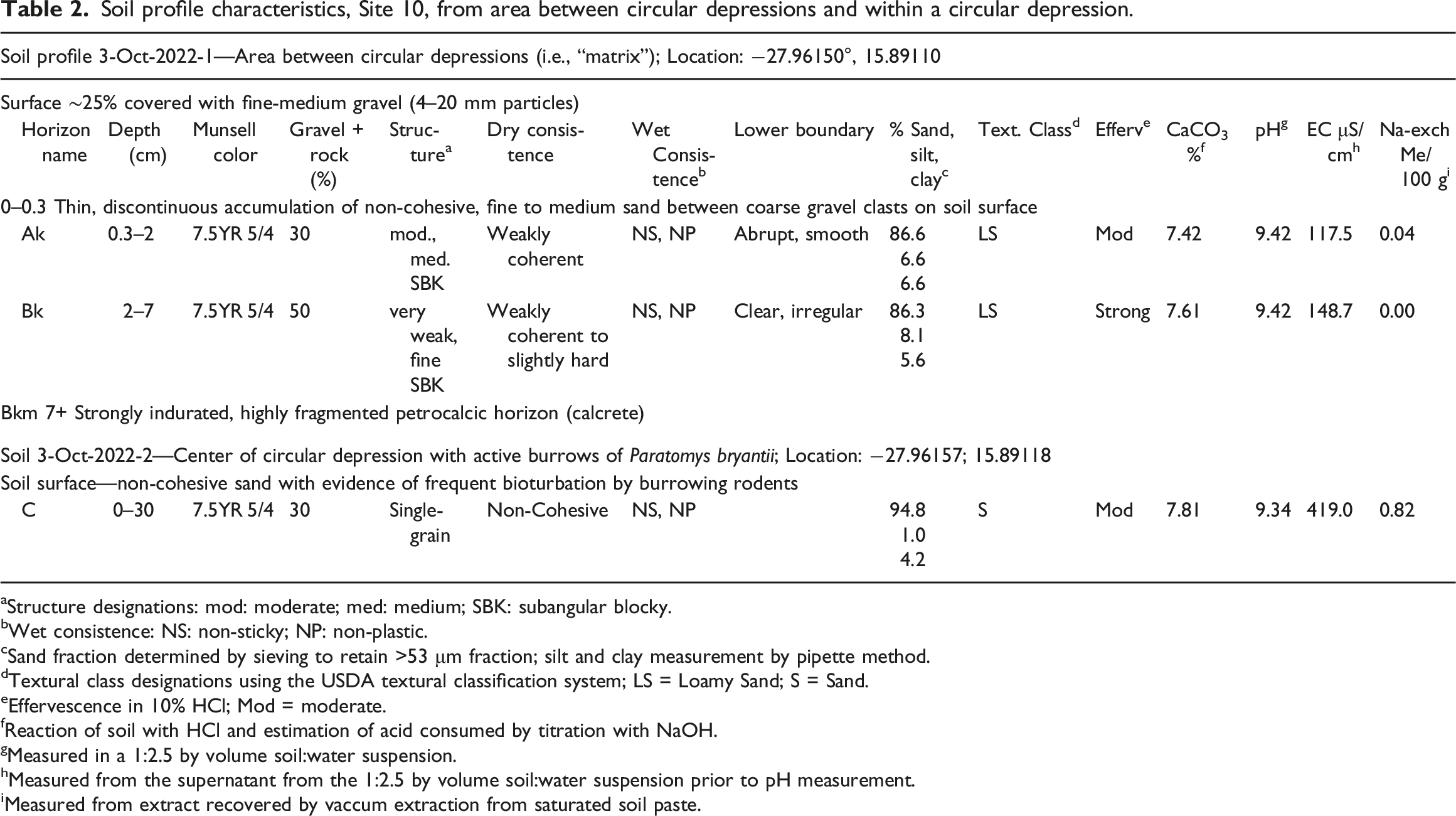
Soil profile characteristics, Site 10, from area between circular depressions and within a circular depression.
aStructure designations: mod: moderate; med: medium; SBK: subangular blocky.
bWet consistence: NS: non-sticky; NP: non-plastic.
cSand fraction determined by sieving to retain >53 μm fraction; silt and clay measurement by pipette method.
dTextural class designations using the USDA textural classification system; LS = Loamy Sand; S = Sand.
eEffervescence in 10% HCl; Mod = moderate.
fReaction of soil with HCl and estimation of acid consumed by titration with NaOH.
gMeasured in a 1:2.5 by volume soil:water suspension.
hMeasured from the supernatant from the 1:2.5 by volume soil:water suspension prior to pH measurement.
iMeasured from extract recovered by vaccum extraction from saturated soil paste.
Results
Characteristics and occurrence
Circular depressions are shallow, concave depressions on planar landscape surfaces. The depressions typically range in diameter from 6 to 10 m, with centers 10–20 cm below the level of the surrounding terrain (Figure 2(a)). They occur widely throughout Tsau//Khaeb (Sperrgebiet) National Park from the Atlantic coastal plain (Sites 8–10, Figure 1), and inland to valley floors, pediments, and basins adjacent to low mountains (Sites 1–7, Figure 1). The features occur exclusively on nearly level to very slightly sloping surfaces with slope inclinations typically 1.5% or less (Table 3). Landscapes dominated by active aeolian sand dunes and sand sheets lack the features. (a) Circular depression ∼9 m wide surrounded by calcrete clasts and occasional basalt and quartz cobbles. Center of depression ∼20 cm lower than surrounding area. Plants within depression are mostly dead remains of Brownanthus sp. (Site #3, 25 Sept 2022). (b) Soil surface within above circular depression; centimeter scale divisions. Largest particles are gravel-sized clasts of quartz, <1 cm diameter. (c) Soil surface outside of depression with large clasts of petrocalcic rubble exceeding 10 cm diameter. The dark cobble in upper left is basalt; small pieces of quartz are in upper right quadrant. (d) Surface of strongly indurated calcrete clast deeply pitted by dissolution weathering. Many of the small, protruding surfaces on the clasts are surfaces of quartz gravels incorporated into the calcrete matrix; their relative insolubility leaves them protruding as the surrounding carbonate-rich matrix dissolves. Locations of circular depressions examined in the field with elevations and slope inclinations of the sites.
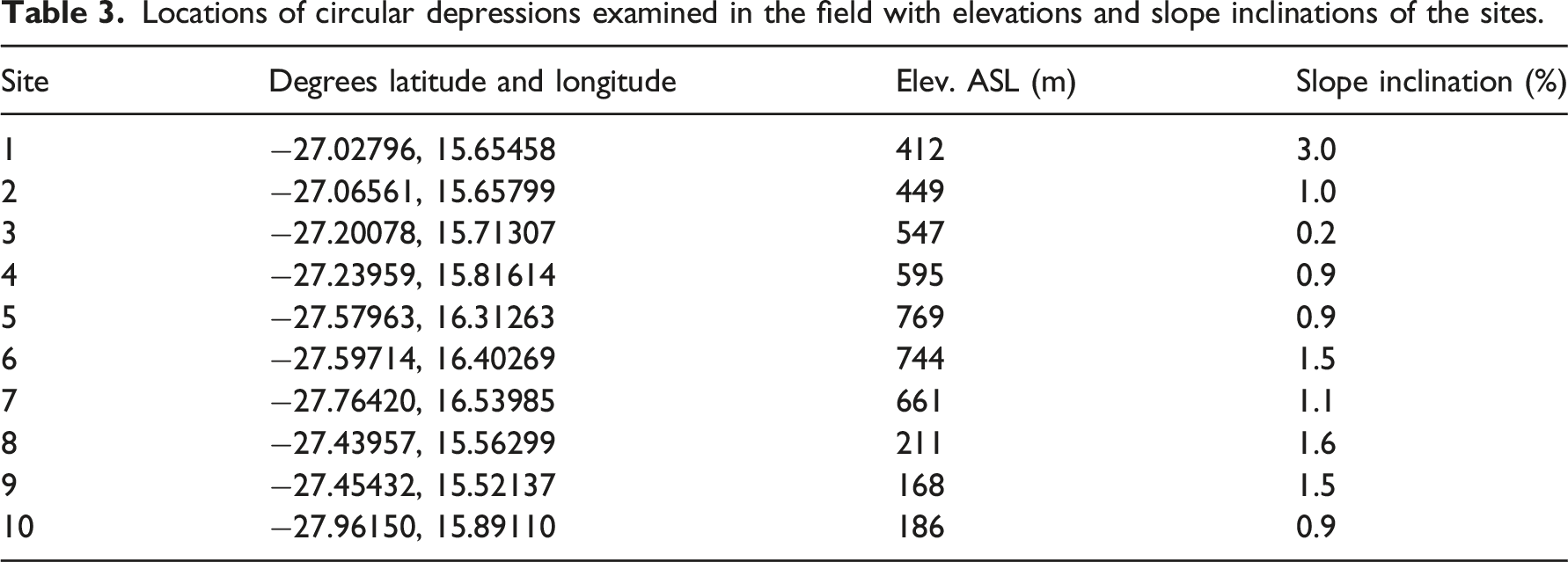
Soils surrounding the depressions commonly contain strongly cemented, but highly fragmented accumulations of pedogenic calcium carbonate (calcrete) at depths as shallow as 10 cm or less beneath thin, gravelly to sandy soils. In such settings, accumulations of cobble-sized calcrete clasts occur on the soil surface immediately surrounding and between circular depressions, but not within the depressions (Figure 2(a)–(c)). Exposed surfaces of strongly indurated calcrete clasts surrounding the perimeter of circular depressions typically have surfaces with deep (>10 mm) surface pitting and etching due to dissolution weathering, indicating a lengthy duration of aerial exposure (Figure 2(d)). In areas of alluvial deposits containing coarse, cobble-sized stones of various rock types, those materials also occur on soil surfaces surrounding circular depressions, but not within depressions (Figure 2(c)). Margins of circular depressions are marked by a sharp transition from the clast-free surfaces within the depression to the presence of large clasts on the soil surface at the perimeter (Figure 2(a)). Soils within circular depressions generally consist of relatively structureless, sandy soil containing relatively fine gravel particles (<1 cm diameter) and an absence of calcrete to the depth of a spade (∼30 cm). Fine gravelly soils of some locations, including the coastal plain and basin floors distant from upland sources of coarse, stony alluvium, commonly have developed in parent materials that that lack large clasts. Nevertheless, circular depressions in such areas can be distinguished as distinct patches of considerably finer-grained materials compared to the coarser gravel surfaces of the immediately surrounding terrain; site 4 (Table 3) exhibited these characteristics.
Vegetation
Circular depressions entirely lacked perennial vegetation at some localities examined in 2021 and 2022. However, when perennial plants were present, vegetation consists almost exclusively of a single species—the small, succulent-stemmed shrub, Brownanthus pseudoschlichtianus (Aizoaceae). In contrast, the areas surrounding circular depressions typically contain several perennial plant species, including small woody shrubs (Salsola sp.), various dwarf shrubs of the Aizoaceae, including Eberlanzia sp. and Stoeberia sp., and Tetraena clavata (Zygophyllaceae), and the stem-succulent Euphorbia gummifera (Euphorbiaceae) (Figure 2(a), 3(a)). (a) Circular depression at Site 8 occupied by Brownanthus pseudoschlichtianus. The darker cobbles about the perimeter are basalt. (Site 8, 29 Sept 2022). (b) Google Earth Pro™ view of area at Site 8 showing dark vegetation at centers of circular depressions. The white arrow points to the circular depression shown in (a). The square shown in red is the 4 ha area used in the analysis of density and spatial pattern (Table 1).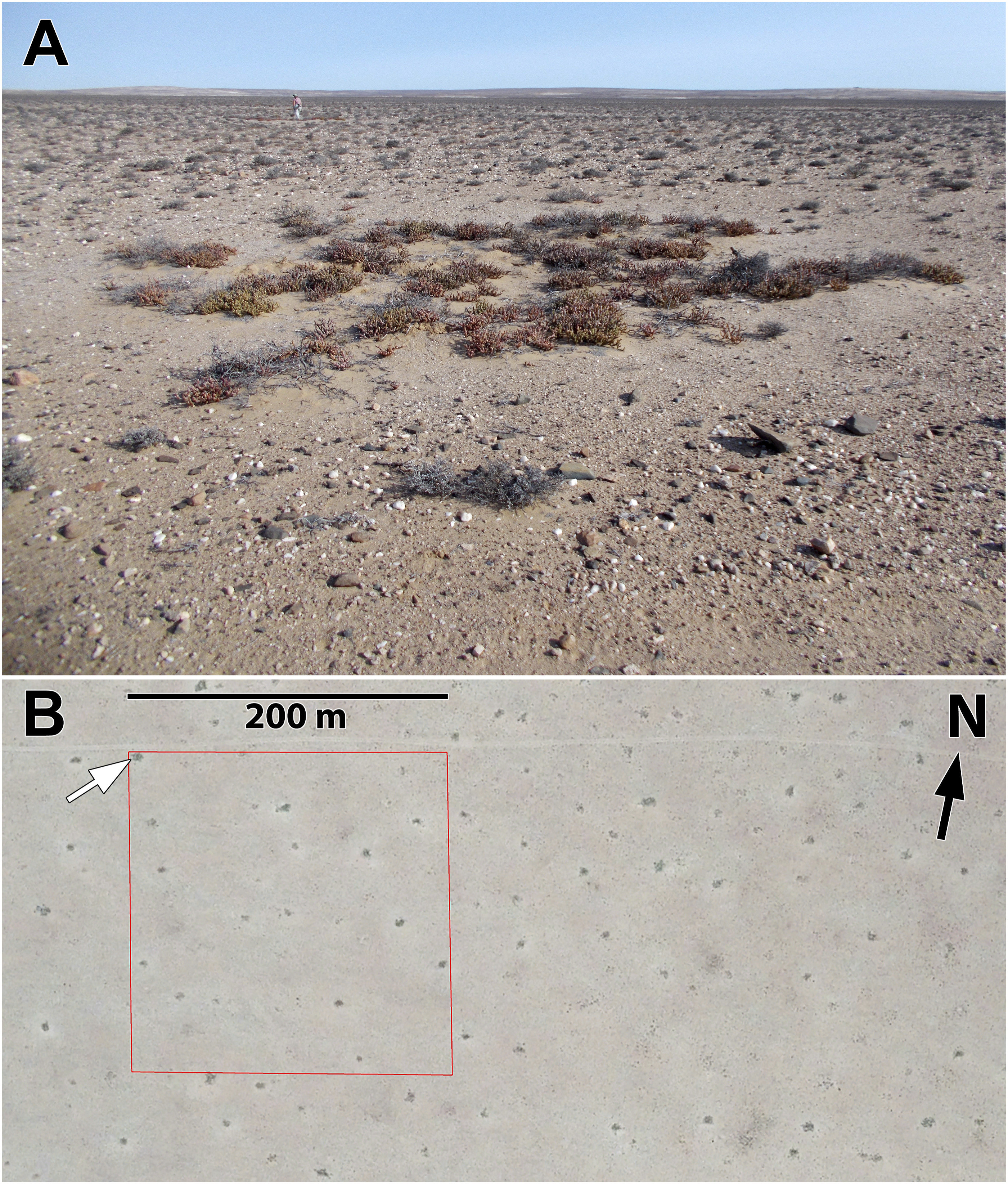
Time series of Google Earth imagery indicate marked, short-term changes in vegetation cover within circular depressions, likely due to the emergence and transient cover provided by short-lived plants. Ground observations at Site 7 in 2021 and 2022 showed the depressions to be largely devoid of vegetation (Figure 4(a)). High-resolution imagery for that location on Google Earth exists for eight dates from March 2003 through May 2023. The March 2003 image indicates little vegetation cover within circular depressions, but the next image from November 2006 shows dense cover of vegetation dotting the area; that vegetation cover is within circular depressions (Figure 4(b)). The southern Namib Desert experienced exceptional rains earlier in 2006, which produced a flush of vegetation in places where vegetation had been almost entirely absent for many years (Muller et al., 2008). However, by the time of the next image on January 2011, those dense vegetation patches had disappeared, and the circular depressions apparently remained relatively barren through the time of ground observations in 2021 and 2022 (Figure 4(c)) and to the most recent imagery in May 2023. The plant species responsible for the short-lived pulse of vegetation cover exhibited in the 2006 imagery are unknown, but may include grasses (Stipagrostis obtusa or S. ciliata) and/or ephemeral members of the Aizoaceae and Asteraceae. (a) Circular depression nearly entirely lacking vegetation (Site 7, 19 Oct 2021) (N. Jürgens photograph, vegetation relevé location 37706). (b) Google Earth Pro™ view in November, 2006 of site 7 showing circular depressions with dark vegetation cover. White arrow indicates the circular depression shown in (a) The red square designates the boundary of the 4 ha plot used for analyses of spatial pattern. (c) Google Earth Pro™ view in October 2018 of same area indicating the disappearance of dense vegetation cover within circular depressions.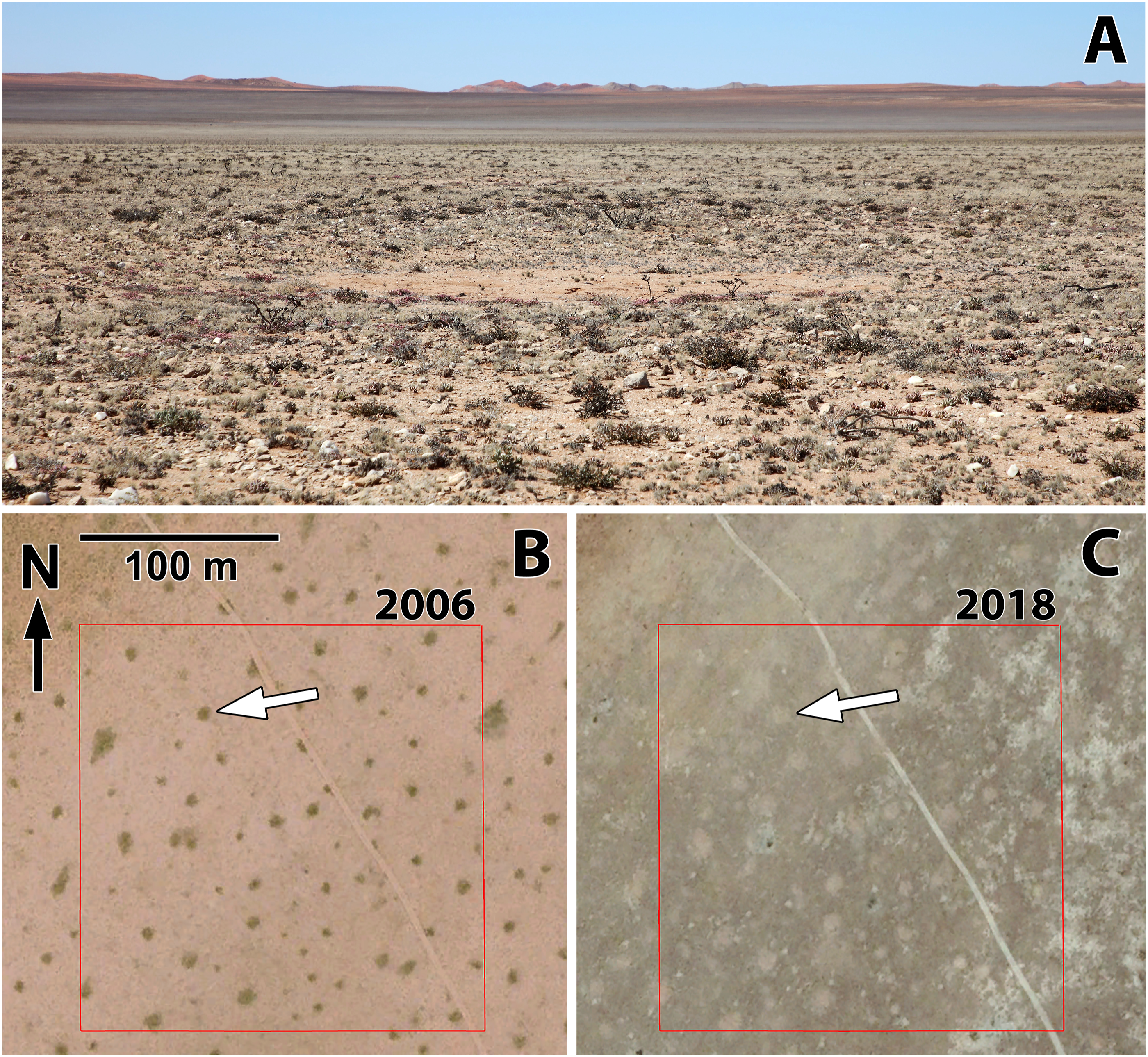
Density and spatial pattern
At the four sites where locations of individual circular depressions were determined from either aerial imagery (Sites 7–9; Figure 4(b), 3(b)) or by mapping on the ground (Site 10; Figure 5(a)), density of circular depressions range from approximately 10–20/ha. Mean distances between centers of nearest neighboring circular depressions correspondingly range from approximately 17 m to slightly more than 22 m. Regardless of density, circular depressions were significantly overdispersed (tending strongly towards even spacing) at all the sites (Table 1). (a) Google Earth Pro™ aerial view of Site 10 showing regularly spaced circular depressions, many containing vegetation cover. The red square designates the boundary of the 1 ha plot within which positional data were recorded on the ground of centers of all circular depressions. (b) Ground photo of circular depression occupied by a multiple burrow openings within a (warren) of Parotomys brantsii at Site 10. The area occupied by the burrow openings is enclosed in a dashed ellipse and small arrows point to five individual burrow openings in the foreground. Additional burrow entrances are distributed throughout the area within the ellipse. (c) Burrow entrance hole within a warren of P. brantsii; centimeter scale divisions. The succulent-stemmed plant is Brownanthus pseudoschlichtianus, stems of which are harvested and eaten by the rodents occupying the burrows.
Dimensions of circular depressions, animal activity, and associated soil modifications
Average diameter of circular depressions at Site 10 was 7.5 m (range = 4.8–11.5 m, n = 7), and measured depths ranged from 10 to 13 cm (n = 6). Site 10 was the only site that exhibited considerable evidence of active or recent occupation of circular depressions by burrowing rodents. Evidence or such occupation was the presence of fresh fecal pellets at burrow entrances, rodent tracks leading to and from the entrances, and burrow openings that had not collapsed. Six of the seven (86%) circular depressions at Site 10 from which diameter measurements were made contained active or recently active burrow systems (Figure 5(b)). Individual depressions with rodent burrows contain a mean of 21.8 entry holes to burrow systems of rodents (range = 8–38, n = 6), and mean diameter of entry holes is 8.3 cm (n = 25 holes; five holes measured in each of five circular depressions; range = 5.0–12.0 cm, 95% C.I. = 7.6–9.0 cm) (Figure 5(b) and (c)). Rodent fecal pellets distributed around entrance holes ranged from 10 to 12 mm long and 4–5 mm diameter.
A single animal standing upright at a burrow entry hole was observed and was visually identified as a Brants’ whistling rat (Parotomys brantsii) before it retreated into the burrow. The soil surface at many entrance holes was littered with remnants of partially eaten, succulent living stems of B. pseudoschlichtianus. The consumption of fresh stem tissues of this plant provides further evidence that the burrow occupants were P. brantsii because this rodent is strictly herbivorous. Parotomys brantsii excavates burrows to depths of 25–35 cm, and individual burrow systems (warrens) contain multiple entrance holes (Jackson, 2000), as was observed at the site (Figure 5(b)). Individual warrens are typically occupied and defended by single individuals (with the exception of females with developing young) (Jackson, 1999). The average diameter of entrance holes and the large number of closely spaced holes within individual circular depressions also match that recorded for P. brantsii at a South African site (Jackson, 2000).
Soils of the surrounding area between circular depressions contain moderately developed soil profiles with A and B horizons that have higher combined silt and clay content than soil from the circular depression (Table 2). Development of structure and durable peds within the A horizon yields a soil surface that is resistance to wind erosion (Figure 6(a)–(c)). In contrast, persistent bioturbation by rodents within the areas of their warrens destroys soil horizon structure, making the fine-grained soil materials highly susceptible to removal by the wind (Figure 6(d); Table 2). Loss of those materials from the areas of concentrated bioturbation contributes to formation of the small, shallow deflation basins. Soil within the circular depression also contains higher concentration of salts, as indicated by measures of E.C. and exchangeable sodium (Table 2). The salt accumulation can be attributed to concentration of these materials due to a long duration of inputs by rodents (i.e., urine, feces) within a restricted area. This greater salinity may be a factor contributing to the distinct vegetation within circular depressions, particularly the occurrence of B. pseudoschlichtianus, which is salt-tolerant. (a) Soil surface at Site 10 located outside of circular depressions. Coarse, white gravels are quartz. Scale divisions in centimeters. (b) Knife inserted into soil outside of circular depression showing subangular blocky structure, imparts resistance to aerial deflation. (c) Soil profile located outside of circular depression showing horizon names (profile description in Table 2). Both the Ak and Bk horizons exhibit moderate structure rendering them resistant to wind erosion. The Bkm horizon is a massive, highly fractured, strongly cemented calcic horizon, but is somewhat obscured in the photograph due to material from upper horizons sliding over it. (d) Soil surface within a circular depression at Site 10 consisting of structureless, non-cohesive sand.
In addition to bioturbation by rodents, considerably larger excavations by predators in search of rodent prey may also play a key role in the formation of another characteristic of many circular depressions—the elimination of large clasts from their surfaces. Near Site 10, a honey badger (Mellivora capensis) was observed hunting by excavating within warrens of P. brantsii (Figure 7(a) and (b)). Such hunting excavations are likely concentrated within the distinct patches containing the warrens, since rodent burrows are absent from intervening areas. At another locale within a warren of P. brantsii, deep excavation attributed to a honey badger brought large fragments of calcrete to the surface, spreading some of that material more than a meter away from the focal point of excavation (Figure 7(c)). The aardvark (Orycteropus afer) is another mammal capable of making large, deep excavations and bringing large clasts to the surface. However, this termite- and ant-eating specialist is absent from the area of study and throughout the extremely arid Namib Desert (Taylor et al., 2016). Consequently, throughout the arid Namib Desert region, excavation by relatively large predators of small mammals, particularly the honey badger, may contribute significantly to the dislodging of large fragments of subsoil calcrete and subsurface stones, and transfering those materials to the surface. The question remains, though, as to how movement of large clasts to the surface in this manner could possibly contribute to net directional displacement of the clasts from within circular depressions that range up to ∼10 m diameter. (a) Honey badger excavating within a burrow system of Parotomys brantsii. The reddish-hued vegetation is Brownanthus pseudoschlichtianus within a circular depression containing the burrows. Fine dust thrown up during the excavation is seen to the left of the honey badger. The black-tailed jackal to the right closely followed the badger at a distance of several meters; this behavior has been described as “kleptoparasitism” in which occasionally the jackal opportunistically seizes prey flushed by the badger (Begg et al., 2022). (b) View of honey badger and accompanying jackal after excavation shown in (a) White arrow points to pit dug by honey badger (Norbert Juergens photographs, 3 Oct 2022 near site 10). (c) Hunting excavation at another location attributed to a honey badger within a burrow system of P. brantsii. The large white fragments displaced to the right are pieces of calcrete removed from deeper levels by the excavation. Some of the fragments were displaced more than 1 m beyond the focal point of excavation (30 Sept 2022, −27.62918°, 16.31263°).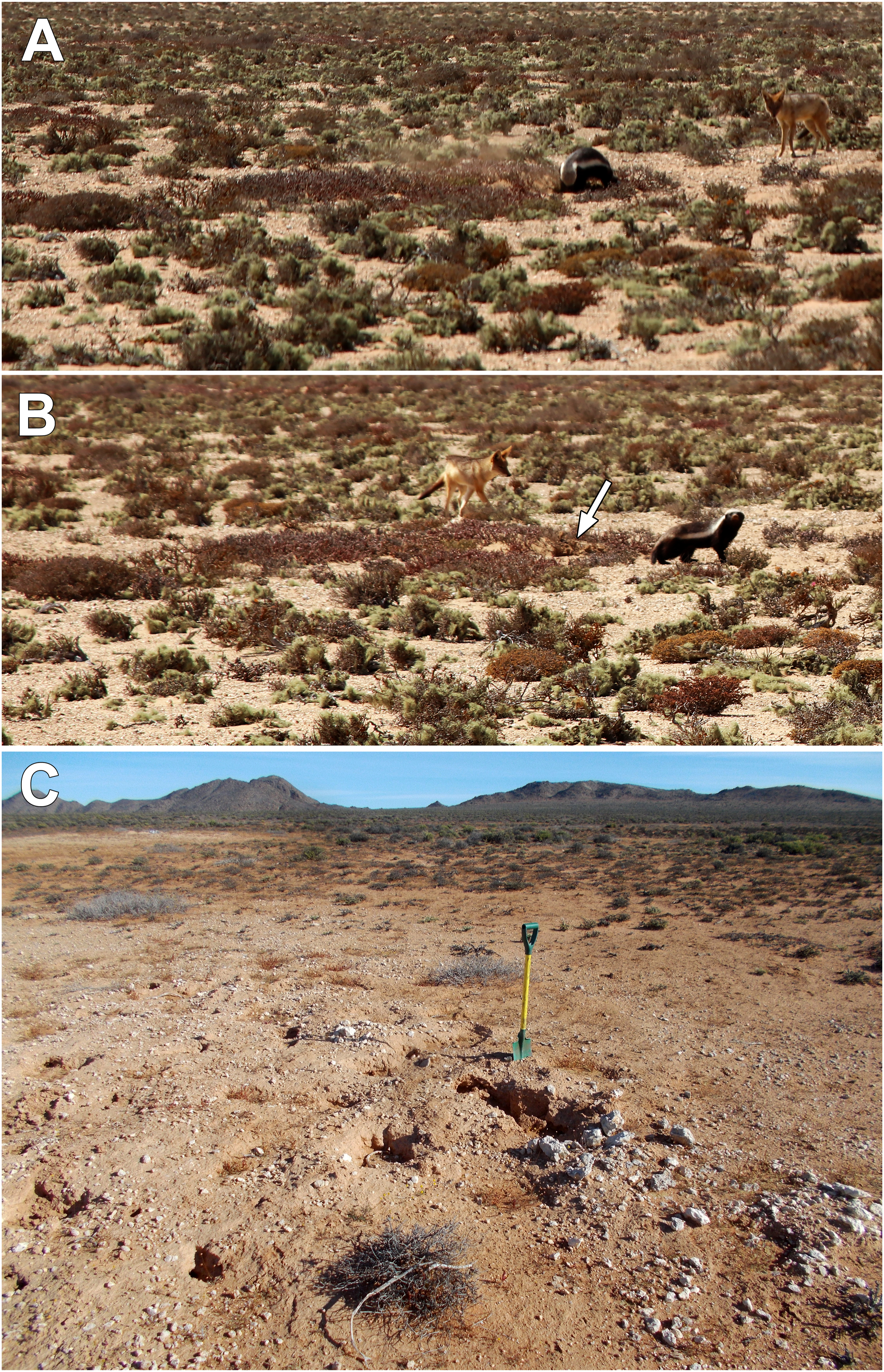
Conceptual model: Two-dimensional random walk of mammal-displaced clasts
Movement of individual, large clasts within well-defined, widely separated patches occupied by burrows of whistling rats or other burrowing rodents can be represented as a two-dimensional random walk. Starting with a general premise that predators such as honey badgers focus their rodent hunting excavations within areas containing burrows occupied by rodents, and a conservative assumption that successive excavation events tend to move large clasts about the surface in random directions, the sequential movement over time of a large clast over the surface takes the form of a two-dimensional random walk (Figure 8). As long as the clast remains within the “arena” occupied by a concentration of occupied rodent burrows (i.e., the zone that elicits hunting excavations), the random walk of that clast will continue through time due to occasional displacement. However, once a clast is displaced to a position outside the boundary of the arena occupied by rodents and associated hunting excavations, it would tend to remain in that position, as it is no longer subject to the same kind of repeated movement. Over time, the collective consequence of the random walks of large clasts brought to the surface is their eventual exclusion from the arena of active excavation, coupled with the removal of large clasts from the subsoil environment to the depth of excavation by large mammals. The number of individual steps taken by a single clast before expulsion from the arena can conceivably range from 1 (e.g., a clast originally positioned on the surface very near the outer boundary, where a single step could remove it beyond that boundary) to a limit of infinity (but with an accompanying, infinitesimally small probability of occurrence). Nevertheless, in this conceptual model, the fate of most objects (large clasts moved about the surface by excavation activities) is their eventual expulsion beyond the outer boundary of the arena of excavation, yielding a patch lacking those objects on the surface and in the soil to the depth of excavation, and a concentration of those objects on the surface beyond the outer boundary of the arena. (Upper) Diagrammatic representation of a two-dimensional random walk within a circular, bounded arena. A uniform step size is shown for simplicity, but step size of an actual clast over time moved about during large excavations would be variable. (Lower) Relationships between linear displacement from a starting point of an object subject to a two-dimensional random walk and the number of steps for three different step sizes (l). The curves represent the mean expected distance moved; displacement of individual objects would be normally distributed about the depicted average, and increase with number of steps. Average number of steps required to linearly displace an object 5 m from the starting point (radius of a large circular depression), given step sizes of 1.0, 0.5, and 0.2 m, ranges from 25 to 625 steps as shown.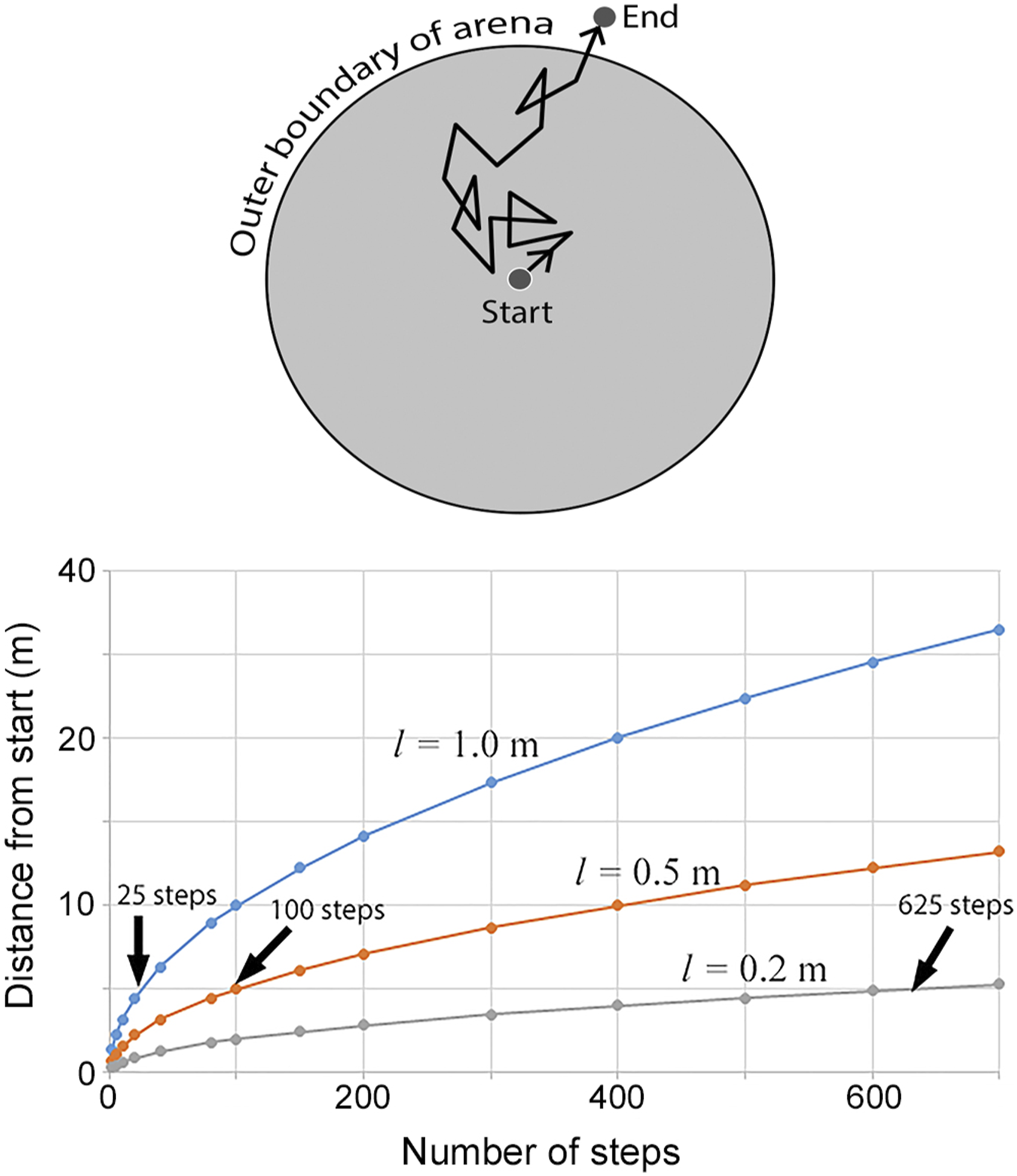
How plausible is this model explanation for the formation of the clast-free interiors of circular depressions in the Namib Desert region? Calculation of the average number of steps within a two-dimensional random walk required to move materials a given distance indicates whether such a process could conceivably generate observed phenomena within a realistic time frame of the late Quaternary (i.e., latest Pleistocene and Holocene).
Within a two-dimensional random walk, a measure of the linear displacement of an object from the starting point is referred to as the root mean square displacement (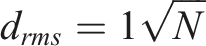
where
The mean number of steps (size 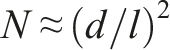
Curvilinear relationships between average distance from the starting point and number of steps in two-dimensional random walks of three different step sizes: 0.2, 0.5, and 1.0 m are shown (Figure 8). These step sizes are well within the range of distances that clasts are laterally displaced in a bout of hunting excavation by a honey badger (Figure 7(c)). The arrows indicate the average number of steps required to move 5 m away from the starting point (i.e., the radius of a large circular depression) for each of the step sizes. It is noted, though, that these relationships depict the mean distance moved away from a starting point as a function of step number, and the actual distribution of distances moved is normally distributed, with the variance increasing with step number.
Discussion
Mechanisms of formation and similar features in other parts of the Namib Desert
Evidence collected from Site 10 indicates that the circular depressions form in association with long-term occupancy of regularly spaced patches by burrowing rodents. The other sites (e.g., 7, 8, and 9) contain circular depressions with the same dimensions, characteristics, and spacing patterns, and consequently, likely represent features that originally formed through the same processes. The burrowing and excavating activities within these patches by mammals, including large predators, function as geomorphic agents in two principal ways in the formation of the depressions: (1) directly, through vertical and horizontal movement of materials during excavation and (2) indirectly through bioturbation, which destroys soil structure and increases the propensity for localized removal of non-cohesive soil materials by the wind, contributing to formation of small deflation basins.
The circular vegetation features in the central Namib Desert described by Louw and Seely (1982) and Cox (1987) are similar to the circular depressions we describe, since they consist of circular patches of sandy soil surrounded by a conspicuous border of coarser gravel, and are spatially overdispersed. Formation of those features was associated with burrowing activity of gerbils (G. setzeri). The patches with active or formerly active rodent burrows studied by Cox (1987) were smaller in diameter (X̅ = 5.9 m) than those occupied by Paratomys brantsii at Site 10 (X̅ = 7.5 m). Additionally, the features studied by Cox (1987) were more closely spaced (X̅ = 11.2 m), and with higher densities (32.3/ha) than the circular depressions at sites 7, 8, 9, and 10 the southern Namib (X̅ spacing = 17.3 m – 23.5 m, densities = 9.8–22.0/ha; Table 1). This contrast likely reflects differences in the sizes of rodent species associated with the formation of the features and the relationship between body size and area occupied by burrow systems. Individual adult body mass of the gerbil G. setzeri ranges from 30 to 40 g, compared to 75-111 g in Parotomys brantsii (Wilson, 2017).
Cox (1987) attributed the zone of coarser gravels surrounding circular vegetation (patches to the directed displacement of soil excavated by the rodents outward from the colony center, yet no detailed observations or data were provided to convincingly demonstrate this claim. Winnowing of fine materials by the wind was cited as the mechanism responsible for eventually concentrating coarser gravels on the surface of materials transported to the perimeter. Slight depressions in some of the vegetated patches were attributed to the collapse of vacant burrow systems from which soil had been previously excavated. Cox (1987) also noted the presence of large fragments of calcrete on the surface of some the circular features and suggested that some of those materials could have been excavated by predators digging for rodent prey. However, no information was provided on the composition or occurrence of larger clasts around the perimeter of the circular depression, or how those materials might accumulate.
The removal of large clasts from the circular areas and accumulation of those materials around the perimeters do not necessarily require a directional bias of transport away from the center during excavations by either rodents, as suggested by Cox (1987) or by larger predators. Over time, an unbiased, two-dimensional random walk of such materials is conceivably capable of eventually yielding a surface free of large clasts, and an accumulation of those materials in a zone surrounding the area of repeated excavation. Similarly, although the directional displacement of soil materials by burrowing rodents outward from the center of a colony as suggested by Cox (1987) could contribute to the eventual collapse of burrows that are eventually abandoned, in the strongly windswept environments of the Namib Desert, such a process would not be essential to lower the center of the areas subject to bioturbation by rodents. Destruction of soil structure by repeated bioturbation creates a non-cohesive mass of soil that is extremely prone to wind erosion, thereby creating a shallow deflation basin. The considerably lower silt content in the soil from a circular depression, and corresponding greater sand content than in soil of the surrounding area (Table 2) likely reflect, in part, the selective winnowing of the finer particles by the wind. Following abandonment by rodents and cessation of bioturbation, continued wind erosion of fines would yield the coarser gravelly lag that is commonly present on the surfaces of circular depressions (Figure 2(b)). Soils outside of circular depressions possess developmental characteristics (structural development of soil horizons associated with higher silt and clay contents and lack of bioturbation) that provide considerably more resistance to wind erosion, as has been observed in other arid environments (Ellwein et al., 2018). Additionally, in the fog-influenced Namib, biogenic soil crusts are also often present (Büdel et al., 2010), stabilizing the soil surface and thereby preventing erosion. Repeated bioturbation by rodents eliminates such crusts, exposing disturbed areas to deflation.
The regular spatial dispersion of circular depressions observed at all sites we investigated in the southern Namib (Table 1), as well as the features examined by Cox (1987) can be attributed to intraspecific, territorial interactions among rodents occupying burrow systems. Warrens of Parotomys brantsii like those at Site 10 (Figure 5(b)) are most frequently occupied by single individuals, with some warrens shared by one or more others (Jackson, 1999). However, the rodents exhibit territorial defense of either entire warrens or the portions of individual warrens they occupy. G. setzeri also exhibits some degree of territorial behavior (Demster et al., 1998). Once an array of regularly spaced burrow systems is established by populations of any of these rodents, particularly within landscapes with shallow, relatively impenetrable calcrete horizons, those locations likely are repeatedly occupied for an extended period due to the greater ease of burrowing and occupation of previously disturbed areas. Consequently, the positioning of burrowing systems remains relatively fixed for long durations of time.
Although the circular vegetation features associated with gerbils in the central Namib described by Cox (1987) are smaller in diameter and are more closely spaced than the circular depressions in the southern Namib, circular depressions similar in size and spacing to those we describe in the southern Namib also occur in other portions of the Namib Desert. At a site approximately 75 km southeast of the area studied by Cox (1987), the ‘calcrete plain’ landscape contains slight, circular depressions consisting of deeper sandy or silty soils and are surrounded by a zone of large fragments of calcrete and other clasts (Jürgens, 2022, Figure 9.6.10, p. 296; −24.03116°, 15.66000°). However, in areas with much younger landforms consisting of fine-grained sediments in wide alluvial plains and apparently lacking substantial formation of pedogenic calcrete, regularly spaced circular depressions occur that lack a surrounding ring of coarse materials (Jürgens, 2022; Figures. 9.6.8 & 9.6.9, p. 295; −22.86751°, 15.06500°). Those structures are similar to those that we observed at one of the 10 sites where we documented the presence of circular depressions in the southern Namib Desert (Site 4, Table 3).
Contribution of large, excavating predators
Honey badgers clearly displace large clasts excavated from relatively deep within the soil profile to the surface (e.g., Figure 7(c)). It is unlikely that displacement of large clasts that commonly surround circular depressions can be attributed to the activities of smaller rodents alone because the mass of single clasts can be more than an order of magnitude greater than that of an individual rodent. However, those materials are readily excavated and laterally displaced by much larger, excavating predators. For example, in the Sonoran Desert region in the USA, McAuliffe and McDonald (2006) documented the excavation and movement of stones up to 20 cm diameter and 2.5 kg by the American badger (Taxidea taxus), a predator with body mass of ∼9 km that, like the honey badger, excavates burrow systems of rodents in search of prey.
The size of honey badgers (90 cm body length; ∼9 kg), combined with powerful forelimbs and long claws, enable them to efficiently dig in search of prey (Vanderhaar and Hwang, 2003). Small mammals are one of the most important food sources for this predator and excavation into rodent burrow systems is a predominant mode of hunting (Kruuk and Mills, 1983). At a site in the Kalahari region of northern South Africa, predation by honey badgers on small mammals was directly correlated with the relative abundance of the rodents (Begg et al., 2003). When abundant, rodents accounted for more than half of food items in the honey badgers’ diets. At that site, the rodents most commonly consumed were two species of gerbils that live in burrow systems. Individual honey badgers forage over large areas (males: 548 km2; females: 139 km2) and make multiple excavations on a daily basis in search of prey. Direct observation by Kruuk and Mills (1983) revealed that an adult male traveled a linear distance of 27 km/day while foraging, digging an average of 1.3 holes/km in search of prey. One female observed, although traveling a shorter distance of 10 km/day, dug an average of 10.2 holes/km traversed. Over time, these excavation activities can significantly affect the landscape, particularly where excavations are focused within areas occupied by rodent burrow systems.
Antiquity of some circular depressions
Processes responsible for formation of circular depressions are apparently ongoing in some areas due to active bioturbation by rodents and associated excavation activities of predators (e.g., Figure 5). However, in some areas, circular depressions lack any signs on the surface of recent occupation by burrowing rodents or other excavation activity and probably formed long ago during periods of less arid conditions when greater vegetation productivity was capable of supporting substantial populations of herbivorous or granivorous rodents. Jürgens (2022, p. 296) also suggested that some of the circular features that occur in the central Namib desert were likely formed under different climatic conditions long ago. Strongly indurated calcrete clasts strewn about the perimeters of circular depressions often exhibit evidence of considerable surface dissolution (Figure 2(d)), which suggests those materials have been aerially exposed for great lengths of time, likely measured in millennia. Weathering of surfaces of carbonate rocks by dissolution is strongly dependent on precipitation. For example, along a precipitation gradient raging from approximately 600 mm yr−1 to 50 mm yr1 in Israel, Ryb et al. (2014) determined that rates of erosion by surface dissolution of relatively level carbonate bedrock surfaces ranged from a corresponding 20 mm ka−1 to 1–3 mm ka−1.
In the southern Namib Desert, bioturbation associated with occupation by burrowing rodents was likely far more prevalent and continuous during less arid climatic intervals of the late Quaternary. Palaeoclimate reconstructions for the arid Atlantic coastal region of South Africa and Namibia based on both marine sediment core and terrestrial pollen records extracted from rock hyrax middens indicate cooler, humid conditions during the last glacial period and warm, arid conditions during the Holocene (Dewar and Stewart, 2016; Lim et al., 2016; Shi et al., 2000), with the late Holocene being the driest period during the last 50,000 years (Chase et al., 2019). Consequently, formation of some of the features, particularly those in extremely barren areas that now lack sufficient vegetation required to support rodent populations, likely date to the late Pleistocene or the early Holocene. For circular depressions with such antiquity, it is possible that the mammal species responsible for original formation of the features may no longer occupy those environments.
Landscape responses and ecological impacts
The circular features briefly mentioned by Louw and Seely (1982) and examined in more detail by Cox (1987) in the central Namib Desert support higher densities of grasses than the surrounding area. Cox demonstrated the higher infiltration capacity within circles and an increased capacity to absorb precipitation as the cause of this difference. In contrast, where perennial vegetation occurs within central depressions at the sites in Tsau ǁKhaeb National Park in the southern Namib, the shrubby, succulent-leaved Brownanthus predominates. This contrast in vegetation response can be attributed to differences between the areas in seasonality of precipitation. The southern Namib region represents the northern extension of the Succulent Karoo Biome that receives cool-season precipitation, where small succulent-leaved shrubs of the Aizoaceae predominate. In contrast, with increased distance to the north, summer precipitation increasingly predominates, leading to a greater occurrence of both perennial and ephemeral grasses as a dominant component of the vegetation. Yet in spite of these differences, circular depressions apparently form due to the actions of burrowing rodents.
Large portions of the southern Namib Desert region consist of extensive planar areas, and the presence of regularly spaced circular depressions provides significant microrelief that may exert considerable influence over landscape processes, particularly surface hydrology and associated soil moisture storage and availability. Consequently, these features may play a prominent role in their influence on both flora and fauna of the region, as indicated by the marked but short-lived vegetation response of vegetation within circular depressions following precipitation (Figure 4(b)).
Similar features in other arid regions
McAuliffe and McDonald (2006) described circular depressions, some of which are similar in size to those in the southern Namib, on old (Pleistocene) alluvial fan surfaces in an extremely arid portion of the Sonoran Desert, USA. Examination of soil profiles indicated those depressions likely formed in locations occupied by rodents and associated bioturbation that develop around the bases of large, long-lived perennial shrubs. Increased aridity associated with major climate changes (i.e., Pleistocene to Holocene) apparently led to mortality of those shrubs in the more xeric interfluve areas, resulting in selective erosion of fine soil materials from the areas of bioturbation, the eventual settling of the residual soil mass, and formation of slight depressions. Accordingly, McAuliffe and McDonald (2006) called these features plant scar depressions. Larger, highly weathered clasts were also distributed about the perimeters of depressions, and the displacement of those objects toward the perimeter of initial bioturbation mounds was attributed to their excavation and movement by larger burrowing predators, particularly the American badger (T. taxus). Like the honey badger of the Old World, the American badger also relies on rodents as a principal prey and excavates and displaces large stones in hunting excavations. McAuliffe and McDonald (2006) suggested that badgers moved stones outward by directional displacement as they excavate, oriented towards the base of a centrally located shrubs and expelling material posteriorly while they dig. However, given the maximum diameters of some of those depressions (6 m), the non-directional displacement of larger clasts involving a two-dimensional random walk over time as proposed in this paper may also have contributed significantly.
Although the circular depressions described in this paper and in McAuliffe and McDonald (2006) in the Sonoran Desert all form in association with localized areas of bioturbation by burrowing rodents, once formed, such features are likely preserved for long durations only in extremely arid environments. For example, in the Sonoran Desert, plant scar depressions are found only in the more arid portions (MAP <100 mm), on Pleistocene alluvial fan surfaces that have minimal slope inclination, where strongly developed stone pavements covered with desert varnish have developed. Such surfaces now generally lack any perennial plants or significant bioturbation. Consequently, lacking biotic activity that would displace surface materials, the depressions have been preserved as microtopographic features for a lengthy duration, likely at least the duration of the entire Holocene or longer. Similarly, in the southern Namib Desert, circular depressions that may have formed long ago, but now lacking significant perennial plant cover and associated animal activity are likely preserved for many millennia, and may represent relict features that actively formed during the late Pleistocene during less arid conditions.
Microtopographic depressions that share characteristics of those described in this paper and McAuliffe and McDonald (2006) may occur in other arid regions of the world, where localized bioturbation by burrowing mammals occurs. For example, a type of circular depression referred to as sorted circles has been described for arid regions of Australia (Mabbutt, 1979). The features include large silcrete clasts distributed about the perimeters of the depressions. Their formation was attributed to expansion and contraction of clay-rich subsoils, and the resulting upward displacement of large clasts to the surface. However, some of these features may also represent localized areas of occupation and movement of subsoil materials by burrowing animals long ago. For example, several species of native marsupial wombats are found in Australia and all create substantial excavations. The southern hairy-nosed wombat (Lasiorhinus latifrons) displaces substantial volumes of soil material from deeper horizons, including calcrete (Löffler and Margules, 1980). The manner in which these and other native burrowing animals in arid zones of Australia may have contributed to different types of microtopographic alteration requires further investigation.
Similarly, other extremely arid regions of the world, including the Atacama of Chile and southern Peruvian Deserts, the more arid portions of the Monte of Argentina, and some of the Middle Eastern and North African desert regions may contain microtopographic features that share characteristics and processes of formation similar to the circular depressions of the Namib. However, for features like these that formed long ago, they might be expected to be found only on older, geomorphically stable surfaces of minimal slope inclination that presently lack significant biotic activity (both plant and animal) that would over time tend to obscure the features.
Further questions
Many questions remain regarding the distribution and formation of circular depressions in the Namib Desert region. To date, there has not been a detailed examination of deep soil profiles through circular depressions. Both the work of Cox (1987) and this study have focused largely on surface characteristics. However, excavation of deep exposures through circular depressions (e.g., >1 m depth) and detailed description of soil profile characteristics and stratigraphy are required to further understand the depth of bioturbation and overall processes involved in formation of the features. That approach was employed by McAuliffe and McDonald (2006) and yielded valuable information on depth of past bioturbation through the identification of old, soil-filled burrow passages (crotovina) and comparison of physical and chemical characteristics with soils of outlying areas. Such an approach also has the potential to yield material that could enable dating of past biotic activity (e.g., radiocarbon dating of organic material contained within crotovina). Additionally, as mentioned in section 4.5 above, investigation across deserts regions of the world on similar features could provide valuable information on general characteristics of landscapes, climate, flora and fauna under which such features develop.
Implications
The circular depressions described in this paper represent persistent environmental legacies of biotic modifications of the physical landscape that in some cases occurred long ago. Despite the antiquity of these structures, they continue to significantly influence the soil heterogenieity and vegetation diversity. Understanding the origin of these features as well as their environmental effects is particularly important in the face of anthropogenic changes to the landscape, (e.g., excavation, mining, and other surface-disturbing activities) potentially exacerbated by the added, predicted increase in aridity in many parts of southern Africa (Hadley Centre, 2001).
Footnotes
Acknowledgements
A special thank-you is given to Bertchen Kohrs of Windhoek, who provided invaluable assistance in obtaining various permits from Namibian governmental ministries. Personnel of the Namibian Ministry of Mines and Energy and the Ministry of Environment, Forestry and Tourism granted the necessary permissions to work in the area. Personnel of Namdeb and Sperrgebiet Diamond Mining provided entry into the area. Mbeytjiua “Ricky” Rikambura (Conservation Scientist) and Kasara Paulus (Assistant Ranger), from the Ministry of Environment & Tourism, who accompanied us for a portion of the trip and provided valuable assistance in locating routes obscured by drifting sand and assistance in freeing a vehicle (twice) deeply mired in sand. Albertha Dausas of the Namibian Ministry of Agriculture, Water & Land Reform in Windhoek provided laboratory services for soil sample analyses.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Federal Ministry of Education and Research (BMBF) of Germany (Grant #FKZ 01LG1201N) (NJ and FG) and the Desert Botanical Garden (JM).